
Flying without a Net: Space Radiation Cancer Risk Predictions without a Gamma-ray Basis

Abstract
:1. Introduction
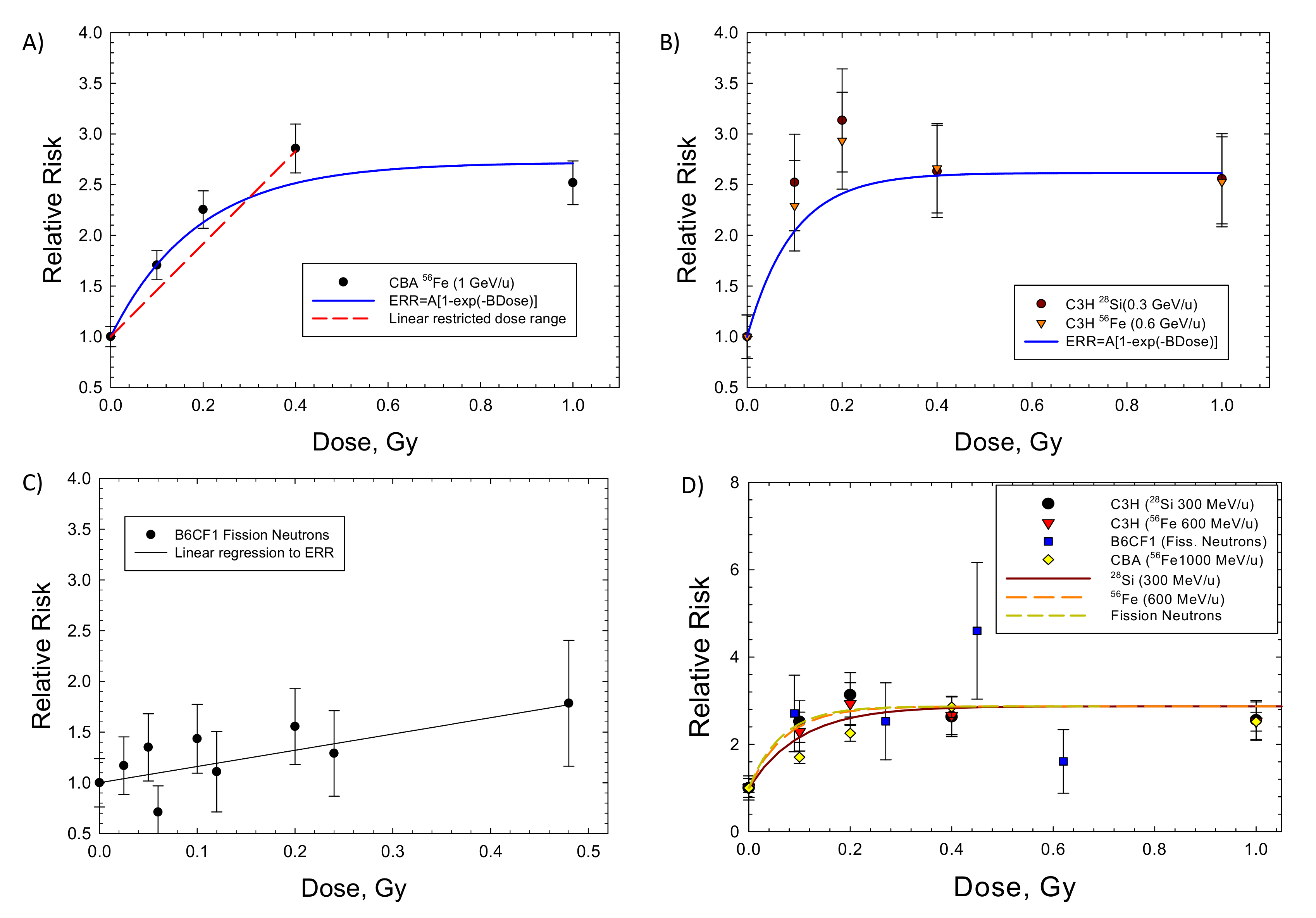
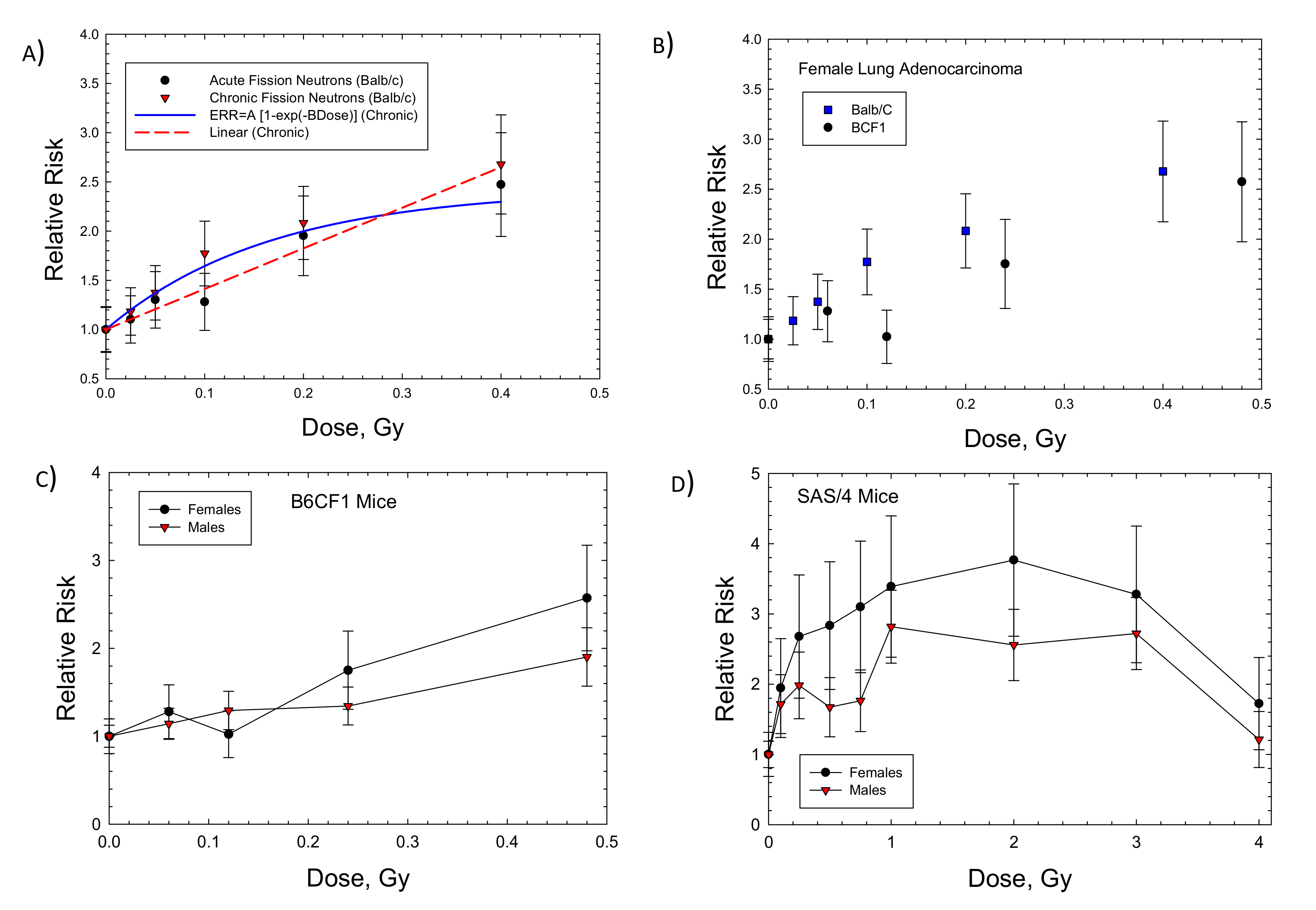
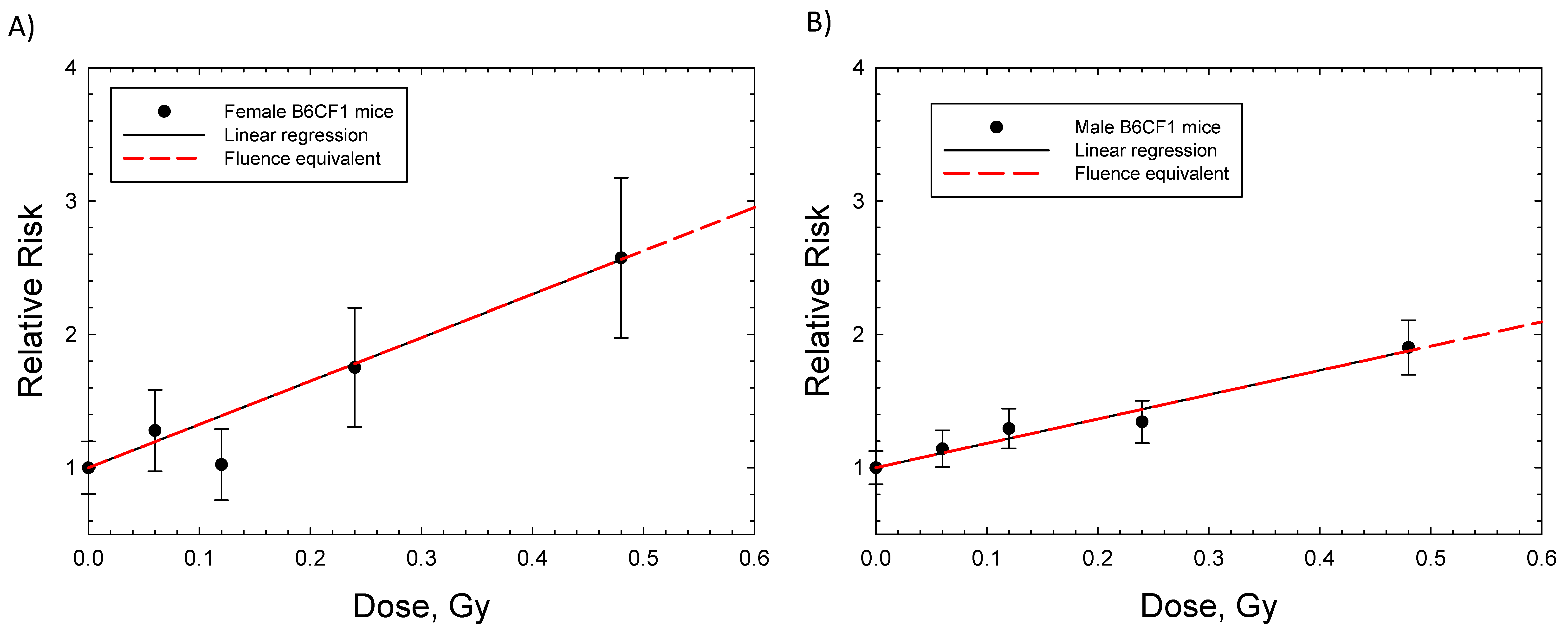
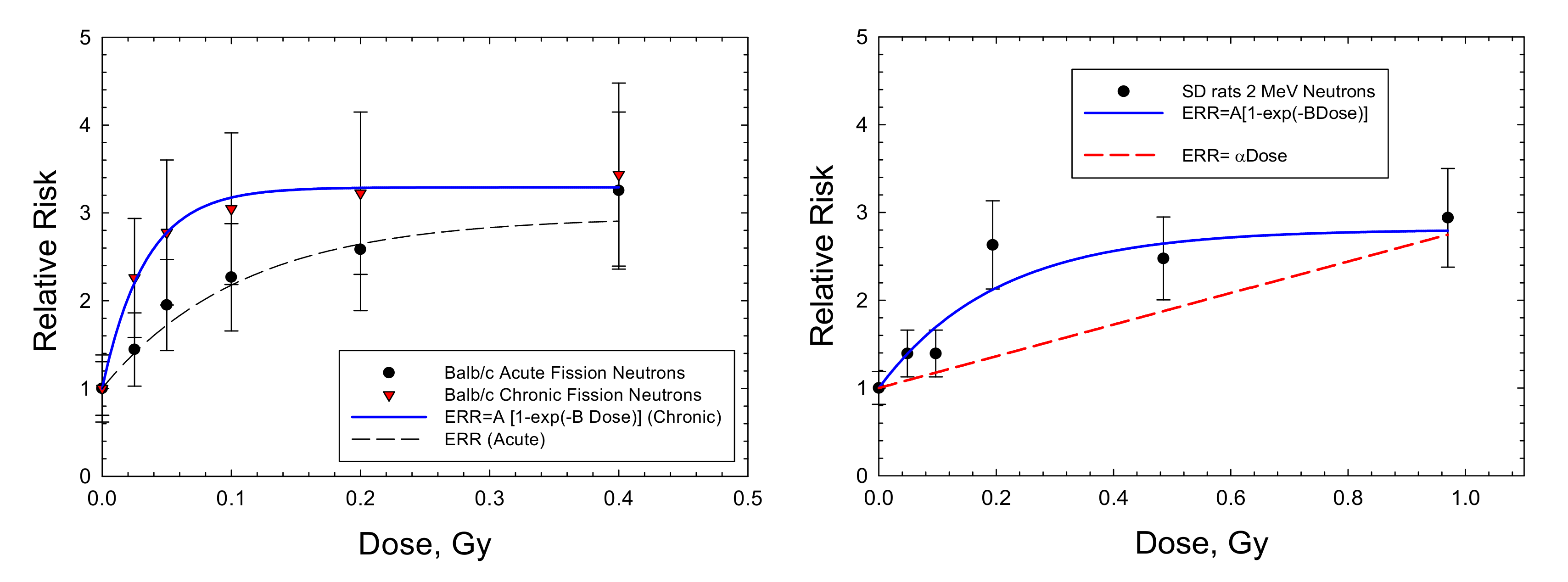
2. Results

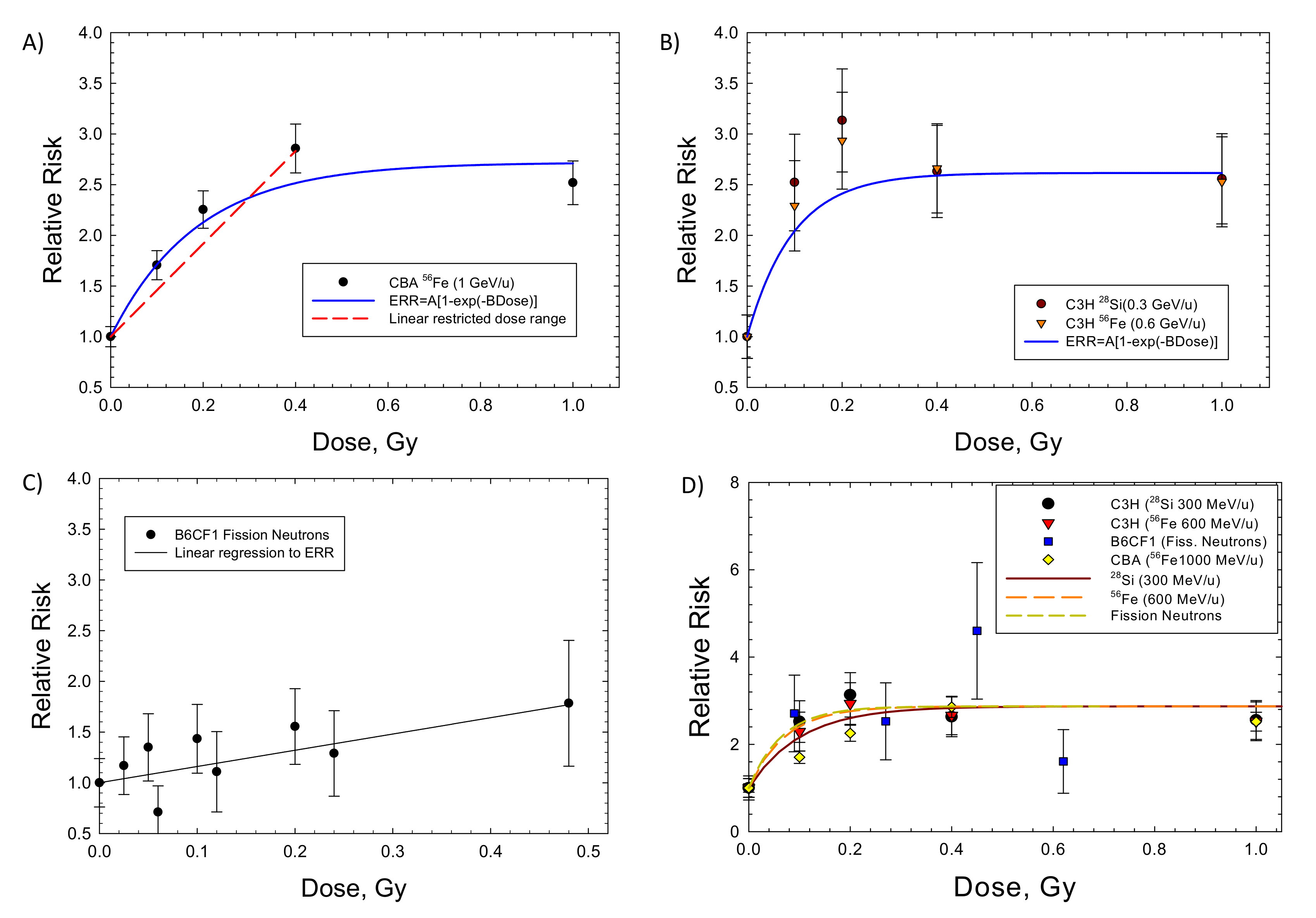{kind=link}
{kind=link}
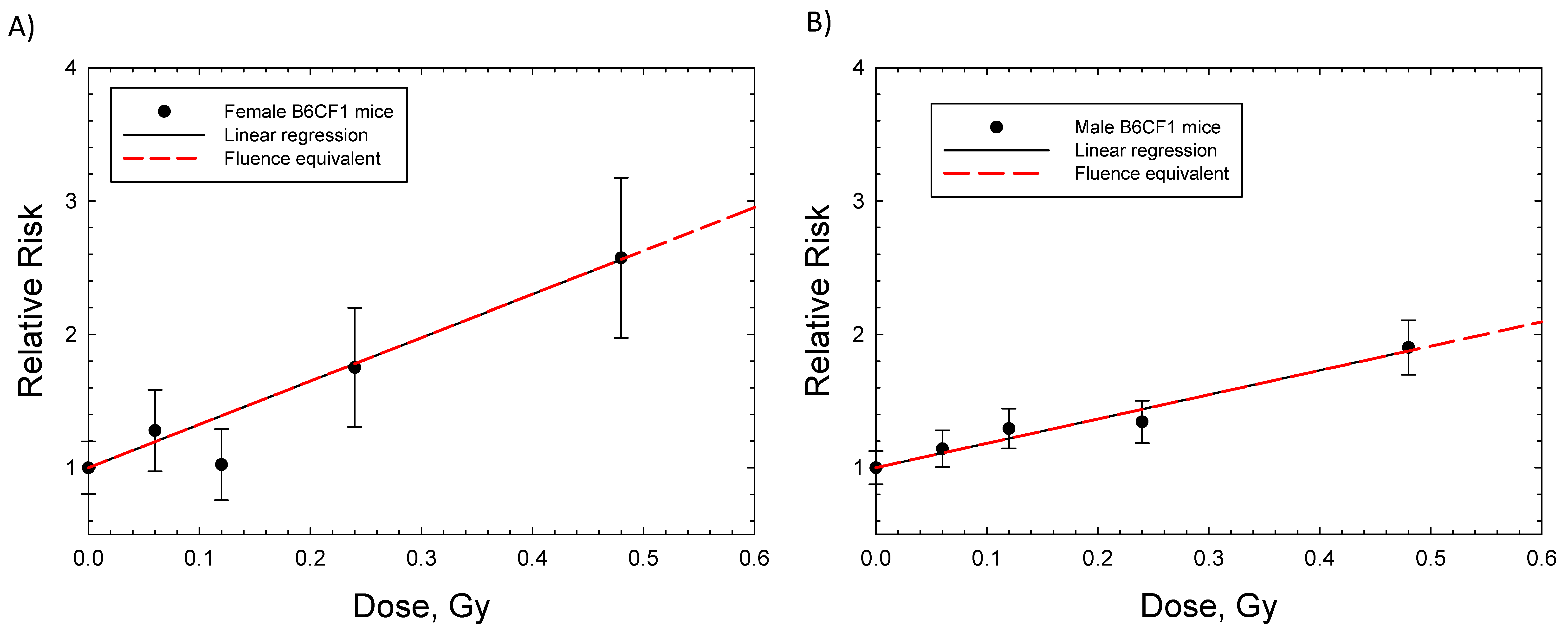{kind=link}
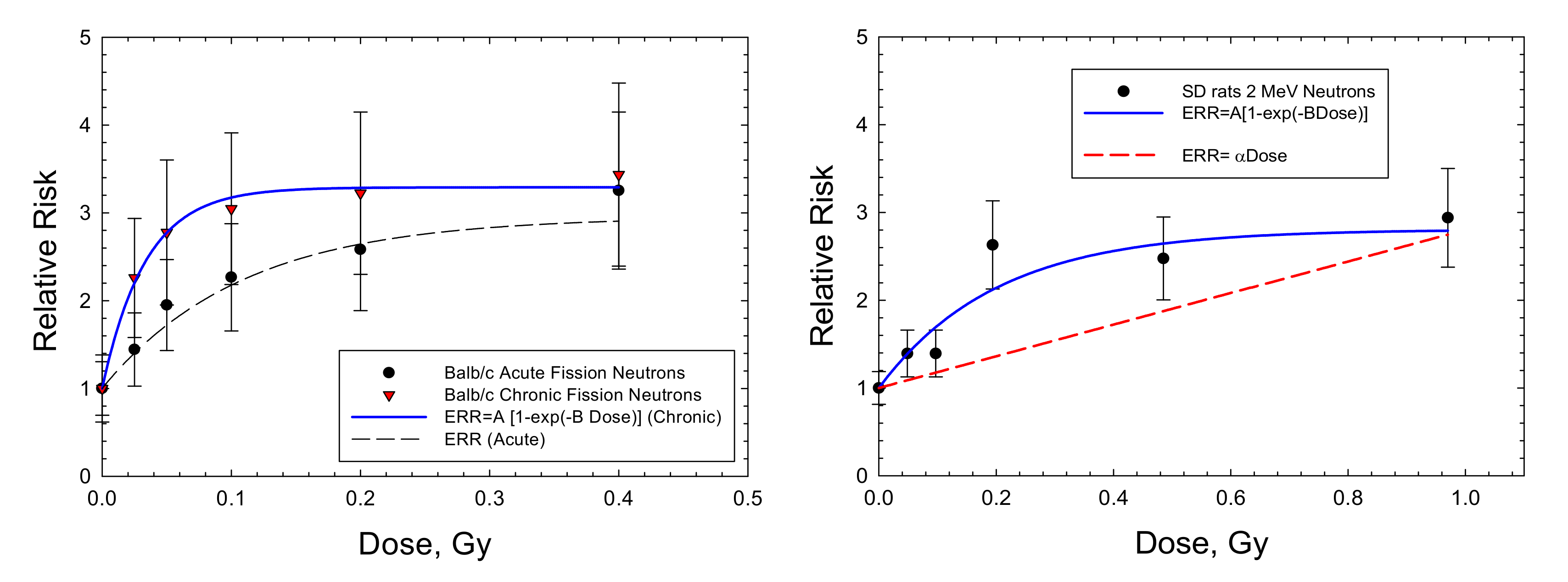{kind=link}
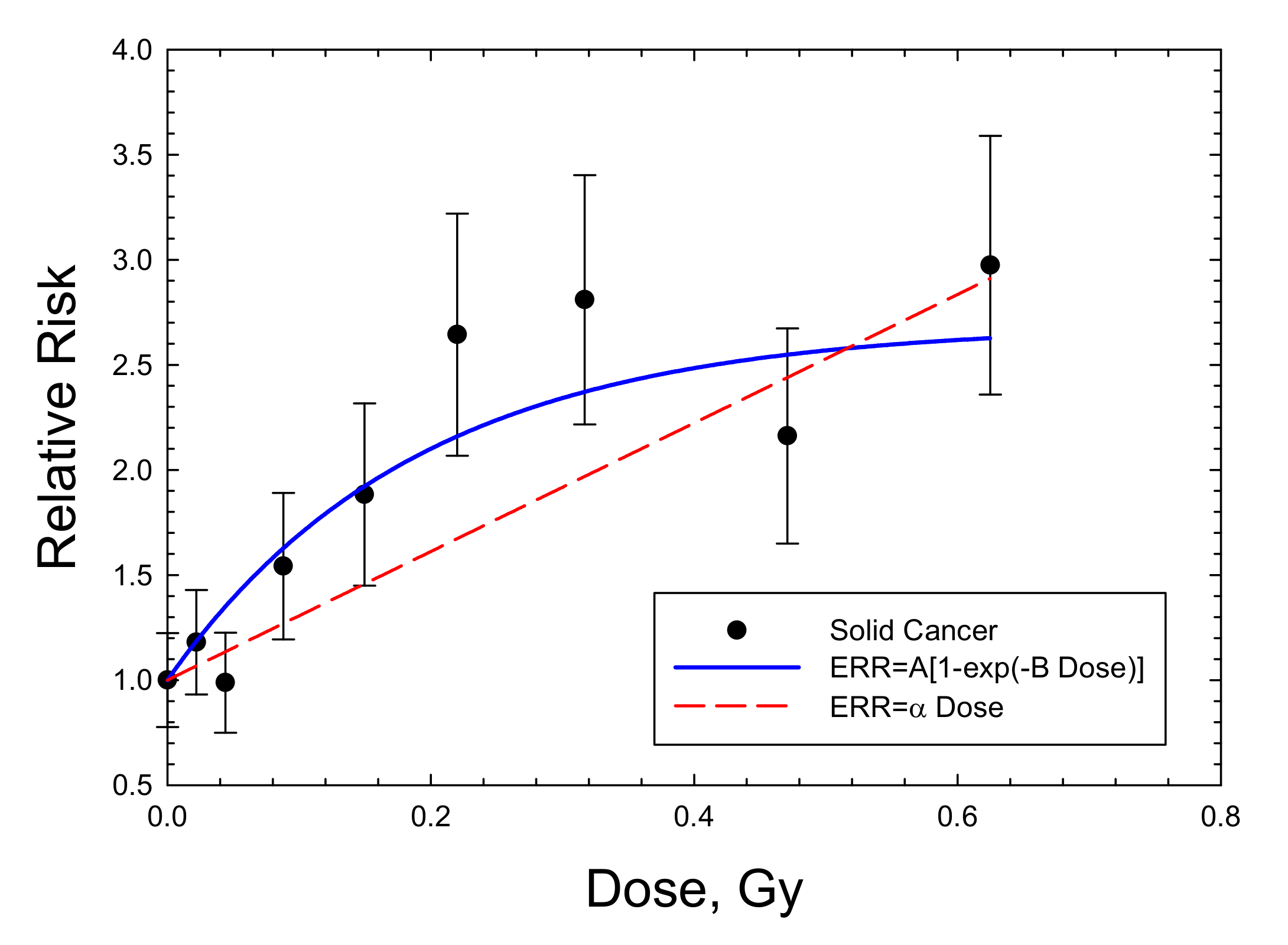{kind=link}
| Model | Tumor | Age at Irradiation | Irradiation Types | Duration of Observations | Reference |
|---|---|---|---|---|---|
| BALB/c/AnNBdf, female mice | Mammary adenocarcinoma | 120 d | FN mean energy 2 MeV, acute and chronic | Lifespan | [6] |
| BALB/c/AnNBdf, female mice | Lung adenocarcinoma | 120 d | FN mean energy 2 MeV, acute and chronic | Lifespan | [6] |
| CBA/CaJ, male mice | Hepatocellular carcinoma | 8–14 weeks | 56Fe (1 GeV/u), acute | Observed to age 800 d | [17] |
| C3H/HeNCrl, male mice | Hepatocellular carcinoma | 8–10 weeks | 56Fe (0.6 GeV/u), 28Si (0.3 GeV/u), acute | Observed to age 800 d | [18] |
| Sprague-Dawley, female rats | Mammary carcinoma | 7 weeks | Fast neutrons mean energy 2.3 MeV, acute | Observed to age 90 weeks | [19] |
| B6CF1 ((C57BL/6 Bd x BALB/c Bd), male mice) | Hepatocellular carcinoma | 16 weeks | FN mean energy 2 MeV, acute and chronic | Observed to 1200 d | [7] |
| B6CF1 (C57BL/6 Bd x BALB/c Bd), female and male Mice) | Lung Adenocarcinoma | 16 weeks | FN mean energy 2 MeV, acute and chronic | Observed to 1200 d | [7] |
| BCF1, male mice | Hepatocellular carcinoma, all solid cancers | 3 months | FN mean energy 0.4 MeV, acute and fractionated | Lifespan | [8] |
| Saturation Model | Linear Model | |||||
|---|---|---|---|---|---|---|
| Tissue: Radiation | A | B, Gy−1 | AB, Gy−1 | Adjusted R2 | α, Linear, Gy−1 | Adjusted R2 |
| Lung Balb/c (F): FN (chronic) | 1.43 ± 0.365 (p < 0.0175) | 6.01 ± 3.29 (p < 0.1415) | 8.59 | 0.893 | 4.12 ± 0.47 (p < 0.003) | 0.883 |
| Lung B6CF1 (F): FN (fractions) | 1.24 ± 1.69 (p < 0.518) | 2.99 ± 7.04 (p < 0.7) | 3.71 | 0.585 | 3.25 ± 0.34 (p < 0.0007) | 0.918 |
| Lung B6CF1 (M): FN (fractions) | - | - | - | NC | 1.83 ± 0.115 (p < 0.0001) | 0.966 |
| Mammary Balb/c: FN (chronic) | 2.29 ± 0.068 (p < 0.0001) | 29.77 ± 3.36 (p < 0.0009) | 68.2 | 0.986 | 4.35 ± 2.85 (p < 0.19) | −1.17 |
| Mammary Balb/c: FN (acute) | 1.95 ± 0.258 (p < 0.0016) | 9.21 ± 2.75 (p < 0.029) | 17.96 | 0.9491 | 5.05 ± 1.21 (p < 0.0096) | 0.557 |
| Mammary SD Rats: 2 MeV neutrons (acute) | 1.81 ± 0.326 (p < 0.0052) | 4.99 ± 2.19 (p < 0.0857) | 9.03 | 0.867 | 1.804 ± 0.596 (p < 0.092) | 0.328 |
| Liver C3H (M): Si (0.3 GeV/u) and Fe (0.6 GeV/u) (acute) | 1.61 ± 0.2 (p < 0.0001) | 10.28 ± 4.9 (p < 0.0691) | 16.6 | 0.799 | 0.45 ± 0.96 (p < 0.652) | −5.14 |
| Liver CBA (M): Fe (1 GeV/u) (acute) | 1.72 ± 0.22 (p < 0.0044) | 5.33 ± 1.79 (p < 0.058) | 9.2 | 0.937 | 4.78 ± 0.53 * (p < 0.0032) | 0.906 |
| Liver B6CF1 (M): FN (acute and fractions) | 1.749 ± 0.362 (p < 0.0019) | 26.02 ± 24.96 (p < 0.332) | 45.5 | −0.23 | 1.603 ± 0.365 (p < 0.0023) | 0.521 |
| All solid B6CF1 (M): FN (acute and fractions) | 1.69 ± 0.39 (p < 0.0034) | 4.64 ± 2.4 (p < 0.097) | 7.84 | 0.825 | 2.69 ± 0.5 (p < 0.006) | 0.503 |
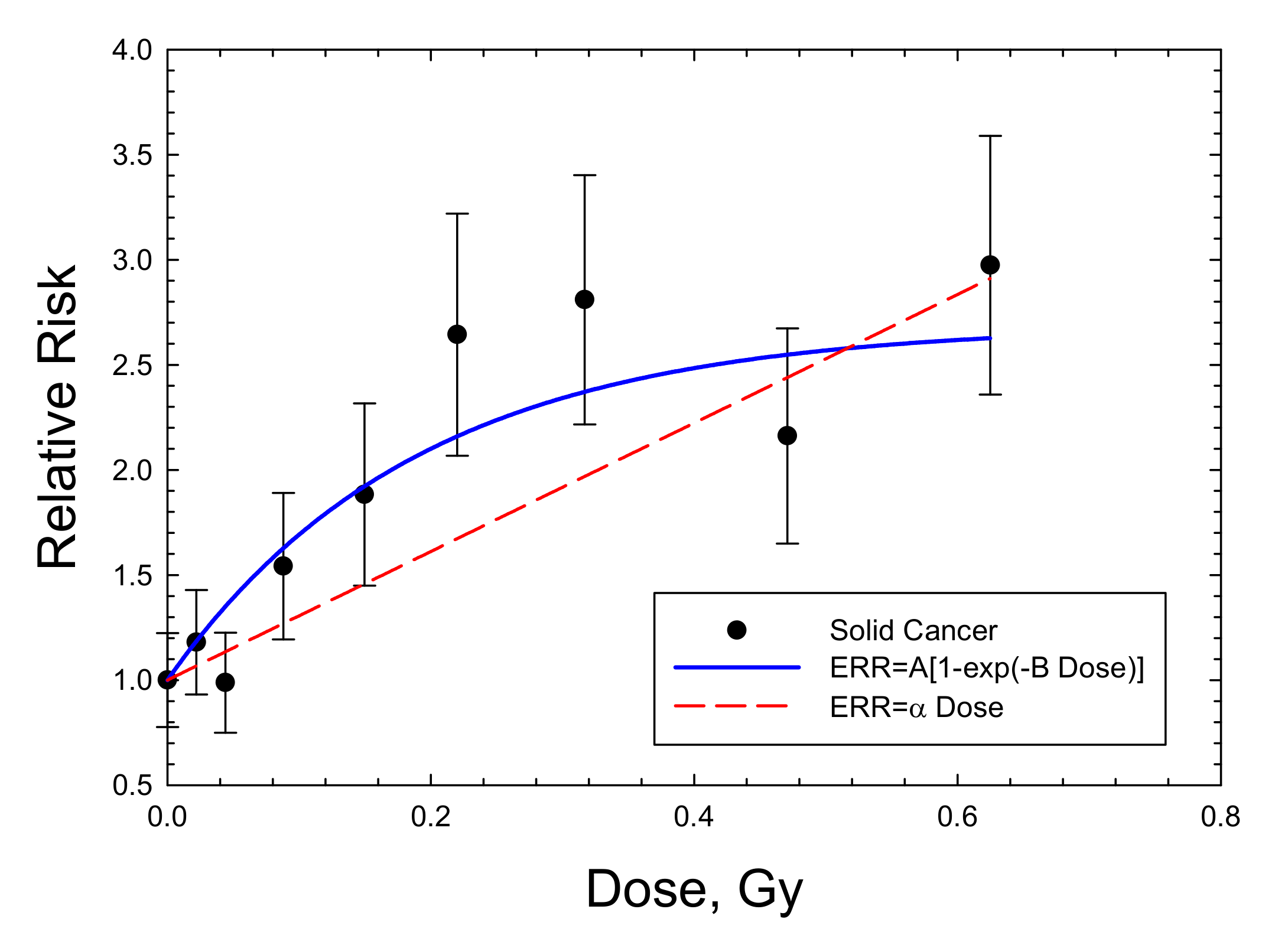
3. Discussion
4. Materials and Methods
Fission Neutron and Space Radiation Organ Exposures
5. Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Monson, R.; Cleaver, J.; Abrams, H.L.; Bingham, E.; Buffler, P.A.; Cardis, E.; Cox, R.; Davis, S.; Hutchinson, F.; Dewey, W.C.; et al. BEIR VII: Health Risks from Exposure to Low Levels of Ionizing Radiation; National Academy of Sciences Committee on the Biological Effects of Radiation; National Academy of Sciences Press: Washington, DC, USA, 2006. [Google Scholar]
- NCRP. Guidance on Radiation Received in Space Activities; NCRP Report 98; National Council on Radiation Protection and Measurements: Bethesda, MD, USA, 1989. [Google Scholar]
- NCRP. Recommendations of Dose Limits for Low Earth Orbit; NCRP Report 132; National Council on Radiation Protection and Measurements: Bethesda, MD, USA, 2000. [Google Scholar]
- Cucinotta, F.A. A new approach to reduce uncertainties in space radiation cancer risk predictions. PLoS ONE 2015, 10, e0120717. [Google Scholar] [CrossRef] [PubMed]
- NCRP. Relative Biological Effectiveness of Radiations of Different Quality; National Council on Radiation Protection and Measurements Report 104; NCRP: Bethesda, MD, USA, 1990. [Google Scholar]
- Ullrich, R.L. Tumor induction in BALB/c mice after fractionated or protracted exposures to fission-spectrum neutrons. Radiat. Res. 1984, 97, 587–597. [Google Scholar] [CrossRef] [PubMed]
- Storer, J.B.; Fry, R.J.M. On the shape of neutron dose-effect curves for radiogenic cancers and life shorting in mice. Radiat. Environ. Biophys. 1995, 34, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Di Majo, V.; Coppola, M.; Rebessi, S.; Saran, A.; Pazzaglia, S.; Pariset, L.; Covelli, V. Neutron-induced tumors in BC3F1 mice: Effects of dose fractionation. Radiat. Res. 1994, 138, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Cucinotta, F.A.; To, K.; Cacao, E. Predictions of space radiation fatality risk for exploration missions. Life Sci. Space Res. 2017, 13, 1–11. [Google Scholar] [CrossRef]
- Hoel, D.G. Comments on the DDREF estimate of the BEIR VII committee. Health Phys. 2015, 108, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Cucinotta, F.A.; Kim, M.Y.; Chappell, L. Space Radiation Cancer Risk Projections and Uncertainties—2012; NASA TP 2013-217375; NASA: Houston, TX, USA, 2013.
- Cucinotta, F.A.; Wilson, J.W. Initiation-promotion model for tumor prevalence from high energy and charge radiation. Phys. Med. Biol. 1994, 39, 1811–1831. [Google Scholar] [CrossRef]
- Alpen, E.L.; Powers-Risius, P.; Curtis, S.B.; DeGuzman, R. Tumorigenic potential of high-Z, high-LET charged particle radiations. Radiat. Res. 1993, 88, 132–143. [Google Scholar] [CrossRef]
- Chang, P.Y.; Cucinotta, F.A.; Bjornstad, K.A.; Bakke, J.; Rosen, C.J.; Du, N.; Fairchild, D.G.; Cacao, E.; Blakely, E.A. Harderian gland tumorigenesis: Low-dose- and LET-Response. Radiat. Res. 2016, 185, 448–459. [Google Scholar] [CrossRef]
- Fry, R.J.M.; Oak Ridge National Lab, Oak Ridge, TN, USA. Personal communication, 1993.
- Coggle, J.E. Lung tumour induction in mice after X-rays and neutrons. Int. J. Radiat. Biol. 1988, 53, 585–598. [Google Scholar] [CrossRef]
- Weil, M.M.; Bedford, J.S.; Bielefeldt-Ohmann, H.; Ray, F.A.; Genik, P.C.; Ehrhart, E.J.; Fallgren, C.M.; Hailu, F.; Battaglia, C.L.; Charles, B.; et al. Incidence of acute myeloid leukemia and hepatocellular carcinoma in mice irradiated with 1 GeV/nucleon 56Fe ions. Radiat. Res. 2009, 172, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Weil, M.M.; Ray, F.A.; Genik, P.C.; Yu, Y.; McCarthy, M.; Fallgren, C.M.; Ullrich, R.L. Effects of 28Si ions, 56Fe ions, and protons on the induction of murine acute myeloid leukemia and hepatocellular carcinoma. PLoS ONE 2014, 9, e104819. [Google Scholar] [CrossRef] [PubMed]
- Imaoka, T.; Nishimura, M.; Daino, K.; Hosoki, A.; Takabatake, M.; Kokubo, T.; Doi, K.; Showler, K.; Nishimura, Y.; Moriyama, H.; et al. Age modifies the effect of 2-MeV fast neutrons on rat mammary carcinogenesis. Radiat. Res. 2017, 188, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Cucinotta, F.A.; Cacao, E.; Kim, M.Y.; Saganti, P.B. Benchmarking risk predictions and uncertainties in the NSCR model of GCR cancer risks with revised low LET risk coefficients. Life Sci. Space Res. 2020, 27, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Cahoon, E.K.; Preston, D.L.; Pierce, D.A.; Grant, E.; Brenner, A.V.; Mabuchi, K.; Utada, M.; Ozasa, K. Lung, laryngeal and other respiratory cancer incidence among Japanese atomic bomb survivors: An updated analysis from 1958 through 2009. Radiat. Res. 2017, 187, 538–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- NCRP. Information Needed to Make Radiation Protection Recommendations for Space Missions beyond Low Earth Orbit; NCRP Report 153; NCRP: Bethesda, MD, USA, 2006. [Google Scholar]
- NCRP. Radiation Protection for Space Activities: Supplement to Previous Recommendations; National Council on Radiation Protection and Measurements Commentary 23; NCRP: Bethesda, MD, USA, 2014. [Google Scholar]
- Cucinotta, F.A.; Schimmerling, W.; Blakely, E.A.; Hei, T. A proposed change to astronaut exposure limits is a giant leap back for radiation orotection. Life Sci. Space Res. 2021, 31, 59–70. [Google Scholar] [CrossRef]
- Goodhead, D.T.; Nikjoo, H. Track structure analysis of ultrasoft X-rays compared to high- and low-LET radiations. Int. J. Radiat. Biol. 1989, 55, 513–529. [Google Scholar] [CrossRef]
- Durante, M.; George, K.; Wu, H.; Cucinotta, F.A. Karyotypes of human lymphocytes exposed to high-energy iron ions. Radiat. Res. 2002, 158, 581–590. [Google Scholar] [CrossRef]
- Kawata, T.; Durante, M.; Furusawa, Y.; George, K.; Ito, H.; Wu, H.; Cucinotta, F.A. Rejoining of isochromatid breaks induced by heavy ions in G2-phase normal human fibroblasts. Radiat. Res. 2001, 156, 598–602. [Google Scholar] [CrossRef]
- Schipler, A.; Iliakis, G. DNA double-strand break complexity levels and their possible contributions to the probability of error-prone processing and repair way choice. Nucleic Acids Res. 2013, 41, 7589–7605. [Google Scholar] [CrossRef] [Green Version]
- Sridharan, D.M.; Asaithamby, A.; Bailey, S.M.; Costes, S.V.; Doetsch, P.W.; Dynan, W.S.; Kronenberg, A.; Rithidech, K.N.; Saha, J.; Snijders, A.M.; et al. Understanding Cancer Development Processes Following HZE Particle Exposure: Roles of ROS, DNA Damage Repair, and Inflammation. Radiat. Res. 2015, 183, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Jella, K.K.; Jaafar, L.; Li, S.; Park, S.; Story, M.D.; Wang, H.; Wang, Y.; Dynan, W.S. Exposure to galactic cosmic radiation compromises DNA repair and increases the potential for oncogenic chromosomal rearrangements in bronchial epithelial cells. Sci. Rep. 2018, 8, 11038. [Google Scholar] [CrossRef] [PubMed]
- Barcellos-Hoff, M.H.; Mao, J.-H. HZE radiation non-targeted effects on the microenvironment that mediate mammary carcinogenesis. Front. Oncol. 2016, 6, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadhim, M.; Salomaa, S.; Wright, E.; Hildebrandt, G.; Belyakov, O.V.; Prise, K.M.; Little, M.P. Non-targeted effects of ionizing radiation- implications for low dose risk. Mutat. Res. 2013, 752, 84–98. [Google Scholar] [CrossRef] [Green Version]
- Illa-Bochaca, I.; Ouyang, H.; Tang, J.; Sebastiano, C.; Mao, J.-H.; Costes, S.V.; Demaria, S.; Barcellos-Hoff, M.H. Densely ionizing radiation acts via the microenvironment to promote aggressive Trp53-null mammary carcinomas. Cancer Res. 2014, 74, 7137–7148. [Google Scholar] [CrossRef] [Green Version]
- McConnel, A.M.; Konda, B.; Kirsch, D.G.; Stripp, B.R. Distal airway epithelial progenitor cells are radiosensitive to high-LET radiation. Sci. Rep. 2016, 6, 3345. [Google Scholar] [CrossRef]
- Peng, Y.; Brown, N.; Finnon, R.; Warner, C.L.; Liu, X.; Genik, P.C.; Callan, M.A.; Ray, F.A.; Borak, T.B.; Badie, C.; et al. Radiation leukemogenesis in mice: Loss of PU.1 on chromosome 2 in CBA and C57BL/6 mice after irradiation with 1 GeV/nucleon 56Fe, X rays or γ rays. Part. I. experimental observations. Radiat. Res. 2009, 171, 478–483. [Google Scholar]
- Ullrich, R.L.; Preston, R.J. Myeloid leukemia in male RFM mice following irradiation with fission spectrum neutrons or γ-rays. Radiat. Res. 1987, 109, 165–170. [Google Scholar] [CrossRef]
- Cucinotta, F.A.; Cacao, E. Predictions of cognitive detriments from galactic cosmic ray exposures to astronauts on exploration missions. Life Sci. Space Res. 2019, 25, 129–135. [Google Scholar] [CrossRef]
- NCRP. Fluence-Based and Microdosimetric Event-Based Methods for Radiation Protection in Space; NCRP Report 137; National Council on Radiation Protection and Measurements: Bethesda, MD, USA, 2001. [Google Scholar]
- NCRP. Extrapolation of Radiation Induced Cancer Risks from Nonhuman Experimental Systems to Humans; NCRP Report 150; National Council on Radiation Protection and Measurements: Bethesda, MD, USA, 2005. [Google Scholar]
- Carnes, B.A.; Olshansky, S.J.; Grahn, D. An interspecies prediction of the risk of radiation induced mortality. Radiat. Res. 1998, 149, 487–492. [Google Scholar] [CrossRef]
- Cucinotta, F.A.; Chappell, L.J.; Kim, M.Y.; Wang, M. Radiation carcinogenesis risk assessments for never-smokers. Health Phys. 2012, 103, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Cacao, E.; Hada, M.; Saganti, P.B.; George, K.A.; Cucinotta, F.A. Relative biological effectiveness of HZE particles for chromosomal aberrations and other Surrogate Cancer Risk Endpoints. PLoS ONE 2016, 11, e0153998. [Google Scholar] [CrossRef] [PubMed]
- Cucinotta, F.A.; Nikjoo, H.; Goodhead, D.T. Applications of amorphous track models in radiobiology. Radiat. Environ. Biophys. 1999, 38, 81–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodhead, D.T. Relationship of radiation track structure to biological effect: A re-interpretation of the parameters of the Katz model. Nucl. Tracks Radiat. Meas. 1989, 116, 177–184. [Google Scholar] [CrossRef]
- Vogel, H.H. Mammary gland neoplasms after fission neutron irradiation. Nature 1969, 222, 1279–1281. [Google Scholar] [CrossRef]
- Imaoka, T.; Nishimura, M.; Kakinuma, S.; Hatano, Y.; Ohmachi, Y.; Yoshinaga, S.; Kawano, A.; Maekawa, A.; Shimada, Y. High relative biological effectiveness of carbon ion irradiation on induction of rat mammary carcinoma and its lack of H-ras and Tp53 mutations. Int. J. Radiat. Oncol. Biol. Phys. 2007, 69, 194–203. [Google Scholar] [CrossRef]
- Dicello, J.F.; Christian, A.; Cucinotta, F.A.; Gridley, D.S.; Kathirithamby, R.; Mann, J.; Markham, A.R.; Moyers, M.F.; Novak, G.R.; Piantadosi, S.; et al. In vivo Mammary tumorigenesis in the Sprague–Dawley rat and microdosimetric correlates. Phys. Med. Biol. 2004, 49, 3817–3830. [Google Scholar] [CrossRef]
- Fry, R.J.M. Experimental radiation carcinogenesis: What have we learned? Radiat. Res. 1981, 87, 224–239. [Google Scholar] [CrossRef] [Green Version]
- Storer, J.B.; Mitchell, T.J.; Fry, R.J.M. Extrapolation of the relative risk of radiogenic neoplasms across mouse strains and to man. Radiat. Res. 1988, 114, 331–353. [Google Scholar] [CrossRef]
- Demant, P. Cancer susceptibility in the mouse: Genetics, biology, and implications for human cancer. Nat. Rev. Genet. 2003, 4, 721–734. [Google Scholar] [CrossRef]
- Evers, B.; Jonkers, J. Mouse models of BRCA1 and BRCA2 deficiency: Past lessons, current understanding and future prospects. Oncogene 2006, 25, 5885–5897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parmar, H.; Cuncha, G.R. Epithelial-stromal interactions in the mouse and human mammary gland in vivo. Endocr. Relat. Cancer 2004, 11, 437–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suman, S.; Kumar, S.; Moon, B.H.; Strawn, S.J.; Thakor, H.; Fan, Z.; Shay, J.W.; Fornace, A.J., Jr.; Datta, K. Relative biological effectiveness of energetic heavy ions for intestinal tumorigenesis shows male preponderance and radiation type and energy dependence in APC 1638N/+ mice. Int. J. Radiat. Oncol. Biol. Phys. 2016, 95, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Edmondson, E.F.; Gatti, D.M.; Ray, F.A.; Garcia, E.L.; Fallgren, C.M.; Kamstock, D.A.; Weil, M.M. Genomic mapping in outbred mice reveals overlap in genetic susceptibility for HZE ion– and γ-ray–induced tumors. Sci. Adv. 2020, 6, eaax5940. [Google Scholar] [CrossRef] [Green Version]
- Ullrich, R.L.; Jernigan, M.C.; Storer, J.B. Neutron carcinogenesis dose and dose-rate effects in BALB/c mice. Radiat. Res. 1977, 72, 487–498. [Google Scholar] [CrossRef]
- Ullrich, R.L. Tumor induction in BALB/c female mice after fission neutrons or γ irradiation. Radiat. Res. 1983, 93, 506–515. [Google Scholar] [CrossRef]
- Grant, E.J.; Brenner, A.; Sugiyama, H.; Sakata, R.; Sadakane, A.; Utada, M.; Cahoon, E.K.; Milder, C.M.; Soda, M.; Cullings, H.M.; et al. Solid cancer incidence among the Life-span study of atomic-bomb survivors: 1958–2009. Radiat. Res. 2017, 187, 513–537. [Google Scholar] [CrossRef] [Green Version]
- Brenner, A.V.; Preston, D.L.; Sakata, R.; Sugiyama, H.; De Gonzalez, A.B.; French, B.; Utada, M.; Cahoon, E.K.; Sadakane, A.; Ozasa, K.; et al. Incidence of breast cancer in the life span study of atomic bomb survivors: 1958–2009. Radiat. Res. 2018, 190, 433–444. [Google Scholar] [CrossRef] [Green Version]
- Arias, E.; Bastian, B.; Xu, J.; Tejada-Vera, B. National Vital Statistics Report, US State Life Tables, 2018; Center of Disease Control and Prevention (CDC): Atlanta, GA, USA, 2021; Volume 70.
- Midthune, D.N.; Fay, M.P.; Clegg, L.X.; Feuer, E.J. Modeling reporting delays and reporting corrections in cancer registry data. J. Am. Stat. Assoc. 2005, 100, 61–70. [Google Scholar] [CrossRef]
- SEER Explorer: An Interactive Website for SEER Cancer Statistics [Internet]. Surveillance Research Program, National Cancer Institute. Available online: https://seer.cancer.gov/explorer/ (accessed on 15 April 2021).
- Pak, S.; Cucinotta, F.A. Comparison between PHITS and GEANT4 simulations of the heavy ion beams at the BEVALAC at LBNL and the booster accelerator at BNL. Life Sci. Space Res. 2021, 29, 38–45. [Google Scholar] [CrossRef]
- Storer, J.B.; Serranoe, J.; Darden, E.B., Jr.; Jernigan, M.C.; Ullrich, R.L. Life shortening in RFM mice and BALB/c mice as a function of radiation quality, dose, and dose rate. Radiat. Res. 1979, 78, 122–161. [Google Scholar] [CrossRef] [PubMed]
- Edwards, A.A.; Dennis, J.A. The calculation of charged particle fluence and LET spectra for the irradiation of biologically significant materials by neutrons. Phys. Med. Biol 1975, 20, 395–409. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.W.; Townsend, L.W.; Shinn, J.L.; Cucinotta, F.A.; Costen, R.C.; Badavi, F.F.; Lamkin, S.L. Galactic Cosmic Ray Transport Methods, Past, Present, and Future. Adv. Space Res. 1994, 10, 841–852. [Google Scholar] [CrossRef]
- Cucinotta, F.A.; Kim, M.Y.; Schneider, S.I.; Hassler, D.M. Description of light ion production cross sections and fluxes on the Mars surface using the QMSFRG model. Radiat. Environ. Biophys. 2007, 46, 101–106. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.Y.; Rusek, A.; Cucinotta, F.A. Issues in ground-based GCR simulation for space radiobiology. Front. Radiat. Oncol. 2015, 5, 122. [Google Scholar] [CrossRef] [Green Version]
| Source of Uncertainty | Conventional Model | DERR Model |
|---|---|---|
| Statistical | Epidemiology data, experimental for gamma-rays, experimental for ions. | Experimental for ions alone. |
| Dosimetry | Epidemiology data, and in accelerator studies with animals. | In accelerator studies with animals. |
| Time period | Epidemiology data, including changes in background cancer rates over decades due to changes in environmental, dietary, and genetic factors. | Small if models developed in reasonable research time period. |
| Genetic and environmental factors | Likely distinct in epidemiology data from modern day radiation workers including astronauts. Considered in experimental design for gamma-rays and ions. | Considered in experimental design for ions. |
| Dose and Dose-rate | Extrapolation of epidemiology data, and experimental data for gamma-rays to risks of ions at chronic low doses. Uncertainty in shape of response at low dose. | Extrapolation of experimental data for ions to chronic low doses. |
| Range of doses and ion types considered | Limited by investments in experiments performed. | Limited by investments in experiments performed. |
| Extrapolation of tissue-specific factors from epidemiology or mouse studies to humans. | For both gamma-rays and ions. Transfer model uncertainties. | For ions. |
| Space dosimetry | Modest uncertainties in organ dose equivalent (<15%). Dosimetric methods developed. | Modest uncertainties in organ dose equivalent (<15%), but dosimetric verification methods have not been developed. |
| Tissue | AT | BT, Gy−1 (Σ0, μm2) |
|---|---|---|
| Lung (Females) | 1.4 ± 0.4 | 5 ± 2 (250) |
| Liver (Males) | 1.65 ± 0.2 | 7.5 ± 3 (400) |
| Breast (Females) | 2.1 ± 0.3 | 20 ± 5 (1000) |
| ΣT, μm−1 | ||
| Lung (Females) | 148 | |
| Lung (Males) | 83 |
| Cancer Site | NSCR-2020 | NSCR-2020 with NTE | DERR Saturation Model | DERR Linear Model |
|---|---|---|---|---|
| Age at Exposure = 35 years | ||||
| Lung Females | 1.98 [0.52, 6.0] | 4.7 [1.4, 10.7] | 0.92 [0.2, 3.1] | 0.42 [0.07,1.43] |
| Lung Males | 0.70 [0.18, 2.0] | 1.64 [0.49, 3.84] | - | 0.24 [0.04,0.83] |
| Breast Females | 4.2 [0.81, 13.2] | 9.7 [2.2, 23.5] | 8.5 [2.1, 21.5] | - |
| Liver Males | 0.11 [0.03, 0.34] | 0.27 [0.08,0.63] | 0.24 [0.05, 0.61] | - |
| Age at Exposure = 50 years | ||||
| Lung Females | 1.99 [0.51, 6.03] | 4.68 [1.4, 10.7] | 0.88 [0.17, 2.89] | 0.41 [0.02,0.23] |
| Lung Males | 0.73 [0.19, 2.24] | 1.57 [0.48, 3.67] | - | 0.24 [0.04, 0.86] |
| Breast Females | 2.98 [0.57, 9.41] | 7.0 [1.53,17.2] | 6.8 [ 1.67, 16.2] | - |
| Liver Males | 0.07 [0.02, 0.22] | 0.17 [0.05, 0.40] | 0.2 [0.05, 0.44] | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cucinotta, F.A. Flying without a Net: Space Radiation Cancer Risk Predictions without a Gamma-ray Basis. Int. J. Mol. Sci. 2022, 23, 4324. https://doi.org/10.3390/ijms23084324
Cucinotta FA. Flying without a Net: Space Radiation Cancer Risk Predictions without a Gamma-ray Basis. International Journal of Molecular Sciences. 2022; 23(8):4324. https://doi.org/10.3390/ijms23084324
Chicago/Turabian StyleCucinotta, Francis A. 2022. "Flying without a Net: Space Radiation Cancer Risk Predictions without a Gamma-ray Basis" International Journal of Molecular Sciences 23, no. 8: 4324. https://doi.org/10.3390/ijms23084324
APA StyleCucinotta, F. A. (2022). Flying without a Net: Space Radiation Cancer Risk Predictions without a Gamma-ray Basis. International Journal of Molecular Sciences, 23(8), 4324. https://doi.org/10.3390/ijms23084324