
Tolerogenic IDO1+CD83− Langerhans Cells in Sentinel Lymph Nodes of Patients with Melanoma

Abstract
:1. Introduction
2. Results
2.1. Clinical Features of Patients with Melanoma
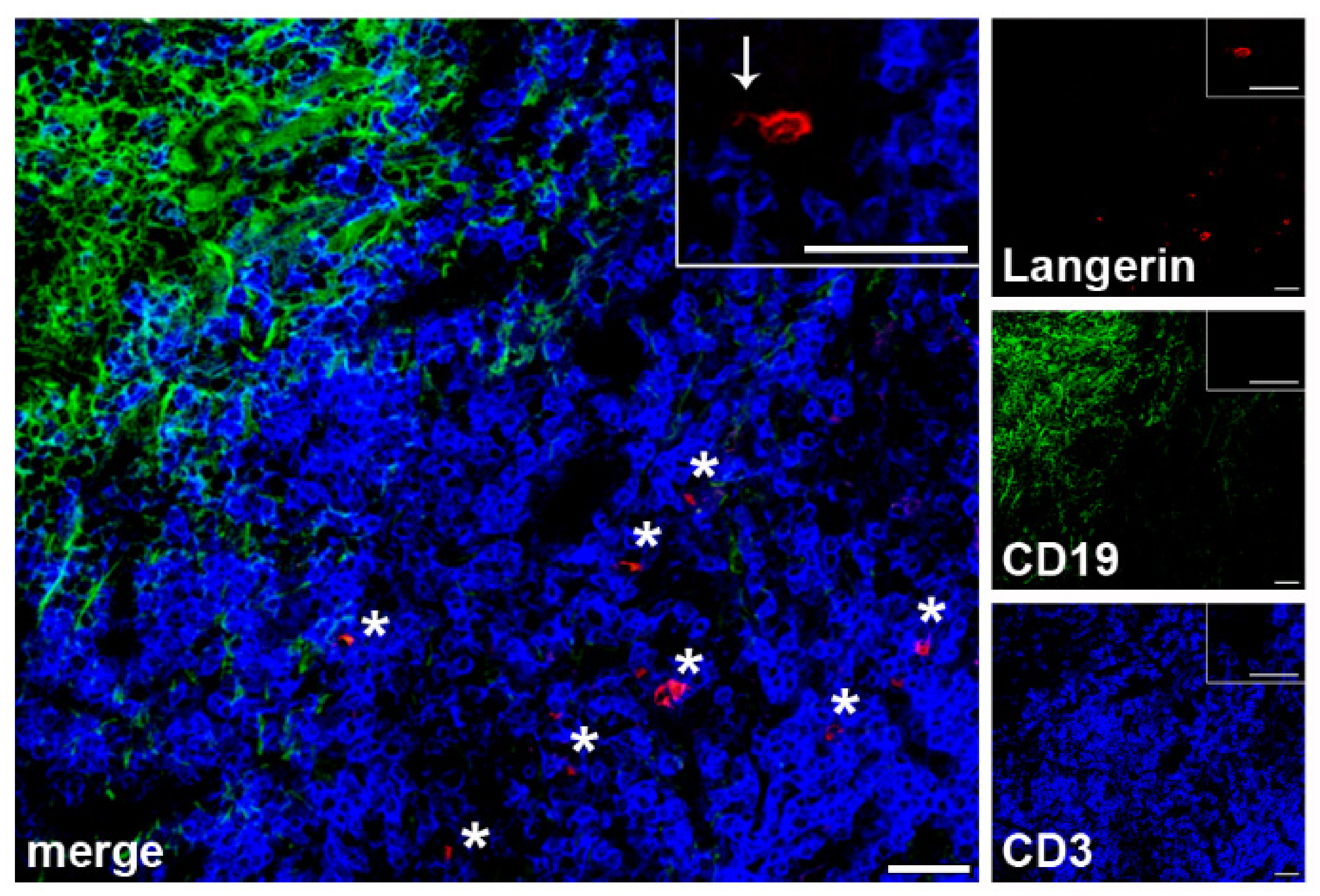
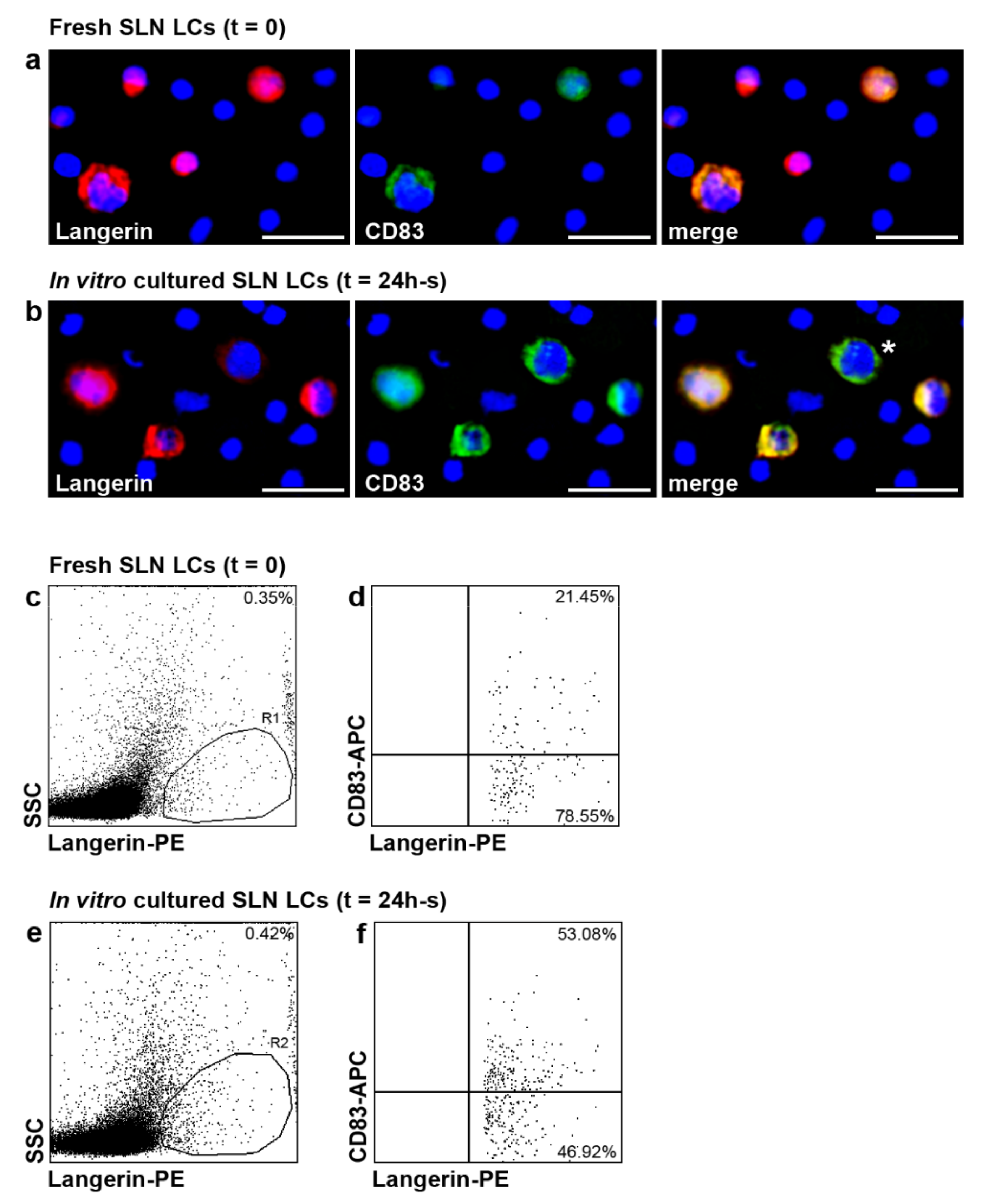
2.2. Localization of LCs within SLN of Patients with Melanoma
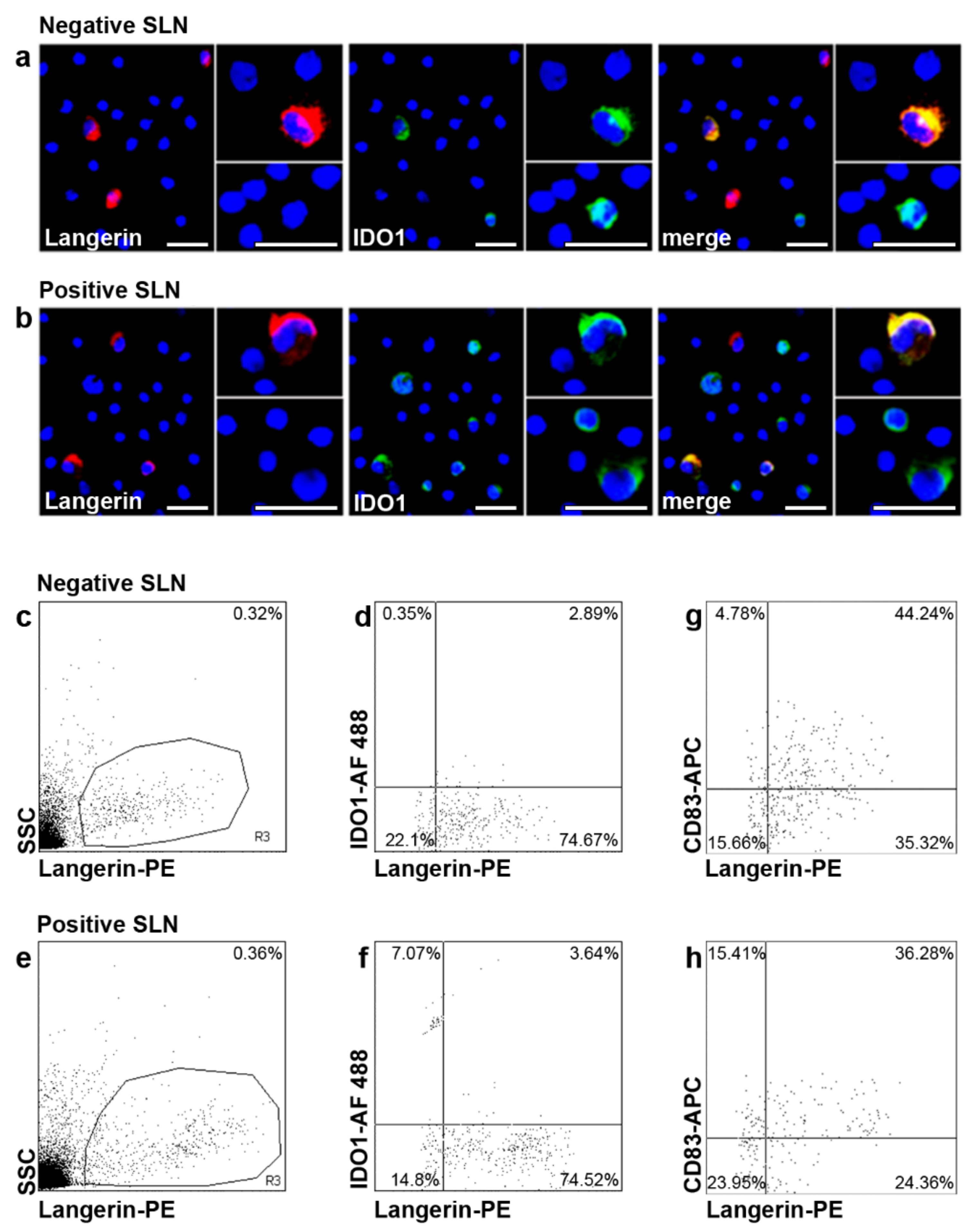
2.3. IDO1 Expression in SLN LC of Patients with Melanoma
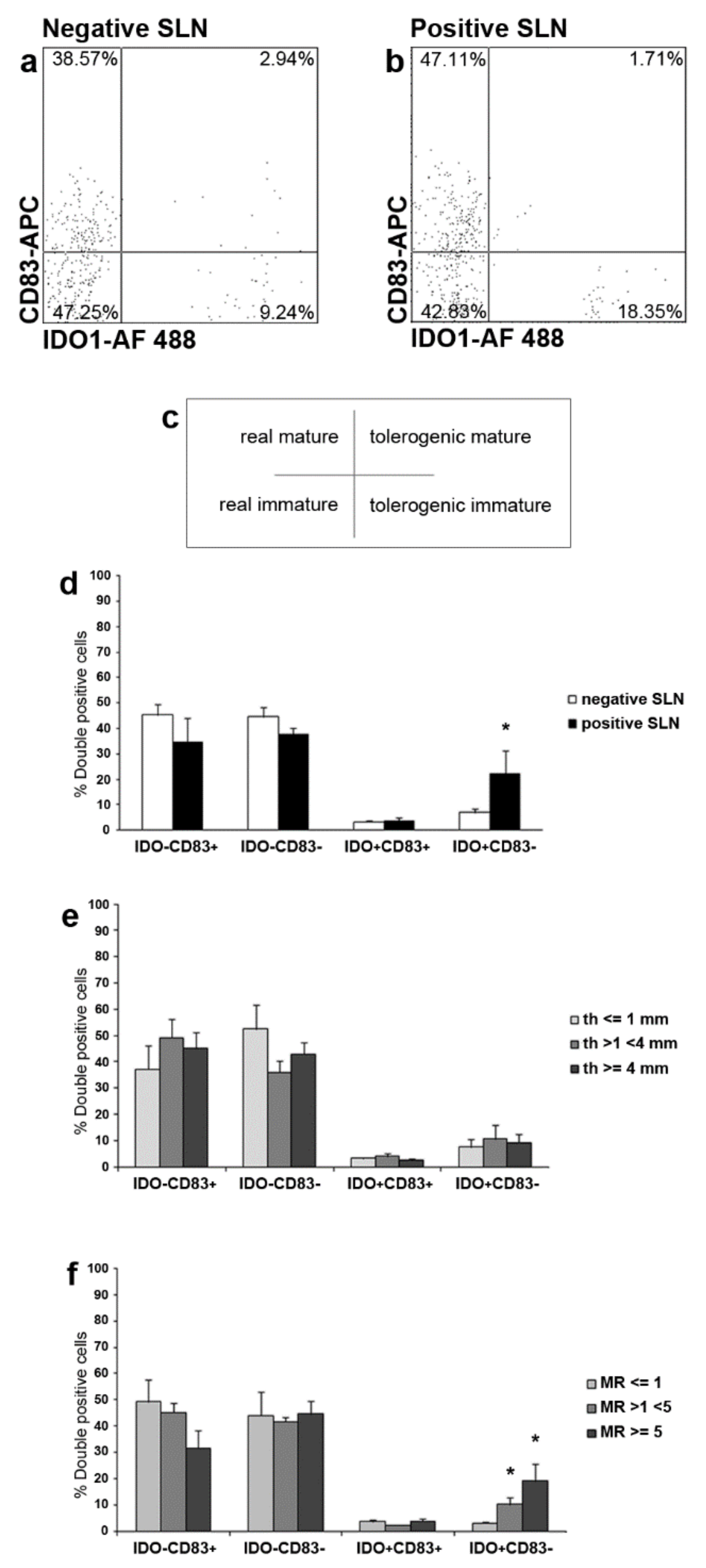
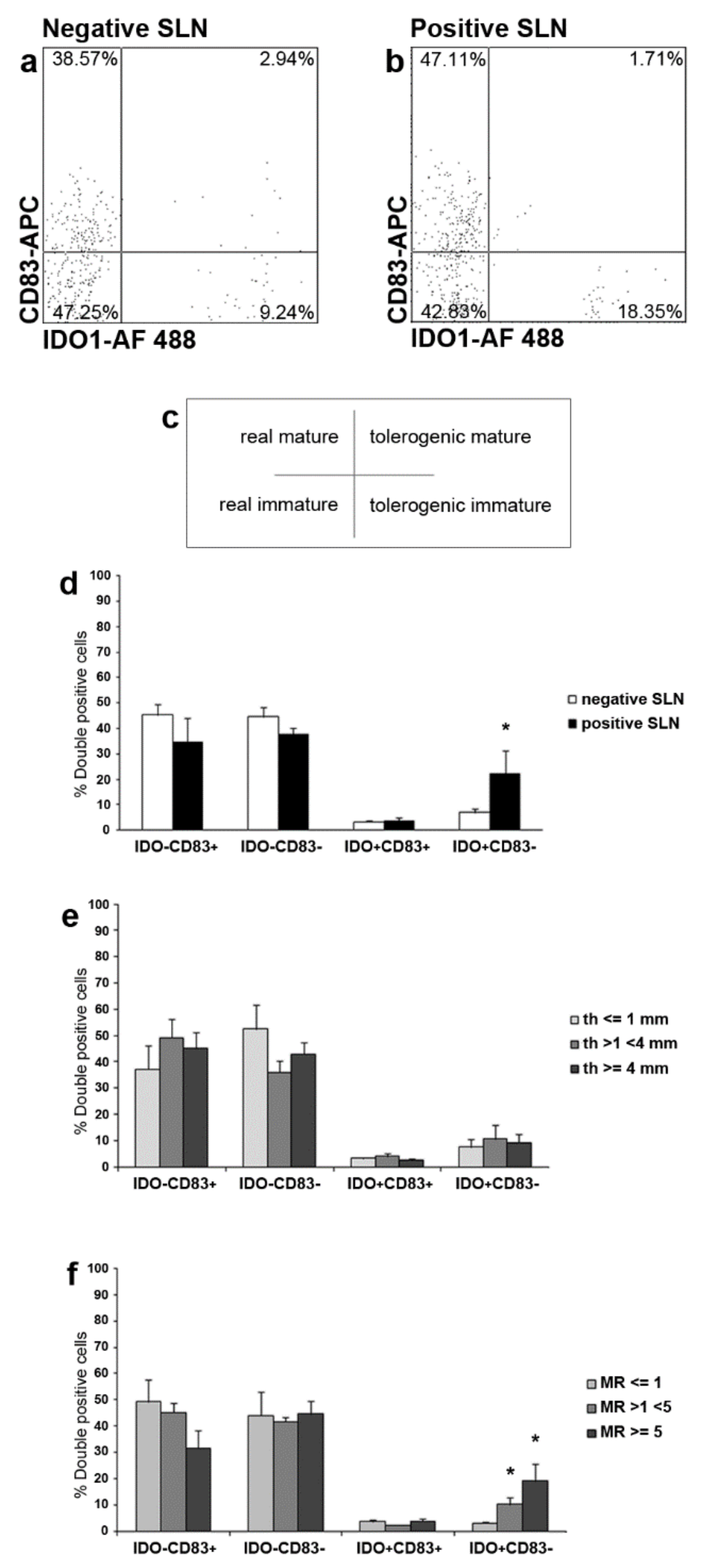
2.4. IDO1 and CD83 Concomitant Expression Analysis Identified Four SLN LCs Subsets
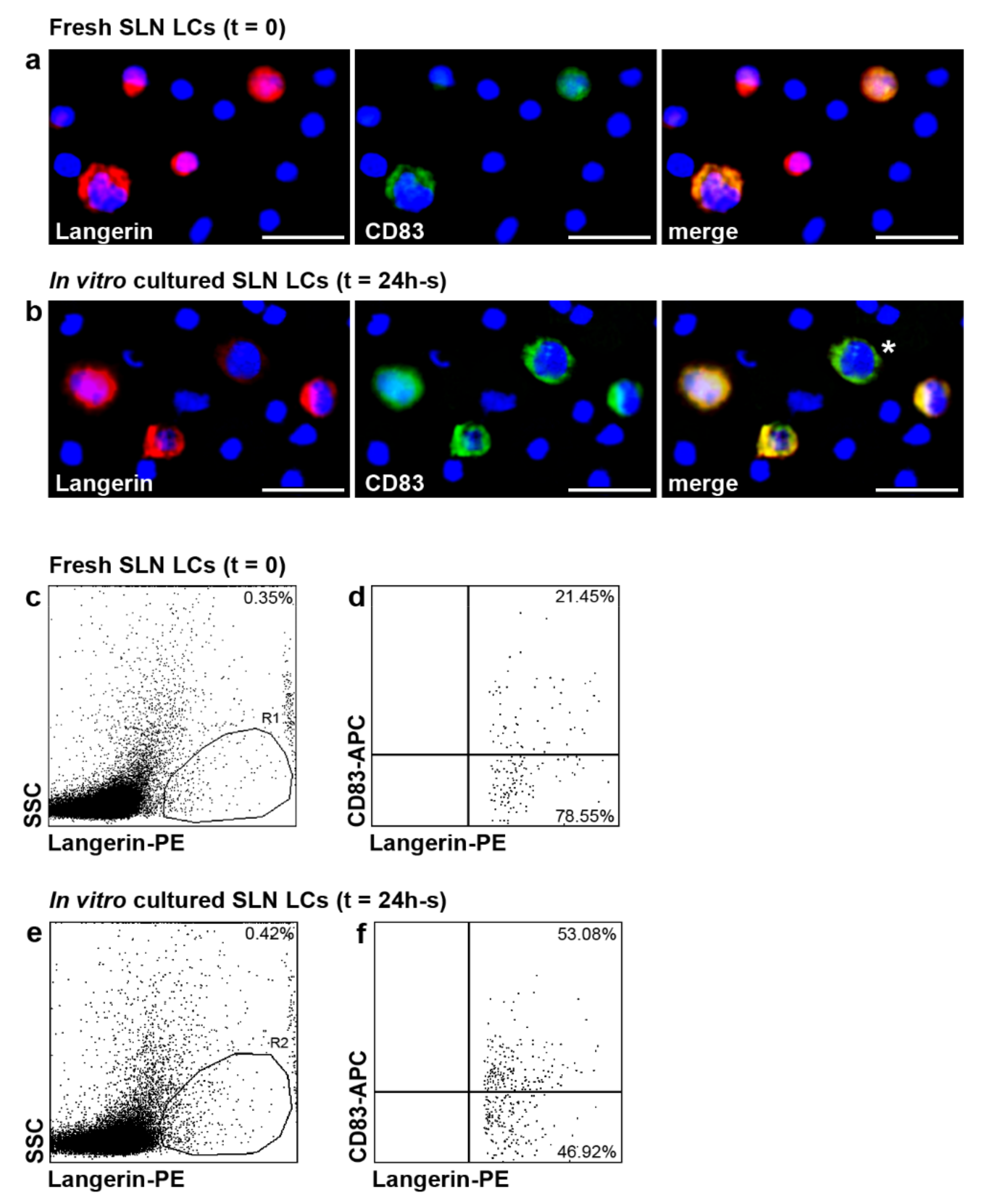
2.5. CD83 Expression in SLN LCs of Patients with Melanoma after In Vitro Stimulation with Inflammatory Cytokines
3. Discussion
4. Materials and Methods
4.1. Human Samples
4.2. Antibodies
4.3. In Vitro Stimulation with Inflammatory Cytokines
4.4. Fluorescent Immunohistochemistry and Immunocytochemistry
4.5. Flow Cytometry Analyses
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schreiber, R.D.; Old, L.J.; Smyth, M.J. Cancer Immunoediting: Integrating Immunity’s Roles in Cancer Suppression and Promotion. Science 2011, 331, 1565–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morton, D.L. Lymphatic mapping and sentinel lymphadenectomy for melanoma: Past, present, and future. Ann. Surg. Oncol. 2001, 8, 22S–28S. [Google Scholar]
- Cochran, A.J.; Huang, R.; Lee, J.; Itakura, E.; Leong, S.P.L.; Essner, R. Tumor-induced immune modulation of sentinel lymph nodes. Nat. Rev. Immunol. 2006, 6, 659–670. [Google Scholar] [CrossRef] [PubMed]
- Gerlini, G.; Tun-Kyi, A.; Dudli, C.; Burg, G.; Pimpinelli, N.; Nestle, F.O. Metastatic melanoma secreted IL-10 down-regulates CD1 molecules on dendritic cells in metastatic tumor lesions. Am. J. Pathol. 2004, 165, 1853–1863. [Google Scholar] [CrossRef] [Green Version]
- Polak, M.E.; Borthwick, N.J.; Gabriel, F.G.; Johnson, P.; Higgins, B.; Hurren, J.; McCormick, D.; Jager, M.J.; Cree, I.A. Mechanisms of local immunosuppression in cutaneous melanoma. Br. J. Cancer 2008, 96, 1879–1887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cochran, A.J.; Morton, D.L.; Stern, S.; Lana, A.M.; Essner, R.; Wen, D.R. Sentinel lymph nodes show profound downregulation of antigen-presenting cells of the paracortex: Implications for tumor biology and treatment. Mod. Pathol. 2001, 14, 604–608. [Google Scholar] [CrossRef]
- Essner, R.; Kojima, M. Dendritic cell function in sentinel nodes. Oncology 2001, 16, 27–31. [Google Scholar]
- Lee, J.H.; Torisu-Itakara, H.; Cochran, A.J.; Kadison, A.; Huynh, Y.; Morton, D.L.; Essner, R. Quantitative analysis of melanoma-induced cytokine-mediated immunosuppression in melanoma sentinel nodes. Clin. Cancer Res. 2005, 11, 107–112. [Google Scholar]
- Angel, C.E.; Chen, C.J.J.; Horlacher, O.C.; Winkler, S.; John, T.; Browning, J.; MacGregor, D.; Cebon, J.; Dunbar, P.R. Distinctive localization of antigen-presenting cells in human lymph nodes. Blood 2009, 113, 1257–1267. [Google Scholar] [CrossRef]
- Gerlini, G.; Urso, C.; Mariotti, G.; Di Gennaro, P.; Palli, D.; Brandani, P.; Salvadori, A.; Pimpinelli, N.; Reali, U.M.; Borgognoni, L. Plasmacytoid dendritic cells represent a major dendritic cell subset in sentinel lymph nodes of melanoma patients and accumulate in metastatic nodes. Clin. Immunol. 2007, 125, 184–193. [Google Scholar] [CrossRef]
- Gerlini, G.; Di Gennaro, P.; Mariotti, G.; Urso, C.; Chiarugi, A.; Caporale, R.; Pimpinelli, N.; Borgognoni, L. Human Langerhans cells are immature in melanoma sentinel lymph nodes. Blood 2012, 119, 4807–4808. [Google Scholar] [CrossRef] [Green Version]
- Van de Ven, R.; van den Hout, M.F.; Lindenberg, J.J.; Sluijter, B.J.; van Leeuwen, P.A.; Lougheed, S.M.; Meijer, S.; van den Tol, M.P.; Scheper, R.J.; de Gruijl, T.D. Characterization of four conventional dendritic cell subsets in human skin-draining lymph nodes in relation to T-cell activation. Blood 2011, 118, 2502–2510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munn, D.H.; Mellor, A.L. Indoleamine 2,3-dioxygenase and tumor-induced tolerance. J. Clin. Investig. 2007, 117, 1147–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munn, D.H.; Mellor, A.L. Indoleamine 2,3 dioxygenase and metabolic control of immune responses. Trends Immunol. 2013, 34, 137–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jandus, C.; Speiser, D.; Romero, P. Recent advances and hurdles in melanoma immunotherapy. Pigment Cell Melanoma Res. 2009, 22, 711–723. [Google Scholar] [CrossRef]
- Jacobs, J.F.; Nierkens, S.; Figdor, C.G.; De Vries, I.J.; Adema, G.J. Regulatory T cells in melanoma: The final hurdle towards effective immunotherapy? Lancet Oncol. 2012, 13, 32–42. [Google Scholar] [CrossRef]
- Di Gennaro, P.; Gerlini, G.; Caporale, R.; Sestini, S.; Brandani, P.; Urso, C.; Pimpinelli, N.; Borgognoni, L. T regulatory cells mediate immunosuppresion by adenosine in peripheral blood, sentinel lymph node and TILs from melanoma patients. Cancer Lett. 2018, 417, 124–130. [Google Scholar] [CrossRef]
- Munn, D.H.; Sharma, M.D.; Lee, J.R.; Jhaver, K.G.; Johnson, T.S.; Keskin, D.B.; Marshall, B.; Chandler, P.; Antonia, S.J.; Burgess, R.; et al. Potential regulatory function of human dendritic cells expressing indoleamine 2,3-dioxygenase. Science 2002, 297, 1867–1870. [Google Scholar] [CrossRef]
- Munn, D.H.; Sharma, M.D.; Hou, D.; Baban, B.; Lee, J.R.; Antonia, S.J.; Messina, J.L.; Chandler, P.; Koni, P.A.; Mellor, A.L. Expression of indoleamine 2,3-dioxygenase by plasmacytoid dendritic cells in tumor-draining lymph nodes. J. Clin. Investig. 2004, 114, 280–290. [Google Scholar] [CrossRef] [Green Version]
- Gerlini, G.; Di Gennaro, P.; Mariotti, G.; Urso, C.; Chiarugi, A.; Pimpinelli, N.; Borgognoni, L. Indoleamine 2,3-dioxygenase+ cells correspond to the BDCA2+ plasmacytoid dendritic cells in human melanoma sentinel nodes. J. Investig. Dermatol. 2010, 130, 898–901. [Google Scholar] [CrossRef] [Green Version]
- Dzionek, A.; Sohma, Y.; Nagafune, J.; Cella, M.; Colonna, M.; Facchetti, F.; Günther, G.; Johnston, I.; Lanzavecchia, A.; Nagasaka, T.; et al. BDCA-2, a novel plasmacytoid dendritic cell-specific type II Ctype lectin, mediates antigen capture and is a potent inhibitor of interferon alpha/beta induction. J. Exp. Med. 2001, 194, 1823–1834. [Google Scholar] [CrossRef] [PubMed]
- Palucka, K.; Banchereau, J. Cancer immunotherapy via dendritic cells. Nat. Rev. Cancer 2012, 12, 265–277. [Google Scholar] [CrossRef] [PubMed]
- Speeckaert, R.; Vermaelen, K.; Van Geel, N.; Autier, P.; Lambert, J.; Haspeslagh, M.; Van Gele, M.; Thielemans, K.; Neyns, B.; Roche, N.; et al. Indoleamine 2,3-dioxygenase, a new prognostic marker in sentinel lymph nodes of melanoma patients. Eur. J. Cancer 2012, 48, 2004–2011. [Google Scholar] [CrossRef]
- Munn, D.H.; Mellor, A.L. IDO in the Tumor Microenvironment: Inflammation, Counter-Regulation, and Tolerance. Trends Immunol. 2016, 37, 193–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prendergast, G.C.; Mondal, A.; Dey, S.; Laury-Kleintop, L.D.; Muller, A.J. Inflammatory Reprogramming with IDO1 Inhibitors: Turning Immunologically Unresponsive ‘Cold’ Tumors ‘Hot’. Trends Cancer 2018, 4, 38–58. [Google Scholar] [CrossRef] [PubMed]
- Brody, J.R.; Costantino, C.L.; Berger, A.C.; Sato, T.; Lisanti, M.P.; Yeo, C.J.; Emmons, R.V.; Witkiewicz, A.K. Expression of indoleamine 2,3-dioxygenase in metastatic malignant melanoma recruits regulatory T cells to avoid immune detection and affects survival. Cell Cycle 2009, 15, 1930–1934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubel, F.; Kern, J.S.; Technau-Hafsi, K.; Uhrich, S.; Thoma, K.; Häcker, G.; Von Bubnoff, N.; Meiss, F.; Von Bubnoff, D. Indoleamine 2,3-Dioxygenase Expression in Primary Cutaneous Melanoma Correlates with Breslow Thickness and Is of Significant Prognostic Value for Progression-Free Survival. J. Investig. Dermatol. 2018, 138, 679–687. [Google Scholar] [CrossRef] [Green Version]
- Von Bubnoff, D.; Bausinger, H.; Matz, H.; Kock, S.; Hacker, G.; Takikawa, O.; Bieber, T.; Hanau, D.; De la Salle, H. Human epidermal Langerhans cells express the immuno regulatory enzyme indoleamine 2,3-dioxygenase. J. Investig. Dermatol. 2004, 123, 298–304. [Google Scholar] [CrossRef] [Green Version]
- Di Gennaro, P.; Romoli, M.R.; Gerlini, G.; D’Amico, M.; Brandani, P.; Pimpinelli, N.; Borgognoni, L. IDO and CD83 expression in human epidermal Langerhans cells. J. Dermatol. Sci. 2014, 73, 172–174. [Google Scholar] [CrossRef]
- Stoitzner, P.; Green, L.K.; Jung, J.Y.; Price, K.M.; Tripp, C.H.; Malissen, B.; Kissenpfennig, A.; Hermans, I.F.; Ronchese, F. Tumor immunotherapy by epicutaneous immunization requires langerhans cells. J. Immunol. 2008, 180, 1991–1998. [Google Scholar] [CrossRef] [Green Version]
- Von Bubnoff, D.; Scheler, M.; Wilms, H.; Fimmers, R.; Bieber, T. Identification of IDO positive and IDO-negative human dendritic cells after activation by various proinflammatory stimuli. J. Immunol. 2011, 186, 6701–6709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, W.; Lee, S.H.; Lu, J. CD83 is preformed inside monocytes, macrophages and dendritic cells, but it is only stably expressed on activated dendritic cells. Biochem. J. 2005, 385, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Qin, Z.; Schwartzkopff, J.; Pradera, F.; Kammertoens, T.; Seliger, B.; Pircher, H.; Blankenstein, T. A critical requirement of interferon gamma-mediated angiostasis for tumor rejection by CD8+ T cells. Cancer Res. 2003, 63, 4095–4100. [Google Scholar] [PubMed]
- Koch, S.; Stroisch, T.J.; Vorac, J.; Herrmann, N.; Leib, N.; Schnautz, S.; Kirins, H.; Förster, I.; Weighardt, H.; Bieber, T. AhR mediates an anti-inflammatory feedback mechanism in human Langerhans cells involving FcεRI and IDO. Allergy 2017, 72, 1686–1693. [Google Scholar] [CrossRef]
- Borgognoni, L.; Urso, C.; Vaggelli, L.; Brandani, P.; Gerlini, G.; Reali, U.M. Sentinel node biopsy procedures with an analysis of recurrence patterns and prognosis in melanoma patients: Technical advantages using computer-assisted gamma probe with adjustable collimation. Melanoma Res. 2004, 14, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Vuylsteke, R.J.; Van Leeuwen, P.A.; Meijer, S.; Wijnands, P.G.; Statius Muller, M.G.; Busch, D.H. Sampling tumor-draining lymph nodes for phenotypic and functional analysis of dendritic cells and T cells. Am. J. Pathol. 2002, 16, 19–26. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patients | Positive SLN | Negative SLN | Total SLN | |
|---|---|---|---|---|
| Stage III | 8 | 10 | ||
| Stage I–II | 36 | 40 | ||
| total | 44 | 50 | ||
| Sex | ||||
| Men | 20 | 4 | 16 | |
| Women | 24 | 4 | 20 | |
| Age (mean, years) | 62 years range 27–80 | 55 years range 21–93 | ||
| Breslow thickness (mean, mm) | 4.19 mm range 1.1–9.8 | 2.18 mm range 0.5–8.1 | ||
| Lesion Type * | ||||
| SSM | 30 | 4 | 26 | |
| NM | 12 | 4 | 8 | |
| ALM | 2 | 2 | ||
| Total | 44 | 8 | 36 | |
| Mitotic rate (mean) | 5.2 (2.0–12) | 1.3 (0.0–9) | ||
| Mean interval ** (mean, days) | 39 days range 0–61 | 41 days range 0–71 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gerlini, G.; Di Gennaro, P.; Pimpinelli, N.; Sestini, S.; Borgognoni, L. Tolerogenic IDO1+CD83− Langerhans Cells in Sentinel Lymph Nodes of Patients with Melanoma. Int. J. Mol. Sci. 2022, 23, 3441. https://doi.org/10.3390/ijms23073441
Gerlini G, Di Gennaro P, Pimpinelli N, Sestini S, Borgognoni L. Tolerogenic IDO1+CD83− Langerhans Cells in Sentinel Lymph Nodes of Patients with Melanoma. International Journal of Molecular Sciences. 2022; 23(7):3441. https://doi.org/10.3390/ijms23073441
Chicago/Turabian StyleGerlini, Gianni, Paola Di Gennaro, Nicola Pimpinelli, Serena Sestini, and Lorenzo Borgognoni. 2022. "Tolerogenic IDO1+CD83− Langerhans Cells in Sentinel Lymph Nodes of Patients with Melanoma" International Journal of Molecular Sciences 23, no. 7: 3441. https://doi.org/10.3390/ijms23073441
APA StyleGerlini, G., Di Gennaro, P., Pimpinelli, N., Sestini, S., & Borgognoni, L. (2022). Tolerogenic IDO1+CD83− Langerhans Cells in Sentinel Lymph Nodes of Patients with Melanoma. International Journal of Molecular Sciences, 23(7), 3441. https://doi.org/10.3390/ijms23073441