Human Milk Oligosaccharides and Bacterial Profile Modulate Infant Body Composition during Exclusive Breastfeeding

 , , , , ,
, , , , , 
Abstract
:
1. Introduction
2. Results
2.1. Participants Characteristics, Anthropometrics and Body Composition
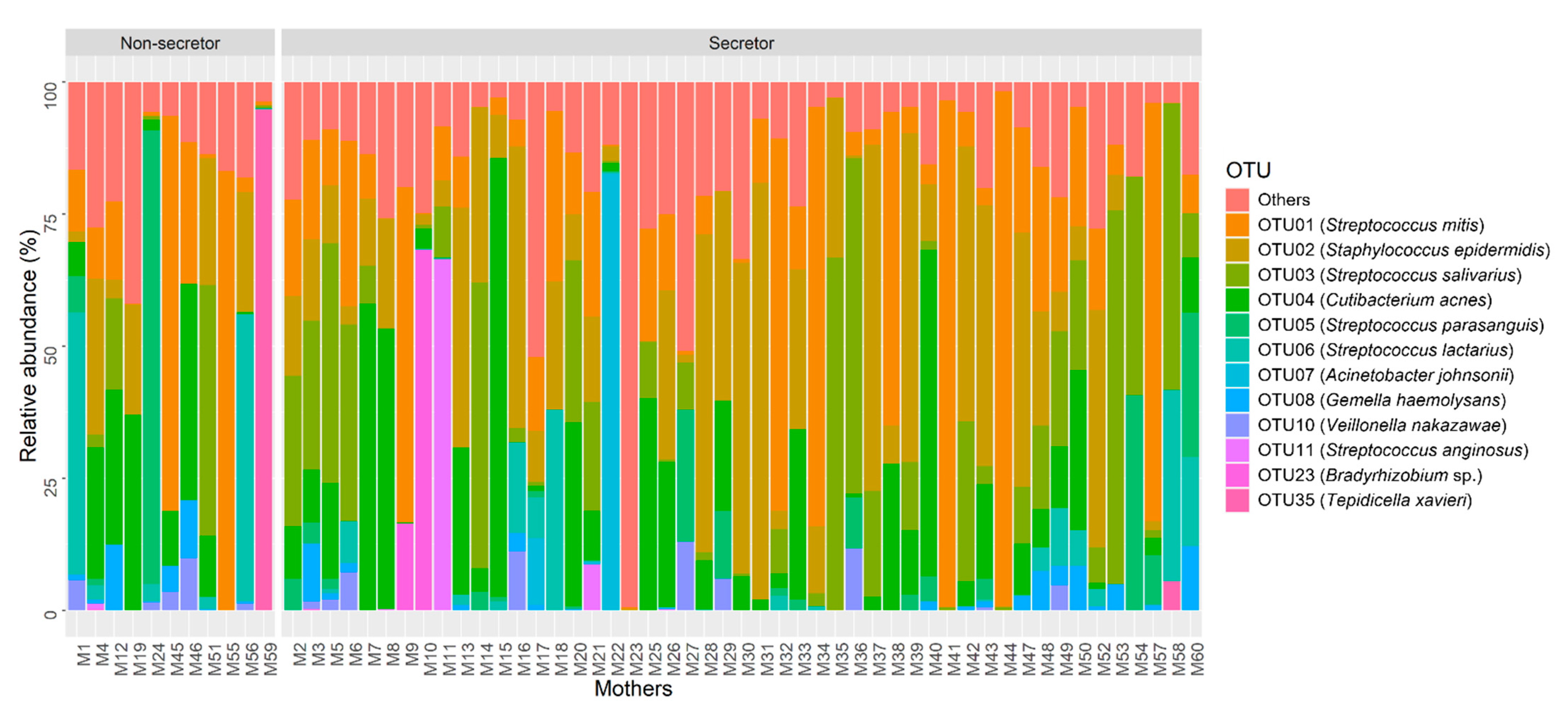
2.2. Human Milk Bacterial Composition
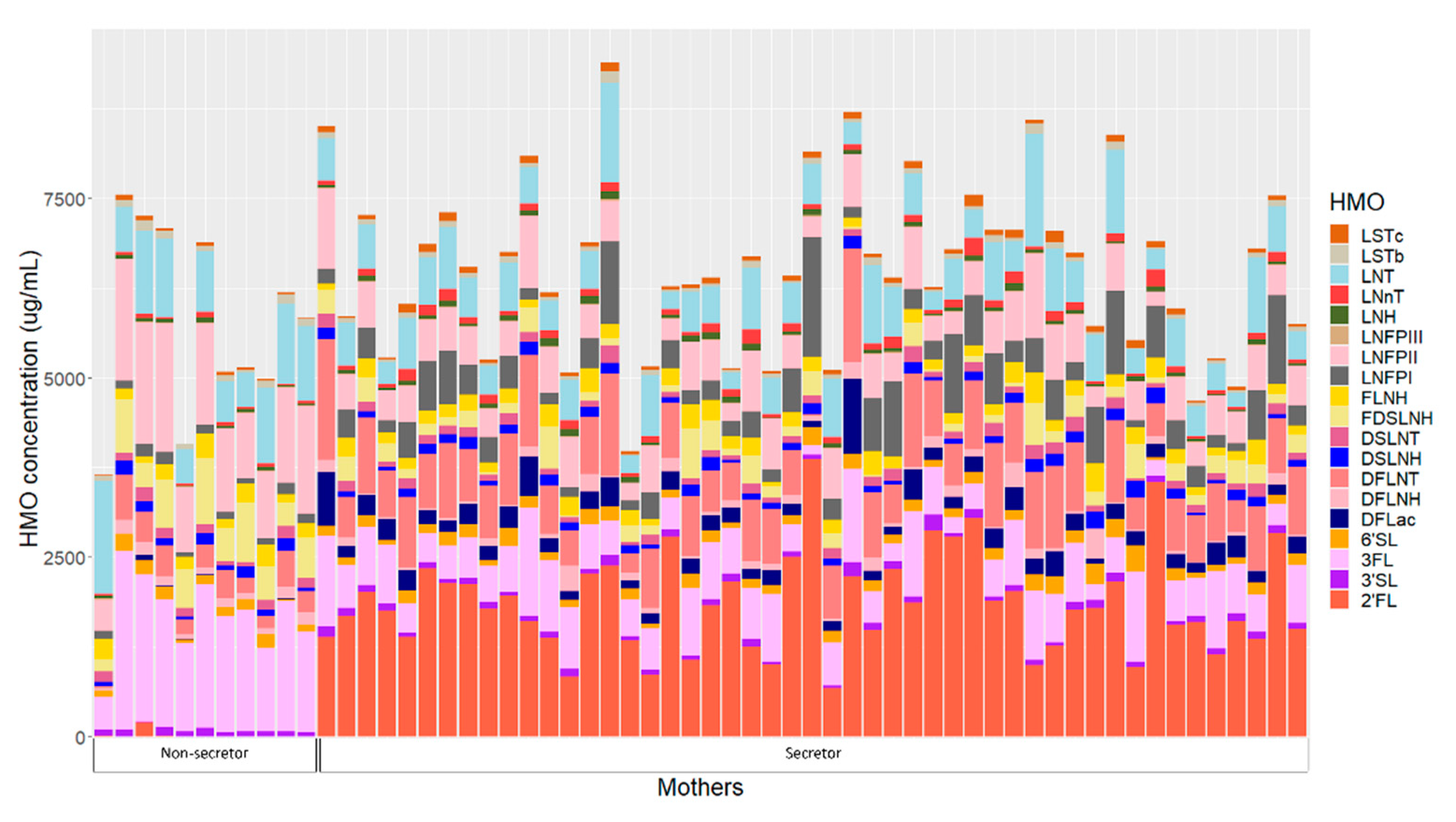
2.3. Human Milk Oligosaccharides
2.4. Maternal Characteristics and Human Milk Oligosaccharides
2.5. Maternal Characteristics and Human Milk Bacterial Profile
2.6. Maternal Characteristics and Infant Body Composition
2.7. Human Milk Oligosaccharide Concentration and Human Milk Bacterial Profile
2.8. Human Milk Bacterial Profile and Infant Body Composition
2.9. Human Milk Oligosaccharide Concentrations and Infant Body Composition
2.10. Human Milk Oligosaccharide Intake and Infant Body Composition
3. Discussion
4. Materials and Methods
4.1. Study Design
4.2. Sample Collection
4.3. Human Milk Oligosaccharides Analysis
4.4. 24-h Milk Intake
4.5. Calculated Daily Intakes of Human Milk Components
4.6. Anthropometry and Body Composition Measurements
4.7. DNA Extraction and Quantitation
4.8. 16S rRNA Gene Amplification and Barcoding
4.9. PacBio Sequencing
4.10. Sequencing Data Processing
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Geddes, D.T.; Gridneva, Z.; Perrella, S.L.; Mitoulas, L.R.; Kent, J.C.; Stinson, L.F.; Lai, C.T.; Sakalidis, V.; Twigger, A.J.; Hartmann, P.E. 25 Years of Research in Human Lactation: From Discovery to Translation. Nutrients 2021, 13, 3071. [Google Scholar] [CrossRef] [PubMed]
- Ballard, O.; Morrow, A.L. Human milk composition: Nutrients and bioactive factors. Orthop. Clin. N. Am. 2013, 60, 49–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savarino, G.; Corsello, A.; Corsello, G. Macronutrient balance and micronutrient amounts through growth and development. Ital. J. Pediatr. 2021, 47, 109. [Google Scholar] [CrossRef]
- Andreas, N.J.; Kampmann, B.; Mehring Le-Doare, K. Human breast milk: A review on its composition and bioactivity. Early Hum. Dev. 2015, 91, 629–635. [Google Scholar] [CrossRef] [PubMed]
- Bode, L. Human milk oligosaccharides: Every baby needs a sugar mama. Glycobiology 2012, 22, 1147–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alderete, T.L.; Autran, C.; Brekke, B.E.; Knight, R.; Bode, L.; Goran, M.I.; Fields, D.A. Associations between human milk oligosaccharides and infant body composition in the first 6 mo of life. Am. J. Clin. Nutr. 2015, 102, 1381–1388. [Google Scholar] [CrossRef]
- Walsh, C.; Lane, J.A.; van Sinderen, D.; Hickey, R.M. Human milk oligosaccharides: Shaping the infant gut microbiota and supporting health. J. Funct. Foods 2020, 72, 104074. [Google Scholar] [CrossRef]
- Sprenger, N.; Lee, L.Y.; De Castro, C.A.; Steenhout, P.; Thakkar, S.K. Longitudinal change of selected human milk oligosaccharides and association to infants’ growth, an observatory, single center, longitudinal cohort study. PLoS ONE 2017, 12, e0171814. [Google Scholar] [CrossRef] [Green Version]
- Gridneva, Z.; Rea, A.; Tie, W.J.; Lai, C.T.; Kugananthan, S.; Ward, L.C.; Murray, K.; Hartmann, P.E.; Geddes, D.T. Carbohydrates in Human Milk and Body Composition of Term Infants during the First 12 Months of Lactation. Nutrients 2019, 11, 1472. [Google Scholar] [CrossRef] [Green Version]
- Wells, J.C.; Chomtho, S.; Fewtrell, M.S. Programming of body composition by early growth and nutrition. Proc. Nutr. Soc. 2007, 66, 423–434. [Google Scholar] [CrossRef] [Green Version]
- Garwolinska, D.; Namiesnik, J.; Kot-Wasik, A.; Hewelt-Belka, W. Chemistry of human breast milk- A comprehensive review of the composition and role of milk Metabolites in child development. J. Agric. Food Chem. 2018, 66, 11881–11896. [Google Scholar] [CrossRef] [PubMed]
- Bode, L. Human milk oligosaccharides: Prebiotics and beyond. Nutr. Rev. 2009, 67 (Suppl. S2), S183–S191. [Google Scholar] [CrossRef] [PubMed]
- Azad, M.B.; Robertson, B.; Atakora, F.; Becker, A.B.; Subbarao, P.; Moraes, T.J.; Mandhane, P.J.; Turvey, S.E.; Lefebvre, D.L.; Sears, M.R.; et al. Human milk oligosaccharide concentrations are associated with multiple fixed and modifiable maternal characteristics, environmental factors, and feeding practices. J. Nutr. 2018, 148, 1733–1742. [Google Scholar] [CrossRef]
- Plows, J.F.; Berger, P.K.; Jones, R.B.; Alderete, T.L.; Yonemitsu, C.; Najera, J.A.; Khwajazada, S.; Bode, L.; Goran, M.I. Longitudinal changes in human milk oligosaccharides (HMOs) over the course of 24 months of lactation. J. Nutr. 2021, 151, 876–882. [Google Scholar] [CrossRef] [PubMed]
- McGuire, M.K.; Meehan, C.L.; McGuire, M.A.; Williams, J.E.; Foster, J.; Sellen, D.W.; Kamau-Mbuthia, E.W.; Kamundia, E.W.; Mbugua, S.; Moore, S.E.; et al. What’s normal? oligosaccharide concentrations and profiles in milk produced by healthy women vary geographically. Am. J. Clin. Nutr. 2017, 105, 1086–1100. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.L.; Alves, R.; Figueiredo, A.; Alves-Santos, N.; Freitas-Costa, N.; Batalha, M.; Yonemitsu, C.; Manivong, N.; Furst, A.; Bode, L.; et al. Human milk oligosaccharide profile variation throughout postpartum in healthy women in a Brazilian cohort. Nutrients 2020, 12, 790. [Google Scholar] [CrossRef] [Green Version]
- Isganaitis, E.; Venditti, S.; Matthews, T.J.; Lerin, C.; Demerath, E.W.; Fields, D.A. Maternal obesity and the human milk metabolome: Associations with infant body composition and postnatal weight gain. Am. J. Clin. Nutr. 2019, 110, 111–120. [Google Scholar] [CrossRef]
- Tonon, K.M.; Abrão, A.C.F.V.; Miranda, A.; Morais, T.B. Maternal and infant factors associated with human milk oligosaccharides concentrations according to secretor and lewis phenotypes. Nutrients 2019, 11, 1358. [Google Scholar] [CrossRef] [Green Version]
- Saben, J.L.; Sims, C.R.; Abraham, A.; Bode, L.; Andres, A. Human milk oligosaccharide concentrations and infant intakes are associated with maternal overweight and obesity and predict infant growth. Nutrients 2021, 13, 446. [Google Scholar] [CrossRef]
- Oliveros, E.; Martin, M.; Torres-Espinola, F.J.; Segura-Moreno, T.; Ramirez, M.; Santos-Fandila, A.; Buck, R.; Rueda, R.; Escudero, M.; Catena, A.; et al. Human milk levels of 2’-fucosyllactose and 6’-sialyllactose are positively associated with infant neurodevelopment and are not impacted by maternal BMI or diabetic status. Nutr. Food Sci. 2021, 4, 100024. [Google Scholar]
- Kok, P.; Seidell, J.C.; Meinders, A.E. The value and limitations of the body mass index (BMI) in the assessment of the health risks of overweight and obesity. Ned. Tijdschr. Voor Geneeskd. 2004, 148, 2379–2382. [Google Scholar]
- Kirmiz, N.; Robinson, R.C.; Shah, I.M.; Barile, D.; Mills, D.A. Milk glycans and their interaction with the infant-gut microbiota. Annu. Rev. Food Sci. Technol. 2018, 9, 429–450. [Google Scholar] [CrossRef] [PubMed]
- Korpela, K.; Salonen, A.; Hickman, B.; Kunz, C.; Sprenger, N.; Kukkonen, K.; Savilahti, E.; Kuitunen, M.; de Vos, W.M. Fucosylated oligosaccharides in mother’s milk alleviate the effects of caesarean birth on infant gut microbiota. Sci. Rep. 2018, 8, 13757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asakuma, S.; Hatakeyama, E.; Urashima, T.; Yoshida, E.; Katayama, T.; Yamamoto, K.; Kumagai, H.; Ashida, H.; Hirose, J.; Kitaoka, M. Physiology of consumption of human milk oligosaccharides by infant gut-associated bifidobacteria. J. Biol. Chem. 2011, 286, 34583–34592. [Google Scholar] [CrossRef] [Green Version]
- Fehr, K.; Moossavi, S.; Sbihi, H.; Boutin, R.C.T.; Bode, L.; Robertson, B.; Yonemitsu, C.; Field, C.J.; Becker, A.B.; Mandhane, P.J.; et al. Breastmilk feeding practices are associated with the co-occurrence of bacteria in mothers’ milk and the infant gut: The CHILD cohort study. Cell Host Microbe 2020, 28, 285–297.e4. [Google Scholar] [CrossRef]
- Corona-Cervantes, K.; Garcia-Gonzalez, I.; Villalobos-Flores, L.E.; Hernandez-Quiroz, F.; Pina-Escobedo, A.; Hoyo-Vadillo, C.; Rangel-Calvillo, M.N.; Garcia-Mena, J. Human milk microbiota associated with early colonization of the neonatal gut in Mexican newborns. PeerJ 2020, 8, e9205. [Google Scholar] [CrossRef]
- Ferretti, P.; Pasolli, E.; Tett, A.; Asnicar, F.; Gorfer, V.; Fedi, S.; Armanini, F.; Truong, D.T.; Manara, S.; Zolfo, M.; et al. Mother-to-infant microbial transmission from different body sites shapes the developing infant gut microbiome. Cell Host Microbe 2018, 24, 133–145.e135. [Google Scholar] [CrossRef]
- Williams, J.E.; Price, W.J.; Shafii, B.; Yahvah, K.M.; Bode, L.; McGuire, M.A.; McGuire, M.K. Relationships among microbial communities, maternal cells, oligosaccharides, and macronutrients in human milk. J. Hum. Lact. 2017, 33, 540–551. [Google Scholar] [CrossRef]
- Aakko, J.; Kumar, H.; Rautava, S.; Wise, A.; Autran, C.; Bode, L.; Isolauri, E.; Salminen, S. Human milk oligosaccharide categories define the microbiota composition in human colostrum. Benef. Microbes 2017, 8, 563–567. [Google Scholar] [CrossRef]
- Hunt, K.M.; Preuss, J.; Nissan, C.; Davlin, C.A.; Williams, J.E.; Shafii, B.; Richardson, A.D.; McGuire, M.K.; Bode, L.; McGuire, M.A. Human milk oligosaccharides promote the growth of staphylococci. Appl. Environ. Microbiol. 2012, 78, 4763–4770. [Google Scholar] [CrossRef] [Green Version]
- Pace, R.M.; Williams, J.E.; Robertson, B.; Lackey, K.A.; Meehan, C.L.; Price, W.J.; Foster, J.A.; Sellen, D.W.; Kamau-Mbuthia, E.W.; Kamundia, E.W.; et al. Variation in human milk compostion is related to differeces in milk and infant fecal microbial communities. Microorganisms 2021, 9, 1153. [Google Scholar] [CrossRef] [PubMed]
- Moossavi, S.; Atakora, F.; Miliku, K.; Sepehri, S.; Robertson, B.; Duan, Q.L.; Becker, A.B.; Mandhane, P.J.; Turvey, S.E.; Moraes, T.J.; et al. Integrated analysis of human milk microbiota with oligosaccharides and fatty acids in the CHILD cohort. Front. Nutr. 2019, 6, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabrera-Rubio, R.; Kunz, C.; Rudloff, S.; García-Mantrana, I.; Crehuá-Gaudiza, E.; Martínez-Costa, C.; Collado, M.C. Association of maternal secretor status and human milk oligosaccharides with milk microbiota: An observational pilot study. J. Pediatr. Gastroenterol. Nutr. 2019, 68, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Ayoub Moubareck, C.; Lootah, M.; Tahlak, M.; Venema, K. Profiles of human milk oligosaccharides and their relations to the milk microbiota of breastfeeding mothers in Dubai. Nutrients 2020, 12, 1727. [Google Scholar] [CrossRef] [PubMed]
- Lagstrom, H.; Rautava, S.; Ollila, H.; Kaljonen, A.; Turta, O.; Makela, J.; Yonemitsu, C.; Gupta, J.; Bode, L. Associations between human milk oligosaccharides and growth in infancy and early childhood. Am. J. Clin. Nutr. 2020, 111, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Larsson, M.W.; Lind, M.V.; Laursen, R.P.; Yonemitsu, C.; Larnkjaer, A.; Molgaard, C.; Michaelsen, K.F.; Bode, L. Human milk oligosaccharide composition is associated with excessive weight gain during exclusive breastfeeding-an explorative study. Front. Pediatr. 2019, 7, 297. [Google Scholar] [CrossRef] [Green Version]
- Binia, A.; Lavalle, L.; Chen, C.; Austin, S.; Agosti, M.; Al-Jashi, I.; Pereira, A.B.; Costeira, M.J.; Silva, M.G.; Marchini, G.; et al. Human milk oligosaccharides, infant growth, and adiposity over the first 4 months of lactation. Pediatr. Res. 2021, 90, 684–693. [Google Scholar] [CrossRef]
- Davis, J.C.; Lewis, Z.T.; Krishnan, S.; Bernstein, R.M.; Moore, S.E.; Prentice, A.M.; Mills, D.A.; Lebrilla, C.B.; Zivkovic, A.M. Growth and morbidity of Gambian infants are influenced by maternal milk oligosaccharides and infant gut microbiota. Sci. Rep. 2017, 7, 40466. [Google Scholar] [CrossRef]
- Butte, N.F.; Garza, C.; Smith, E.O.; Nichols, B.L. Human milk intake and growth in exclusively breast-fed infants. J. Pediatr. 1984, 104, 187–195. [Google Scholar] [CrossRef]
- Lingwood, B.E.; Storm van Leeuwen, A.-M.; Carberry, A.E.; Fitzgerald, E.C.; Callaway, L.K.; Colditz, P.B.; Ward, L.C. Prediction of fat-free mass and percentage of body fat in neonates using bioelectrical impedance analysis and anthropometric measures: Validation against the PEA POD. Br. J. Nutr. 2012, 107, 1545–1552. [Google Scholar] [CrossRef]
- Lyons-Reid, J.; Derraik, J.G.B.; Ward, L.C.; Tint, M.T.; Kenealy, T.; Cutfield, W.S. Bioelectrical impedance analysis for assessment of body composition in infants and young children-A systematic literature review. Clin. Obes. 2021, 11, e12441. [Google Scholar] [CrossRef] [PubMed]
- Lyons, K.E.; Ryan, C.A.; Dempsey, E.M.; Ross, R.P.; Stanton, C. Breast milk, a source of beneficial microbes and associated benefits for infant health. Nutrients 2020, 12, 1039. [Google Scholar] [CrossRef] [PubMed]
- Berger, P.K.; Plows, J.F.; Jones, R.B.; Alderete, T.L.; Yonemitsu, C.; Poulsen, M.; Ryoo, J.H.; Peterson, B.S.; Bode, L.; Goran, M.I. Human milk oligosaccharide 2’-fucosyllactose links feedings at 1 month to cognitive development at 24 months in infants of normal and overweight mothers. PLoS ONE 2020, 15, e0228323. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.E.; McGuire, M.K.; Meehan, C.L.; McGuire, M.A.; Brooker, S.L.; Kamau-Mbuthia, E.W.; Kamundia, E.W.; Mbugua, S.; Moore, S.E.; Prentice, A.M.; et al. Key genetic variants associated with variation of milk oligosaccharides from diverse human populations. Genomics 2021, 113, 1867–1875. [Google Scholar] [CrossRef] [PubMed]
- Han, S.M.; Derraik, J.G.B.; Binia, A.; Sprenger, N.; Vickers, M.H.; Cutfield, W.S. Maternal and infant factors influencing human milk oligosaccharide composition: Beyond maternal genetics. J. Nutr. 2021, 151, 1383–1393. [Google Scholar] [CrossRef] [PubMed]
- LeMay-Nedjelski, L.; Yonemitsu, C.; Asbury, M.R.; Butcher, J.; Ley, S.H.; Hanley, A.J.; Kiss, A.; Unger, S.; Copeland, J.K.; Wang, P.W.; et al. Oligosaccharides and microbiota in human milk are interrelated at 3 months postpartum in a cohort of women with a high prevalence of gestational impaired glucose tolerance. J. Nutr. 2021, 151, 3431–3441. [Google Scholar] [CrossRef] [PubMed]
- Saben, J.L.; Abraham, A.; Bode, L.; Sims, C.R.; Andres, A. Third-trimester glucose homeostasis in healthy women is differentially associated with human milk oligosaccharide composition at 2 months postpartum by secretor phenotype. Nutrients 2020, 12, 2209. [Google Scholar] [CrossRef]
- Cheema, A.S.; Stinson, L.F.; Rea, A.; Lai, C.T.; Payne, M.S.; Murray, K.; Geddes, D.T.; Gridneva, Z. Human milk lactose, insulin, and glucose relative to infant body composition during exclusive breastfeeding. Nutrients 2021, 13, 3724. [Google Scholar] [CrossRef]
- Cortes-Macias, E.; Selma-Royo, M.; Garcia-Mantrana, I.; Calatayud, M.; Gonzalez, S.; Martinez-Costa, C.; Collado, M.C. Maternal diet shapes the breast milk microbiota composition and diversity: Impact of mode of delivery and antibiotic exposure. J. Nutr. 2021, 151, 330–340. [Google Scholar] [CrossRef]
- Cabrera-Rubio, R.; Mira-Pascual, L.; Mira, A.; Collado, M.C. Impact of mode of delivery on the milk microbiota composition of healthy women. J. Dev. Orig. Health Dis. 2016, 7, 54–60. [Google Scholar] [CrossRef] [Green Version]
- Cabrera-Rubio, R.; Collado, M.C.; Laitinen, K.; Salminen, S.; Isolauri, E.; Mira, A. The human milk microbiome changes over lactation and is shaped by maternal weight and mode of delivery. Am. J. Clin. Nutr. 2012, 96, 544–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khodayar-Pardo, P.; Mira-Pascual, L.; Collado, M.C.; Martinez-Costa, C. Impact of lactation stage, gestational age and mode of delivery on breast milk microbiota. J. Perinatol. 2014, 34, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Hermansson, H.; Kumar, H.; Collado, M.C.; Salminen, S.; Isolauri, E.; Rautava, S. Breast milk microbiota Is shaped by mode of delivery and intrapartum antibiotic exposure. Front. Nutr. 2019, 6, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dizdar, E.A.; Sari, F.N.; Degirmencioglu, H.; Canpolat, F.E.; Oguz, S.S.; Uras, N.; Dilmen, U. Effect of mode of delivery on macronutrient content of breast milk. J. Matern.-Fetal Neonatal Med. 2014, 27, 1099–1102. [Google Scholar] [CrossRef] [PubMed]
- Samuel, T.M.; Binia, A.; de Castro, C.A.; Thakkar, S.K.; Billeaud, C.; Agosti, M.; Al-Jashi, I.; Costeira, M.J.; Marchini, G.; Martinez-Costa, C.; et al. Impact of maternal characteristics on human milk oligosaccharide composition over the first 4 months of lactation in a cohort of healthy European mothers. Sci. Rep. 2019, 9, 11767. [Google Scholar] [CrossRef] [PubMed]
- Kota, S.K.; Gayatri, K.; Jammula, S.; Kota, S.K.; Krishna, S.V.; Meher, L.K.; Modi, K.D. Endocrinology of parturition. Indian J. Endocrinol. Metab. 2013, 17, 50–59. [Google Scholar] [CrossRef]
- Kiriakopoulos, N.; Grigoriadis, S.; Maziotis, E.; Philippou, A.; Rapani, A.; Giannelou, P.; Tsioulou, P.; Sfakianoudis, K.; Kontogeorgi, A.; Bakas, P.; et al. Investigating stress response during vaginal delivery and elective cesarean section through assessment of levels of cortisol, interleukin 6 (IL-6), growth hormone (GH) and insulin-like growth factor 1 (IGF-1). J. Clin. Med. 2019, 8, 1112. [Google Scholar] [CrossRef] [Green Version]
- Iversen, D.S.; Kesmodel, U.S.; Ovesen, P.G. Associations between parity and maternal BMI in a population-based cohort study. Acta Obstet. Gynecol. Scand. 2018, 97, 694–700. [Google Scholar] [CrossRef] [Green Version]
- Reis-Santos, B.; Barros, F.C.; Horta, B.L. Is there a causal effect of parity on body composition: A birth cohort study. BMC Public Health 2018, 18, 215. [Google Scholar] [CrossRef]
- Prentice, A.; Jarjou, L.M.; Drury, P.J.; Dewit, O.; Crawford, M.A. Breast-milk fatty acids of rural Gambian mothers: Effects of diet and maternal parity. J. Pediatr. Gastroenterol. Nutr. 1989, 8, 486–490. [Google Scholar] [CrossRef]
- Léké, A.; Grognet, S.; Deforceville, M.; Goudjil, S.; Chazal, C.; Kongolo, G.; Dzon, B.E.; Biendo, M. Macronutrient composition in human milk from mothers of preterm and term neonates is highly variable during the lactation period. Clin. Nutr. Exp. 2019, 26, 59–72. [Google Scholar] [CrossRef] [Green Version]
- Conde-Agudelo, A.; Rosas-Bermudez, A.; Castano, F.; Norton, M.H. Effects of birth spacing on maternal, perinatal, infant, and child health: A systematic review of causal mechanisms. Stud. Fam. Plan. 2012, 43, 93–114. [Google Scholar] [CrossRef] [PubMed]
- Collado, M.C.; Laitinen, K.; Salminen, S.; Isolauri, E. Maternal weight and excessive weight gain during pregnancy modify the immunomodulatory potential of breast milk. Pediatr. Res. 2012, 72, 77–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soeorg, H.; Metsvaht, T.; Eelmae, I.; Merila, M.; Treumuth, S.; Huik, K.; Jurna-Ellam, M.; Ilmoja, M.L.; Lutsar, I. The role of breast milk in the colonization of neonatal gut and skin with coagulase-negative staphylococci. Pediatr. Res. 2017, 82, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Efuntoye, M.O.; Adetosoye, A.I. Enterotoxigenicity and drug sensitivity of staphylococci from children aged five years and below with sporadic diarrhoea. East Afr. Med. J. 2003, 80, 656–659. [Google Scholar] [CrossRef] [PubMed]
- Lundell, A.C.; Adlerberth, I.; Lindberg, E.; Karlsson, H.; Ekberg, S.; Aberg, N.; Saalman, R.; Hock, B.; Steinkasserer, A.; Hesselmar, B.; et al. Increased levels of circulating soluble CD14 but not CD83 in infants are associated with early intestinal colonization with Staphylococcus aureus. Clin. Exp. Allergy 2007, 37, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Adlerberth, I.; Strachan, D.P.; Matricardi, P.M.; Ahrne, S.; Orfei, L.; Aberg, N.; Perkin, M.R.; Tripodi, S.; Hesselmar, B.; Saalman, R.; et al. Gut microbiota and development of atopic eczema in 3 European birth cohorts. J. Allergy Clin. Immunol. 2007, 120, 343–350. [Google Scholar] [CrossRef]
- Kalliomaki, M.; Collado, M.C.; Salminen, S.; Isolauri, E. Early differences in fecal microbiota composition in children may predict overweight. Am. J. Clin. Nutr. 2008, 87, 534–538. [Google Scholar] [CrossRef]
- van den Bogert, B.; Erkus, O.; Boekhorst, J.; de Goffau, M.; Smid, E.J.; Zoetendal, E.G.; Kleerebezem, M. Diversity of human small intestinal streptococcus and veillonella populations. FEMS Microbiol. Ecol. 2013, 85, 376–388. [Google Scholar] [CrossRef] [Green Version]
- Zoetendal, E.G.; Raes, J.; van den Bogert, B.; Arumugam, M.; Booijink, C.C.; Troost, F.J.; Bork, P.; Wels, M.; de Vos, W.M.; Kleerebezem, M. The human small intestinal microbiota is driven by rapid uptake and conversion of simple carbohydrates. ISME J. 2012, 6, 1415–1426. [Google Scholar] [CrossRef]
- Ng, S.K.; Hamilton, I.R. Lactate metabolism by Veillonella parvula. J. Bacteriol. 1971, 105, 999–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monnard, C.R.; Dulloo, A.G. Polyunsaturated fatty acids as modulators of fat mass and lean mass in human body composition regulation and cardiometabolic health. Obes. Rev. 2021, 22 (Suppl. S2), e13197. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, H.; Zhang, B.; Popkin, B.M.; Du, S. Elevated Fat Intake Increases Body Weight and the Risk of Overweight and Obesity among Chinese Adults: 1991-2015 Trends. Nutrients 2020, 12, 3272. [Google Scholar] [CrossRef]
- Marcobal, A.; Barboza, M.; Froehlich, J.W.; Block, D.E.; German, J.B.; Lebrilla, C.B.; Mills, D.A. Consumption of human milk oligosaccharides by gut-related microbes. J. Agric. Food Chem. 2010, 58, 5334–5340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, L.; Kiewiet, M.B.G.; Logtenberg, M.J.; Groeneveld, A.; Nauta, A.; Schols, H.A.; Walvoort, M.T.C.; Harmsen, H.J.M.; de Vos, P. Effects of different human milk oligosaccharides on growth of bifidobacteria in monoculture and co-culture with faecalibacterium prausnitzii. Front. Microbiol. 2020, 11, 569700. [Google Scholar] [CrossRef]
- Lin, A.E.; Autran, C.A.; Szyszka, A.; Escajadillo, T.; Huang, M.; Godula, K.; Prudden, A.R.; Boons, G.J.; Lewis, A.L.; Doran, K.S.; et al. Human milk oligosaccharides inhibit growth of group B streptococcus. J. Biol. Chem. 2017, 292, 11243–11249. [Google Scholar] [CrossRef] [Green Version]
- Kumar, H.; du Toit, E.; Kulkarni, A.; Aakko, J.; Linderborg, K.M.; Zhang, Y.; Nicol, M.P.; Isolauri, E.; Yang, B.; Collado, M.C.; et al. Distinct patterns in human milk microbiota and fatty acid profiles across specific geographic locations. Front. Microbiol. 2016, 7, 1619. [Google Scholar] [CrossRef] [Green Version]
- Lackey, K.A.; Williams, J.E.; Meehan, C.L.; Zachek, J.A.; Benda, E.D.; Price, W.J.; Foster, J.A.; Sellen, D.W.; Kamau-Mbuthia, E.W.; Kamundia, E.W.; et al. What’s normal? Microbiomes in human milk and infant feces are related to each other but vary geographically: The INSPIRE study. Front. Nutr. 2019, 6, 45. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Gallego, C.; Morales, J.M.; Monleon, D.; du Toit, E.; Kumar, H.; Linderborg, K.M.; Zhang, Y.; Yang, B.; Isolauri, E.; Salminen, S.; et al. Human Breast Milk NMR Metabolomic Profile across Specific Geographical Locations and Its Association with the Milk Microbiota. Nutrients 2018, 10, 1355. [Google Scholar] [CrossRef] [Green Version]
- Seferovic, M.D.; Mohammad, M.; Pace, R.M.; Engevik, M.; Versalovic, J.; Bode, L.; Haymond, M.; Aagaard, K.M. Maternal diet alters human milk oligosaccharide composition with implications for the milk metagenome. Sci. Rep. 2020, 10, 22092. [Google Scholar] [CrossRef]
- Torres Roldan, V.D.; Urtecho, S.M.; Gupta, J.; Yonemitsu, C.; Carcamo, C.P.; Bode, L.; Ochoa, T.J. Human milk oligosaccharides and their association with late-onset neonatal sepsis in Peruvian very-low-birth-weight infants. Am. J. Clin. Nutr. 2020, 112, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Rousseaux, A.; Brosseau, C.; Le Gall, S.; Piloquet, H.; Barbarot, S.; Bodinier, M. Human milk oligosaccharides: Their effects on the host and their potential as therapeutic agents. Front. Immunol. 2021, 12, 680911. [Google Scholar] [CrossRef] [PubMed]
- Borewicz, K.; Gu, F.; Saccenti, E.; Arts, I.C.W.; Penders, J.; Thijs, C.; van Leeuwen, S.S.; Lindner, C.; Nauta, A.; van Leusen, E.; et al. Correlating infant faecal microbiota composition and human milk oligosaccharide consumption by microbiota of one-month old breastfed infants. Mol. Nutr. Food. Res. 2019, 63, e1801214. [Google Scholar] [CrossRef] [PubMed]
- Perrella, S.; Gridneva, Z.; Lai, C.T.; Stinson, L.; George, A.; Bilston-John, S.; Geddes, D. Human milk composition promotes optimal infant growth, development and health. Semin. Perinatol. 2021, 45, 151380. [Google Scholar] [CrossRef] [PubMed]
- Newburg, D.S.; Morelli, L. Human milk and infant intestinal mucosal glycans guide succession of the neonatal intestinal microbiota. Pediatr. Res. 2015, 77, 115–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padilha, M.; Danneskiold-Samsoe, N.B.; Brejnrod, A.; Hoffmann, C.; Cabral, V.P.; Iaucci, J.M.; Sales, C.H.; Fisberg, R.M.; Cortez, R.V.; Brix, S.; et al. The human milk microbiota is modulated by maternal diet. Microorganisms 2019, 7, 502. [Google Scholar] [CrossRef] [Green Version]
- Jorgensen, J.M.; Young, R.; Ashorn, P.; Ashorn, U.; Chaima, D.; Davis, J.C.C.; Goonatilleke, E.; Kumwenda, C.; Lebrilla, C.B.; Maleta, K.; et al. Associations of human milk oligosaccharides and bioactive proteins with infant growth and development among Malawian mother-infant dyads. Am. J. Clin. Nutr. 2020, 113, 209–220. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, Z.; Wu, Y.; Wang, Y.; Wang, J.; Zhou, L.; Ni, Z.; Hao, L.; Yang, N.; Yang, X. Early feeding of larger volumes of formula milk is associated with greater body weight or overweight in later infancy. Nutr. J. 2018, 17, 12. [Google Scholar] [CrossRef] [Green Version]
- Giugliani, E.R.J. Growth in exclusively breastfed infants. J. Pediatr. 2019, 95 (Suppl. S1), 79–84. [Google Scholar] [CrossRef]
- Puccio, G.; Alliet, P.; Cajozzo, C.; Janssens, E.; Corsello, G.; Sprenger, N.; Wernimont, S.; Egli, D.; Gosoniu, L.; Steenhout, P. Effects of infant formula with human milk oligosaccharides on growth and morbidity: A randomized multicenter trial. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 624–631. [Google Scholar] [CrossRef] [Green Version]
- Marriage, B.J.; Buck, R.H.; Goehring, K.C.; Oliver, J.S.; Williams, J.A. Infants fed a lower calorie formula with 2’FL show growth and 2’FL uptake like breast-fed infants. J. Pediatr. Gastroenterol. Nutr. 2015, 61, 649–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eriksen, K.G.; Christensen, S.H.; Lind, M.V.; Michaelsen, K.F. Human milk composition and infant growth. Curr. Opin. Clin. Nutr. Metab. Care 2018, 21, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Butte, N.F.; Wong, W.W.; Hopkinson, J.M.; Smith, E.O.; Ellis, K.J. Infant feeding mode affects early growth and body composition. Pediatrics 2000, 106, 1355–1366. [Google Scholar] [CrossRef] [PubMed]
- Gridneva, Z.; Rea, A.; Hepworth, A.R.; Ward, L.C.; Lai, C.T.; Hartmann, P.E.; Geddes, D.T. Relationships between breastfeeding patterns and maternal and infant body composition over the first 12 months of lactation. Nutrients 2018, 10, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hull, H.R.; Dinger, M.K.; Knehans, A.W.; Thompson, D.M.; Fields, D.A. Impact of maternal body mass index on neonate birthweight and body composition. Am. J. Obstet. Gynecol. 2008, 198, 416.e1–416.e6. [Google Scholar] [CrossRef] [PubMed]
- Parschat, K.; Melsaether, C.; Japelt, K.R.; Jennewein, S. Clinical evaluation of 16-week supplementation with 5HMO-mix in healthy-term human infants to determine tolerability, safety, and effect on growth. Nutrients 2021, 13, 2871. [Google Scholar] [CrossRef]
- Frese, S.A.; Hutton, A.A.; Contreras, L.N.; Shaw, C.A.; Palumbo, M.C.; Casaburi, G.; Xu, G.; Davis, J.C.C.; Lebrilla, C.B.; Henrick, B.M.; et al. Persistence of Supplemented Bifidobacterium longum subsp. infantis EVC001 in Breastfed Infants. MSphere 2017, 2, e00501-17. [Google Scholar] [CrossRef] [Green Version]
- Smilowitz, J.T.; Moya, J.; Breck, M.A.; Cook, C.; Fineberg, A.; Angkustsiri, K.; Underwood, M.A. Safety and tolerability of Bifidobacterium longum subspecies infantis EVC001 supplementation in healthy term breastfed infants: A phase I clinical trial. BMC Pediatr. 2017, 17, 133. [Google Scholar] [CrossRef]
- Tahir, M.J.; Ejima, K.; Li, P.; Demerath, E.W.; Allison, D.B.; Fields, D.A. Associations of breastfeeding or formula feeding with infant anthropometry and body composition at 6 months. Matern. Child Nutr. 2021, 17, e13105. [Google Scholar] [CrossRef]
- Bell, K.A.; Wagner, C.L.; Feldman, H.A.; Shypailo, R.J.; Belfort, M.B. Associations of infant feeding with trajectories of body composition and growth. Am. J. Clin. Nutr. 2017, 106, 491–498. [Google Scholar] [CrossRef]
- Ramsay, D.T.; Mitoulas, L.R.; Kent, J.C.; Cregan, M.D.; Doherty, D.A.; Larsson, M.; Hartmann, P.E. Milk flow rates can be used to identify and investigate milk ejection in women expressing breast milk using an electric breast pump. Breastfeed. Med. 2006, 1, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Kent, J.C.; Hepworth, A.R.; Sherriff, J.L.; Cox, D.B.; Mitoulas, L.R.; Hartmann, P.E. Longitudinal changes in breastfeeding patterns from 1 to 6 months of lactation. Breastfeed. Med. 2013, 8, 401–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neville, M.C.; Keller, R.; Seacat, J.; Lutes, V.; Neifert, M.; Casey, C.; Allen, J.; Archer, P. Studies in human lactation: Milk volumes in lactating women during the onset of lactation and full lactation. Am. J. Clin. Nutr. 1988, 48, 1375–1386. [Google Scholar] [CrossRef] [PubMed]
- Gridneva, Z.; Hepworth, A.R.; Ward, L.C.; Lai, C.T.; Hartmann, P.E.; Geddes, D.T. Determinants of body composition in breastfed infants using bioimpedance spectroscopy and ultrasound skinfolds-methods comparison. Pediatr. Res. 2017, 81, 423–433. [Google Scholar] [CrossRef]
- VanItallie, T.B.; Yang, M.U.; Heymsfield, S.B.; Funk, R.C.; Boileau, R.A. Height-normalized indices of the body’s fat-free mass and fat mass: Potentially useful indicators of nutritional status. Am. J. Clin. Nutr. 1990, 52, 953–959. [Google Scholar] [CrossRef] [Green Version]
- WHO. Child Growth Standards Software. WHO Anthro Survey Analyser and Other Tools (Version 3.2.2). Available online: https://www.who.int/childgrowth/software/en/ (accessed on 16 June 2021).
- Cheema, A.S.; Lai, C.T.; Dymock, M.; Rae, A.; Geddes, D.T.; Payne, M.S.; Stinson, L.F. Impact of expression mode and timing of sample collection, relative to milk ejection, on human milk bacterial DNA profiles. J. Appl. Microbiol. 2021, 131, 988–995. [Google Scholar] [CrossRef]
- Stinson, L.; Hallingstrom, M.; Barman, M.; Viklund, F.; Keelan, J.; Kacerovsky, M.; Payne, M.; Jacobsson, B. Comparison of Bacterial DNA Profiles in Mid-Trimester Amniotic Fluid Samples From Preterm and Term Deliveries. Front. Microbiol. 2020, 11, 415. [Google Scholar] [CrossRef] [Green Version]
- Cheema, A.S.; Stinson, L.F.; Lai, C.T.; Geddes, D.T.; Payne, M.S. DNA extraction method influences human milk bacterial profiles. J. Appl. Microbiol. 2020, 130, 142–156. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienne, Austria, 2021. [Google Scholar]
- Ho, N.T.; Li, F.; Wang, S.; Kuhn, L. metamicrobiomeR: An R package for analysis of microbiome relative abundance data using zero-inflated beta GAMLSS and meta-analysis across studies using random effects models. BMC Bioinform. 2019, 20, 188. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Maternal Predictor | Secretor | Non-Secretor | Interaction | ||||
|---|---|---|---|---|---|---|---|
| Parameter Estimate | SE | p-Value | Parameter Estimate | SE | p-Value | p-Value | |
| 2′FL | |||||||
| FM (%) | 0.43 | 0.16 | 0.01 | 0.07 | 0.03 | 0.01 | 0.03 |
| Log (FMI (kg/m2)) | 6.05 | 3.00 | 0.05 | 1.25 | 0.53 | 0.02 | 0.23 |
| Log (FM/FFM) | 7.85 | 4.11 | 0.06 | 1.59 | 0.61 | 0.01 | 0.14 |
| 3′SL | |||||||
| Log (FM (kg)) | −0.34 | 1.48 | 0.82 | −7.13 | 3.03 | 0.02 | 0.05 |
| FM (%) | −0.04 | 0.08 | 0.66 | −0.37 | 0.12 | <0.001 | 0.02 |
| Log (FMI (kg/m2)) | −0.63 | 1.53 | 0.68 | −5.88 | 2.56 | 0.03 | 0.08 |
| Log (FM/FFM) | −2.81 | 2.04 | 0.17 | −8.36 | 2.68 | <0.001 | 0.11 |
| FLNH | |||||||
| Age | −0.12 | 0.24 | 0.63 | 1.80 | 0.69 | 0.01 | 0.01 |
| Log (Weight (kg)) | 14.32 | 5.43 | 0.01 | −36.11 | 22.83 | 0.12 | 0.04 |
| Log (BMI (kg/m2)) | 13.81 | 5.99 | 0.03 | −15.11 | 15.95 | 0.35 | 0.10 |
| Log (FM (kg)) | 8.37 | 3.35 | 0.02 | −16.06 | 8.88 | 0.08 | 0.01 |
| Log (FMI (kg/m2)) | 7.86 | 3.50 | 0.03 | −11.22 | 7.82 | 0.16 | 0.03 |
| Entire cohort | |||||||
| 2′FL | |||||||
| Log (Weight (kg)) | 3.63 | 1.52 | 0.02 | - | - | - | - |
| Log (BMI (kg/m2)) | 2.37 | 1.04 | 0.03 | - | - | - | - |
| Log (FM (kg)) | 1.68 | 0.61 | 0.01 | - | - | - | - |
| Log (LNH) | |||||||
| Log (BMI (kg/m2)) | 0.73 | 0.27 | 0.01 | - | - | - | - |
| FFMI (kg/m2) | 0.05 | 0.02 | 0.02 | - | - | - | - |
| Maternal Predictor | Secretor | Non-Secretor | Interaction | ||||
|---|---|---|---|---|---|---|---|
| Parameter Estimate | SE | p-Value | Parameter Estimate | SE | p-Value | p-Value | |
| OTU03 (Streptococcus salivarius) | |||||||
| Log (Weight (kg)) | −2.24 | 0.82 | 0.01 | 6.01 | 5.07 | 0.24 | 0.12 |
| Log (BMI (kg/m2)) | −2.52 | 0.89 | 0.01 | 4.44 | 2.8 | 0.12 | 0.02 |
| Log (FM (kg)) | −1.37 | 0.5 | 0.01 | 2.63 | 2 | 0.19 | 0.06 |
| FM (%) | −0.06 | 0.03 | 0.04 | 0.12 | 0.08 | 0.17 | 0.05 |
| Log (FMI (kg/m2)) | −1.42 | 0.52 | 0.01 | 2.25 | 1.51 | 0.14 | 0.02 |
| Log (FM/FFM) | −1.33 | 0.64 | 0.04 | 2.66 | 1.96 | 0.18 | 0.06 |
| OTU06 (Streptococcus lactarius) | |||||||
| Age | 0.03 | 0.04 | 0.41 | 0.82 | 0.26 | <0.001 | <0.001 |
| OTU08 (Gemella haemolysans) | |||||||
| Log (BMI (kg/m2)) | −0.69 | 1.28 | 0.59 | 5.43 | 2.39 | 0.03 | 0.03 |
| FFMI (kg/m2) | −0.05 | 0.11 | 0.64 | 0.55 | 0.24 | 0.03 | 0.03 |
| Log (FMI (kg/m2)) | −0.37 | 0.74 | 0.62 | 2.62 | 1.22 | 0.04 | 0.04 |
| OTU10 (Veillonella nakazawae) | |||||||
| FFMI (kg/m2) | 0.12 | 0.06 | 0.04 | 0.42 | 0.19 | 0.03 | 0.14 |
| Entire cohort | |||||||
| OUT01 (Streptococcus mitis) | |||||||
| Age | 0.07 | 0.03 | 0.04 | - | - | - | - |
| OTU02 (Staphylococcus epidermidis) | |||||||
| Log (Weight (kg)) | −1.54 | 0.69 | 0.03 | - | - | - | - |
| FFM (kg) | −0.04 | 0.02 | 0.02 | - | - | - | - |
| Log (FM (kg)) | −1.04 | 0.43 | 0.02 | - | - | - | - |
| Log (FMI (kg/m2)) | −0.9 | 0.43 | 0.04 | - | - | - | - |
| OTU10 (Veillonella nakazawae) | |||||||
| FFM (kg) | 0.04 | 0.02 | 0.03 | - | - | - | - |
| Maternal Predictors | Secretor | Non-Secretor | Interaction | ||||
|---|---|---|---|---|---|---|---|
| Parameter Value | SE | p-Value | Parameter Value | SE | p-Value | p-Value | |
| Infant weight (kg) | |||||||
| FFM (kg) | 0.01 | 0.01 | 0.51 | −0.26 | 0.12 | 0.04 | 0.03 |
| FFMI (kg/m2) | −0.00 | 0.04 | 0.98 | −0.47 | 0.14 | <0.001 | <0.001 |
| Infant length (cm) | |||||||
| FFM (kg) | 0.10 | 0.04 | 0.01 | −0.66 | 0.36 | 0.08 | 0.04 |
| FFMI (kg/m2) | 0.19 | 0.13 | 0.16 | −1.04 | 0.47 | 0.03 | 0.02 |
| Infant BMI (kg/m2) | |||||||
| FFMI (kg/m2) | −0.11 | 0.07 | 0.11 | −0.63 | 0.2 | <0.001 | 0.01 |
| Infant log (FFM (kg)) | |||||||
| FFM (kg) | 0.00 | 0.00 | 0.33 | −0.04 | 0.02 | 0.03 | 0.02 |
| FFMI (kg/m2) | 0.00 | 0.01 | 0.83 | −0.06 | 0.02 | <0.001 | <0.001 |
| Infant log (FFMI (kg/m2)) | |||||||
| Log (BMI (kg/m2)) | −0.08 | 0.07 | 0.23 | −0.34 | 0.15 | 0.03 | 0.13 |
| FFM (kg) | 0.00 | 0.00 | 0.97 | −0.02 | 0.01 | 0.03 | 0.03 |
| FFMI (kg/m2) | −0.00 | 0.01 | 0.70 | −0.05 | 0.01 | <0.001 | <0.001 |
| Infant FM (kg) | |||||||
| FFMI (kg/m2) | 0.01 | 0.02 | 0.44 | −0.16 | 0.06 | 0.01 | 0.01 |
| Infant FMI (kg/m2) | |||||||
| FFMI (kg/m2) | 0.03 | 0.05 | 0.60 | −0.42 | 0.16 | 0.01 | 0.01 |
| Entire cohort | |||||||
| Infant length (cm) | |||||||
| Log (Weight (kg)) | 3.48 | 1.69 | 0.04 | - | - | - | - |
| Infant BMI (kg/m2) | |||||||
| Log (BMI (kg/m2)) | −2.16 | 0.9 | 0.02 | - | - | - | - |
| Log (FM (kg)) | −1.24 | 0.51 | 0.02 | - | - | - | - |
| FM (%) | −0.06 | 0.03 | 0.03 | - | - | - | - |
| Log (FMI (kg/m2)) | −1.33 | 0.51 | 0.01 | - | - | - | - |
| Infant FM (%) | |||||||
| Age | −0.17 | 0.07 | 0.02 | - | - | - | - |
| Infant FM/FFM | |||||||
| Age | −0.00 | 0.00 | 0.02 | - | - | - | - |
| HMO (Concentrations) | Secretor | Non-Secretor | Interaction | ||||
|---|---|---|---|---|---|---|---|
| Parameter Estimate | SE | p-Value | Parameter Estimate | SE | p-Value | p-Value | |
| OTU02 (Staphylococcus epidermidis) | |||||||
| 3′SL | −0.12 | 0.05 | 0.01 | 0.09 | 0.11 | 0.45 | 0.10 |
| OTU04 (Cutibacterium acnes) | |||||||
| 6′SL | −0.01 | 0.02 | 0.71 | 0.01 | 0.05 | 0.03 | 0.04 |
| OTU05 (Streptococcus parasanguis) | |||||||
| Log (DFLac) | 0.03 | 0.37 | 0.93 | 3.09 | 1.17 | 0.01 | 0.02 |
| 3′SL | 0.05 | 0.05 | 0.34 | −2.80 | 0.78 | <0.001 | <0.001 |
| 6′SL | −0.02 | 0.05 | 0.60 | −0.20 | 0.09 | 0.04 | 0.10 |
| Log (LNFP I) | −0.11 | 0.31 | 0.72 | −9.49 | 2.40 | <0.001 | <0.001 |
| Log (LSTb) | 0.05 | 0.41 | 0.90 | −17.23 | 7.23 | 0.02 | 0.02 |
| Log (LSTc) | 0.11 | 0.55 | 0.84 | −1.71 | 0.81 | 0.04 | 0.07 |
| Log (LNH) | −0.04 | 0.36 | 0.91 | −4.32 | 1.37 | <0.001 | <0.001 |
| DSLNH | 0.00 | 0.01 | 0.73 | −0.02 | 0.01 | 0.04 | 0.05 |
| OTU06 (Streptococcus lactarius) | |||||||
| 2′FL | 0.02 | 0.04 | 0.65 | 26.81 | 11.68 | 0.03 | 0.03 |
| Log (LNnT) | 0.28 | 0.55 | 0.61 | −1.89 | 0.58 | <0.001 | 0.01 |
| Log (FDSLNH) | −0.03 | 0.18 | 0.85 | −2.15 | 0.69 | <0.001 | <0.001 |
| OTU08 (Gemella haemolysans) | |||||||
| 2′FL | −0.01 | 0.04 | 0.83 | 0.98 | 0.33 | <0.001 | <0.001 |
| 3′SL | 0.06 | 0.09 | 0.51 | −0.25 | 0.11 | 0.03 | 0.04 |
| Log (LNnT) | −0.19 | 0.39 | 0.63 | 1.22 | 0.54 | 0.03 | 0.04 |
| OTU10 (Veillonella nakazawae) | |||||||
| Log (DFLac) | −1.81 | 0.85 | 0.04 | −0.33 | 0.35 | 0.35 | 0.11 |
| Entire cohort | |||||||
| OTU02 (Staphylococcus epidermidis) | |||||||
| FLNH | −0.04 | 0.02 | 0.03 | - | - | - | - |
| OTU03 (Streptococcus salivarius) | |||||||
| FLNH | −0.04 | 0.02 | 0.02 | - | - | - | - |
| OTU04 (Cutibacterium acnes) | |||||||
| Log (LNnT) | 0.71 | 0.28 | 0.01 | - | - | - | - |
| OTU08 (Gemella haemolysans) | |||||||
| Log (DFLac) | 0.47 | 0.19 | 0.02 | - | - | - | - |
| OTU10 (Veillonella nakazawae) | |||||||
| Log (LNFP III) | −0.83 | 0.36 | 0.03 | - | - | - | - |
| DSLNT | −0.01 | 0.00 | <0.001 | - | - | - | - |
| Log (FDSLNH) | −0.43 | 0.17 | 0.02 | - | - | - | - |
| Human Milk OTUs | Secretor | Non-Secretor | Interaction | Log (Infant Milk Intake) | ||||
|---|---|---|---|---|---|---|---|---|
| Parameter Value | SE | p-Value | Parameter Value | SE | p-Value | p-Value | p-Value | |
| Head circumference (cm) | ||||||||
| OTU01 (Streptococcus mitis) | 0.01 | 0.01 | 0.56 | −0.03 | 0.01 | 0.01 | 0.03 | 0.03 |
| Length-for-age z-score | ||||||||
| OTU01 (Streptococcus mitis) | −0.00 | 0.01 | 0.61 | −0.02 | 0.01 | 0.03 | 0.15 | 0.03 |
| BMI-for-age z-score | ||||||||
| OTU05 (Streptococcus parasanguis) | −0.02 | 0.02 | 0.42 | 0.01 | 0.01 | 0.02 | 0.16 | 0.03 |
| Head circumference-for-age z-score | ||||||||
| OTU01 (Streptococcus mitis) | 0.00 | 0.01 | 0.63 | −0.03 | 0.01 | <0.001 | 0.01 | 0.10 |
| Entire cohort | ||||||||
| Weight (kg) | ||||||||
| OTU05 (Streptococcus parasanguis) | 0.02 | 0.01 | <0.001 | - | - | - | - | <0.001 |
| Length (cm) | ||||||||
| OTU02 (Staphylococcus epidermidis) | −0.04 | 0.02 | 0.04 | - | - | - | - | <0.001 |
| Weight-for-length z-score | ||||||||
| OTU02 (Staphylococcus epidermidis) | 0.02 | 0.01 | 0.02 | - | - | - | - | NA |
| OTU06 (Streptococcus lactarius) | 0.02 | 0.01 | 0.04 | - | - | - | - | NA |
| Weight-for-age z-score | ||||||||
| OTU05 (Streptococcus parasanguis) | 0.01 | 0.01 | 0.03 | - | - | - | - | <0.001 |
| BMI-for-age z-score | ||||||||
| OTU06 (Streptococcus lactarius) | 0.02 | 0.01 | 0.01 | - | - | - | - | NA |
| Log fat-free mass (kg) | ||||||||
| OTU05 (Streptococcus parasanguis) | 0.00 | 0.00 | <0.001 | - | - | - | - | <0.001 |
| Log fat-free mass index (kg/m2) | ||||||||
| OTU05 (Streptococcus parasanguis) | 0.00 | 0.00 | 0.01 | - | - | - | - | <0.001 |
| Fat mass (kg) | ||||||||
| OTU05 (Streptococcus parasanguis) | 0.01 | 0.00 | 0.03 | - | - | - | - | <0.001 |
| OTU06 (Streptococcus lactarius) | 0.01 | 0.00 | 0.03 | - | - | - | - | 0.01 |
| Fat mass index (kg/m2) | ||||||||
| OTU05 (Streptococcus parasanguis) | 0.01 | 0.01 | 0.03 | - | - | - | - | <0.001 |
| OTU06 (Streptococcus lactarius) | 0.02 | 0.01 | 0.02 | - | - | - | - | 0.03 |
| Fat mass (%) | ||||||||
| OTU06 (Streptococcus lactarius) | 0.05 | 0.03 | 0.04 | - | - | - | - | NA |
| Fat mass to fat-free mass ratio | ||||||||
| OTU06 (Streptococcus lactarius) | 0.00 | 0.00 | 0.03 | - | - | - | - | NA |
| HMO Concentration | Secretor | Non-Secretor | Interaction | ||||
|---|---|---|---|---|---|---|---|
| Parameter Value | SE | p-Value | Parameter Value | SE | p-Value | p-Value | |
| Weight (kg) | |||||||
| DFLNT | 0.03 | 0.03 | 0.42 | 0.30 | 0.14 | 0.04 | 0.06 |
| Length (cm) | |||||||
| DFLNT | −0.03 | 0.11 | 0.80 | 0.95 | 0.38 | 0.01 | 0.02 |
| FLNH | −0.02 | 0.05 | 0.67 | −0.22 | 0.09 | 0.02 | 0.06 |
| Weight-for-age z-score | |||||||
| DFLNT | 0.03 | 0.04 | 0.39 | 0.33 | 0.16 | 0.04 | 0.07 |
| Length-for-age z-score | |||||||
| DFLNT | −0.01 | 0.05 | 0.82 | 0.43 | 0.19 | 0.03 | 0.03 |
| FLNH | −0.01 | 0.02 | 0.76 | −0.09 | 0.04 | 0.04 | 0.09 |
| Log fat-free mass (kg) | |||||||
| 3′SL | 0.01 | 0.01 | 0.04 | −0.01 | 0.01 | 0.41 | 0.13 |
| Fat mass (kg) | |||||||
| DFLNT | 0.01 | 0.01 | 0.58 | 0.12 | 0.06 | 0.04 | 0.06 |
| Entire cohort | |||||||
| Weight (kg) | |||||||
| FLNH | −0.03 | 0.01 | 0.04 | - | - | - | - |
| Log (DFLNH) | 0.24 | 0.11 | 0.03 | - | - | - | - |
| Length (cm) | |||||||
| Log (LNnT) | −1.11 | 0.5 | 0.03 | - | - | - | - |
| Log (DFLNH) | 1.1 | 0.36 | <0.001 | - | - | - | - |
| Length-for-age z-score | |||||||
| Log (LNnT) | −0.51 | 0.23 | 0.03 | - | - | - | - |
| Log (DFLNH) | 0.44 | 0.17 | 0.01 | - | - | - | - |
| Log fat-free mass (kg) | |||||||
| Log (DFLNH) | 0.04 | 0.02 | 0.02 | - | - | - | - |
| Fat mass (%) | |||||||
| Log (LNFP III) | −1.6 | 0.64 | 0.02 | - | - | - | - |
| Fat mass to fat-free mass ratio | |||||||
| Log (LNFP III) | −0.03 | 0.01 | 0.02 | - | - | - | - |
| HMOs (Intake) | Secretor | Non-Secretor | Interaction | ||||
|---|---|---|---|---|---|---|---|
| Parameter Value | SE | p-Value | Parameter Value | SE | p-Value | p-Value | |
| Weight (kg) | |||||||
| Log (3′SL) | 0.88 | 0.28 | <0.001 | −0.31 | 0.36 | 0.39 | 0.01 |
| 6′SL | −0.01 | 0.03 | 0.86 | −0.09 | 0.04 | 0.03 | 0.11 |
| Length (cm) | |||||||
| Log (3′SL) | 2.59 | 1.01 | 0.01 | −0.58 | 1.16 | 0.62 | 0.05 |
| Body mass index (kg/m2) | |||||||
| Log (FDSLNH) | −0.17 | 0.21 | 0.44 | −2.87 | 1.06 | 0.01 | 0.02 |
| Weight-for-length z-score | |||||||
| Log (FDSLNH) | −0.22 | 0.14 | 0.13 | −1.54 | 0.61 | 0.02 | 0.04 |
| Weight-for-age z-score | |||||||
| Log (3′SL) | 0.86 | 0.35 | 0.02 | −0.20 | 0.39 | 0.62 | 0.05 |
| 6′SL | 0.01 | 0.04 | 0.70 | −0.11 | 0.04 | 0.01 | 0.03 |
| BMI-for-age z-score | |||||||
| Log (FDSLNH) | −0.09 | 0.14 | 0.52 | −1.50 | 0.56 | 0.01 | 0.02 |
| Log fat-free mass (kg) | |||||||
| Log (3′SL) | 0.14 | 0.04 | <0.001 | −0.05 | 0.05 | 0.34 | 0.01 |
| Log fat-free mass index (kg/m2) | |||||||
| Log (3′SL) | 0.09 | 0.03 | 0.01 | −0.04 | 0.04 | 0.32 | 0.01 |
| Log (FDSLNH) | (−)0.00 | 0.01 | 0.09 | −0.19 | 0.07 | 0.01 | 0.02 |
| Fat mass (kg) | |||||||
| 6′SL | 0.00 | 0.01 | 0.83 | −0.04 | 0.02 | 0.03 | 0.07 |
| Fat mass index (kg/m2) | |||||||
| 6′SL | 0.00 | 0.04 | 0.90 | −0.01 | 0.00 | 0.04 | 0.09 |
| Entire cohort | |||||||
| Weight (kg) | |||||||
| 2′FL | 0.00 | 0.00 | 0.03 | - | - | - | - |
| 3FL | 0.09 | 0.03 | 0.01 | - | - | - | - |
| Log (DFLac) | 0.33 | 0.14 | 0.02 | - | - | - | - |
| Log (DFLNH) | 0.28 | 0.11 | 0.01 | - | - | - | - |
| Length (cm) | |||||||
| 3FL | 0.20 | 0.08 | 0.01 | - | - | - | - |
| Log (DFLNH) | 1.22 | 0.35 | <0.001 | - | - | - | - |
| Body mass index (kg/m2) | |||||||
| Log (DFLac) | 0.51 | 0.23 | 0.03 | - | - | - | - |
| Log (LSTb) | 0.83 | 0.39 | 0.04 | - | - | - | - |
| DFLNT | 0.12 | 0.05 | 0.04 | - | - | - | - |
| Weight-for-age z-score | |||||||
| 3FL | 0.10 | 0.04 | 0.01 | - | - | - | - |
| Log (DFLNH) | 0.28 | 0.13 | 0.03 | - | - | - | - |
| Length-for-age z-score | |||||||
| 3FL | 0.08 | 0.04 | 0.04 | - | - | - | - |
| Log (DFLNH) | 0.48 | 0.17 | 0.01 | - | - | - | - |
| BMI-for-age z-score | |||||||
| Log (DFLac) | 0.28 | 0.13 | 0.04 | - | - | - | - |
| DFLNT | 0.07 | 0.03 | 0.03 | - | - | - | - |
| Log fat-free mass (kg) | |||||||
| 3FL | 0.01 | 0.00 | 0.01 | - | - | - | - |
| Log (DFLac) | 0.05 | 0.02 | 0.02 | - | - | - | - |
| Log (DFLNH) | 0.04 | 0.02 | 0.01 | - | - | - | - |
| Log fat-free mass index (kg/m2) | |||||||
| 3FL | 0.01 | 0.00 | 0.04 | - | - | - | - |
| Log (DFLac) | 0.04 | 0.01 | 0.02 | - | - | - | - |
| DFLNT | 0.01 | 0.00 | 0.03 | - | - | - | - |
| Fat mass (kg) | |||||||
| 2′FL | 0.02 | 0.01 | 0.04 | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheema, A.S.; Gridneva, Z.; Furst, A.J.; Roman, A.S.; Trevenen, M.L.; Turlach, B.A.; Lai, C.T.; Stinson, L.F.; Bode, L.; Payne, M.S.; et al. Human Milk Oligosaccharides and Bacterial Profile Modulate Infant Body Composition during Exclusive Breastfeeding. Int. J. Mol. Sci. 2022, 23, 2865. https://doi.org/10.3390/ijms23052865
Cheema AS, Gridneva Z, Furst AJ, Roman AS, Trevenen ML, Turlach BA, Lai CT, Stinson LF, Bode L, Payne MS, et al. Human Milk Oligosaccharides and Bacterial Profile Modulate Infant Body Composition during Exclusive Breastfeeding. International Journal of Molecular Sciences. 2022; 23(5):2865. https://doi.org/10.3390/ijms23052865
Chicago/Turabian StyleCheema, Ali S., Zoya Gridneva, Annalee J. Furst, Ana S. Roman, Michelle L. Trevenen, Berwin A. Turlach, Ching T. Lai, Lisa F. Stinson, Lars Bode, Matthew S. Payne, and et al. 2022. "Human Milk Oligosaccharides and Bacterial Profile Modulate Infant Body Composition during Exclusive Breastfeeding" International Journal of Molecular Sciences 23, no. 5: 2865. https://doi.org/10.3390/ijms23052865
APA StyleCheema, A. S., Gridneva, Z., Furst, A. J., Roman, A. S., Trevenen, M. L., Turlach, B. A., Lai, C. T., Stinson, L. F., Bode, L., Payne, M. S., & Geddes, D. T. (2022). Human Milk Oligosaccharides and Bacterial Profile Modulate Infant Body Composition during Exclusive Breastfeeding. International Journal of Molecular Sciences, 23(5), 2865. https://doi.org/10.3390/ijms23052865