
Quantification of Species-Preferential Micropylar Chemoattraction in Arabidopsis by Fluorescein Diacetate Staining of Pollen Tubes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
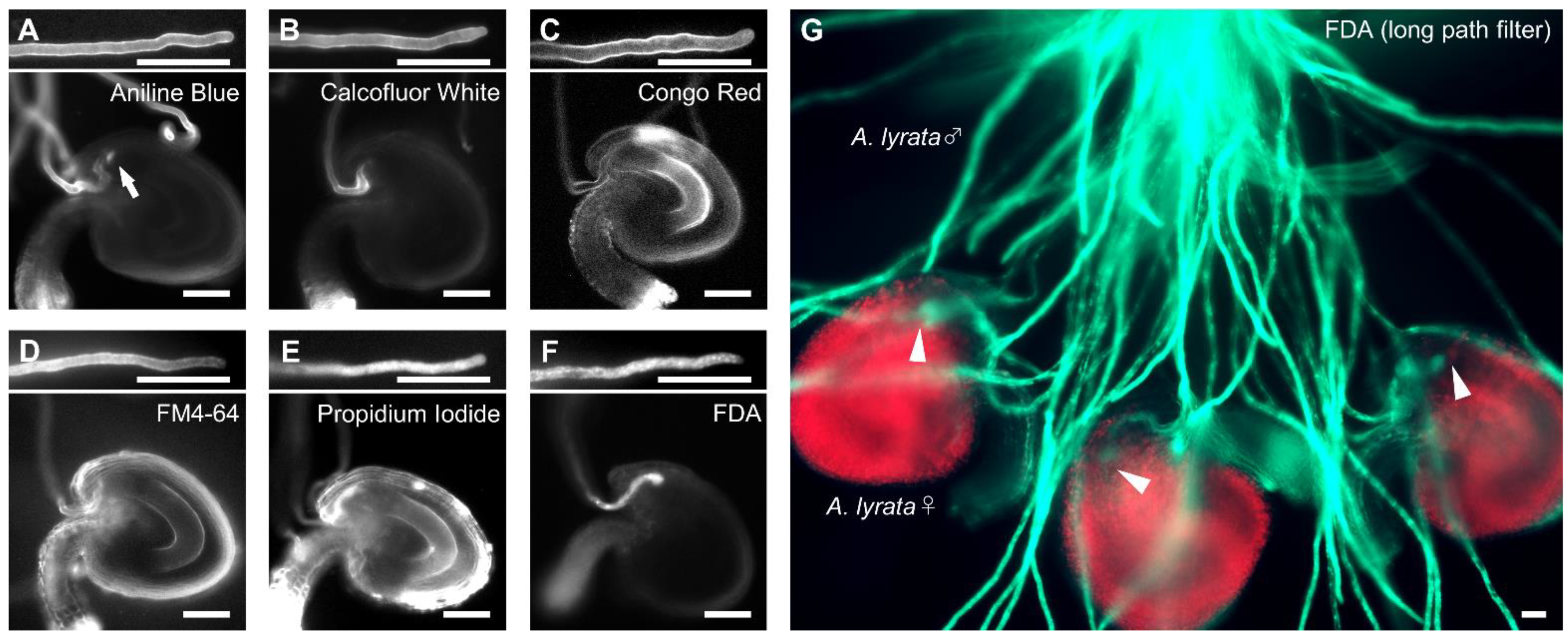
2.1. FDA Staining Enables to Directly Observe Pollen Tube Attracted into Ovules
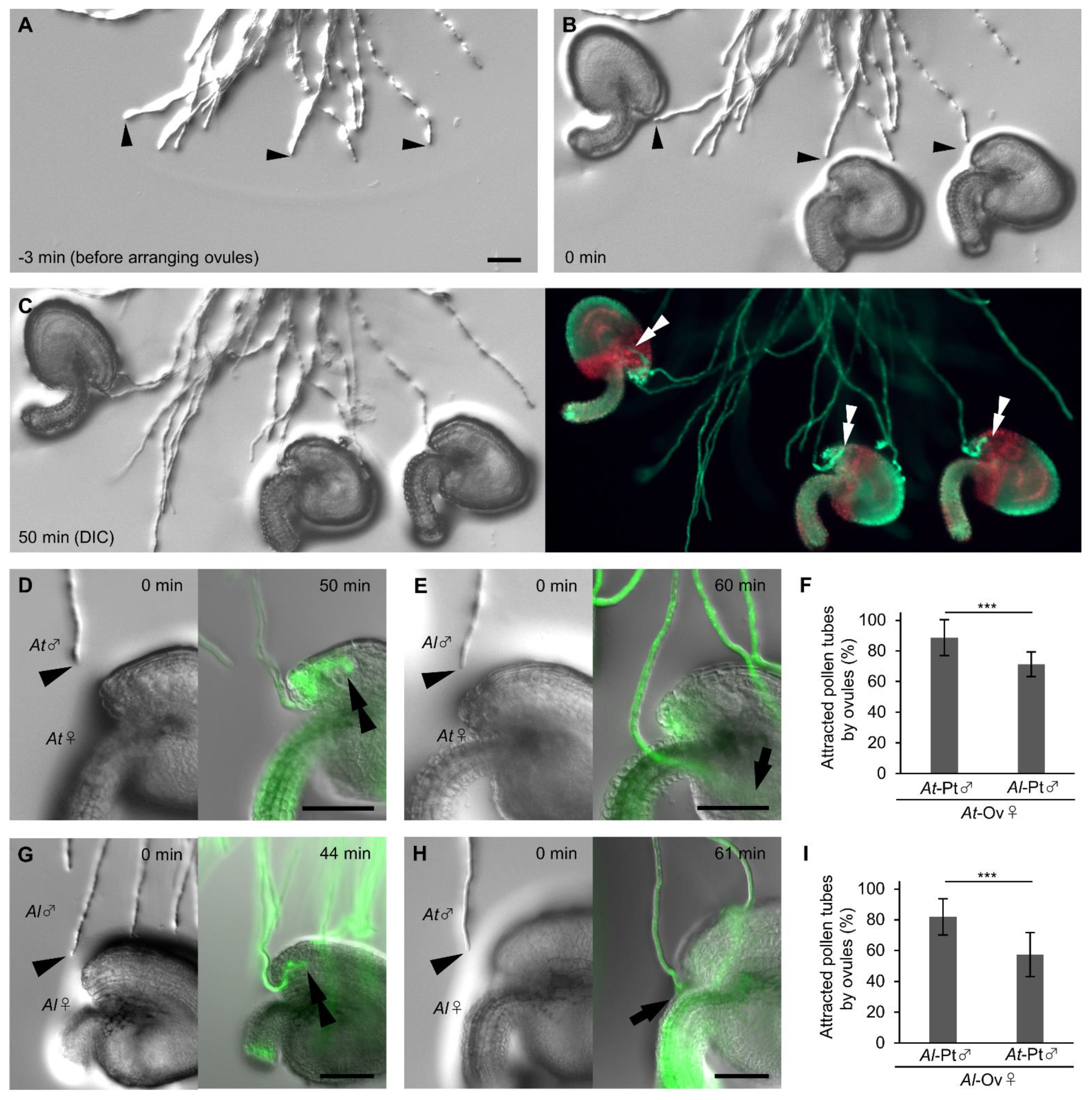
2.2. Arabidopsis Ovules Attract Pollen Tubes in a Species-Preferential Manner
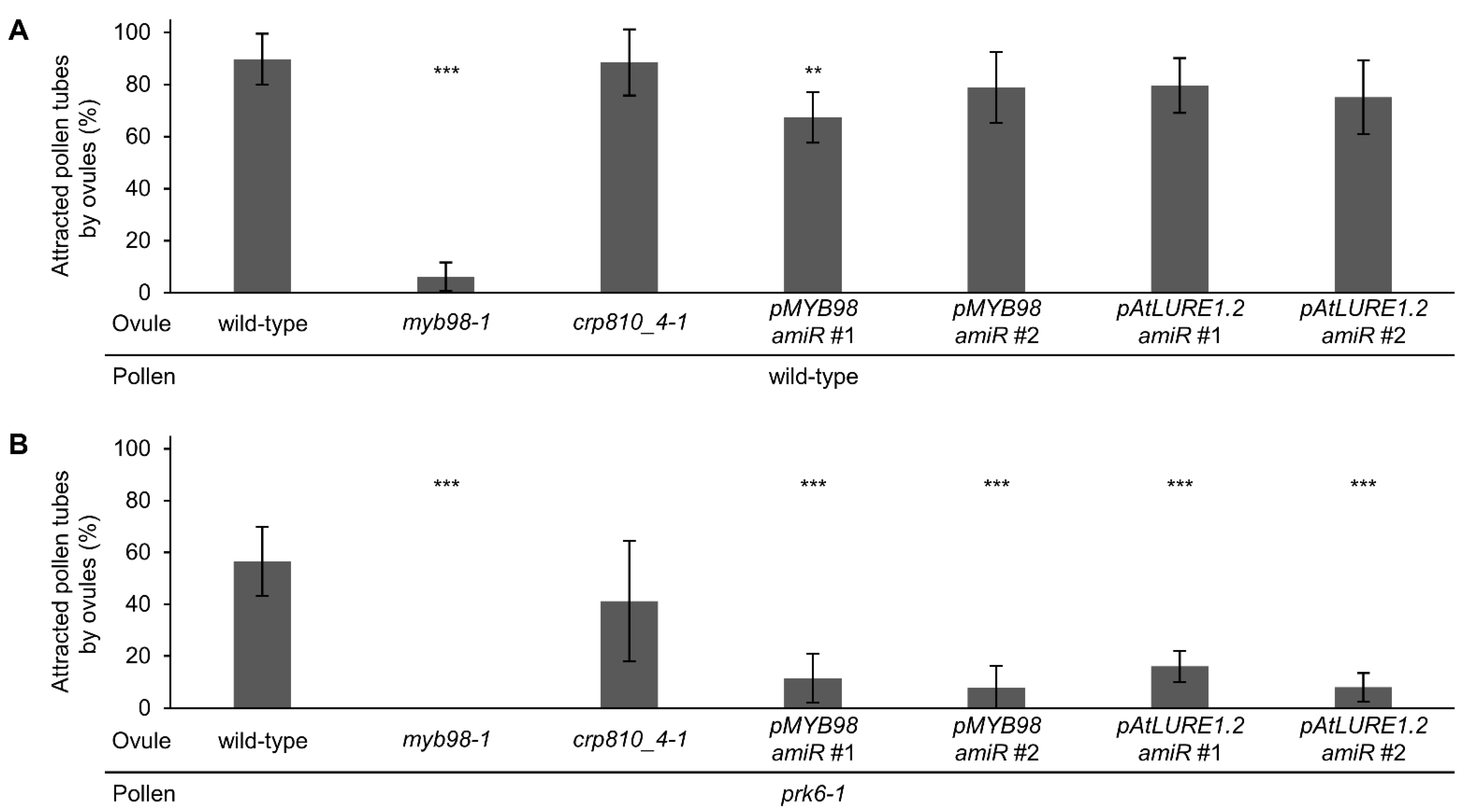
2.3. Semi In Vivo Pollen Tube Attraction Is Regulated by PRK6 and CRP810_1, CRP810_2, CRP810_3, and CRP810_4 Proteins
2.4. The Ovule Competition Assay Is More Sensitive for Detecting Species Differences in the Attraction Signal
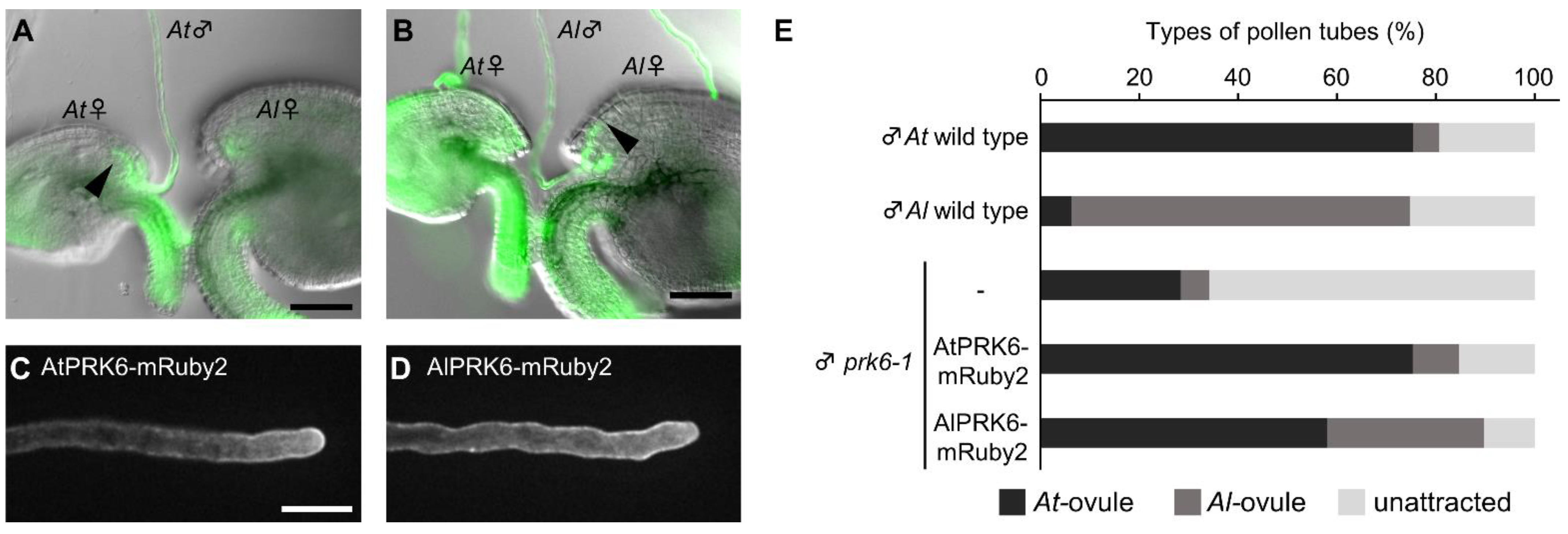
2.5. PRK6 and an Unidentified Receptor(s) of A. thaliana and A. lyrata Recognize Attractants of Their Own Species
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Growth Conditions
4.3. Pollen Tube-Ovule Interaction Assay
4.4. Chemical Florescence Staining of Pollen Tubes
4.5. Vector Construction and Plant Transformation
4.6. Localization Analysis of AtPRK6-mRuby and AlPRK6-mRuby
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lowry, D.B.; Modliszewski, J.L.; Wright, K.M.; Wu, C.A.; Willis, J.H. The strength and genetic basis of reproductive isolating barriers in flowering plants. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 3009–3021. [Google Scholar] [CrossRef] [PubMed]
- Williams, E.; Kaul, V.; Rouse, J.; Palser, B. Overgrowth of Pollen Tubes in Embryo Sacs of Rhododendron Following Interspecific Pollinations. Aust. J. Bot. 1986, 34, 413–423. [Google Scholar] [CrossRef]
- Lausser, A.; Kliwer, I.; Srilunchang, K.-O.; Dresselhaus, T. Sporophytic control of pollen tube growth and guidance in maize. J. Exp. Bot. 2010, 61, 673–682. [Google Scholar] [CrossRef]
- Meng, J.-G.; Zhang, M.-X.; Yang, W.-C.; Li, H.-J. TICKET attracts pollen tubes and mediates reproductive isolation between relative species in Brassicaceae. Sci. China Life Sci. 2019, 62, 1413–1419. [Google Scholar] [CrossRef]
- Zhong, S.; Liu, M.; Wang, Z.; Huang, Q.; Hou, S.; Xu, Y.-C.; Ge, Z.; Song, Z.; Huang, J.; Qiu, X.; et al. Cysteine-rich peptides promote interspecific genetic isolation in Arabidopsis. Science 2019, 364. [Google Scholar] [CrossRef]
- Escobar-Restrepo, J.-M.; Huck, N.; Kessler, S.; Gagliardini, V.; Gheyselinck, J.; Yang, W.-C.; Grossniklaus, U. The FERONIA Receptor-like Kinase Mediates Male-Female Interactions During Pollen Tube Reception. Science 2007, 317, 656–660. [Google Scholar] [CrossRef]
- Shimizu, K.K. Ecology meets molecular genetics in Arabidopsis. Popul. Ecol. 2002, 44, 221–233. [Google Scholar] [CrossRef]
- Kikuchi, S.; Kino, H.; Tanaka, H.; Tsujimoto, H. Pollen Tube Growth in Cross Combinations between Torenia fournieri and Fourteen Related Species. Breed. Sci. 2007, 57, 117–122. [Google Scholar] [CrossRef][Green Version]
- Fujii, S.; Tsuchimatsu, T.; Kimura, Y.; Ishida, S.; Tangpranomkorn, S.; Shimosato-Asano, H.; Iwano, M.; Furukawa, S.; Itoyama, W.; Wada, Y.; et al. A stigmatic gene confers interspecies incompatibility in the Brassicaceae. Nat. Plants 2019, 5, 731–741. [Google Scholar] [CrossRef]
- Higashiyama, T.; Yabe, S.; Sasaki, N.; Nishimura, Y.; Miyagishima, S.-Y.; Kuroiwa, H.; Kuroiwa, T. Pollen Tube Attraction by the Synergid Cell. Science 2001, 293, 1480–1483. [Google Scholar] [CrossRef]
- Okuda, S.; Tsutsui, H.; Shiina, K.; Sprunck, S.; Takeuchi, H.; Yui, R.; Kasahara, R.D.; Hamamura, Y.; Mizukami, A.; Susaki, D.; et al. Defensin-like polypeptide LUREs are pollen tube attractants secreted from synergid cells. Nature 2009, 458, 357–361. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, H.; Higashiyama, T. A Species-Specific Cluster of Defensin-Like Genes Encodes Diffusible Pollen Tube Attractants in Arabidopsis. PLoS Biol. 2012, 10, e1001449. [Google Scholar] [CrossRef]
- Márton, M.L.; Cordts, S.; Broadhvest, J.; Dresselhaus, T. Micropylar Pollen Tube Guidance by Egg Apparatus 1 of Maize. Science 2005, 307, 573–576. [Google Scholar] [CrossRef] [PubMed]
- Márton, M.L.; Fastner, A.; Uebler, S.; Dresselhaus, T. Overcoming Hybridization Barriers by the Secretion of the Maize Pollen Tube Attractant ZmEA1 from Arabidopsis Ovules. Curr. Biol. 2012, 22, 1194–1198. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, H.; Higashiyama, H.T.T. Tip-localized receptors control pollen tube growth and LURE sensing in Arabidopsis. Nature 2016, 531, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Liang, L.; Xue, Y.; Jia, P.F.; Chen, W.; Zhang, M.X.; Wang, Y.C.; Li, H.J.; Yang, W.C. A receptor heteromer mediates the male perception of female attractants in plants. Nature 2016, 531, 241–244. [Google Scholar] [CrossRef]
- Kanaoka, M.M.; Kawano, N.; Matsubara, Y.; Susaki, D.; Okuda, S.; Sasaki, N.; Higashiyama, T. Identification and characterization of TcCRP1, a pollen tube attractant from Torenia concolor. Ann. Bot. 2011, 108, 739–747. [Google Scholar] [CrossRef]
- Higashiyama, T.; Inatsugi, R.S.; Sakamoto, S.; Sasaki, N.; Mori, T.; Kuroiwa, H.; Nakada, T.; Nozaki, H.; Kuroiwa, T.; Nakano, A. Species Preferentiality of the Pollen Tube Attractant Derived from the Synergid Cell of Torenia fournieri. Plant Physiol. 2006, 142, 481–491. [Google Scholar] [CrossRef]
- Jones-Rhoades, M.W.; Borevitz, J.O.; Preuss, D. Genome-Wide Expression Profiling of the Arabidopsis Female Gametophyte Identifies Families of Small, Secreted Proteins. PLoS Genet. 2007, 3, e171. [Google Scholar] [CrossRef]
- Punwani, J.A.; Rabiger, D.S.; Drews, G.N. MYB98 Positively Regulates a Battery of Synergid-Expressed Genes Encoding Filiform Apparatus–Localized Proteins. Plant Cell 2007, 19, 2557–2568. [Google Scholar] [CrossRef]
- Susaki, D.; Suzuki, T.; Maruyama, D.; Ueda, M.; Higashiyama, T.; Kurihara, D. Dynamics of the cell fate specifications during female gametophyte development in Arabidopsis. PLoS Biol. 2021, 19, e3001123. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, H. The role of diverse LURE-type cysteine-rich peptides as signaling molecules in plant reproduction. Peptides 2021, 142, 170572. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, W.; Nagae, T.T.; Takeuchi, H.; Zhang, H.; Han, Z.; Higashiyama, T.; Chai, J. Structural basis for receptor recognition of pollen tube attraction peptides. Nat. Commun. 2017, 8, 1331. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Wang, Z.; Hou, S.; Wang, L.; Huang, Q.; Gu, H.; Dresselhaus, T.; Zhong, S.; Qu, L.-J. AtLURE1/PRK6-mediated signaling promotes conspecific micropylar pollen tube guidance. Plant Physiol. 2021, 186, 865–873. [Google Scholar] [CrossRef]
- Palanivelu, R.; Preuss, D. Distinct short-range ovule signals attract or repel Arabidopsis thaliana pollen tubes in vitro. BMC Plant Biol. 2006, 6, 7. [Google Scholar] [CrossRef]
- Kandasamy, M.K.; Nasrallah, J.B.; Nasrallah, M.E. Pollen-pistil interactions and developmental regulation of pollen tube growth in Arabidopsis. Development 1994, 120, 3405–3418. [Google Scholar] [CrossRef]
- Chebli, Y.; Kaneda, M.; Zerzour, R.; Geitmann, A. The Cell Wall of the Arabidopsis Pollen Tube—Spatial Distribution, Recycling, and Network Formation of Polysaccharides. Plant Physiol. 2012, 160, 1940–1955. [Google Scholar] [CrossRef]
- Palanivelu, R.; Brass, L.; Edlund, A.F.; Preuss, D. Pollen Tube Growth and Guidance Is Regulated by POP2, an Arabidopsis Gene that Controls GABA Levels. Cell 2003, 114, 47–59. [Google Scholar] [CrossRef]
- Rounds, C.M.; Lubeck, E.; Hepler, P.K.; Winship, L.J. Propidium Iodide Competes with Ca2+ to Label Pectin in Pollen Tubes and Arabidopsis Root Hairs. Plant Physiol. 2011, 157, 175–187. [Google Scholar] [CrossRef]
- Parton, R.M.; Fischer-Parton, S.; Watahiki, M.K.; Trewavas, A.J. Dynamics of the apical vesicle accumulation and the rate of growth are related in individual pollen tubes. J. Cell Sci. 2001, 114, 2685–2695. [Google Scholar] [CrossRef]
- Yang, H. Fluorescein diacetate used as a vital stain for labeling living pollen tubes. Plant Sci. 1986, 44, 59–63. [Google Scholar] [CrossRef]
- Wood, P.J.; Fulcher, R.; Stone, B.A. Studies on the specificity of interaction of cereal cell wall components with Congo Red and Calcofluor. Specific detection and histochemistry of (1→3),(1→4),-β-D-glucan. J. Cereal Sci. 1983, 1, 95–110. [Google Scholar] [CrossRef]
- Evans, N.; Hoyne, P.; Stone, B. Characteristics and specificity of the interaction of a fluorochrome from aniline blue (sirofluor) with polysaccharides. Carbohydr. Polym. 1984, 4, 215–230. [Google Scholar] [CrossRef]
- Iwano, M.; Ngo, Q.A.; Entani, T.; Shiba, H.; Nagai, T.; Miyawaki, A.; Isogai, A.; Grossniklaus, U.; Takayama, S. Cytoplasmic Ca2+ changes dynamically during the interaction of the pollen tube with synergid cells. Development 2012, 139, 4202–4209. [Google Scholar] [CrossRef] [PubMed]
- Rotman, B.; Papermaster, B.W. Membrane properties of living mammalian cells as studied by enzymatic hydrolysis of fluorogenic esters. Proc. Natl. Acad. Sci. USA 1966, 55, 134–141. [Google Scholar] [CrossRef]
- Kasahara, R.D.; Portereiko, M.F.; Sandaklie-Nikolova, L.; Rabiger, D.S.; Drews, G.N. MYB98Is Required for Pollen Tube Guidance and Synergid Cell Differentiation inArabidopsis. Plant Cell 2005, 17, 2981–2992. [Google Scholar] [CrossRef]
- Boavida, L.C.; McCormick, S. TECHNICAL ADVANCE: Temperature as a determinant factor for increased and reproducible in vitro pollen germination in Arabidopsis thaliana. Plant J. 2007, 52, 570–582. [Google Scholar] [CrossRef]
- Schwab, R.; Ossowski, S.; Riester, M.; Warthmann, N.; Weigel, D. Highly Specific Gene Silencing by Artificial MicroRNAs inArabidopsis. Plant Cell 2006, 18, 1121–1133. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagae, T.T.; Takeuchi, H.; Higashiyama, T. Quantification of Species-Preferential Micropylar Chemoattraction in Arabidopsis by Fluorescein Diacetate Staining of Pollen Tubes. Int. J. Mol. Sci. 2022, 23, 2722. https://doi.org/10.3390/ijms23052722
Nagae TT, Takeuchi H, Higashiyama T. Quantification of Species-Preferential Micropylar Chemoattraction in Arabidopsis by Fluorescein Diacetate Staining of Pollen Tubes. International Journal of Molecular Sciences. 2022; 23(5):2722. https://doi.org/10.3390/ijms23052722
Chicago/Turabian StyleNagae, Takuya T., Hidenori Takeuchi, and Tetsuya Higashiyama. 2022. "Quantification of Species-Preferential Micropylar Chemoattraction in Arabidopsis by Fluorescein Diacetate Staining of Pollen Tubes" International Journal of Molecular Sciences 23, no. 5: 2722. https://doi.org/10.3390/ijms23052722
APA StyleNagae, T. T., Takeuchi, H., & Higashiyama, T. (2022). Quantification of Species-Preferential Micropylar Chemoattraction in Arabidopsis by Fluorescein Diacetate Staining of Pollen Tubes. International Journal of Molecular Sciences, 23(5), 2722. https://doi.org/10.3390/ijms23052722