
RNA Packaging in the Cystovirus Bacteriophages: Dynamic Interactions during Capsid Maturation


Abstract
1. Introduction
2. The Classification of Cystoviridae Family
3. Virion Structure: Protein Components and Their Viral Location
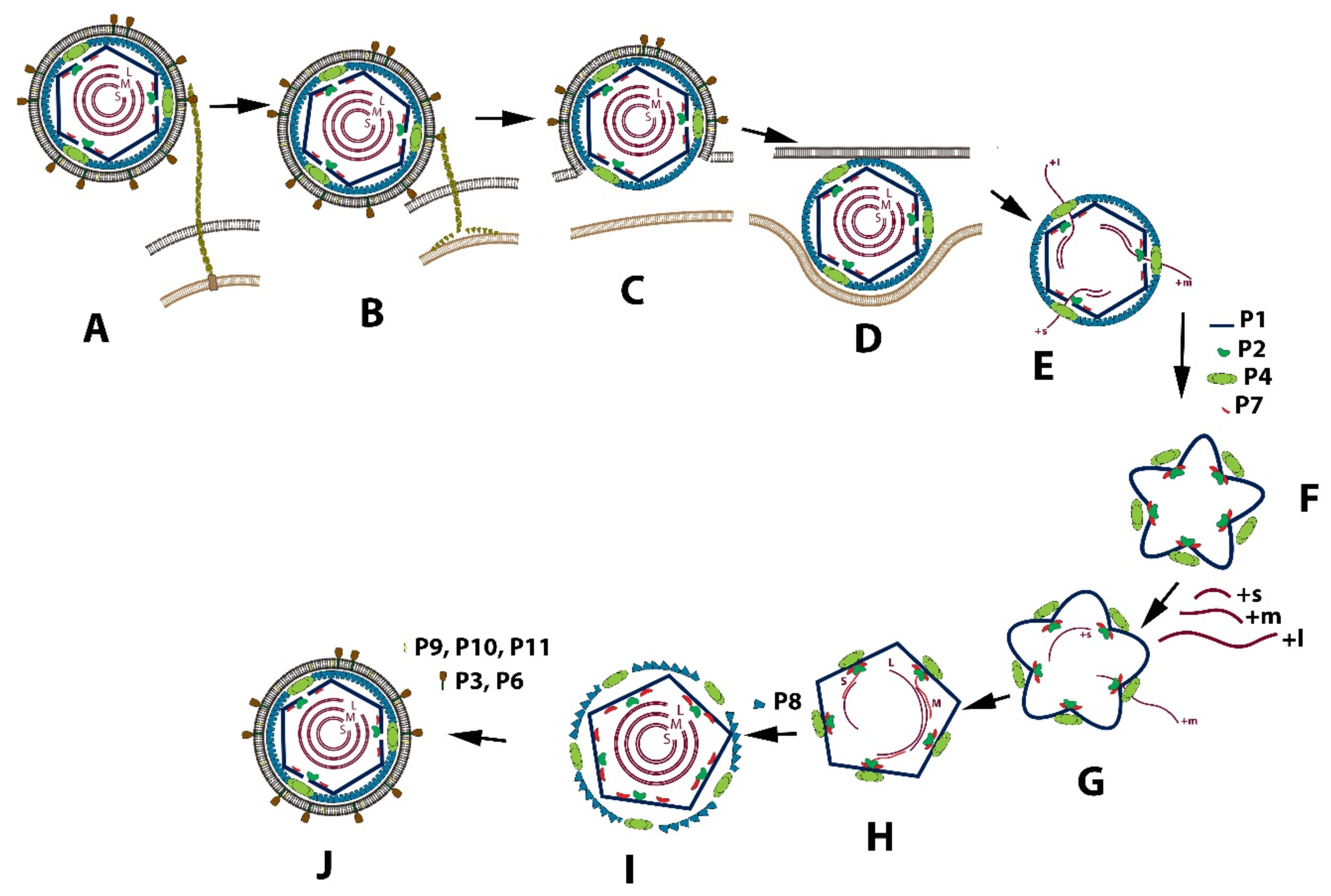
3.1. Replication Cycle
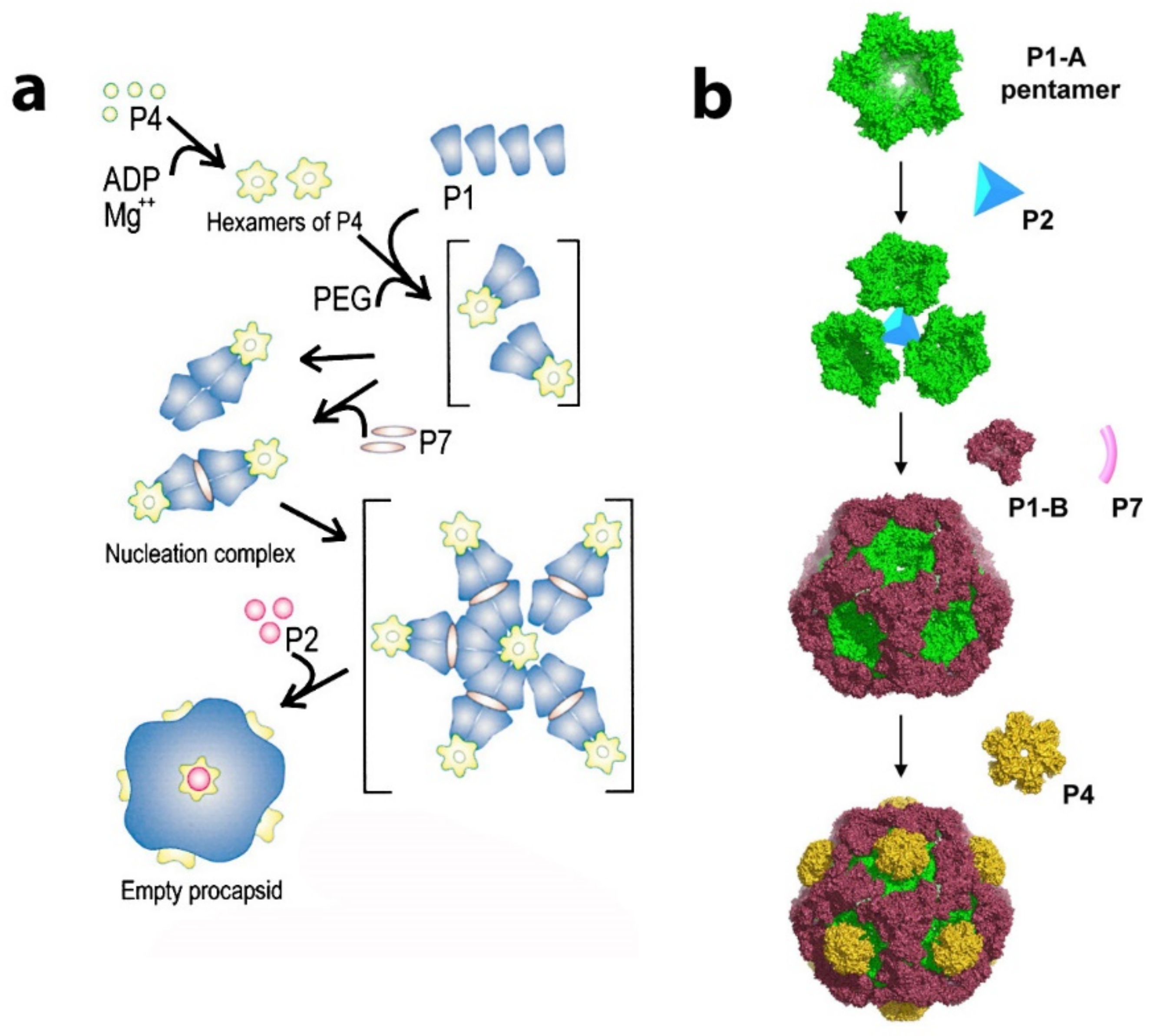
3.2. Procapsid
3.3. Nucleocapsid
3.4. Outer Envelope
4. Genome Organization
5. Viral RNA Transcription and Packaging Signals
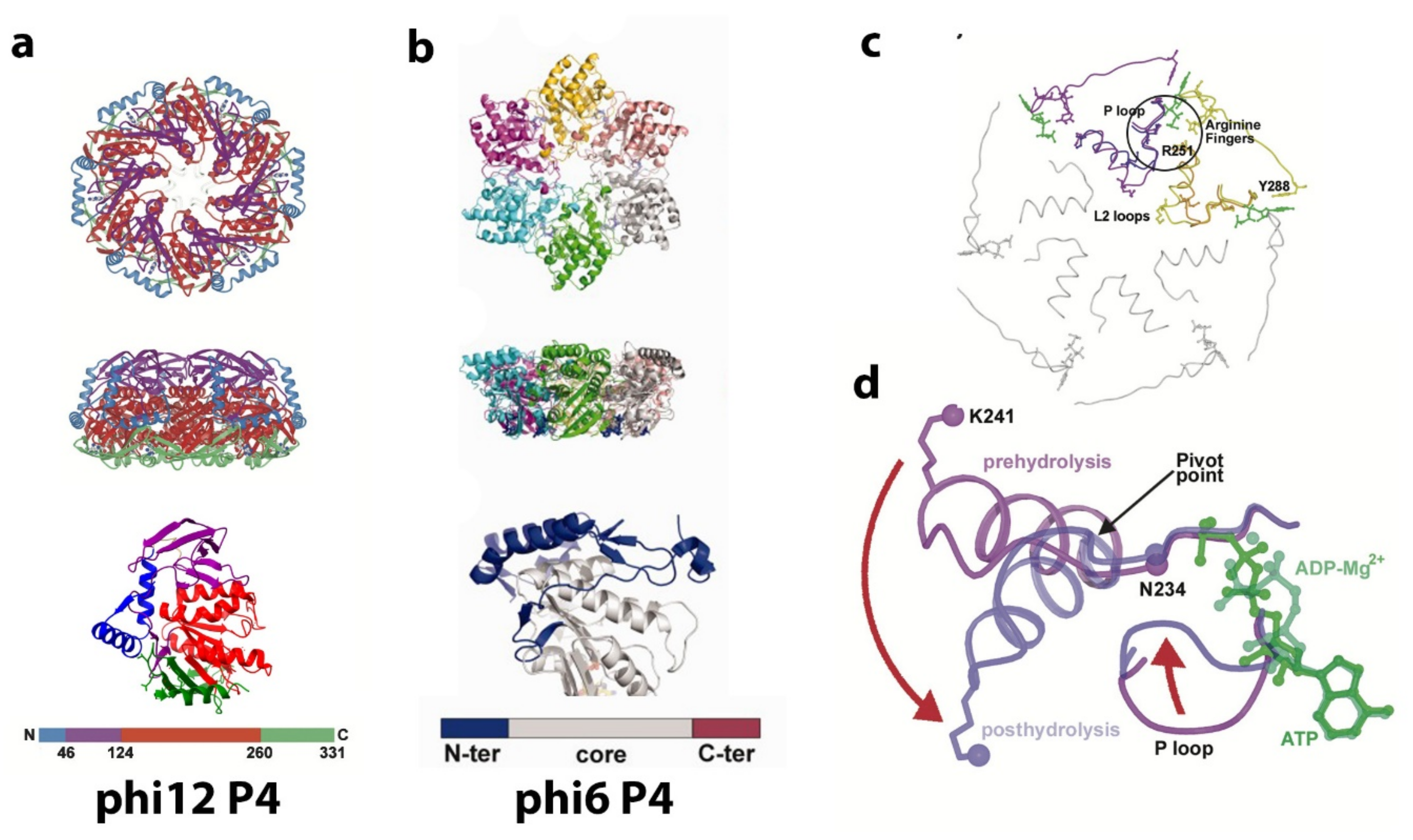
6. Maturation of the PC Mediated by Protein P1
7. P4 Portals: Occupancy and Translational Motion in the PC and NC- Implications for RNA Packaging and Viral Maturation
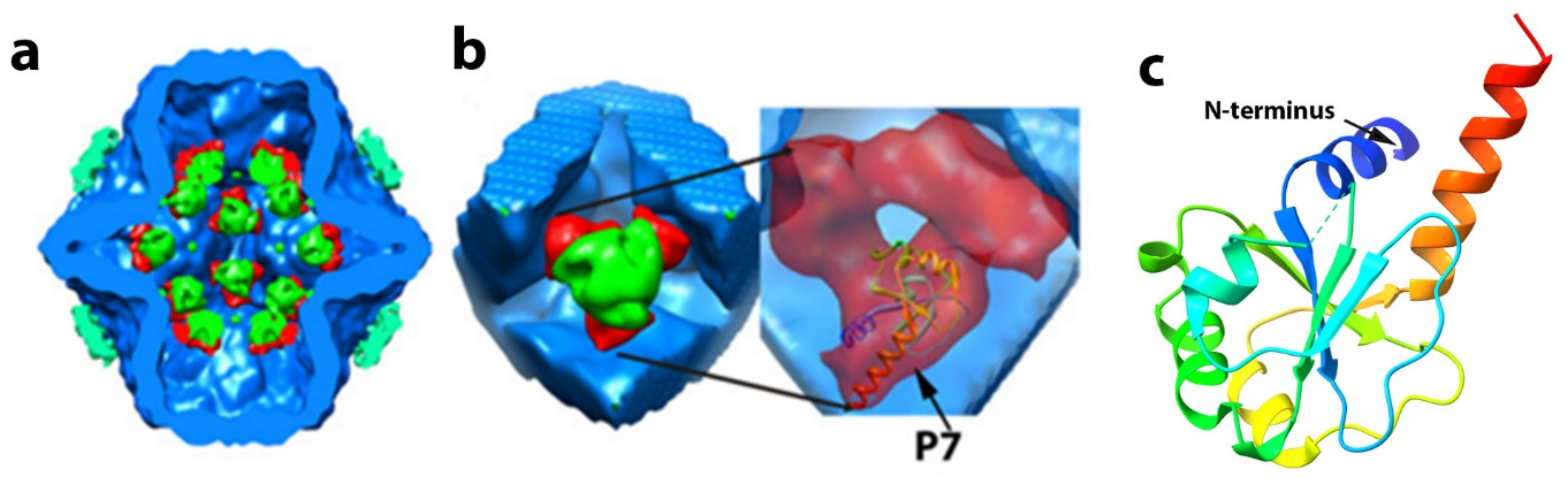
8. The Precise Function and Positioning of Protein P7
9. Final Maturation of the Nucleocapsid and Virus Particle
10. Conclusions and Suggestions for Additional Areas of Investigation
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Vidaver, A.K.; Koski, R.K.; Van Etten, J.L. Bacteriophage phi6: A Lipid-Containing Virus of Pseudomonas phaseolicola. J. Virol. 1973, 11, 799–805. [Google Scholar] [CrossRef]
- Poranen, M.M.; Mäntynen, S.; Ictv Report, C. ICTV Virus Taxonomy Profile: Cystoviridae. J. Gen. Virol. 2017, 98, 2423–2424. [Google Scholar] [CrossRef]
- Mindich, L.; Qiao, X.; Qiao, J.; Onodera, S.; Romantschuk, M.; Hoogstraten, D. Isolation of additional bacteriophages with genomes of segmented double-stranded RNA. J. Bacteriol. 1999, 181, 4505–4508. [Google Scholar] [CrossRef] [PubMed]
- Roine, E.; Nunn, D.N.; Paulin, L.; Romantschuk, M. Characterization of genes required for pilus expression in Pseudomonas syringae pathovar phaseolicola. J. Bacteriol. 1996, 178, 410–417. [Google Scholar] [CrossRef][Green Version]
- Romantschuk, M.; Bamford, D.H. Function of pili in bacteriophage phi 6 penetration. J. Gen. Virol. 1985, 66 Pt 11, 2461–2469. [Google Scholar] [CrossRef]
- Mindich, L.; Cohen, J.; Weisburd, M. Isolation of nonsense suppressor mutants in Pseudomonas. J. Bacteriol. 1976, 126, 177–182. [Google Scholar] [CrossRef]
- Qiao, X.; Sun, Y.; Qiao, J.; Di Sanzo, F.; Mindich, L. Characterization of Phi2954, a newly isolated bacteriophage containing three dsRNA genomic segments. BMC Microbiol. 2010, 10, 55. [Google Scholar] [CrossRef] [PubMed]
- Mäntynen, S.; Laanto, E.; Kohvakka, A.; Poranen, M.M.; Bamford, J.K.H.; Ravantti, J.J. New enveloped dsRNA phage from freshwater habitat. J. Gen. Virol. 2015, 96, 1180–1189. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Lu, S.; Shen, W.; Zhao, X.; Shen, M.; Tan, Y.; Li, G.; Li, M.; Wang, J.; Hu, F.; et al. Characterization of the first double-stranded RNA bacteriophage infecting Pseudomonas aeruginosa. Sci. Rep. 2016, 6, 38795. [Google Scholar] [CrossRef]
- Caldentey, J.; Bamford, D.H. The lytic enzyme of the Pseudomonas phage phi 6. Purification and biochemical characterization. Biochim. Biophys. Acta 1992, 1159, 44–50. [Google Scholar] [CrossRef]
- Bamford, D.H.; Palva, E.T. Structure of the lipid-containing bacteriophage phi 6. Disruption by Triton X-100 treatment. Biochim. Biophys. Acta 1980, 601, 245–259. [Google Scholar] [CrossRef]
- Bamford, D.H.; Mindich, L. Electron microscopy of cells infected with nonsense mutants of bacteriophage phi 6. Virology 1980, 107, 222–228. [Google Scholar] [CrossRef]
- Mindich, L.; Davidoff-Abelson, R. The characterization of a 120 S particle formed during phi 6 infection. Virology 1980, 103, 386–391. [Google Scholar] [CrossRef]
- Ktistakis, N.T.; Lang, D. The dodecahedral framework of the bacteriophage phi 6 nucleocapsid is composed of protein P1. J. Virol. 1987, 61, 2621–2623. [Google Scholar] [CrossRef]
- Olkkonen, V.M.; Bamford, D.H. The nucleocapsid of the lipid-containing double-stranded RNA bacteriophage phi 6 contains a protein skeleton consisting of a single polypeptide species. J. Virol. 1987, 61, 2362–2367. [Google Scholar] [CrossRef]
- Borodavka, A.; Desselberger, U.; Patton, J.T. Genome packaging in multi-segmented dsRNA viruses: Distinct mechanisms with similar outcomes. Curr. Opin. Virol. 2018, 33, 106–112. [Google Scholar] [CrossRef]
- Huiskonen, J.T.; Jäälinoja, H.T.; Briggs, J.A.; Fuller, S.D.; Butcher, S.J. Structure of a hexameric RNA packaging motor in a viral polymerase complex. J. Struct. Biol. 2007, 158, 156–164. [Google Scholar] [CrossRef]
- De Haas, F.; Paatero, A.O.; Mindich, L.; Bamford, D.H.; Fuller, S.D. A symmetry mismatch at the site of RNA packaging in the polymerase complex of dsRNA bacteriophage phi6. J. Mol. Biol. 1999, 294, 357–372. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Cheng, R.H.; Berriman, J.; Rice, W.J.; Stokes, D.L.; Katz, A.; Morgan, D.G.; Gottlieb, P. Three-dimensional structure of the enveloped bacteriophage phi12: An incomplete T = 13 lattice is superposed on an enclosed T = 1 shell. PLoS ONE 2009, 4, e6850. [Google Scholar] [CrossRef]
- Mindich, L. Precise packaging of the three genomic segments of the double-stranded-RNA bacteriophage phi6. Microbiol. Mol. Biol. Rev. 1999, 63, 149–160. [Google Scholar] [CrossRef]
- Nemecek, D.; Boura, E.; Wu, W.; Cheng, N.; Plevka, P.; Qiao, J.; Mindich, L.; Heymann, J.B.; Hurley, J.H.; Steven, A.C. Subunit folds and maturation pathway of a dsRNA virus capsid. Structure 2013, 21, 1374–1383. [Google Scholar] [CrossRef]
- Huiskonen, J.T.; de Haas, F.; Bubeck, D.; Bamford, D.H.; Fuller, S.D.; Butcher, S.J. Structure of the bacteriophage phi6 nucleocapsid suggests a mechanism for sequential RNA packaging. Structure 2006, 14, 1039–1048. [Google Scholar] [CrossRef]
- Veesler, D.; Johnson, J.E. Cystovirus maturation at atomic resolution. Structure 2013, 21, 1266–1268. [Google Scholar] [CrossRef] [PubMed]
- Butcher, S.J.; Dokland, T.; Ojala, P.M.; Bamford, D.H.; Fuller, S.D. Intermediates in the assembly pathway of the double-stranded RNA virus phi6. EMBO J. 1997, 16, 4477–4487. [Google Scholar] [CrossRef]
- Qiao, X.; Qiao, J.; Mindich, L. Stoichiometric packaging of the three genomic segments of double-stranded RNA bacteriophage phi6. Proc. Natl. Acad. Sci. USA 1997, 94, 4074–4079. [Google Scholar] [CrossRef] [PubMed]
- Mindich, L. Packaging, replication and recombination of the segmented genome of bacteriophage Phi6 and its relatives. Virus Res. 2004, 101, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Nemecek, D.; Cheng, N.; Qiao, J.; Mindich, L.; Steven, A.C.; Heymann, J.B. Stepwise expansion of the bacteriophage ϕ6 procapsid: Possible packaging intermediates. J. Mol. Biol. 2011, 414, 260–271. [Google Scholar] [CrossRef][Green Version]
- Oliveira, L.M.; Ye, Z.; Katz, A.; Alimova, A.; Wei, H.; Herman, G.T.; Gottlieb, P. Component tree analysis of cystovirus φ6 nucleocapsid Cryo-EM single particle reconstructions. PLoS ONE 2018, 13, e0188858. [Google Scholar] [CrossRef]
- Ilca, S.L.; Sun, X.; El Omari, K.; Kotecha, A.; de Haas, F.; DiMaio, F.; Grimes, J.M.; Stuart, D.I.; Poranen, M.M.; Huiskonen, J.T. Multiple liquid crystalline geometries of highly compacted nucleic acid in a dsRNA virus. Nature 2019, 570, 252–256. [Google Scholar] [CrossRef]
- Gottlieb, P.; Strassman, J.; Qiao, X.Y.; Frucht, A.; Mindich, L. In vitro replication, packaging, and transcription of the segmented double-stranded RNA genome of bacteriophage phi 6: Studies with procapsids assembled from plasmid-encoded proteins. J. Bacteriol. 1990, 172, 5774–5782. [Google Scholar] [CrossRef]
- Poranen, M.M.; Paatero, A.O.; Tuma, R.; Bamford, D.H. Self-assembly of a viral molecular machine from purified protein and RNA constituents. Mol. Cell 2001, 7, 845–854. [Google Scholar] [CrossRef]
- Katz, G.; Wei, H.; Alimova, A.; Katz, A.; Morgan, D.G.; Gottlieb, P. Protein P7 of the cystovirus φ6 is located at the three-fold axis of the unexpanded procapsid. PLoS ONE 2012, 7, e47489. [Google Scholar] [CrossRef] [PubMed]
- Poranen, M.M.; Butcher, S.J.; Simonov, V.M.; Laurinmäki, P.; Bamford, D.H. Roles of the minor capsid protein P7 in the assembly and replication of double-stranded RNA bacteriophage phi6. J. Mol. Biol. 2008, 383, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, P.; Wei, H.; Potgieter, C.; Toporovsky, I. Characterization of phi 12, a bacteriophage related to phi 6: Nucleotide sequence of the small and middle double-stranded RNA. Virology 2002, 293, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Hoogstraten, D.; Qiao, X.; Sun, Y.; Hu, A.; Onodera, S.; Mindich, L. Characterization of phi8, a bacteriophage containing three double-stranded RNA genomic segments and distantly related to Phi6. Virology 2000, 272, 218–224. [Google Scholar] [CrossRef]
- Qiao, X.; Qiao, J.; Onodera, S.; Mindich, L. Characterization of phi 13, a bacteriophage related to phi 6 and containing three dsRNA genomic segments. Virology 2000, 275, 218–224. [Google Scholar] [CrossRef]
- Hu, G.B.; Wei, H.; Rice, W.J.; Stokes, D.L.; Gottlieb, P. Electron cryo-tomographic structure of cystovirus phi 12. Virology 2008, 372, 1–9. [Google Scholar] [CrossRef]
- Leo-Macias, A.; Katz, G.; Wei, H.; Alimova, A.; Katz, A.; Rice, W.J.; Diaz-Avalos, R.; Hu, G.B.; Stokes, D.L.; Gottlieb, P. Toroidal surface complexes of bacteriophage ϕ12 are responsible for host-cell attachment. Virology 2011, 414, 103–109. [Google Scholar] [CrossRef]
- Bamford, D.H.; Romantschuk, M.; Somerharju, P.J. Membrane fusion in prokaryotes: Bacteriophage phi 6 membrane fuses with the Pseudomonas syringae outer membrane. EMBO J. 1987, 6, 1467–1473. [Google Scholar] [CrossRef]
- Semancik, J.S.; Vidaver, A.K.; Van Etten, J.L. Characterization of segmented double-helical RNA from bacteriophage phi6. J. Mol. Biol. 1973, 78, 617–625. [Google Scholar] [CrossRef]
- McGraw, T.; Mindich, L.; Frangione, B. Nucleotide sequence of the small double-stranded RNA segment of bacteriophage phi 6: Novel mechanism of natural translational control. J. Virol. 1986, 58, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Mindich, L.; Nemhauser, I.; Gottlieb, P.; Romantschuk, M.; Carton, J.; Frucht, S.; Strassman, J.; Bamford, D.H.; Kalkkinen, N. Nucleotide sequence of the large double-stranded RNA segment of bacteriophage phi 6: Genes specifying the viral replicase and transcriptase. J. Virol. 1988, 62, 1180–1185. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, P.; Potgieter, C.; Wei, H.; Toporovsky, I. Characterization of phi12, a bacteriophage related to phi6: Nucleotide sequence of the large double-stranded RNA. Virology 2002, 295, 266–271. [Google Scholar] [CrossRef]
- Mäntynen, S.; Sundberg, L.R.; Poranen, M.M. Recognition of six additional cystoviruses: Pseudomonas virus phi6 is no longer the sole species of the family Cystoviridae. Arch. Virol. 2018, 163, 1117–1124. [Google Scholar] [CrossRef]
- Frilander, M.; Poranen, M.; Bamford, D.H. The large genome segment of dsRNA bacteriophage phi6 is the key regulator in the in vitro minus and plus strand synthesis. RNA 1995, 1, 510–518. [Google Scholar]
- Qiao, X.; Sun, Y.; Qiao, J.; Mindich, L. The role of host protein YajQ in the temporal control of transcription in bacteriophage Phi6. Proc. Natl. Acad. Sci. USA 2008, 105, 15956–15960. [Google Scholar] [CrossRef]
- Qiao, X.; Sun, Y.; Qiao, J.; Mindich, L. Interaction of a host protein with core complexes of bacteriophage phi6 to control transcription. J. Virol. 2010, 84, 4821–4825. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Qiao, J.; Mindich, L. The template specificity of bacteriophage Phi6 RNA polymerase. J. Virol. 2013, 87, 10190–10194. [Google Scholar] [CrossRef]
- Gottlieb, P.; Qiao, X.; Strassman, J.; Frilander, M.; Mindich, L. Identification of the packaging regions within the genomic RNA segments of bacteriophage phi 6. Virology 1994, 200, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Pirttimaa, M.J.; Bamford, D.H. RNA secondary structures of the bacteriophage phi6 packaging regions. RNA 2000, 6, 880–889. [Google Scholar] [CrossRef]
- Onodera, S.; Qiao, X.; Qiao, J.; Mindich, L. Isolation of a mutant that changes genomic packaging specificity in phi6. Virology 1998, 252, 438–442. [Google Scholar] [CrossRef]
- Qiao, J.; Qiao, X.; Sun, Y.; Mindich, L. Isolation and analysis of mutants of double-stranded-RNA bacteriophage phi6 with altered packaging specificity. J. Bacteriol. 2003, 185, 4572–4577. [Google Scholar] [CrossRef] [PubMed]
- El Omari, K.; Sutton, G.; Ravantti, J.J.; Zhang, H.; Walter, T.S.; Grimes, J.M.; Bamford, D.H.; Stuart, D.I.; Mancini, E.J. Plate tectonics of virus shell assembly and reorganization in phage φ8, a distant relative of mammalian reoviruses. Structure 2013, 21, 1384–1395. [Google Scholar] [CrossRef]
- Kainov, D.E.; Butcher, S.J.; Bamford, D.H.; Tuma, R. Conserved intermediates on the assembly pathway of double-stranded RNA bacteriophages. J. Mol. Biol. 2003, 328, 791–804. [Google Scholar] [CrossRef]
- Juuti, J.T.; Bamford, D.H. Protein P7 of phage phi6 RNA polymerase complex, acquiring of RNA packaging activity by in vitro assembly of the purified protein onto deficient particles. J. Mol. Biol. 1997, 266, 891–900. [Google Scholar] [CrossRef]
- Yang, H.; Makeyev, E.V.; Bamford, D.H. Comparison of polymerase subunits from double-stranded RNA bacteriophages. J. Virol. 2001, 75, 11088–11095. [Google Scholar] [CrossRef][Green Version]
- Makeyev, E.V.; Bamford, D.H. Replicase activity of purified recombinant protein P2 of double-stranded RNA bacteriophage phi6. EMBO J. 2000, 19, 124–133. [Google Scholar] [CrossRef]
- Juuti, J.T.; Bamford, D.H.; Tuma, R.; Thomas, G.J., Jr. Structure and NTPase activity of the RNA-translocating protein (P4) of bacteriophage phi 6. J. Mol. Biol. 1998, 279, 347–359. [Google Scholar] [CrossRef]
- Poranen, M.M.; Tuma, R. Self-assembly of double-stranded RNA bacteriophages. Virus Res. 2004, 101, 93–100. [Google Scholar] [CrossRef]
- Sun, X.; Bamford, D.H.; Poranen, M.M. Probing, by self-assembly, the number of potential binding sites for minor protein subunits in the procapsid of double-stranded RNA bacteriophage Φ6. J. Virol. 2012, 86, 12208–12216. [Google Scholar] [CrossRef]
- Nemecek, D.; Heymann, J.B.; Qiao, J.; Mindich, L.; Steven, A.C. Cryo-electron tomography of bacteriophage phi6 procapsids shows random occupancy of the binding sites for RNA polymerase and packaging NTPase. J. Struct. Biol. 2010, 171, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Sen, A.; Heymann, J.B.; Cheng, N.; Qiao, J.; Mindich, L.; Steven, A.C. Initial location of the RNA-dependent RNA polymerase in the bacteriophage Phi6 procapsid determined by cryo-electron microscopy. J. Biol. Chem. 2008, 283, 12227–12231. [Google Scholar] [CrossRef] [PubMed]
- Mancini, E.J.; Kainov, D.E.; Grimes, J.M.; Tuma, R.; Bamford, D.H.; Stuart, D.I. Atomic snapshots of an RNA packaging motor reveal conformational changes linking ATP hydrolysis to RNA translocation. Cell 2004, 118, 743–755. [Google Scholar] [CrossRef] [PubMed]
- El Omari, K.; Meier, C.; Kainov, D.; Sutton, G.; Grimes, J.M.; Poranen, M.M.; Bamford, D.H.; Tuma, R.; Stuart, D.I.; Mancini, E.J. Tracking in atomic detail the functional specializations in viral RecA helicases that occur during evolution. Nucleic Acids Res. 2013, 41, 9396–9410. [Google Scholar] [CrossRef]
- Catalano, C.E. The terminase enzyme from bacteriophage lambda: A DNA-packaging machine. Cell Mol. Life Sci. 2000, 57, 128–148. [Google Scholar] [CrossRef]
- Moore, S.D.; Prevelige, P.E., Jr. DNA packaging: A new class of molecular motors. Curr. Biol. 2002, 12, R96–R98. [Google Scholar] [CrossRef][Green Version]
- Smith, D.E.; Tans, S.J.; Smith, S.B.; Grimes, S.; Anderson, D.L.; Bustamante, C. The bacteriophage straight phi29 portal motor can package DNA against a large internal force. Nature 2001, 413, 748–752. [Google Scholar] [CrossRef]
- Chen, H.; Meisburger, S.P.; Pabit, S.A.; Sutton, J.L.; Webb, W.W.; Pollack, L. Ionic strength-dependent persistence lengths of single-stranded RNA and DNA. Proc. Natl. Acad. Sci. USA 2012, 109, 799–804. [Google Scholar] [CrossRef]
- Kainov, D.E.; Pirttimaa, M.; Tuma, R.; Butcher, S.J.; Thomas, G.J., Jr.; Bamford, D.H.; Makeyev, E.V. RNA packaging device of double-stranded RNA bacteriophages, possibly as simple as hexamer of P4 protein. J. Biol. Chem. 2003, 278, 48084–48091. [Google Scholar] [CrossRef]
- Kainov, D.E.; Lísal, J.; Bamford, D.H.; Tuma, R. Packaging motor from double-stranded RNA bacteriophage phi12 acts as an obligatory passive conduit during transcription. Nucleic Acids Res. 2004, 32, 3515–3521. [Google Scholar] [CrossRef][Green Version]
- Lísal, J.; Lam, T.T.; Kainov, D.E.; Emmett, M.R.; Marshall, A.G.; Tuma, R. Functional visualization of viral molecular motor by hydrogen-deuterium exchange reveals transient states. Nat. Struct. Mol. Biol. 2005, 12, 460–466. [Google Scholar] [CrossRef]
- Paatero, A.O.; Mindich, L.; Bamford, D.H. Mutational analysis of the role of nucleoside triphosphatase P4 in the assembly of the RNA polymerase complex of bacteriophage phi6. J. Virol. 1998, 72, 10058–10065. [Google Scholar] [CrossRef]
- Krishnan, A.; Burroughs, A.M.; Iyer, L.M.; Aravind, L. Comprehensive classification of ABC ATPases and their functional radiation in nucleoprotein dynamics and biological conflict systems. Nucleic Acids Res. 2020, 48, 10045–10075. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, P.; Strassman, J.; Mindich, L. Protein P4 of the bacteriophage phi 6 procapsid has a nucleoside triphosphate-binding site with associated nucleoside triphosphate phosphohydrolase activity. J. Virol. 1992, 66, 6220–6222. [Google Scholar] [CrossRef]
- Walker, J.E.; Saraste, M.; Runswick, M.J.; Gay, N.J. Distantly related sequences in the alpha- and beta-subunits of ATP synthase, myosin, kinases and other ATP-requiring enzymes and a common nucleotide binding fold. EMBO J. 1982, 1, 945–951. [Google Scholar] [CrossRef] [PubMed]
- Carpino, J. Structure and Function in Bacteriophage Phi6. Ph.D. Thesis, The City University of New York, New York, NY, USA, 2014. [Google Scholar]
- Lísal, J.; Kainov, D.E.; Lam, T.T.; Emmett, M.R.; Wei, H.; Gottlieb, P.; Marshall, A.G.; Tuma, R. Interaction of packaging motor with the polymerase complex of dsRNA bacteriophage. Virology 2006, 351, 73–79. [Google Scholar] [CrossRef][Green Version]
- Day, L.A.; Mindich, L. The molecular weight of bacteriophage phi 6 and its nucleocapsid. Virology 1980, 103, 376–385. [Google Scholar] [CrossRef]
- Gottlieb, P.; Strassman, J.; Frucht, A.; Qiao, X.Y.; Mindich, L. In vitro packaging of the bacteriophage phi 6 ssRNA genomic precursors. Virology 1991, 181, 589–594. [Google Scholar] [CrossRef]
- Nemecek, D.; Qiao, J.; Mindich, L.; Steven, A.C.; Heymann, J.B. Packaging accessory protein P7 and polymerase P2 have mutually occluding binding sites inside the bacteriophage 6 procapsid. J. Virol. 2012, 86, 11616–11624. [Google Scholar] [CrossRef]
- Alimova, A.; Wei, H.; Katz, A.; Spatz, L.; Gottlieb, P. The ϕ6 cystovirus protein P7 becomes accessible to antibodies in the transcribing nucleocapsid: A probe for viral structural elements. PLoS ONE 2015, 10, e0122160. [Google Scholar]
- Eryilmaz, E.; Benach, J.; Su, M.; Seetharaman, J.; Dutta, K.; Wei, H.; Gottlieb, P.; Hunt, J.F.; Ghose, R. Structure and dynamics of the P7 protein from the bacteriophage phi 12. J. Mol. Biol. 2008, 382, 402–422. [Google Scholar] [CrossRef]
- Alphonse, S.; Arnold, J.J.; Bhattacharya, S.; Wang, H.; Kloss, B.; Cameron, C.E.; Ghose, R. Cystoviral polymerase complex protein P7 uses its acidic C-terminal tail to regulate the RNA-directed RNA polymerase P2. J. Mol. Biol. 2014, 426, 2580–2593. [Google Scholar] [CrossRef] [PubMed]
- Jäälinoja, H.T.; Huiskonen, J.T.; Butcher, S.J. Electron cryomicroscopy comparison of the architectures of the enveloped bacteriophages phi6 and phi8. Structure 2007, 15, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Lyytinen, O.L.; Starkova, D.; Poranen, M.M. Microbial production of lipid-protein vesicles using enveloped bacteriophage phi6. Microb. Cell. Fact. 2019, 18, 29. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Molar Weight, KDa 1 | Copies per Virion 2 | Function |
|---|---|---|---|
| P1 | 93.0 | 120 or 60 dimers | Major structural protein |
| P2 | 74.8 | 12 | RNA-directed RNA polymerase |
| P4 | 35.0 | 72 (12 hexamers) | NTPase packaging motor, transcription |
| P7 | 17.2 | 60 | Assembly cofactor; packaging cofactor |
| P5/P11 | 24.0 | Lytic endopeptidase; murein peptidase | |
| P8 | 15.8 | 600 (200 trimers); 720 | Shell |
| P6 | 21.0 | Integral membrane protein | |
| P3 | 84.0 | Binding protein | |
| P9 | 9.5 | Major envelope protein; | |
| P10 | 6.0 | Putative holin protein |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gottlieb, P.; Alimova, A. RNA Packaging in the Cystovirus Bacteriophages: Dynamic Interactions during Capsid Maturation. Int. J. Mol. Sci. 2022, 23, 2677. https://doi.org/10.3390/ijms23052677
Gottlieb P, Alimova A. RNA Packaging in the Cystovirus Bacteriophages: Dynamic Interactions during Capsid Maturation. International Journal of Molecular Sciences. 2022; 23(5):2677. https://doi.org/10.3390/ijms23052677
Chicago/Turabian StyleGottlieb, Paul, and Aleksandra Alimova. 2022. "RNA Packaging in the Cystovirus Bacteriophages: Dynamic Interactions during Capsid Maturation" International Journal of Molecular Sciences 23, no. 5: 2677. https://doi.org/10.3390/ijms23052677
APA StyleGottlieb, P., & Alimova, A. (2022). RNA Packaging in the Cystovirus Bacteriophages: Dynamic Interactions during Capsid Maturation. International Journal of Molecular Sciences, 23(5), 2677. https://doi.org/10.3390/ijms23052677