
BdGUCD1 and Cyclic GMP Are Required for Responses of Brachypodium distachyon to Fusarium pseudograminearum in the Mechanism Involving Jasmonate

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
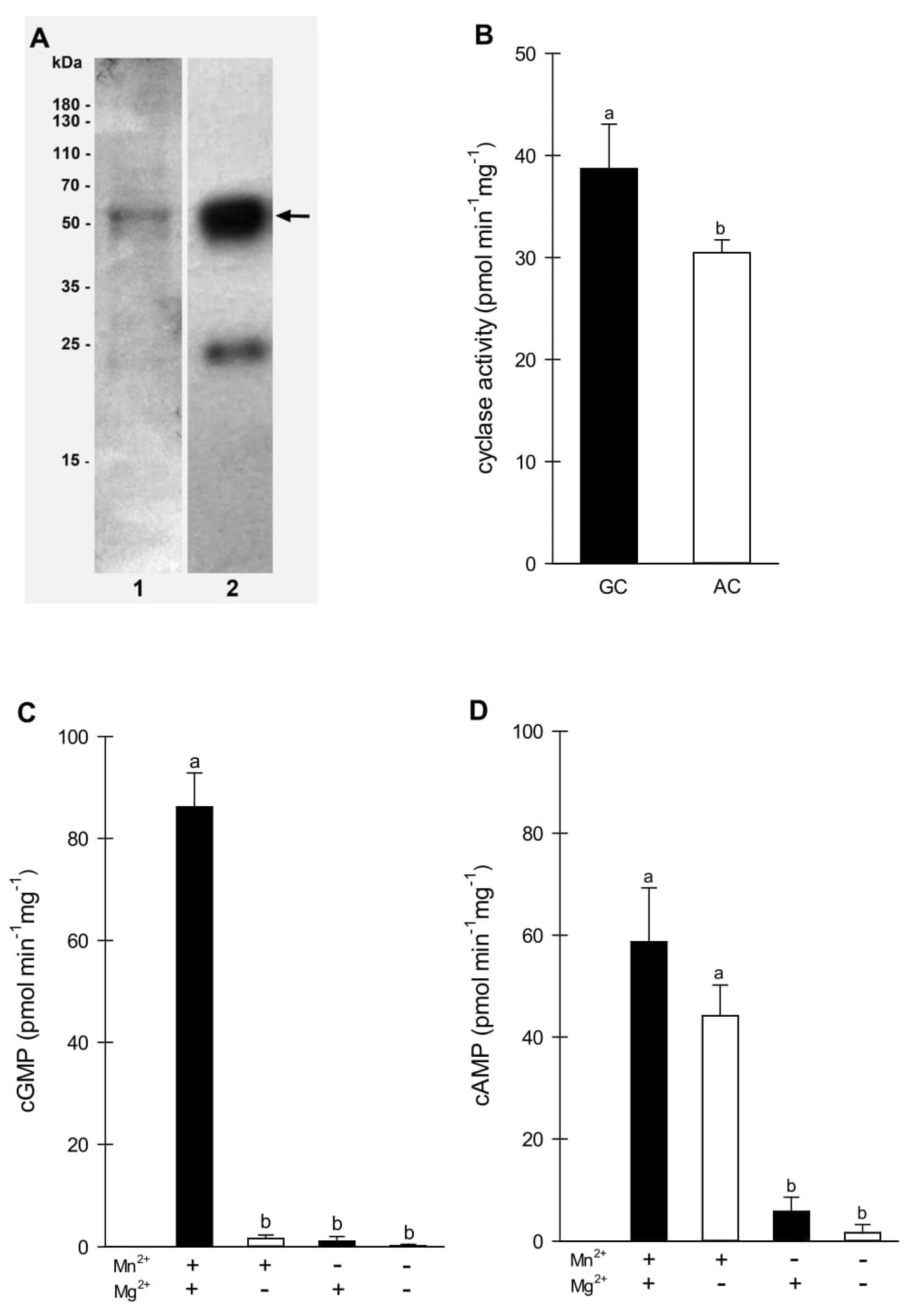
2.1. Sequence Analysis of BdGUCD1
2.2. In Vitro Analysis of the AC and GC Activity of BdGUCD1
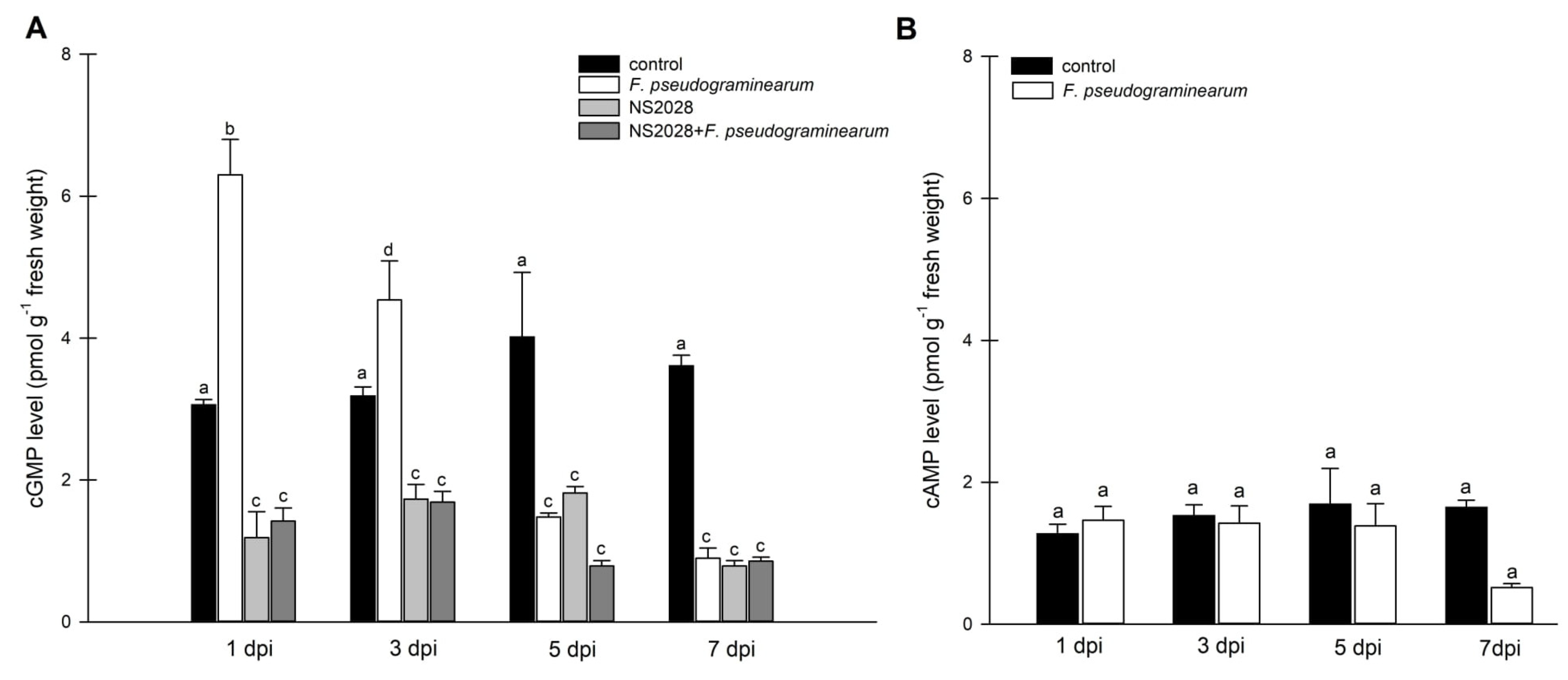
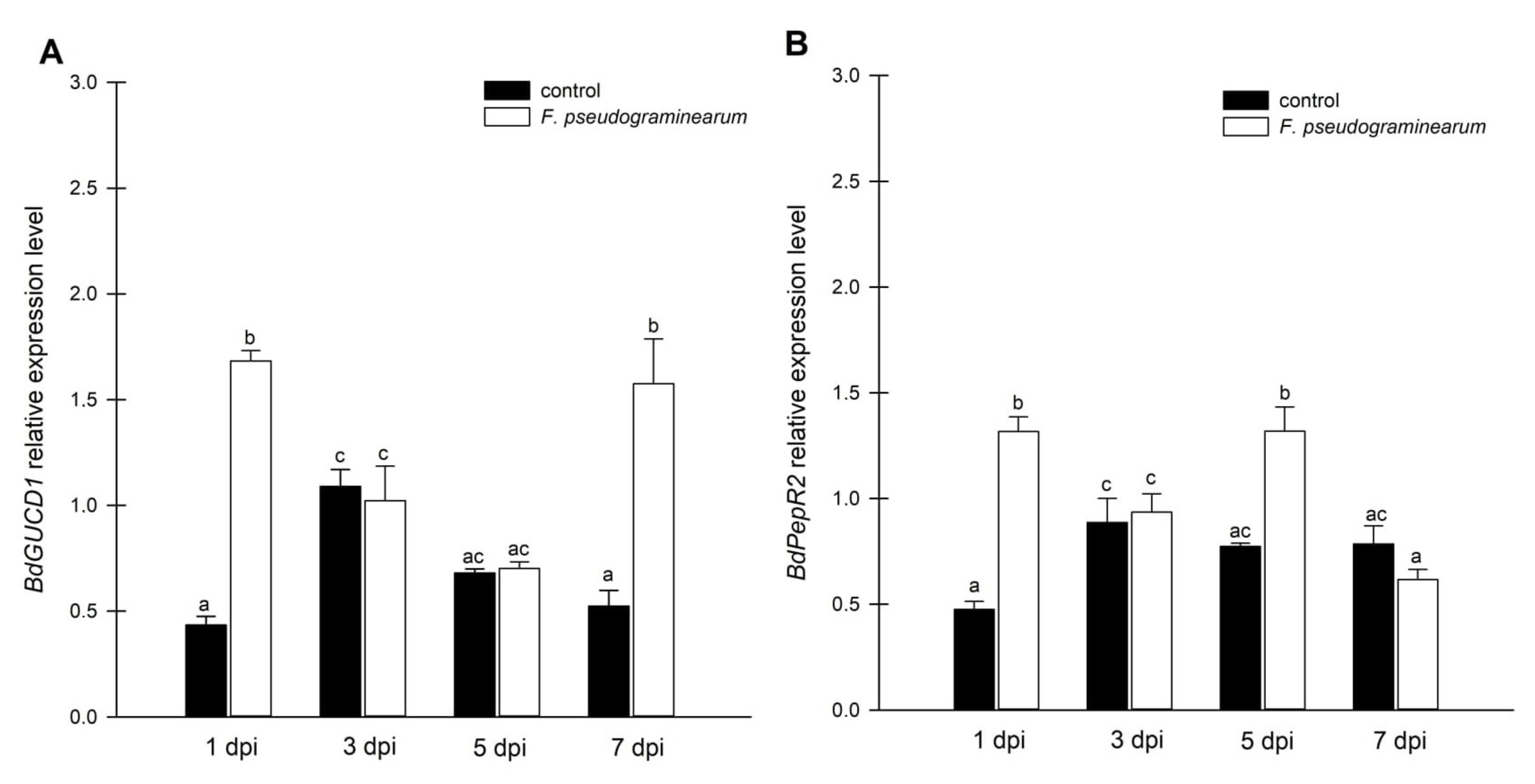
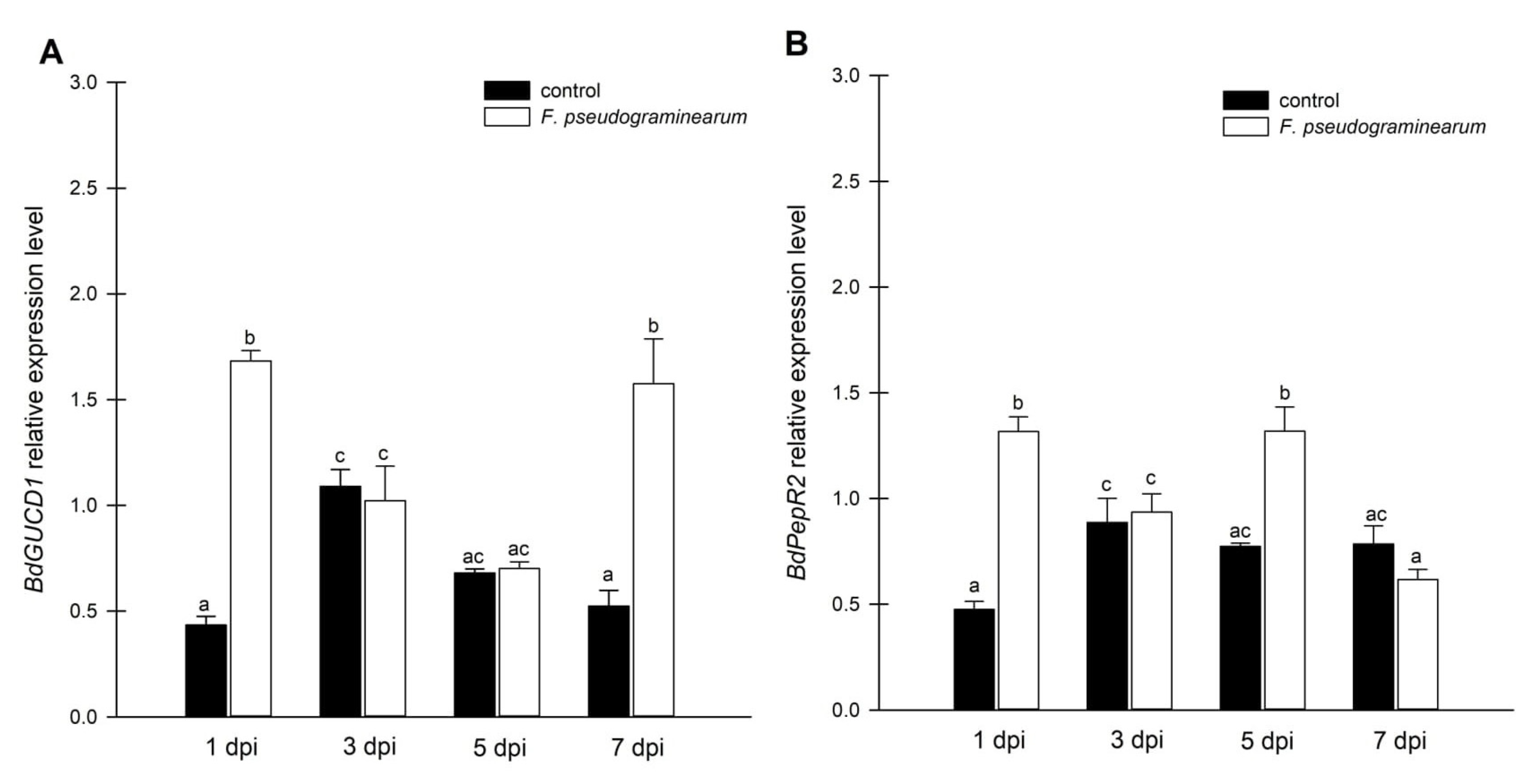
2.3. Effect of F. pseudograminearum Treatment on BdGUCD1 and BdPepR2 Expression and Endogenous Levels of Cyclic Nucleotides
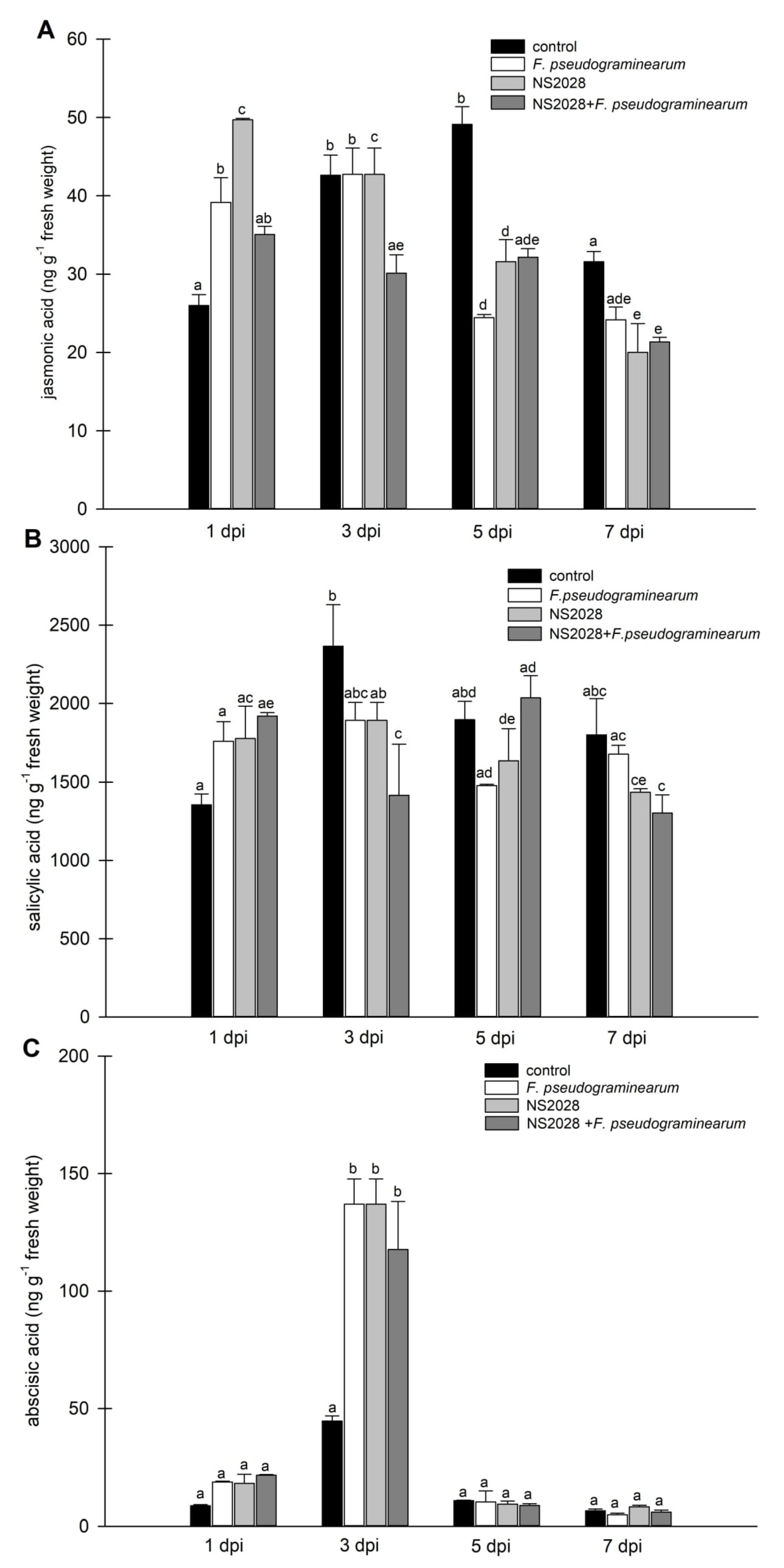
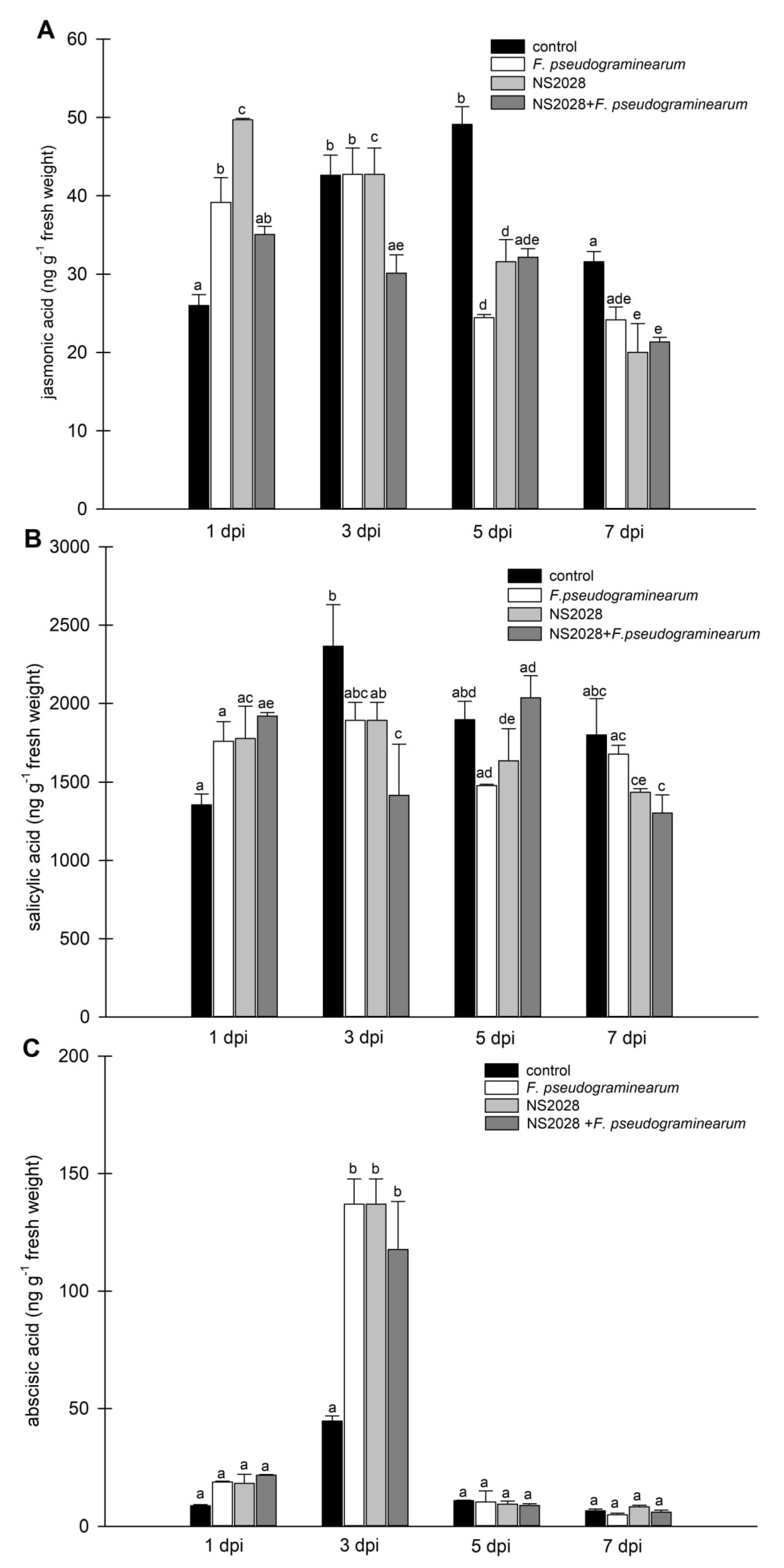
2.4. Effect of F. pseudograminearum Treatment on Phytohormone Levels
2.5. Impact of cGMP on Phytohormone Levels during Infection
3. Materials and Methods
3.1. Construction of the Expression Vector, Expression and Purification of Recombinant Protein
3.2. Determination of Guanylyl and Adenylyl Cyclase Activity
3.3. Plant Material and Pathogen Inoculation
3.4. Determination of Endogenous Levels of Phytohormones
3.5. Determination of Endogenous Levels of Cyclic Nucleotides
3.6. Gene Expression Analysis
3.7. Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Pauline, S. Guanylyl cyclases across the tree of life. Front. Biosci. 2005, 10, 1485–1498. [Google Scholar] [CrossRef] [Green Version]
- Świeżawska, B.; Duszyn, M.; Jaworski, K.; Szmidt-Jaworska, A. Downstream Targets of Cyclic Nucleotides in Plants. Front. Plant Sci. 2018, 9, 1428. [Google Scholar] [CrossRef] [Green Version]
- Kwezi, L.; Meier, S.; Mungur, L.; Ruzvidzo, O.; Irving, H.; Gehring, C. The Arabidopsis thaliana Brassinosteroid Receptor (AtBRI1) Contains a Domain that Functions as a Guanylyl Cyclase In Vitro. PLoS ONE 2007, 2, e449. [Google Scholar] [CrossRef] [Green Version]
- Świeżawska, B.; Jaworski, K.; Duszyn, M.; Pawełek, A.; Szmidt-Jaworska, A. The Hippeastrum hybridum PepR1 gene (HpPepR1) encodes a functional guanylyl cyclase and is involved in early response to fungal infection. J. Plant Physiol. 2017, 216, 100–107. [Google Scholar] [CrossRef]
- Świeżawska, B.; Jaworski, K.; Szewczuk, P.; Pawełek, A.; Szmidt-Jaworska, A. Identification of a Hippeastrum hybridum guanylyl cyclase responsive to wounding and pathogen infection. J. Plant Physiol. 2015, 189, 77–86. [Google Scholar] [CrossRef]
- Qi, Z.; Verma, R.; Gehring, C.; Yamaguchi, Y.; Zhao, Y.; Ryan, C.A.; Berkowitz, G.A. Ca2+ signaling by plant Arabidopsis thaliana Pep peptides depends on AtPepR1, a receptor with guanylyl cyclase activity, and cGMP-activated Ca2+ channels. Proc. Natl. Acad. Sci. USA 2010, 107, 21193–21198. [Google Scholar] [CrossRef] [Green Version]
- Malukani, K.K.; Ranjan, A.; Hota, S.J.; Patel, H.K.; Sonti, R.V. Dual Activities of Receptor-Like Kinase OsWAKL21.2 Induce Immune Responses. Plant Physiol. 2020, 183, 1345–1363. [Google Scholar] [CrossRef]
- Gehring, C.A.; Turek, I. Cyclic Nucleotide Monophosphates and Their Cyclases in Plant Signaling. Front. Plant Sci. 2017, 8, 1704. [Google Scholar] [CrossRef] [Green Version]
- Isner, J.-C.; Nühse, T.; Maathuis, F.J.M. The cyclic nucleotide cGMP is involved in plant hormone signalling and alters phosphorylation of Arabidopsis thaliana root proteins. J. Exp. Bot. 2012, 63, 3199–3205. [Google Scholar] [CrossRef] [Green Version]
- Miras-Moreno, B.; Zhang, L.; Senizza, B.; Lucini, L. A metabolomics insight into the Cyclic Nucleotide Monophosphate signaling cascade in tomato under non-stress and salinity conditions. Plant Sci. 2021, 309, 110955. [Google Scholar] [CrossRef]
- Dubovskaya, L.V.; Bakakina, Y.S.; Kolesneva, E.V.; Sodel, D.L.; McAinsh, M.R.; Hetherington, A.M.; Volotovski, I.D. cGMP-dependent ABA-induced stomatal closure in the ABA-insensitive Arabidopsis mutantabi1-1. New Phytol. 2011, 191, 57–69. [Google Scholar] [CrossRef]
- Shen, Q.; Zhan, X.; Yang, P.; Li, J.; Chen, J.; Tang, B.; Wang, X.; Hong, Y. Dual Activities of Plant cGMP-Dependent Protein Kinase and Its Roles in Gibberellin Signaling and Salt Stress. Plant Cell 2019, 31, 3073–3091. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Jia, H.; Wang, J. cGMP and ethylene are involved in maintaining ion homeostasis under salt stress in Arabidopsis roots. Plant Cell Rep. 2014, 33, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Meier, S.; Ruzvidzo, O.; Morse, M.; Donaldson, L.; Kwezi, L.; Gehring, C.A. The Arabidopsis Wall Associated Kinase-Like 10 Gene Encodes a Functional Guanylyl Cyclase and Is Co-Expressed with Pathogen Defense Related Genes. PLoS ONE 2010, 5, e8904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Silva, M.N.; Carvalho, S.M.P.; Rodrigues, A.M.; Gómez-Cadenas, A.; António, C.; Vasconcelos, M.W. Defence-related pathways, phytohormones and primary metabolism are key players in kiwifruit plant tolerance to Pseudomonas syringae pv. actinidiae. Plant Cell Environ. 2021, 45, 528–541. [Google Scholar] [CrossRef]
- Buhrow, L.M.; Liu, Z.; Cram, D.; Sharma, T.; Foroud, N.A.; Pan, Y.; Loewen, M.C. Wheat transcriptome profiling reveals abscisic and gibberellic acid treatments regulate early-stage phytohormone defense signaling, cell wall fortification, and metabolic switches following Fusarium graminearum-challenge. BMC Genom. 2021, 22, 798. [Google Scholar] [CrossRef]
- Hemelíková, N.; Žukauskaitė, A.; Pospíšil, T.; Strnad, M.; Doležal, K.; Mik, V. Caged Phytohormones: From Chemical Inactivation to Controlled Physiological Response. J. Agric. Food Chem. 2021, 69, 12111–12125. [Google Scholar] [CrossRef]
- Ngou, B.P.M.; Jones, J.D.; Ding, P. Plant immune networks. Trends Plant Sci. 2021, 27, 255–273. [Google Scholar] [CrossRef]
- Kazan, K.; Gardiner, D.M. Fusarium crown rot caused byFusarium pseudograminearumin cereal crops: Recent progress and future prospects. Mol. Plant Pathol. 2018, 19, 1547–1562. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Ali, M.L.; Taylor, J.; Liu, J.; Sun, G.; Liu, W.; Masilimany, P.; Gulati-Sakhuja, A.; Pauls, K.P. A guanylyl cyclase-like gene is associated with Gibberella ear rot resistance in maize (Zea mays L.). Theor. Appl. Genet. 2007, 116, 465–479. [Google Scholar] [CrossRef]
- Ludidi, N.; Gehring, C. Identification of a Novel Protein with Guanylyl Cyclase Activity in Arabidopsis thaliana. J. Biol. Chem. 2003, 278, 6490–6494. [Google Scholar] [CrossRef] [Green Version]
- Szmidt-Jaworska, A.; Jaworski, K.; Pawełek, A.; Kopcewicz, J. Molecular Cloning and Characterization of a Guanylyl Cyclase, PnGC-1, Involved in Light Signaling in Pharbitis nil. J. Plant Growth Regul. 2009, 28, 367–380. [Google Scholar] [CrossRef]
- Wong, A.; Gehring, C.; Irving, H.R. Conserved Functional Motifs and Homology Modeling to Predict Hidden Moonlighting Functional Sites. Front. Bioeng. Biotechnol. 2015, 3, 82. [Google Scholar] [CrossRef] [Green Version]
- Wong, A.; Tian, X.; Gehring, C.; Marondedze, C. Discovery of Novel Functional Centers with Rationally Designed Amino Acid Motifs. Comput. Struct. Biotechnol. J. 2018, 16, 70–76. [Google Scholar] [CrossRef]
- Duszyn, M.; Świeżawska-Boniecka, B.; Wong, A.; Jaworski, K.; Szmidt-Jaworska, A. In Vitro Characterization of Guanylyl Cyclase BdPepR2 from Brachypodium distachyon Identified through a Motif-Based Approach. Int. J. Mol. Sci. 2021, 22, 6243. [Google Scholar] [CrossRef] [PubMed]
- Świeżawska-Boniecka, B.; Duszyn, M.; Hammer, K.; Wong, A.; Szmidt-Jaworska, A.; Jaworski, K. Brachypodium distachyon ERECTA-like1 protein kinase is a functional guanylyl cyclase. Front. Biosci. 2021, 13, 249. [Google Scholar] [CrossRef]
- Rahman, H.; Wang, X.-Y.; Xu, Y.-P.; He, Y.-H.; Cai, X.-Z. Characterization of tomato protein kinases embedding guanylate cyclase catalytic center motif. Sci. Rep. 2020, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Meier, S.; Madeo, L.; Ederli, L.; Donaldson, L.; Pasqualini, S.; Gehring, C. Deciphering cGMP signatures and cGMP-dependent pathways in plant defence. Plant Signal. Behav. 2009, 4, 307–309. [Google Scholar] [CrossRef] [Green Version]
- Saijo, Y.; Loo, E.; Yasuda, S. Pattern recognition receptors and signaling in plant-microbe interactions. Plant J. 2018, 93, 592–613. [Google Scholar] [CrossRef]
- Hussain, J.; Chen, J.; Locato, V.; Sabetta, W.; Behera, S.; Cimini, S.; Griggio, F.; Martínez-Jaime, S.; Graf, A.; Bouneb, M.; et al. Constitutive cyclic GMP accumulation in Arabidopsis thaliana compromises systemic acquired resistance induced by an avirulent pathogen by modulating local signals. Sci. Rep. 2016, 6, 36423. [Google Scholar] [CrossRef]
- Donaldson, L.; Ludidi, N.; Knight, M.; Gehring, C.; Denby, K. Salt and osmotic stress cause rapid increases inArabidopsis thalianacGMP levels. FEBS Lett. 2004, 569, 317–320. [Google Scholar] [CrossRef] [Green Version]
- Van Loon, L.; Rep, M.; Pieterse, C. Significance of Inducible Defense-related Proteins in Infected Plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, H.; Kanayama, Y.; Zheng, M.S.; Kusano, T.; Hase, S.; Ikegami, M.; Shah, J. Antagonistic Interactions between the SA and JA Signaling Pathways in Arabidopsis Modulate Expression of Defense Genes and Gene-for-Gene Resistance to Cucumber Mosaic Virus. Plant Cell Physiol. 2004, 45, 803–809. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Zhou, S.; Yang, D.; Fan, Z. Revealing Shared and Distinct Genes Responding to JA and SA Signaling in Arabidopsis by Meta-Analysis. Front. Plant Sci. 2020, 11, 908. [Google Scholar] [CrossRef]
- Desmond, O.J.; Edgar, C.I.; Manners, J.M.; Maclean, D.J.; Schenk, P.; Kazan, K. Methyl jasmonate induced gene expression in wheat delays symptom development by the crown rot pathogen Fusarium pseudograminearum. Physiol. Mol. Plant Pathol. 2005, 67, 171–179. [Google Scholar] [CrossRef]
- Desmond, O.J.; Manners, J.M.; Schenk, P.M.; Maclean, D.J.; Kazan, K. Gene expression analysis of the wheat response to infection by Fusarium pseudograminearum. Physiol. Mol. Plant Pathol. 2008, 73, 40–47. [Google Scholar] [CrossRef]
- Powell, J.J.; Carere, J.; Sablok, G.; Fitzgerald, T.L.; Stiller, J.; Colgrave, M.L.; Gardiner, D.M.; Manners, J.M.; Vogel, J.P.; Henry, R.J.; et al. Transcriptome analysis of Brachypodium during fungal pathogen infection reveals both shared and distinct defense responses with wheat. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef]
- Powell, J.J.; Carere, J.; Fitzgerald, T.L.; Stiller, J.; Covarelli, L.; Xu, Q.; Gubler, F.; Colgrave, M.L.; Gardiner, D.M.; Manners, J.M.; et al. The Fusarium crown rot pathogenFusarium pseudograminearumtriggers a suite of transcriptional and metabolic changes in bread wheat (Triticum aestivum L.). Ann. Bot. 2016, 119, 853–867. [Google Scholar] [CrossRef] [Green Version]
- Ton, J.; Flors, V.; Mauch-Mani, B. The multifaceted role of ABA in disease resistance. Trends Plant Sci. 2009, 14, 310–317. [Google Scholar] [CrossRef]
- Maksimov, I.V. Abscisic acid in the plants-pathogen interaction. Russ. J. Plant Physiol. 2009, 56, 742–752. [Google Scholar] [CrossRef]
- Boba, A.; Kostyn, K.; Kozak, B.; Wojtasik, W.; Preisner, M.; Prescha, A.; Gola, E.M.; Lysh, D.; Dudek, B.; Szopa, J.; et al. Fusarium oxysporum infection activates the plastidial branch of the terpenoid biosynthesis pathway in flax, leading to increased ABA synthesis. Planta 2020, 251, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Buhrow, L.M.; Cram, D.; Tulpan, D.; Foroud, N.A.; Loewen, M.C. Exogenous Abscisic Acid and Gibberellic Acid Elicit Opposing Effects on Fusarium graminearum Infection in Wheat. Phytopathology 2016, 106, 986–996. [Google Scholar] [CrossRef] [Green Version]
- Penson, S.P.; Schuurink, R.C.; Fath, A.; Gubler, F.; Jacobsen, J.V.; Jones, R.L. cGMP Is Required for Gibberellic Acid-Induced Gene Expression in Barley Aleurone. Plant Cell 1996, 8, 2325–2333. [Google Scholar] [CrossRef]
- Wang, Y.H.; Gehring, C.; Cahill, D.M.; Irving, H.R.; Gehring, C. Plant natriuretic peptide active site determination and effects on cGMP and cell volume regulation. Funct. Plant Biol. 2007, 34, 645–653. [Google Scholar] [CrossRef]
- Neill, S.J.; Desikan, R.; Clarke, A.; Hancock, J.T. Nitric Oxide Is a Novel Component of Abscisic Acid Signaling in Stomatal Guard Cells. Plant Physiol. 2002, 128, 13–16. [Google Scholar] [CrossRef]
- Jiao, C.; Gu, Z. Cyclic GMP mediates abscisic acid-stimulated isoflavone synthesis in soybean sprouts. Food Chem. 2019, 275, 439–445. [Google Scholar] [CrossRef]
- Isner, J.-C.; Maathuis, F.J.M. cGMP signalling in plants: From enigma to main stream. Funct. Plant Biol. 2018, 45, 93–101. [Google Scholar] [CrossRef]
- Zhao, Y.; Qi, Z.; Berkowitz, G.A. Teaching an Old Hormone New Tricks: Cytosolic Ca2+ Elevation Involvement in Plant Brassinosteroid Signal Transduction Cascades. Plant Physiol. 2013, 163, 555–565. [Google Scholar] [CrossRef] [Green Version]
- Pagnussat, G.C.; Lanteri, M.L.; Lamattina, L. Nitric Oxide and Cyclic GMP Are Messengers in the Indole Acetic Acid-Induced Adventitious Rooting Process. Plant Physiol. 2003, 132, 1241–1248. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Fan, L.W.; Wu, W.H. Evidences for involvement of endogenous cAMP in Arabidopsis defense responses to Verticillium toxins. Cell Res. 2005, 15, 585–592. [Google Scholar] [CrossRef]
- Lu, M.; Zhang, Y.; Tang, S.; Pan, J.; Yu, Y.; Han, J.; Li, Y.; Du, X.; Nan, Z.; Sun, Q. AtCNGC2 is involved in jasmonic acid-induced calcium mobilization. J. Exp. Bot. 2016, 67, 809–819. [Google Scholar] [CrossRef] [Green Version]
- Świeżawska, B.; Duszyn, M.; Kwiatkowski, M.; Jaworski, K.; Pawełek, A.; Szmidt-Jaworska, A. Brachypodium distachyon triphosphate tunnel metalloenzyme 3 is both a triphosphatase and an adenylyl cyclase upregulated by mechanical wounding. FEBS Lett. 2020, 594, 1101–1111. [Google Scholar] [CrossRef]
- Pu, C.-H.; Lin, S.-K.; Chuang, W.-C.; Shyu, T.-H. Modified QuEChERS method for 24 plant growth regulators in grapes using LC-MS/MS. J. Food Drug Anal. 2018, 26, 637–648. [Google Scholar] [CrossRef] [Green Version]
- Van Damme, T.; Blancquaert, D.; Couturon, P.; Van Der Straeten, D.; Sandra, P.; Lynen, F. Wounding stress causes rapid increase in concentration of the naturally occurring 2′,3′-isomers of cyclic guanosine- and cyclic adenosine monophosphate (cGMP and cAMP) in plant tissues. Phytochemistry 2014, 103, 59–66. [Google Scholar] [CrossRef]
- Pfaffl, M.W. Relative expression software tool (REST(C)) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, 36e. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duszyn, M.; Świeżawska-Boniecka, B.; Skorupa, M.; Jaworski, K.; Szmidt-Jaworska, A. BdGUCD1 and Cyclic GMP Are Required for Responses of Brachypodium distachyon to Fusarium pseudograminearum in the Mechanism Involving Jasmonate. Int. J. Mol. Sci. 2022, 23, 2674. https://doi.org/10.3390/ijms23052674
Duszyn M, Świeżawska-Boniecka B, Skorupa M, Jaworski K, Szmidt-Jaworska A. BdGUCD1 and Cyclic GMP Are Required for Responses of Brachypodium distachyon to Fusarium pseudograminearum in the Mechanism Involving Jasmonate. International Journal of Molecular Sciences. 2022; 23(5):2674. https://doi.org/10.3390/ijms23052674
Chicago/Turabian StyleDuszyn, Maria, Brygida Świeżawska-Boniecka, Monika Skorupa, Krzysztof Jaworski, and Adriana Szmidt-Jaworska. 2022. "BdGUCD1 and Cyclic GMP Are Required for Responses of Brachypodium distachyon to Fusarium pseudograminearum in the Mechanism Involving Jasmonate" International Journal of Molecular Sciences 23, no. 5: 2674. https://doi.org/10.3390/ijms23052674
APA StyleDuszyn, M., Świeżawska-Boniecka, B., Skorupa, M., Jaworski, K., & Szmidt-Jaworska, A. (2022). BdGUCD1 and Cyclic GMP Are Required for Responses of Brachypodium distachyon to Fusarium pseudograminearum in the Mechanism Involving Jasmonate. International Journal of Molecular Sciences, 23(5), 2674. https://doi.org/10.3390/ijms23052674