
Mast Cells in the Mammalian Testis and Epididymis—Animal Models and Detection Methods

 , , and
, , and
Abstract
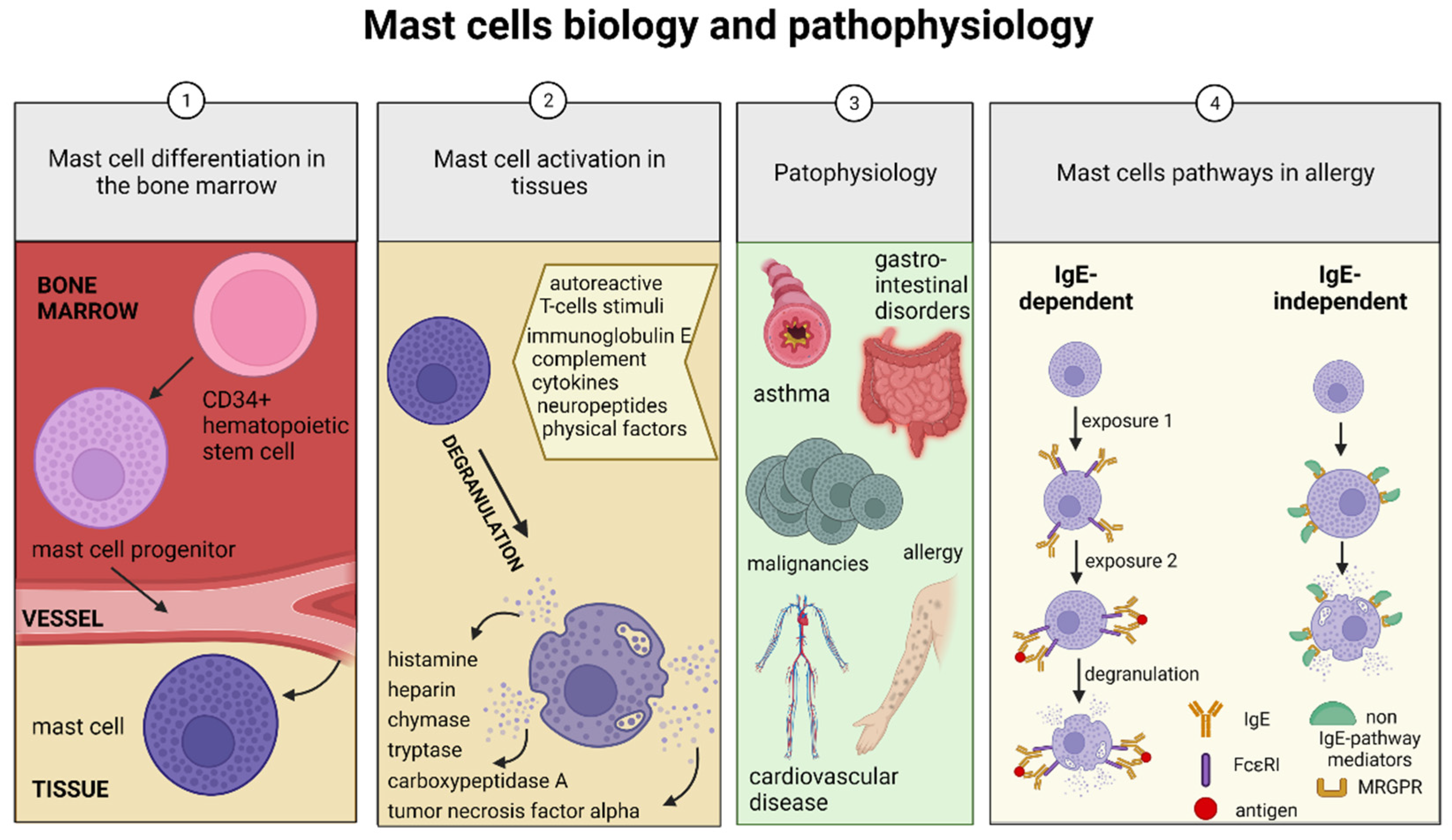
1. Introduction
2. Mast Cell Disorders
2.1. Allergies
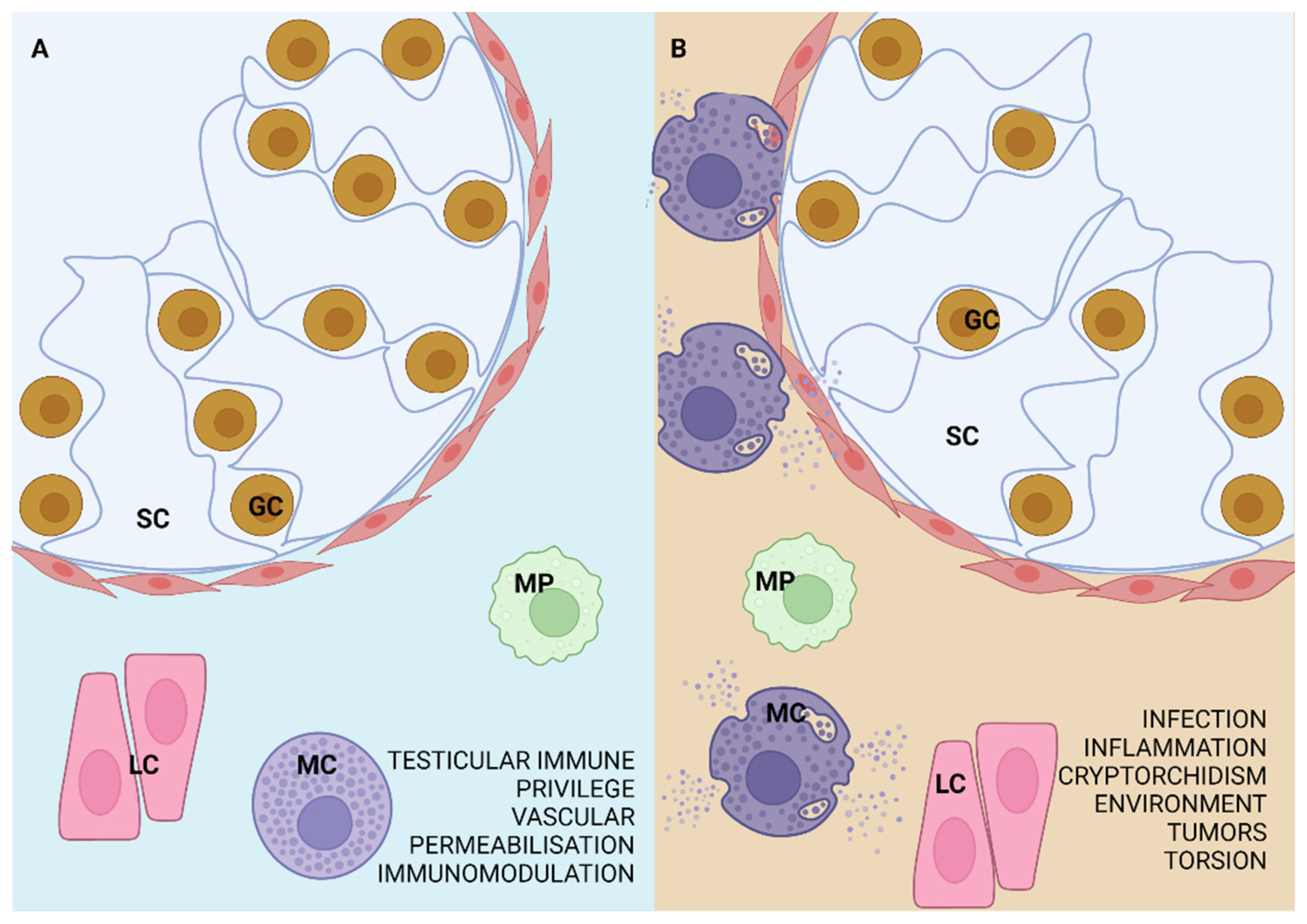
2.2. Contribution of Mast Cells to the Pathology of the Mammalian Testis and Epididymis
3. Evolutionarily Conserved Mast Cells
4. Mast Cell Detection Methods in the Mammalian Testis and Epididymis
5. Mast Cells in Mammalian Testes and Epididymides
5.1. Rodents (Rat, Mouse, Hamster, Other)
5.2. Domestic (Sus scrofa domestica) and Wild Boar (Sus scrofa ferrus)
5.3. Non-Human Primates
6. Experimental Models Investigating Mast Cells in Mammalian Testes
6.1. Gonadal Effects of Medications
6.2. Mast Cell Antagonists
6.3. Experimental Autoimmune Orchitis (EAO)
6.4. Stress
6.5. Hormones
6.6. Genetically Altered Animals
6.7. Cryptorchidism

{kind=link}
{kind=link}
{kind=link}
| Animal | Strain | Experiment | Analysis | Result—MC | Result—ST | Result—Interstitium | Fertility * | Ref. |
|---|---|---|---|---|---|---|---|---|
| ALKYLATING AGENTS | ||||||||
| RAT | Wistar | EDS daily PND 5–16 | PND 6–108 | MC in interstitium at PND 17–35 and increased number of MCs under the TA PND 17–108 | valuolae in SCs, thick basal lamina from PND 11; atrophic ST with no GCs or lumen from PND 28; no recovery after treatment | no LC from PND 11 | NA | [69] |
| Sprague-Dawley | EDS single dose | 0–49 days after treatment | MCs in interstitium, max. 21 days after EDS | ST volume decrease: lowest 14 days after treatment; control levels 35 days after treatment | no LC on day 3 after EDS, recovered 50% by day 28 after EDS | NA | [65] | |
| Sprague-Dawley | 1. EDS treatment ± testosterone implant 2. immunization against oxytocin | 3–10 weeks after treatment | 1. MCs in interstitium 21 and 70 days after EDS treatment 2. no MCs in interstitium after oxytocin administration | 1. 3–4× increased GC degeneration 2. 2× increased GC degeneration | 1. LC depletion | 1. transient subfertility in 50% | [66] | |
| Sprague-Dawley | EDS ± testosterone | 3–41 days after treatment | MC increase in interstitium 21–41 days post-treatment (EDS); slower increase with testosterone | NA | LC depletion, initial macrophage increase (3–10 days), then depletion, prevented by testosterone | NA | [53] | |
| Wistar | 1. hypophysectomy ± EDS (adult) OR GnRH antagonist (prepubertal); colchicine 2. GnRH antagonist | 1–30 days after treatment | 1. MC proliferation on day 20 by EDS, MC number increase 15–30 day (control) and 15–50 day in hypophysectomiy + EDS 2. prolif. and diff. MC PND 23–30 | NA | 1. mitosis-3rd and 18–22nd day 2. immature LC and mitotic figures increase, normal by 30th day | NA | [61] | |
| Sprague Dawley | cyclophosphamide ± ZnO NP | after 4 weeks of treatment | MC number increase in interstitium; ZnO NP reduced (near normal MC number) | ZnO NP reduced ST and GC atrophy, separation of germinal epithelium from BM, tubular wall vacuolization, sperm abnormalities | wide interstitium, vascular congestion, acidophilic material, clusters of dark LC; ZnO NP reduced | NA | [57] | |
| MOUSE | NMRI | cyclophosphamide ± EP for 35 days | after treatment | MC number increase in interstitium; EP reduced | germinal epithelium disarrangement and thickening; EP reduces | LC depletion; EP reduced | NA | [84] |
| INFLAMMATION | ||||||||
| RAT | Wistar and Sprague-Dawley | EAO (3 injections every 15 days) | 7–80 days after 1st injection | MC degranulation, number elevation after 80 days, 2× Sprague-Dawley and 5× Wistar | aspermatogenesis at day 80 | mononuclear cell infiltrate at day 80 | NA | [67] |
| Wistar | EAO | 50 & 80 days after EAO | MC number and degranulation increase; MC in interstitium | almost complete loss of GC | granuloma formation; PAR2+ cell number increase | NA | [47] | |
| Wistar | testis puncture Ø 3 mm (=EAO) followed by skin suture ± ketotifen | 1–30 days after treatment | ketotifen reduced MC number and degranulation increase in interstitium | GC depletion and ST fibrosis; ketotifen (does not prevent damage but) retains regenerative capacity | NA | [74] | ||
| Wistar | EAO ± ketotifen | 56 days after first treatment | ketotifen reduced MC number increase | ketotifen reduced severity of histopathological testicular damage | ketotifen reduced testicular damage | NA | [117] | |
| MOUSE | C57BL6/N or C57BL/6J | EAEO | 30–80 days after 1st immunisation | MC number increase in interstitium from low to high-grade EAEO | severe EAEO: ST diameter reduction, germ cell sloughing, SC-only tubules | macrophage and leukocyte number increase, fibrosis around ST | NA | [77] |
| TORSION AND STRESS | ||||||||
| RAT | Sprague–Dawley | torsion ± ketotifen | 30 days after treatment | ketotifen reduced increased MC numbers—contralateral testis | ketotifen reduced germinal epithelium sloughing, ST atrophy, walls fibrosis—contralateral testis | interstitial edema | NA | [117] |
| Sprague-Dawley | torsion-detorsion ± VIP (2 or 4 h); prepubertal | after torsion | MC degranulation; VIP—protective effect (2 h); no MCs in interstitium | histological abnormalities; VIP reduced (2 h torsion) | histological abnormalities; VIP reduced (2 h torsion) | NA | [48] | |
| Sprague-Dawley | torsion; contralateral testis analysis | 10, 30, and 80 days after torsion | maximum increase in MC number 30 days post-op | germinal epithelium sloughing (spermatocyte and spermatid apoptosis), ST atrophy, fibrosis, diameter, and Johnsen score decrease; possible reversibility | interstitial edema, T-lymphocyte, and macrophage number elevation | NA | [64] | |
| MOUSE | dYY | torsion; contralateral testis analysis | 4–24 weeks after torsion | MC number increase in interstitium, maximum 8 weeks post-op | no histological changes | no histological changes | NA | [75] |
| RAT | Wistar | 1. immobilisation cold stress 3h 3 days PND 15, 30, 45 ± VIP 2. β-endorphin + immobilisation PND 45 ± VIP | after treatment | 1. VIP reduced MC degranulation and maturation 2. VIP reduced MC degranulation and maturation | NA | 1. VIP prevented focal LC depletion | NA | [72] |
| Wistar | torsion/detorsion ± hypothermia for 30 or 90′ prior to detorsion | 8 weeks after operation | torsion/detorsion significantly increased MC number | hypothermia increased Johansen score, reduced by torsion/detorsion | hypothermia ameliorated interstitial edema | NA | [120] | |
| HORMONE TREATMENT | ||||||||
| RAT | Wistar | EB on PND 1 | PND 45 and 90 | strong increase of MC number, interstitium at PND 45 | maturation arrest at the level of pachytene spermatocytes | increased proportion of interstitium (fibrosis and edema) | NA | [44] |
| Wistar | 1. EB on PND 1 2. testosterone propionate on PND 1 | PND 45 | 1. MC number increase after estrogen in the testis 2. no effect | 1. impaired spermatogenesis | 1. immature interstitial cells | NA | [70] | |
| Wistar | EB on PND 1 | PND 15–90 | MC in interstitium on PND 17, increase in number by PND 45 and mature by PND 90 | NA | mature LC from PND 90 | NA | [45] | |
| Wistar | 1. EB treatment on PND 1 2. EDS (adult) | 1. PND 35–70 2. 5 days after treatment | 1. strong increase of MC number, interstitium at PND 35–70 testicular serotonin increase 2. no effect on MCs | NA | 1. LC depletion on PND 35; normal number on PND 70 2. LC absent | NA | [46] | |
| ETHANOL | ||||||||
| RAT | Wistar | ethanol and ethanol extract of Bauhinia forficata | 31 days after 1st treatment | decreased only the number of degranulated MCs | no effect | NA | NA | [68] |
| UchB (Wistar) | ethanol 100 days | after treatment | no difference in MC number (testis); increase in total number of degranulated MCs (epidydimis) | NA | NA | NA | [49] | |
| Wistar | ethanol 54 days | after treatment | no difference (testis); increase of MC number and degranulation (cauda & initial segment) | ethanol: mature spermatid number, mobile sperm count reduction; abnormal seminiferous tubule morphology | NA | NA | [51] | |
| GENETIC ALTERATIONS | ||||||||
| MOUSE | WT and transgenic AROM+ | AROM+ alteration | 4-, 9-, and 15- month-old | MC number increase in interstitium and during aging | spermatogenic disruption progression during aging (GC depletion to absence at 15 months) | LC hyperplasia and hypertrophy (4 months), giant multinucleated macrophage number progression; LC adenomas (9- and 15-month-old) | NA | [76] |
| Mt-hAMH and C57BL/6 | Mt-hAMH alteration | 5-month-old | abundant MC in the interstitium | lower length of ST, vacuolization of Sertoli cells, loss of GC | LC depletion | infertility after 3 rounds of consecutive pairing | [125] | |
| WT (C57BL/6) and RORα-deficient | RORα-deficiency alteration | 10–12-week-old | MC in the interstitium and within the seminiferous tubule near BM | ST diameter and germinal epithelium height decrease (GC apoptosis), basal membrane irregularities, hypospermatogenesis | LC vacuolization number reduction | NA | [78] | |
| INFECTION | ||||||||
| RAT | Wistar | C. trahomatis inoculation | 3–90 days after infection | MCs in inflammatory lesions of the epididymis 30 days after | germinal epithelium loss, spermatid giant cells after 7–70 days | interstitial fibrosis after 7–70 days | NA | [128] |
| DEER MOUSE | Trypanosoma brucei infection | 0–10 weeks after infection | MC number increase in the interstitium | ST diameter decrease (loss of GC), increase with time | LC accumulation, mononuclear cell infiltration, increase with time | NA | [129] | |
| CRYPTORCHIDISM | ||||||||
| RAT | Wistar | experimental UDT (newborn) ± ketotifen, then peripubertal, pubertal, or adult bilateral orchidectomy | after orchidectomy | experimental UDT increased, and ketotifen decreased MC number in scrotal and abdominal testis | UDT: ST diameters decreased, ST basement membranes thickened, and spermatogenesis decreased—both testes; ketotifen prevented | interstitial fibrosis; ketotifen prevented | NA | [73] |
| Sprague-Dawley | PND 15–17 unilateral cryptorchidism | 15 days after treatment | MC number increase in cryptorchid (higher) and scrotal (lower) testis | ST atrophy, diameter reduction, degenerative changes, GC disconnection from BM, BM thickening, destruction in tight junctions between SCs, SCs, and spermatogenic cells, decomposition of cytoplasmic bridges between spermatogenic cells | perivascular and interstitial fibrosis, edema, congestion, hemorrhage | NA | [127] | |
| BOAR | domestic | cryptorchidism (unilateral and bilateral) | 9-month-old | abundant MCs in bilateral cryptorchid testes interstitium | NA | unilateral: fibrosis and LC degeneration in abdominal testis bilateral: advanced fibrosis, immature LC and LC degeneration | NA | [87] |
| OTHER | ||||||||
| RAT | Sprague-Dawley | topical histamine, ritanserin, ketanserin, histamine, and substance 48/80 | 30 min after treatment | MC degranulation after histamine and substance 48/80 (dose-dependant) | NA | NA | NA | [92] |
| Wistar | acyclovir i.p. 15 consecutive days (3 doses) | 18 days after last treatment | increase of MC number in the testicular interstitium and peritubular area with higher doses | ST diameter, epithelial height (cell loss), tubular differentiation index, spermiogenesis index, repopulation index (higher dose) decrease | LC atrophy, connective tissue increase | lower pregnancy rate | [50] | |
| Wistar | 1. Cl2MDP-lp injection PND 5, 10, 15, 20, 25 2. Cl2MDP ± hCG & hFSH PND 18 for 6 days | 1. PND 10–35 2. PND 27 | 1. MC in interstitium 10 and 15 days after treatment | NA | 1. short-term LC depletion after treatment PND 5–15 and long term LC depletion after treatment on PND 20–25 2. LC depletion, no changes after hCG and hFSH treatment | NA | [71] | |
| Wistar | males with no mating experience and colony breeders analyzed at PND 40–170 | PND 40, 60, 90, 120 | serotonin + MC increase PND 40- 90; MC number at peak on PND 90 | NA | NA | NA | [97] | |
| Sprague-Dawley | genistein ± DEHP in utero | PND 60, 120, 180 | genistein + DEHP: MC marker increase on PND 120 | genistein + DEHP: disruption in Sertoli cell function, different stage spermatogonia change | DEHP: LC number decrease PND 120 | NA | [56] | |
7. Experimental Models Investigating Mast Cells in Mammalian Epididymides
8. Discussion
9. Conclusions
- Histological assessment, including toluidine blue stain, should always be included in studies analyzing testicular and epididymal mast cells, as a standard method that stains all mast cell subtypes, regardless of protein content.
- Depending on the effect and antibody used, a few fixation methods should be optimized at the beginning of the study due to the mast cell subtype specificity.
- When investigating animal models, whole testes should be used, including the tunica albuginea, for in most mammals, mast cells reside right underneath. When found in the interstitium, the seminiferous tubule wall thickness should be commented upon.
- With respect to practicality, domestic boars kept under non-variable conditions are proposed to resemble human testicular mast cell distribution better than rodents.
- No exclusively testis- or epididymis-specific mast cell markers have been found yet, although the characterization of other organ-specific mast cell markers is known.
- 3D in vitro models are promising, although they still need significant development in order to incorporate mast cells and the tunica albuginea, if possible. Further efforts need to be made to develop a suitable human-origin testicular cell line combination.
10. Materials and Methods
10.1. Samples
10.2. Immunohistochemistry
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Benoist, C.; Mathis, D. Mast cells in autoimmune disease. Nature 2002, 420, 875–878. [Google Scholar] [CrossRef]
- Eurell, J.A.C.; Frappier, B.L.; Dellmann, H.D. Dellmann’s Textbook of Veterinary Histology; with CD; John Wiley & Sons: Hoboken, NJ, USA, 2006. [Google Scholar]
- Zhang, Z.; Kurashima, Y. Two Sides of the Coin: Mast Cells as a Key Regulator of Allergy and Acute/Chronic Inflammation. Cells 2021, 10, 1615. [Google Scholar] [CrossRef]
- Gentek, R.; Ghigo, C.; Hoeffel, G.; Bulle, M.J.; Msallam, R.; Gautier, G.; Launay, P.; Chen, J.; Ginhoux, F.; Bajénoff, M. Hemogenic Endothelial Fate Mapping Reveals Dual Developmental Origin of Mast Cells. Immunity 2018, 48, 1160–1171.e5. [Google Scholar] [CrossRef]
- Akula, S.; Hellman, L.; Avilés, F.X.; Wernersson, S. Analysis of the mast cell expressed carboxypeptidase A3 and its structural and evolutionary relationship to other vertebrate carboxypeptidases. Dev. Comp. Immunol. 2022, 127, 104273. [Google Scholar] [CrossRef]
- Rodewald, H.R.; Dessing, M.; Dvorak, A.M.; Galli, S.J. Identification of a committed precursor for the mast cell lineage. Science 1996, 271, 818–822. [Google Scholar] [CrossRef]
- Iemura, A.; Tsai, M.; Ando, A.; Wershil, B.K.; Galli, S.J. The c-kit ligand, stem cell factor, promotes mast cell survival by suppressing apoptosis. Am. J. Pathol. 1994, 144, 321–328. [Google Scholar]
- Cruse, G.; Metcalfe, D.D.; Olivera, A. Functional deregulation of KIT: Link to mast cell proliferative diseases and other neoplasms. Immunol. Allergy Clin. N. Am. 2014, 34, 219–237. [Google Scholar] [CrossRef]
- Vajravelu, B.N.; Al-maqtari, T. c-Kit—The Novel Receptor: Physiological Roles, Downstream Signaling and Implications in Cancer. J. Clin. Med. Res. 2021, 2, 1–36. [Google Scholar] [CrossRef]
- Gurish, M.F.; Tao, H.; Abonia, J.P.; Arya, A.; Friend, D.S.; Parker, C.M.; Austen, K.F. Intestinal mast cell progenitors require CD49dbeta7 (alpha4beta7 integrin) for tissue-specific homing. J. Exp. Med. 2001, 194, 1243–1252. [Google Scholar] [CrossRef]
- Galli, S.J.; Hammel, I. Mast cell and basophil development. Curr. Opin. Hematol. 1994, 1, 33–39. [Google Scholar]
- Ribatti, D. The Staining of Mast Cells: A Historical Overview. Int. Arch. Allergy Immunol. 2018, 176, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Fong, M.; Crane, J.S. Histology, Mast Cells; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2021. [Google Scholar]
- Krystel-Whittemore, M.; Dileepan, K.N.; Wood, J.G. Mast Cell: A Multi-Functional Master Cell. Front. Immunol. 2016, 6, 620. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, S.P.; Drucker, I.; Reshef, T.; Kirshenbaum, A.S.; Metcalfe, D.D.; Mekori, Y.A. Activated T lymphocytes induce degranulation and cytokine production by human mast cells following cell-to-cell contact. J. Leukoc. Biol. 1998, 63, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Haidl, G.; Duan, Y.G.; Chen, S.J.; Kohn, F.M.; Schupe, H.C.; Allam, J.P. The role of mast cells in male infertility. Expert Rev. Clin. Immunol. 2011, 7, 627–634. [Google Scholar] [CrossRef]
- Reber, L.L.; Marichal, T.; Galli, S.J. New models for analyzing mast cell functions in vivo. Trends Immunol. 2012, 33, 613–625. [Google Scholar] [CrossRef]
- Welle, M. Development, significance, and heterogeneity of mast cells with particular regard to the mast cell-specific proteases chymase and tryptase. J. Leukoc. Biol. 1997, 61, 233–245. [Google Scholar] [CrossRef]
- Moon, T.C.; St Laurent, C.D.; Morris, K.E.; Marcet, C.; Yoshimura, T.; Sekar, Y.; Befus, A.D. Advances in mast cell biology: New understanding of heterogeneity and function. Mucosal Immunol. 2010, 3, 111–128. [Google Scholar] [CrossRef]
- Wilcock, A.; Bahri, R.; Bulfone-Paus, S.; Arkwright, P.D. Mast cell disorders: From infancy to maturity. Allergy 2019, 74, 53–63. [Google Scholar] [CrossRef]
- Gaudenzio, N.; Sibilano, R.; Starkl, P.; Tsai, M.; Galli, S.J.; Reber, L.L. Analyzing the Functions of Mast Cells In Vivo Using ‘Mast Cell Knock-in’ Mice. J. Vis. Exp. 2015, 99, e52753. [Google Scholar]
- Franke, K.; Wang, Z.; Zuberbier, T.; Babina, M. Cytokines Stimulated by IL-33 in Human Skin Mast Cells: Involvement of NF-κB and p38 at Distinct Levels and Potent Co-Operation with FcεRI and MRGPRX2. Int. J. Mol. Sci. 2021, 22, 3580. [Google Scholar] [CrossRef]
- Galli, S.J.; Tsai, M. IgE and mast cells in allergic disease. Nat. Med. 2012, 18, 693–704. [Google Scholar] [CrossRef] [PubMed]
- Nagata, K.; Nishiyama, C. IL-10 in Mast Cell-Mediated Immune Responses: Anti-Inflammatory and Proinflammatory Roles. Int. J. Mol. Sci. 2021, 22, 4972. [Google Scholar] [CrossRef] [PubMed]
- Quan, P.L.; Sabaté-Brescó, M.; Gou, Y.; Martín, M.; Gastaminza, G. The Multifaceted Mas-Related G Protein-Coupled Receptor Member X2 in Allergic Diseases and Beyond. Int. J. Mol. Sci. 2021, 22, 4421. [Google Scholar] [CrossRef] [PubMed]
- Navinés-Ferrer, A.; Serrano-Candelas, E.; Lafuente, A.; Muñoz-Cano, R.; Martín, M.; Gastaminza, G. MRGPRX2-mediated mast cell response to drugs used inperioperative procedures and anaesthesia. Sci. Rep. 2018, 8, 11628. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, Y.; Che, D.; Zeng, Y.; Wu, Y.; Qin, Q.; Wang, N. Baicalin induces Mrgprb2-dependent pseudo-allergy in mice. Immunol. Lett. 2020, 226, 55–61. [Google Scholar] [CrossRef]
- Ogasawara, H.; Noguchi, M. Therapeutic Potential of MRGPRX2 Inhibitors on Mast Cells. Cells 2021, 10, 2906. [Google Scholar] [CrossRef]
- Ježek, D.; Banek, L.; Dolovski, S.; Kos, M.; Hittmair, A.; Schulze, W. Mast cells in human foetal, cryptorchid and adult infertile testis. Period Biol. 1998, 100, W2. [Google Scholar]
- Meineke, V.; Frunghieri, M.B.; Jessberger, B.; Vogt, H.; Mayerhofer, A. Human testicular mast cells contain tryptase: Increased mast cell number and altered distribution in the testes of infertile men. Fertil. Steril. 2000, 74, 239–244. [Google Scholar] [CrossRef]
- Zhou, Z.; Shi, S.; Wang, S. The development and chronological changes of mast cell in testes of rats. Chin. J. Histochem. Cytochem. 1998, 7, 26–30. [Google Scholar]
- Zhao, S.; Zhu, W.; Xue, S.; Han, D. Testicular defense systems: Immune privilege and innate immunity. Cell. Mol. Immunol. 2014, 11, 428–437. [Google Scholar] [CrossRef]
- Elieh Ali Komi, D.; Shafaghat, F.; Haidl, G. Significance of mast cells in spermatogenesis, implantation, pregnancy, and abortion: Cross talk and molecular mechanisms. Am. J. Reprod. Immunol. 2020, 83, e13228. [Google Scholar] [CrossRef] [PubMed]
- Frungieri, M.B.; Weidinger, S.; Meineke, V.; Köhn, F.M.; Mayerhofer, A. Proliferative action of mast-cell tryptase is mediated by PAR2, COX2, prostaglandins, and PPARγ: Possible relevance to human fibrotic disorders. Proc. Natl. Acad. Sci. USA 2002, 99, 15072–15077. [Google Scholar] [CrossRef] [PubMed]
- Crivellato, E.; Travan, L.; Ribatti, D. The phylogenetic profile of mast cells. Methods Mol. Biol. 2015, 1220, 11–27. [Google Scholar] [PubMed]
- Wong, G.; Zhuo, L.; Kimata, K.; Lam, B.; Satoh, N.; Stevens, R. Ancient origin of mast cells. Biochem. Biophys. Res. Commun. 2014, 451, 314–318. [Google Scholar] [CrossRef]
- Dharani, P.; Kumary, S.U.; Sundaram, V.; Joseph, C.; Ramesh, G. Morphology of the Interstitial Tissue of Active and Resting Testis of the Guinea Fowl. Int. J. Morphol. 2017, 35, 1359–1362. [Google Scholar] [CrossRef][Green Version]
- Wight, P.A.L.; Mackenzie, G.M. The mast cells of Gallus domesticus. Cells Tissues Organs 1970, 75, 263–275. [Google Scholar] [CrossRef]
- Minucci, S.; di Matteo, L.; Chieffi, P.; Pierantoni, R.; Fasano, S. 17β-estradiol effects on mast cell number and spermatogonial mitotic index in the testis of the frog, Rana esculenta. J. Exp. Zool. 1997, 278, 93–100. [Google Scholar] [CrossRef]
- Matteo, L.D.; Vitiello, I.I.; Minucci, S. Effects of sex steroid hormones and their antagonists on mast cell number in the testis of the frog, Rana esculenta. Zygote 2000, 8, 225–234. [Google Scholar] [CrossRef]
- Izzo, G.; d’Istria, M.; Serino, I.; Minucci, S. Inhibition of the increased 17beta-estradiol-induced mast cell number by melatonin in the testis of the frog Rana esculenta, in vivo and in vitro. J. Exp. Biol. 2004, 207, 437–441. [Google Scholar] [CrossRef]
- Baccari, G.C.; Pinelli, C.; Santillo, A.; Minucci, S.; Rastogi, R.K. Mast cells in nonmammalian vertebrates: An overview. Int. Rev. Cell Mol. Biol. 2011, 290, 1–53. [Google Scholar]
- Guerrero, S.M.; Calderón, M.L.; de Pérez, G.R.; Pinilla, M.P.R. Morphology of the male reproductive duct system of Caiman crocodilus (Crocodylia, Alligatoridae). Ann. Anat.-Anat. Anz. 2004, 186, 235–245. [Google Scholar] [CrossRef]
- Gaytan, F.; Bellido, C.; Lucena, M.C.; Paniagua, R. Increased number of mast cells in the testis of neonatally estrogenized rats. Arch. Androl. 1986, 16, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Gaytan, F.; Bellido, C.; Carrera, G.; Aguilar, E. Differentiation of mast cells during postnatal development of neonatally estrogen-treated rats. Cell Tissue Res. 1990, 259, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, R.; Antón, F.; Bellido, C.; Aguilar, E.; Gaytan, F. Testicular serotonin is related to mast cells but not to Leydig cells in the rat. J. Endocrinol. 1995, 146, 15–21. [Google Scholar] [CrossRef]
- Iosub, R.; Klug, J.; Fijak, M.; Schneider, E.; Fröhlich, S.; Blumbach, K.; Wennemuth, G.; Sommerhoff, C.P.; Steinhoff, M.; Meinhardt, A. Development of testicular inflammation in the rat involves activation of proteinase-activated receptor-2. J. Pathol. 2006, 208, 686–698. [Google Scholar] [CrossRef] [PubMed]
- Can, C.; Töre, F.; Tunçel, N.; Uysal, O.; Gürer, F.; Ak, D.; Tunçel, M. Protective effect of vasoactive intestinal peptide on testicular torsion-detorsion injury: Association with heparin-containing mast cells. Urology 2004, 63, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Mendes, L.O.; Amorim, J.P.A.; Teixeira, G.R.; Chuffa, L.G.A.; Fioruci, B.A.; Pimentel, T.A.; de Mello, W., Jr.; Padovani, C.R.; Pereira, S.; Martinez, M.; et al. Mast Cells and Ethanol Consumption: Interactions in the Prostate, Epididymis and Testis of UChB Rats. Am. J. Reprod. Immunol. 2011, 66, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Movahed, E.; Nejati, V.; Sadrkhanlou, R.; Ahmadi, A. Toxic effect of acyclovir on testicular tissue in rats. Iran. J. Reprod. Med. 2013, 11, 111–118. [Google Scholar]
- Paula Franco Punhagui, A.; Rodrigues Vieira, H.; Eloisa Munhoz De Lion Siervo, G.; da Rosa, R.; Scantamburlo Alves Fernandes, G. Ethanol exposure during peripubertal period increases the mast cell number and impairs meiotic and spermatic parameters in adult male rats. Microsc. Res. Tech. 2016, 79, 541–549. [Google Scholar] [CrossRef]
- Wrobel, K.H.; Sinowatz, F.; Mademann, R. Intertubular topography in the bovine testis. Cell Tissue Res. 1981, 217, 289–310. [Google Scholar] [CrossRef]
- Wang, J.; Wreford, N.G.M.; Lan, H.Y.; Atkins, R.; Hedger, M.P. Leukocyte Populations of the Adult Rat Testis Following Removal of the Leydig Cells by Treatment With Ethane Dimethane Sulfonate and Subcutaneous Testosterone Implants. Biol. Reprod. 1994, 51, 551–561. [Google Scholar] [CrossRef] [PubMed]
- Majeed, S.K. Mast cell distribution in rats. Arzneimittelforschung 1994, 44, 370–374. [Google Scholar] [PubMed]
- Majeed, S.K. Mast cell distribution in mice. Arzneimittelforschung 1994, 44, 1170–1173. [Google Scholar]
- Jones, S.; Boisvert, A.; Duong, T.B.; Francois, S.; Thrane, P.; Culty, M. Disruption of Rat Testis Development Following Combined In Utero Exposure to the Phytoestrogen Genistein and Antiandrogenic Plasticizer Di-(2-Ethylhexyl) Phthalate. Biol. Reprod. 2014, 91, 1–14. [Google Scholar] [CrossRef]
- Anan, H.H.; Zidan, R.A.; Abd, E.; Baset, S.A.; Ali, M.M. Ameliorative effect of zinc oxide nanoparticles on cyclophosphamide induced testicular injury in adult rat. Tissue Cell 2018, 54, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Anton, F.; Morales, C.; Aguilar, R.; Bellido, C.; Aguilar, E.; Gaytán, F. A comparative study of mast cells and eosinophil leukocytes in the mammalian testis. Zentralbl Veterinarmed A 1998, 45, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Irani, A.M.; Schwartz, L.B. Mast cell heterogeneity. Clin. Exp. Allergy J. Br. Soc. Allergy Clin. Immunol. 1989, 19, 143–155. [Google Scholar] [CrossRef]
- Spicer, S.S. A correlative study of the histochemical properties of rodent acid mucopolysaccharides. J. Histochem. Cytochem. 1960, 8, 18–35. [Google Scholar] [CrossRef]
- Gaytan, F.; Aceitero, J.; Lucena, C.; Aguilar, E.; Pinilla, L.; Garnelo, P.; Bellido, C. Simultaneous Proliferation and Differentiation of Mast Cells and Leydig Cells in the Rat Testis. J. Androl. 1992, 13, 387–397. [Google Scholar]
- Fritz, F.J.; Pabst, R. Numbers and heterogeneity of mast cells in the male genital tract of the rat. Int. Arch. Allergy Appl. Immunol. 1989, 88, 360–362. [Google Scholar] [CrossRef]
- Klatt, E.C.; Lukes, R.J.; Meyer, P.R. Benign and malignant mast cell proliferations. Diagnosis and separation using a pH-dependent toluidine blue stain in tissue section. Cancer 1983, 51, 1119–1124. [Google Scholar] [CrossRef]
- Rodriguez, M.G.; Rodriguez, M.G.; Rival, C.; Theas, M.S.; Lustig, L. Immunohistopathology of the contralateral testis of rats undergoing experimental torsion of the spermatic cord. Asian J. Androl. 2006, 8, 576–583. [Google Scholar] [CrossRef] [PubMed]
- Jackson, A.E.; O’Leary, P.C.; Ayers, M.M.; de Kretser, D.M. The Effects of Ethylene Dimethane Suiphonate (EDS) on Rat Leydig Cells: Evidence to Support a Connective Tissue Origin of Leydig Cells. Biol. Reprod. 1986, 35, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, R.M.; Fraser, H.M.; Ratnasooriya, W.D. Assessment of the role of Leydig cell products other than testosterone in spermatogenesis and fertility in adult rats. Int. J. Androl. 1988, 11, 507–523. [Google Scholar] [CrossRef] [PubMed]
- Lustig, L.; Rodriguez, M.; Denduchis, B.; Suescun, O. Testicular mast cells in autoimmune orchitis. Am. J. Reprod. Immunol. 1995, 33, 1. [Google Scholar]
- Sampaio, C.F.; Lucchetta, N.R.; Punhagui, A.P.F.; Banedetti, P.R.; Arakawa, N.S.; Seiva, F.R.F.; Fernandes, G.S.A. Alcohol extract of Bauhinia forficata link reduces lipid peroxidation in the testis and epididymis of adult Wistar rats. Microsc. Res. Tech. 2019, 82, 345–351. [Google Scholar] [CrossRef]
- Zaidi, A.; Lendon, R.G.; Dixon, J.S.; Morris, I.D. Abnormal development of the testis after administration of the Leydig cell cytotoxic ethylene-1,2-dimethanesulphonate to the immature rat. J. Reprod. Fertil. 1988, 82, 381–392. [Google Scholar] [CrossRef]
- Gaytan, F.; Carrera, G.; Pinilla, L.; Aguilar, R.; Bellido, C. Mast cells in the testis, epididymis and accessory glands of the rat: Effects of neonatal steroid treatment. J. Androl. 1989, 10, 351–358. [Google Scholar] [CrossRef]
- Gaytán, F.; Bellido, C.; Aguilar, E.; Rooijen, N.V. Requirement for testicular macrophages in Leydig cell proliferation and differentiation during prepubertal development in rats. J. Reprod. Fertil. 1994, 102, 393–399. [Google Scholar] [CrossRef]
- Tunçel, N.; Gürer, F.; Aral, E.; Uzuner, K.; Aydin, Y.; Bayçu, C. The effect of vasoactive intestinal peptide (VIP) on mast cell invasion/degranulation in testicular interstitium of immobilized + cold stressed and beta-endorphin-treated rats. Peptides 1996, 17, 817–824. [Google Scholar] [CrossRef]
- Acikgoz, A.; Asci, R.; Aydin, O.; Çavuş, H.; Donmez, G.; Buyukalpelli, R. The role of ketotifen in the prevention of testicular damage in rats with experimental unilateral undescended testes. Drug Des. Devel. Ther. 2014, 8, 2089–2097. [Google Scholar] [CrossRef] [PubMed]
- Khramtsova, Y.S.; Artashyan, O.S.; Yushkov, B.G.; Volkova, Y.L.; Nezgovorova, N.Y. The influence of mast cells on reparative regeneration of tissues characterized by various degrees of immune privilege. Cell Tissue Biol. 2016, 10, 378–386. [Google Scholar] [CrossRef]
- Qo, S. Mast cell induction to the mouse testicular interstitium. Nihon Hinyokika Gakkai Zasshi 1994, 85, 747–752. [Google Scholar] [PubMed]
- Li, X.; Strauss, L.; Kaatrasalo, A.; Mayerhofer, A.; Huhtaniemi, I.; Santti, R.; Mäkelä, S.; Poutanen, M. Transgenic Mice Expressing P450 Aromatase as a Model for Male Infertility Associated with Chronic Inflammation in the Testis. Endocrinology 2006, 147, 1271–1277. [Google Scholar] [CrossRef] [PubMed]
- Nour, N. Involvement of Activin and Follistatin in the Pathogenesis of Chronic Testicular Inflammation in Mice. Ph.D. Thesis, Monash University, Melbourne, VIC, Australia, 2016. [Google Scholar]
- Sayed, R.K.A.; Mokhtar, D.M.; Fernández-Ortiz, M.; Escames, G.; Acuña-Castroviejo, D. Retinoid-related orphan nuclear receptor alpha (RORα)-deficient mice display morphological testicular defects. Lab. Investig. 2019, 99, 1835–1849. [Google Scholar] [CrossRef] [PubMed]
- Dhyana, R.; Rajendranath, N.; Pramod Kumar, D.; Raghavender, K.B.P. Microanatomical Studies on the Testis of Domestic Pig (Sus scrofa domestica). Int. J. Sci. Environ. Tech. 2016, 5, 2226–2231. [Google Scholar]
- Machado-Neves, M.; Neto, M.J.O.; Miranda, D.C.; Souza, A.C.F.; Castro, M.M.; Sertorio, M.N.; Carvalho, T.F.; Matta, S.L.P.; Freitas, M.B. Dietary Exposure to Tebuconazole Affects Testicular and Epididymal Histomorphometry in Frugivorous Bats. Bull Environ. Contam. Toxicol. 2018, 101, 197–204. [Google Scholar] [CrossRef]
- Souza, A.C.F.; Santos, F.C.; Bastos, D.S.S. Reproductive functions in Desmodus rotundus: A comparison between seasons in a morphological context. PLoS ONE 2018, 13, e0205023. [Google Scholar] [CrossRef]
- Menezes, T.P.; Hill, E.; de Alencar Moura, A.; Lobo, M.D.P.; Monteiro-Moreira, A.C.O.; Breton, S.; Machado-Neves, M. Pattern of protein expression in the epididymis of Oligoryzomys nigripes (Cricetidae, Sigmodontinae). Cell Tissue Res. 2018, 372, 135–147. [Google Scholar] [CrossRef]
- Frungieri, M.B.; Gonzalez-Calvar, S.I.; Rubio, M.; Ozu, M.; Lustig, L.; Calandra, R.S. Serotonin in Golden Hamster Testes: Testicular Levels, Immunolocalization and Role during Sexual Development and Photoperiodic Regression-Recrudescence Transition. Neuroendocrinology 1999, 69, 299–308. [Google Scholar] [CrossRef]
- Bakhtiary, Z.; Shahrooz, R.; Ahmadi, A.; Soltanalinejad, F. Ethyl Pyruvate Ameliorates The Damage Induced by Cyclophosphamide on Adult Mice Testes. Int. J. Fertil. Steril. 2016, 10, 79–86. [Google Scholar] [PubMed]
- Ohanian, C.; Rodriguez, H.; Piriz, H.; Martino, I.; Rieppi, G.; Garófalo, E.G.; Roca, R.A. Studies on the contractile activity and ultrastructure of the boar testicular capsule. Reproduction 1979, 57, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Kangawa, A.; Otake, M.; Enya, S.; Yoshida, T.; Shibata, M. Histological Changes of the Testicular Interstitium during Postnatal Development in Microminipigs. Toxicol. Pathol. 2019, 47, 469–482. [Google Scholar] [CrossRef] [PubMed]
- Pinart, E.; Bonet, S.; Briz, M.; Sancho, S.; García, N.; Badia, E. Cytology of the interstitial tissue in scrotal and abdominal testes of post-puberal boars. Tissue Cell 2001, 33, 8–24. [Google Scholar] [CrossRef]
- Goyal, H.O. Morphology of the bovine epididymis. Am. J. Anat. 1985, 172, 155–172. [Google Scholar] [CrossRef]
- Abd-Elmaksoud, A. Morphological, Glycohistochemical, and Immunohistochemical Studies on the Embryonic and Adult Bovine Testis. Ph.D. Thesis, LMU München, Faculty of Veterinary Medicine, Munich, Germany, 2005. [Google Scholar]
- Alkafafy, M. Glycohistochemical, Immunohistochemical and Ultrastructural Studies of the Bovine Epididymis. Ph.D. Thesis, LMU München, Faculty of Veterinary Medicine, Munich, Germany, 2005. [Google Scholar]
- Elzoghby, I.M.A.; Sosa, G.A.; Mona, N.A.H.; Manshawy, A.A. Postnatal development of the epididymis in the sheep. Benha Vet. Med. J. 2014, 26, 8. [Google Scholar]
- Collin, O.; Damber, J.E.; Bergh, A. 5-Hydroxytryptamine--a local regulator of testicular blood flow and vasomotion in rats. J. Reprod. Fertil. 1996, 106, 17–22. [Google Scholar] [CrossRef][Green Version]
- Combs, J.W.; Lagunoff, D.; Benditt, E.P. Differentiation and proliferation of embryonic mast cells of the rat. J. Cell Biol. 1965, 25, 577–592. [Google Scholar] [CrossRef]
- Yamanaka, K.; Fujisawa, M.; Tanaka, H.; Okada, H.; Arakawa, S.; Kamidono, S. Significance of human testicular mast cells and their subtypes in male infertility. Hum. Reprod. 2000, 15, 1543–1547. [Google Scholar] [CrossRef]
- Horny, H.-P.; Sotlar, K.; Valent, P. Mastocytosis: Immunophenotypical features of the transformed mast cells are unique among hematopoietic cells. Immunol. Allergy Clin. N. Am. 2014, 34, 315–321. [Google Scholar] [CrossRef]
- Unni, S.K.; Modi, D.N.; Pathak, S.G.; Dhabalia, J.V.; Bhartiya, D. Stage-specific localization and expression of c-kit in the adult human testis. J. Histochem. Cytochem. 2009, 57, 861–869. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Ospina, G. Serotonin Concentration, Synthesis, Cell Origin, and Targets in the Rat Caput Epididymis During Sexual Maturation and Variations Associated With Adult Mating Status: Morphological and Biochemical Studies. J. Androl. 2007, 28, 136–149. [Google Scholar]
- Matsuda, A.; Okayama, Y.; Ebihara, N.; Yokoi, N.; Gao, P.; Hamuro, J.; Hopkin, J.M.; Kinoshita, S. High-affinity IgE receptor-beta chain expression in human mast cells. J. Immunol. Methods 2008, 336, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Simoes, J.P.; Schoning, P. Canine mast cell tumors: A comparison of staining techniques. J. Vet. Diagn. Investig. 1994, 6, 458–465. [Google Scholar] [CrossRef]
- Teodosio, C.; Mayado, A.; Sánchez-Muñoz, L.; Morgado, J.M.; Jara-Acevedo, M.; Álvarez-Twose, I.; García-Montero, A.C.; Matito, A.; Caldas, C.; Escribano, L.; et al. The immunophenotype of mast cells and its utility in the diagnostic work-up of systemic mastocytosis. J. Leukoc. Biol. 2015, 97, 49–59. [Google Scholar] [CrossRef]
- Kritikou, E.; Depuydt, M.A.C.; de Vries, M.R.; Mulder, K.E.; Govaert, A.M.; Smit, M.D.; van Duijn, J.; Foks, A.C.; Wezel, A.; Smeets, H.J.; et al. Flow Cytometry-Based Characterization of Mast Cells in Human Atherosclerosis. Cells 2019, 8, 334. [Google Scholar] [CrossRef]
- Digital_Histology. Available online: https://digitalhistology.org/organs-systems/reproductive/male/testis/testis-proper/testis-proper-1/ (accessed on 15 January 2021).
- Mayerhofer, A.; Bartke, A.; Amador, A.G.; Began, T. Histamine affects testicular steroid production in the golden hamster. Endocrinology 1989, 125, 2212–2214. [Google Scholar] [CrossRef]
- Frungieri, M.B.; Urbanski, H.F.; Höhne-Zell, B.; Mayerhofer, A. Neuronal Elements in the Testis of the Rhesus Monkey: Ontogeny, Characterization and Relationship to Testicular Cells. Neuroendocrinology 2000, 71, 43–50. [Google Scholar] [CrossRef]
- Schmid, N.; Stöckl, J.B.; Flenkenthaler, F.; Dietrich, K.G.; Schwarzer, J.U.; Köhn, F.M.; Drummer, C.; Fröhlich, T.; Arnold, G.J.; Behr, R.; et al. Characterization of a non-human primate model for the study of testicular peritubular cells-comparison with human testicular peritubular cells. Mol. Hum. Reprod. 2018, 24, 401–410. [Google Scholar] [CrossRef]
- Hussein, M.R.; Abou-Deif, E.S.; Bedaiwy, M.A.; Said, T.M.; Mustafa, M.G.; Nada, E.; Ezat, A.; Agarwal, A. Phenotypic characterization of the immune and mast cell infiltrates in the human testis shows normal and abnormal spermatogenesis. Fertil. Steril. 2005, 83, 1447–1453. [Google Scholar] [CrossRef]
- Apa, D.D.; Çayan, S.; Polat, A.; Akbay, E. Mast Cells and Fibrosis on Testicular Biopsies in Male Infertility. Arch. Androl. 2002, 48, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Jezek, D.; Banek, L.; Hittmair, A.; Pezerović-Panijan, R.; Goluža, T.; Schulze, W. Mast cells in testicular biopsies of infertile men with ‘mixed atrophy’ of seminiferous tubules. Andrologia 1999, 31, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Shanafelt, T.D.; Lin, T.; Geyer, S.M.; Zent, C.S.; Leung, N.; Kabat, B.; Bowen, D.; Grever, M.R.; Byrd, J.C.; Kay, N.E. Pentostatin, cyclophosphamide, and rituximab regimen in older patients with chronic lymphocytic leukemia. Cancer 2007, 109, 2291–2298. [Google Scholar] [CrossRef] [PubMed]
- Mulvihill, J.J.; Garlow, T.J. Reproductive Outcomes among Men Treated for Cancer. In Male-Mediated Developmental Toxicity; The Royal Society of Chemistry: London, UK, 2007; Chapter 2; pp. 7–14. [Google Scholar]
- Kitamura, Y.; Yokoyama, M.; Matsuda, H.; Ohno, T.; Mori, K.J. Spleen colony-forming cell as common precursor for tissue mast cells and granulocytes. Nature 1981, 291, 159–160. [Google Scholar] [CrossRef]
- Narayana, K. A purine nucleoside analogue-acyclovir [9-(2-hydroxyethoxymethyl)-9h-guanine] reversibly impairs testicular functions in mouse. J. Toxicol. Sci. 2008, 33, 61–70. [Google Scholar] [CrossRef]
- Fletcher, C.; Bean, B.; McLeod, D.C. Evaluation of Oral Acyclovir Therapy. Drug Intell. Clin. Pharm. 1985, 19, 518–524. [Google Scholar] [CrossRef]
- Le Bigot, J.F.; Begue, J.M.; Kiechel, J.R.; Guillouzo, A. Species differences in metabolism of ketotifen in rat, rabbit and man: Demonstration of similar pathways in vivo and in cultured hepatocytes. Life Sci. 1987, 40, 883–890. [Google Scholar] [CrossRef]
- Schill, W.B.; Schneider, J.; Ring, J. The use of ketotifen, a mast cell blocker, for treatment of oligo- and asthenozoospermia. Andrologia 1986, 18, 570–573. [Google Scholar] [CrossRef]
- Oliva, A.; Multigner, L. Ketotifen improves sperm motility and sperm morphology in male patients with leukocytospermia and unexplained infertility. Fertil. Steril. 2006, 85, 240–243. [Google Scholar] [CrossRef]
- Azadi, L.; Abbasi, H.; Deemeh, M.R.; Tavalaee, M.; Arbabian, M.; Pilevarian, A.A.; Nasr-Esfahani, M.H. Zaditen (Ketotifen), as mast cell blocker, improves sperm quality, chromatin integrity and pregnancy rate after varicocelectomy. Int. J. Androl. 2011, 34, 446–452. [Google Scholar] [CrossRef]
- Moreno, D.; Sobarzo, C.M.; Lustig, L.; Rodríguez Peña, M.G.; Guazzone, V.A. Effect of ketotifen fumarate on experimental autoimmune orchitis and torsion of the spermatic cord. Asian J. Androl. 2020, 22, 112–117. [Google Scholar] [PubMed]
- Iosub, I.R. Pathomechanism of Testicular Inflammation in Rat Involves Activation of Proteinase Activated Receptor 2. Ph.D. Thesis, University Of Giessen, Giesen, Germany, 2006. [Google Scholar]
- Windschüttl, S.; Nettersheim, D.; Schlatt, S.; Huber, A.; Welter, H.; Schwarzer, J.U.; Köhn, F.M.; Schorle, H.; Mayerhofer, A. Are testicular mast cells involved in the regulation of germ cells in man? Andrology 2014, 2, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Karakuş, S.C.; Süzen, A.; Yenisey, Ç.; Ertürk, N.; Epikmen, T.; İpek, E.; User, I.R.; Abas, B.I. The effect of hypothermia in a rat testicular torsion/detorsion model. J. Pediatr. Urol. 2021, 17, 291.e1–291.e8. [Google Scholar] [CrossRef]
- Nocka, K.; Tan, J.C.; Chiu, E.; Chu, T.Y.; Ray, P.; Traktman, P.; Besmer, P. Molecular bases of dominant negative and loss of function mutations at the murine c-kit/white spotting locus: W37, Wv, W41 and W. EMBO J. 1990, 9, 1805–1813. [Google Scholar] [CrossRef] [PubMed]
- Lotinun, S.; Krishnamra, N. Disruption of c-Kit Signaling in KitW-sh/W-sh Growing Mice Increases Bone Turnover. Sci. Rep. 2016, 6, 31515. [Google Scholar] [CrossRef]
- Loveland, K.L.; Schlatt, S. Stem cell factor and c-kit in the mammalian testis: Lessons originating from Mother Nature’s gene knockouts. J. Endocrinol. 1997, 153, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Meyer, N.; Jiao, Q.; Scheffel, J.; Zimmermann, C.; Metz, M.; Zenclussen, A.; Maurer, M.; Siebenhaar, F. Chymase-Cre; Mcl-1fl/fl Mice Exhibit Reduced Numbers of Mucosal Mast Cells. Front. Immunol. 2019, 10, 2399. [Google Scholar] [CrossRef] [PubMed]
- Mendis-Handagama, S.M.; Ariyaratne, H.B.; Fectau, K.A.; Grizzle, J.M.; Jayasundera, N.K. Comparison of testis structure, function and thyroid hormone levels in control C57BL/6 mice and anti-mullerian hormone over expressing mice. Histol. Histopathol. 2010, 25, 901–908. [Google Scholar]
- Mechlin, C.W.; Levesque, J.; Feustel, P.; Kogan, B. A Mast cell numbers negatively correlate with fibrosis in cryptorchid testes. J. Pediatr. Urol. 2014, 10, 527–531. [Google Scholar] [CrossRef]
- Bostancierİ, N.; Duran, İ.; Yüncü, M. Investigation of Testicular Mast Cells and Fibrosis in Rats With Experimental Unilateral Cryptorchidism. Bozok Tıp Dergisi 2021, 11, 7. [Google Scholar]
- Jantos, C.; Baumgärtner, W.; Durchfeld, B.; Schiefer, H. Experimental epididymitis due to Chlamydia trachomatis in rats. Infect. Immun. 1992, 60, 2324–2328. [Google Scholar] [CrossRef] [PubMed]
- Anosa, V.O.; Kaneko, J.J. Pathogenesis of Trypanosoma brucei infection in deer mice (Peromyscus maniculatus). Ultrastructural pathology of the spleen, liver, heart, and kidney. Vet. Pathol. 1984, 21, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Fijak, M.; Meinhardt, A. The testis in immune privilege. Immunol. Rev. 2006, 213, 66–81. [Google Scholar] [CrossRef] [PubMed]
- Mruk, D.D. Emergent roles for intercellular adhesion molecule-1 in the restructuring of the blood-testis barrier during spermatogenesis in the mammal. Histol. Histopathol. 2016, 31, 159–166. [Google Scholar] [PubMed]
- Li, N.; Tang, E.I.; Cheng, C.Y. Regulation of blood-testis barrier by actin binding proteins and protein kinases. Reproduction 2016, 151, R29–R41. [Google Scholar] [CrossRef] [PubMed]
- Mruk, D.D.; Cheng, C.Y. The Mammalian Blood-Testis Barrier: Its Biology and Regulation. Endocr. Rev. 2015, 36, 564–591. [Google Scholar] [CrossRef] [PubMed]
- Mayerhofer, A.; Walenta, L.; Mayer, C.; Eubler, K.; Welter, H. Human testicular peritubular cells, mast cells and testicular inflammation. Andrologia 2018, 50, e13055. [Google Scholar] [CrossRef] [PubMed]
- Schulze, M.; Buder, S.; Rüdiger, K.; Beyerbach, M.; Waberski, D. Influences on semen traits used for selection of young AI boars. Anim. Reprod. Sci. 2014, 148, 164–170. [Google Scholar] [CrossRef]
- Sancho, S.; Pinart, E.; Briz, M.; Garcia-Gil, N.; Badia, E.; Bassols, J.; Kádár, E.; Pruneda, A.; Bussalleu, E.; Yeste, M.; et al. Semen quality of postpubertal boars during increasing and decreasing natural photoperiods. Theriogenology 2004, 62, 1271–1282. [Google Scholar] [CrossRef]
- Kozdrowski, R.; Dubiel, A. The effect of season on the properties of wild boar (Sus scrofa L.) semen. Anim Reprod Sci. 2004, 80, 281–289. [Google Scholar] [CrossRef]
- Frossi, B.; Mion, F.; Sibilano, R.; Danelli, L.; Pucillo, C.E.M. Is it time for a new classification of mast cells? What do we know about mast cell heterogeneity? Immunol. Rev. 2018, 282, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Claman, H.N.; Choi, K.L.; Sujansky, W.; Vatter, A.E. Mast cell “disappearance” in chronic murine graft-vs-host disease (GVHD)-ultrastructural demonstration of “phantom mast cells”. J. Immunol. 1986, 137, 2009–2013. [Google Scholar] [PubMed]
- Walls, A.F.; Amalinei, C. Detection of Mast Cells and Basophils by Immunohistochemistry. Methods Mol. Biol. 2020, 2163, 263–280. [Google Scholar] [PubMed]
- Oliver, E.; Stukenborg, J.B. Rebuilding the human testis in vitro. Andrology 2020, 8, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Lara, N.D.L.E.M.; Sakib, S.; Dobrinski, I. Regulation of Cell Types Within Testicular Organoids. Endocrinology 2021, 162, bqab033. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Himelreich-Perić, M.; Katušić-Bojanac, A.; Hohšteter, M.; Sinčić, N.; Mužić-Radović, V.; Ježek, D. Mast Cells in the Mammalian Testis and Epididymis—Animal Models and Detection Methods. Int. J. Mol. Sci. 2022, 23, 2547. https://doi.org/10.3390/ijms23052547
Himelreich-Perić M, Katušić-Bojanac A, Hohšteter M, Sinčić N, Mužić-Radović V, Ježek D. Mast Cells in the Mammalian Testis and Epididymis—Animal Models and Detection Methods. International Journal of Molecular Sciences. 2022; 23(5):2547. https://doi.org/10.3390/ijms23052547
Chicago/Turabian StyleHimelreich-Perić, Marta, Ana Katušić-Bojanac, Marko Hohšteter, Nino Sinčić, Vedrana Mužić-Radović, and Davor Ježek. 2022. "Mast Cells in the Mammalian Testis and Epididymis—Animal Models and Detection Methods" International Journal of Molecular Sciences 23, no. 5: 2547. https://doi.org/10.3390/ijms23052547
APA StyleHimelreich-Perić, M., Katušić-Bojanac, A., Hohšteter, M., Sinčić, N., Mužić-Radović, V., & Ježek, D. (2022). Mast Cells in the Mammalian Testis and Epididymis—Animal Models and Detection Methods. International Journal of Molecular Sciences, 23(5), 2547. https://doi.org/10.3390/ijms23052547