
Cytokine-Induced JAK2-STAT3 Activates Tissue Regeneration under Systemic or Local Inflammation


Abstract
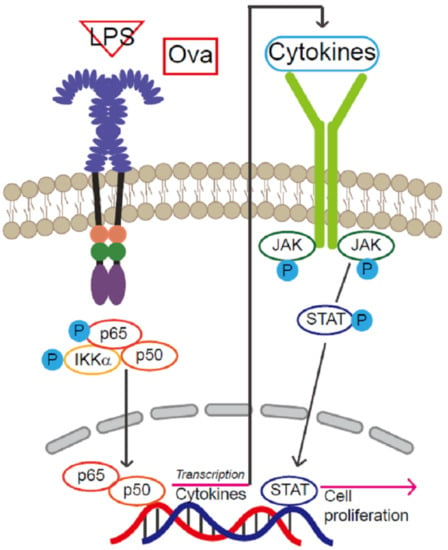
:
1. Introduction
2. Results
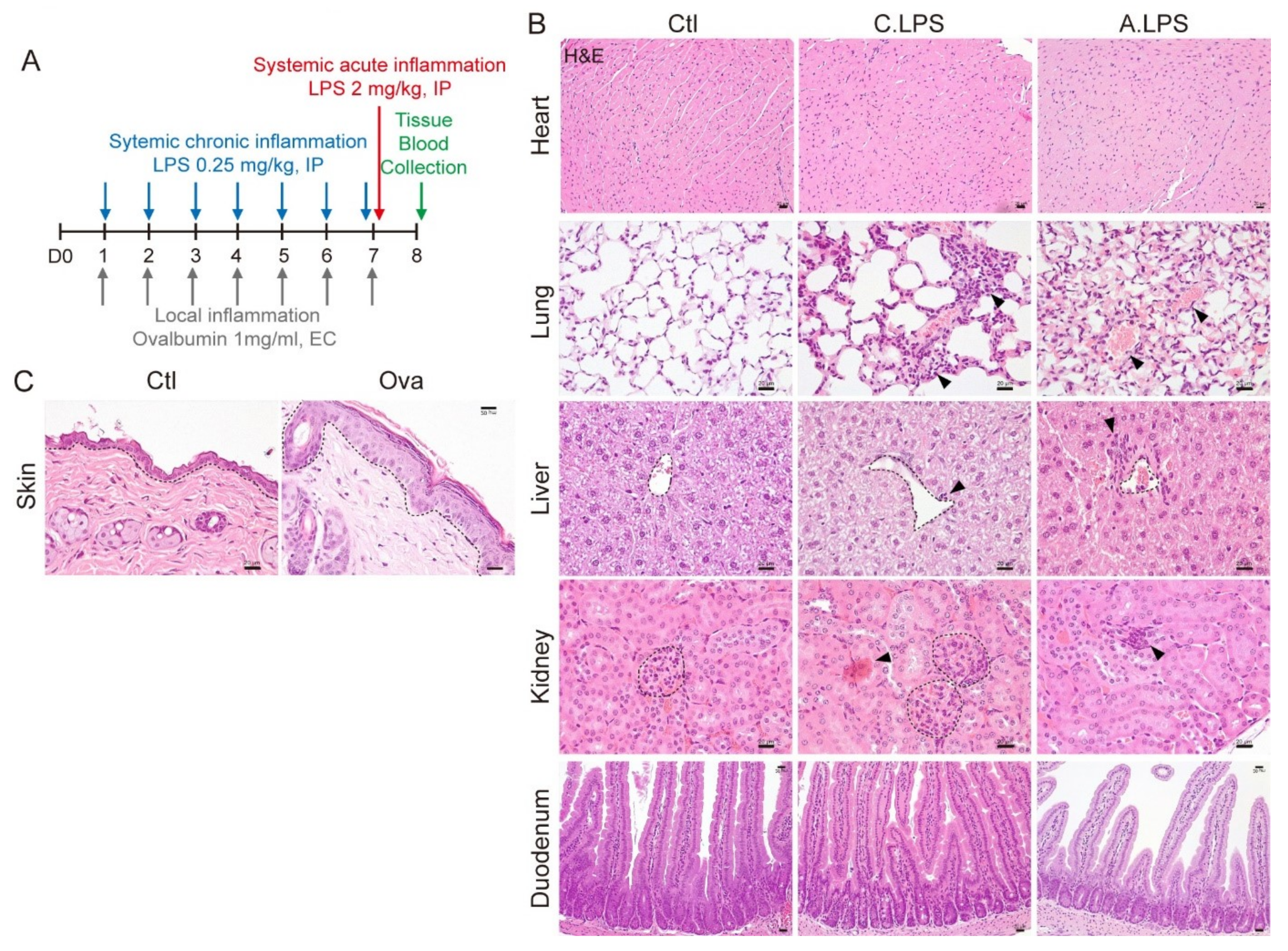
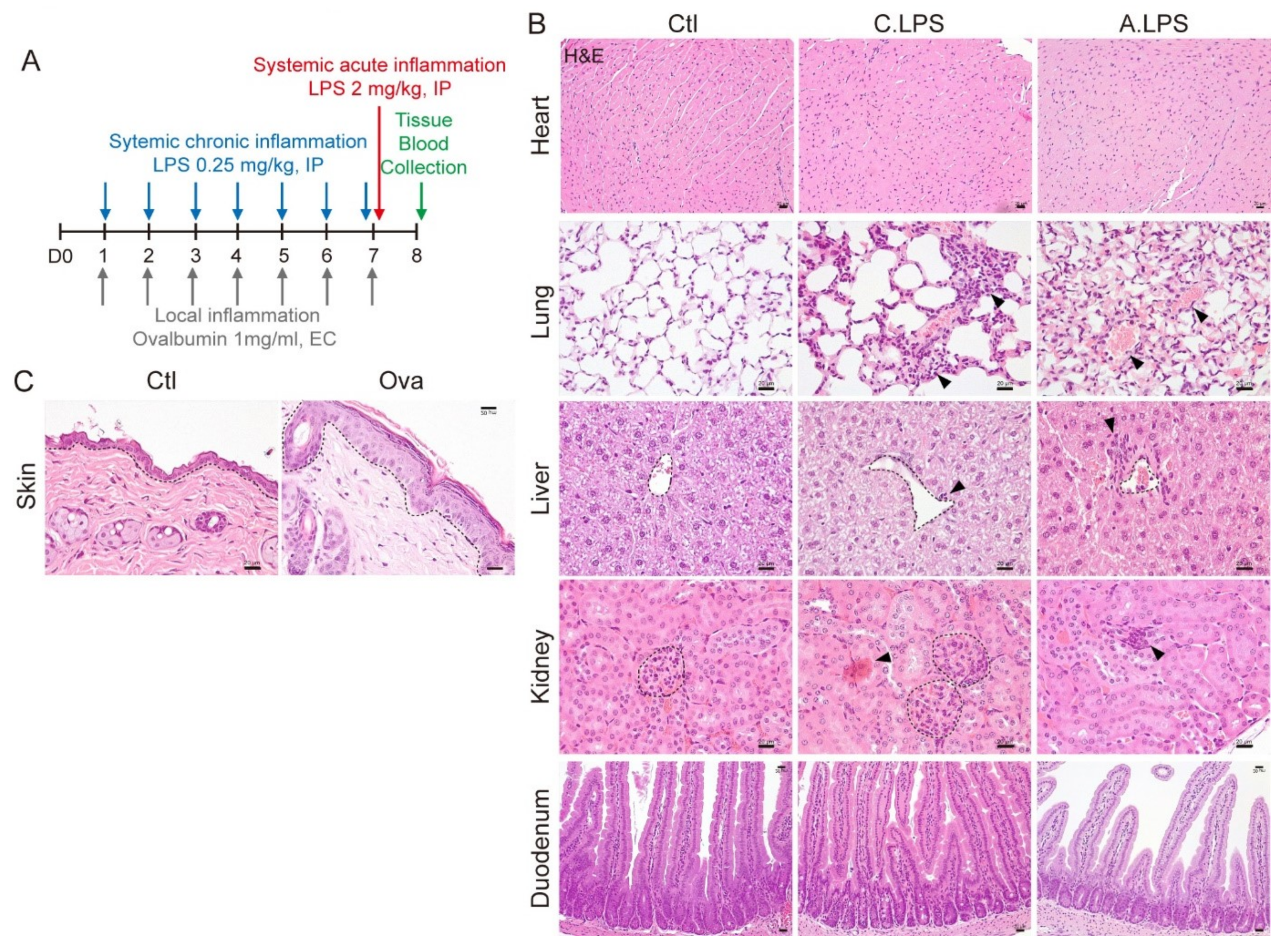
2.1. Validation of Systemic and Local Inflammation Mouse Models Using Histopathology
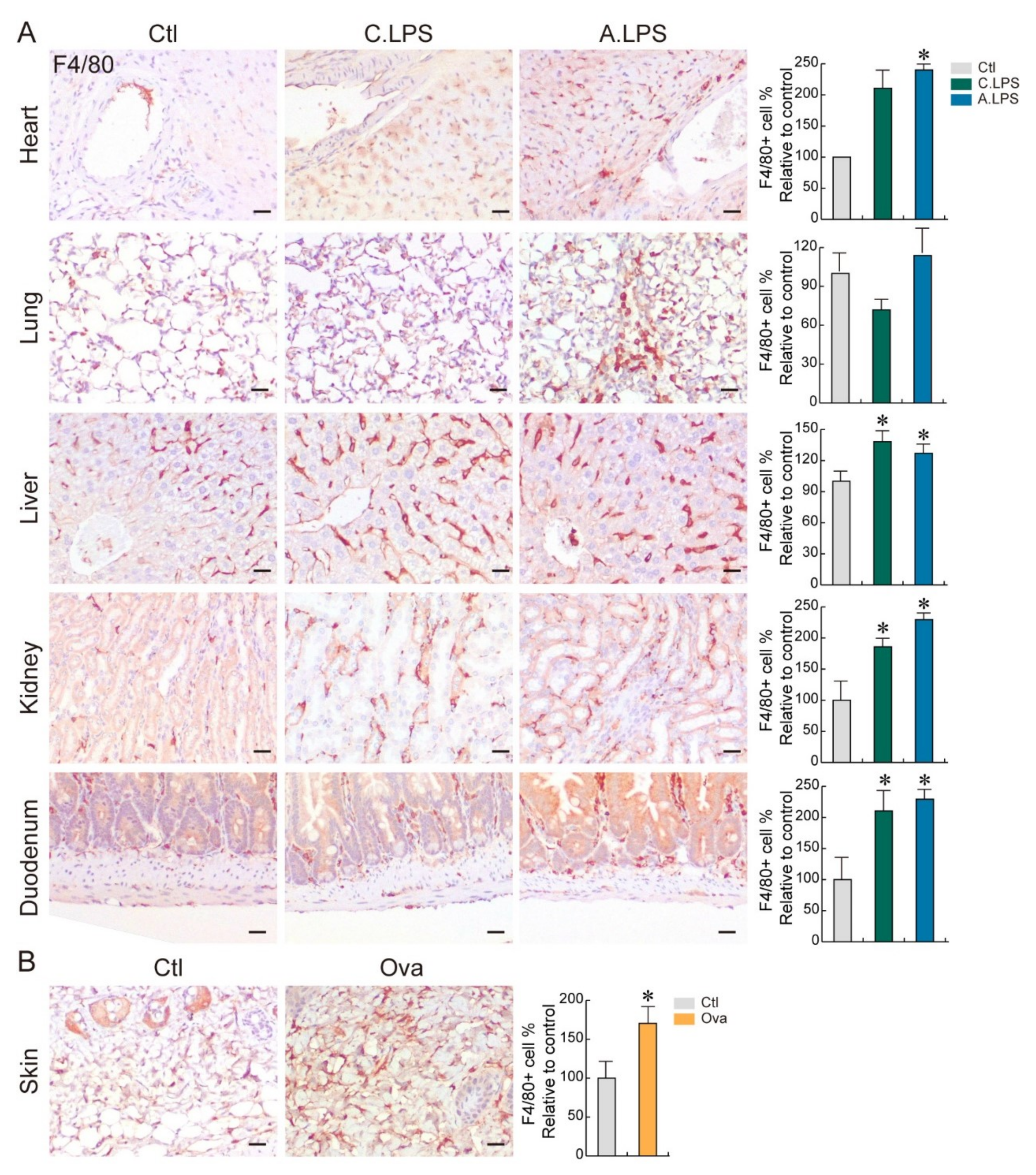
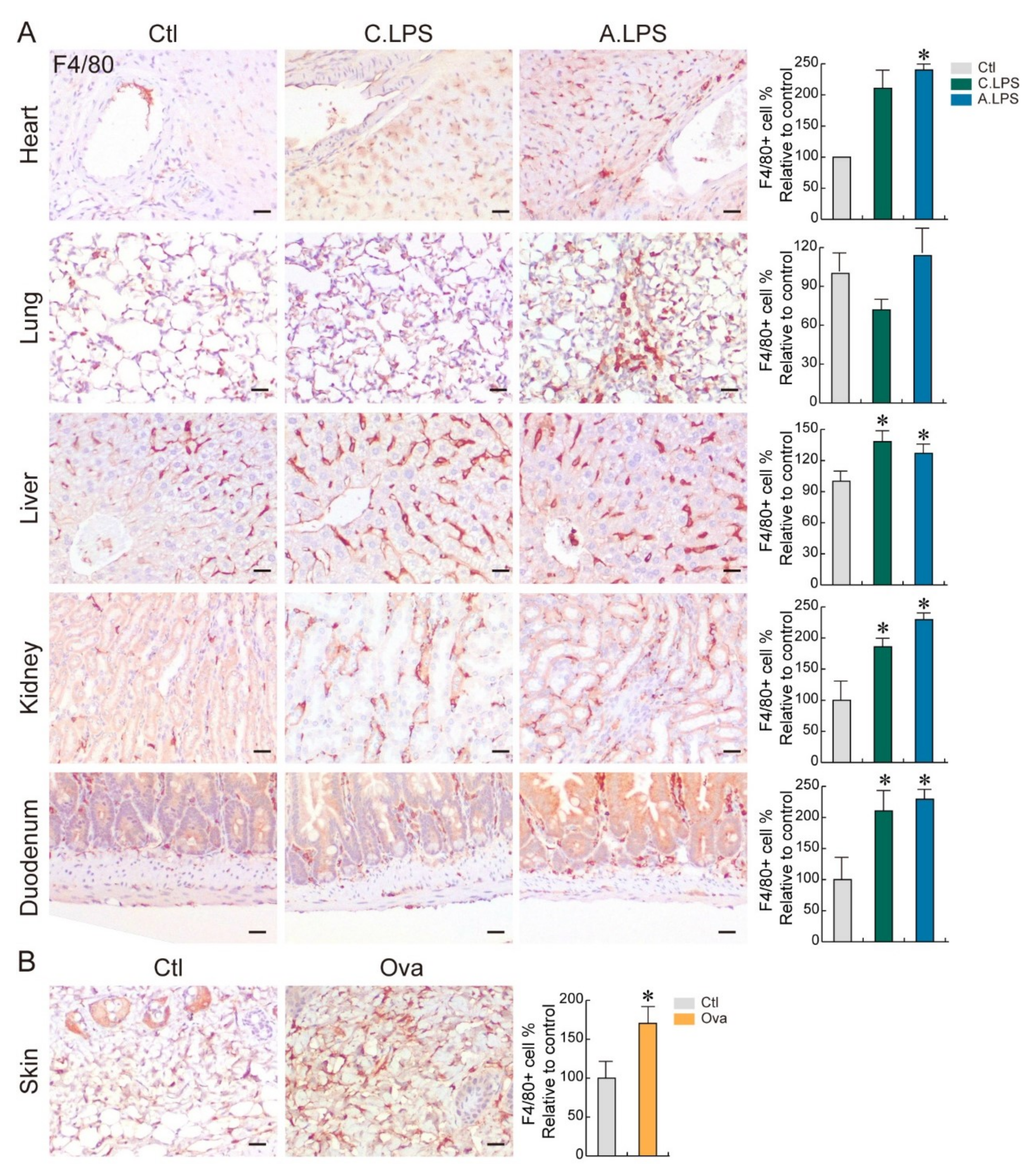
2.2. Detection of Immune Cells through Immunohistochemistry to Confirm Inflammation
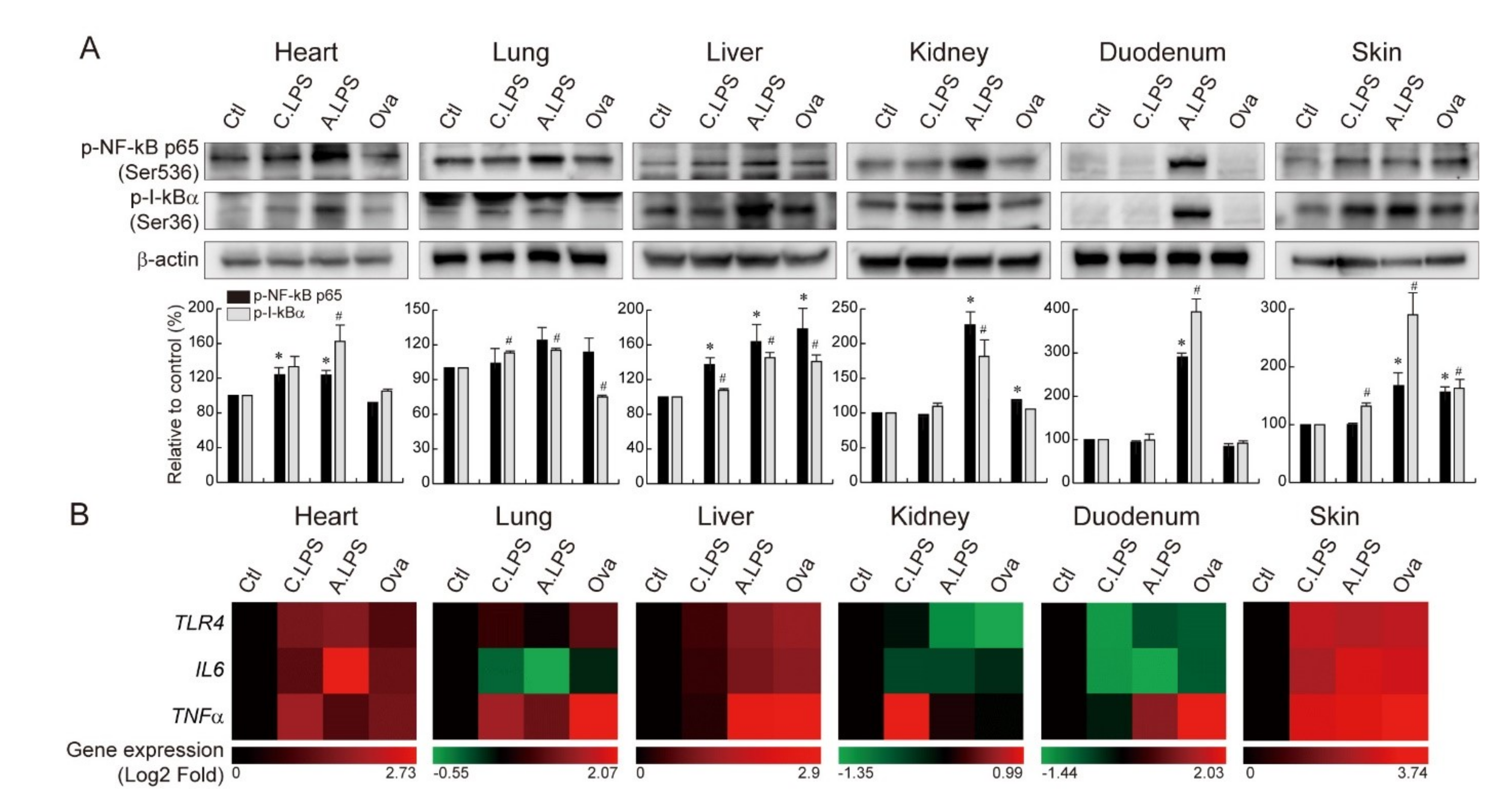
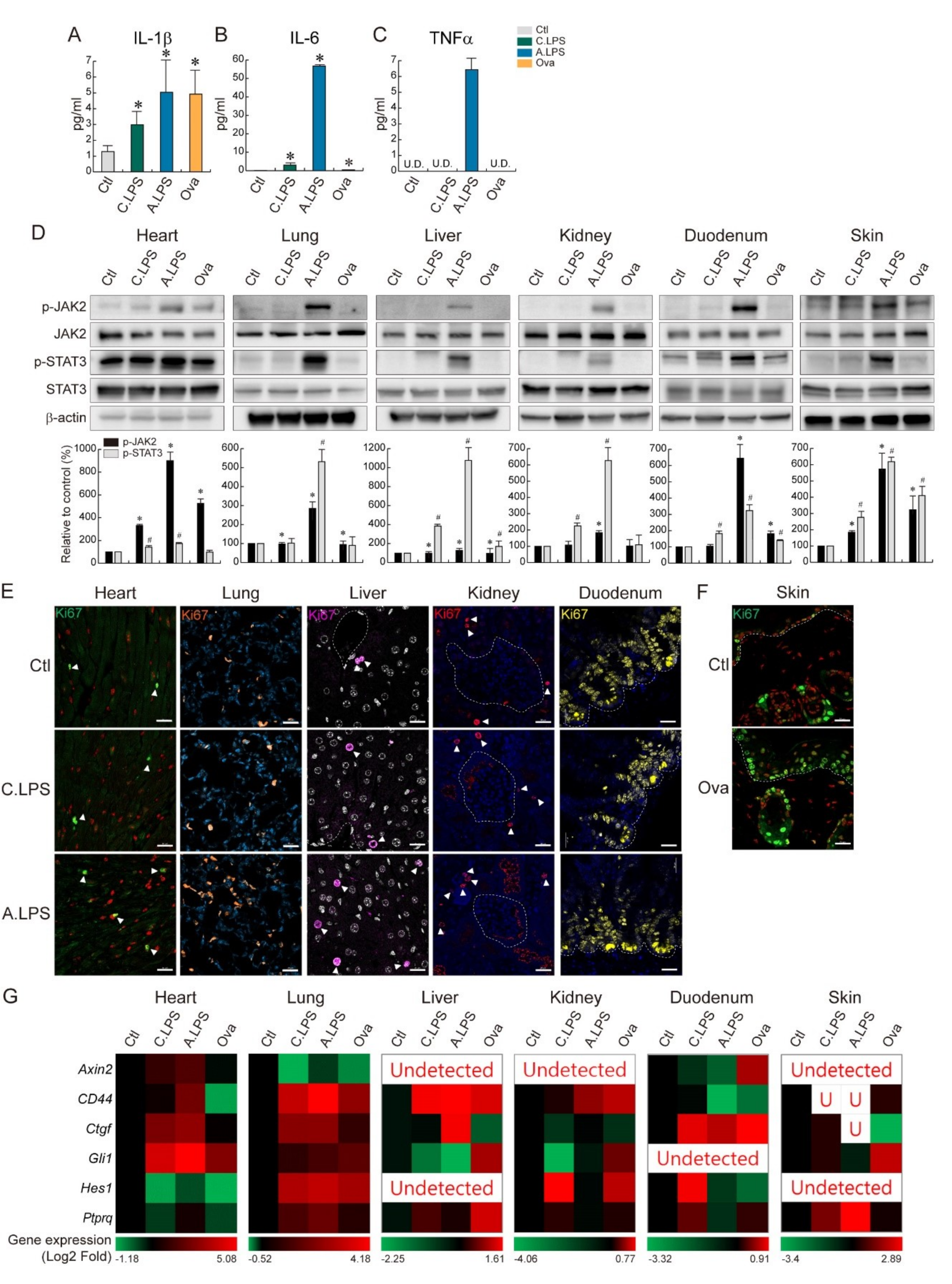
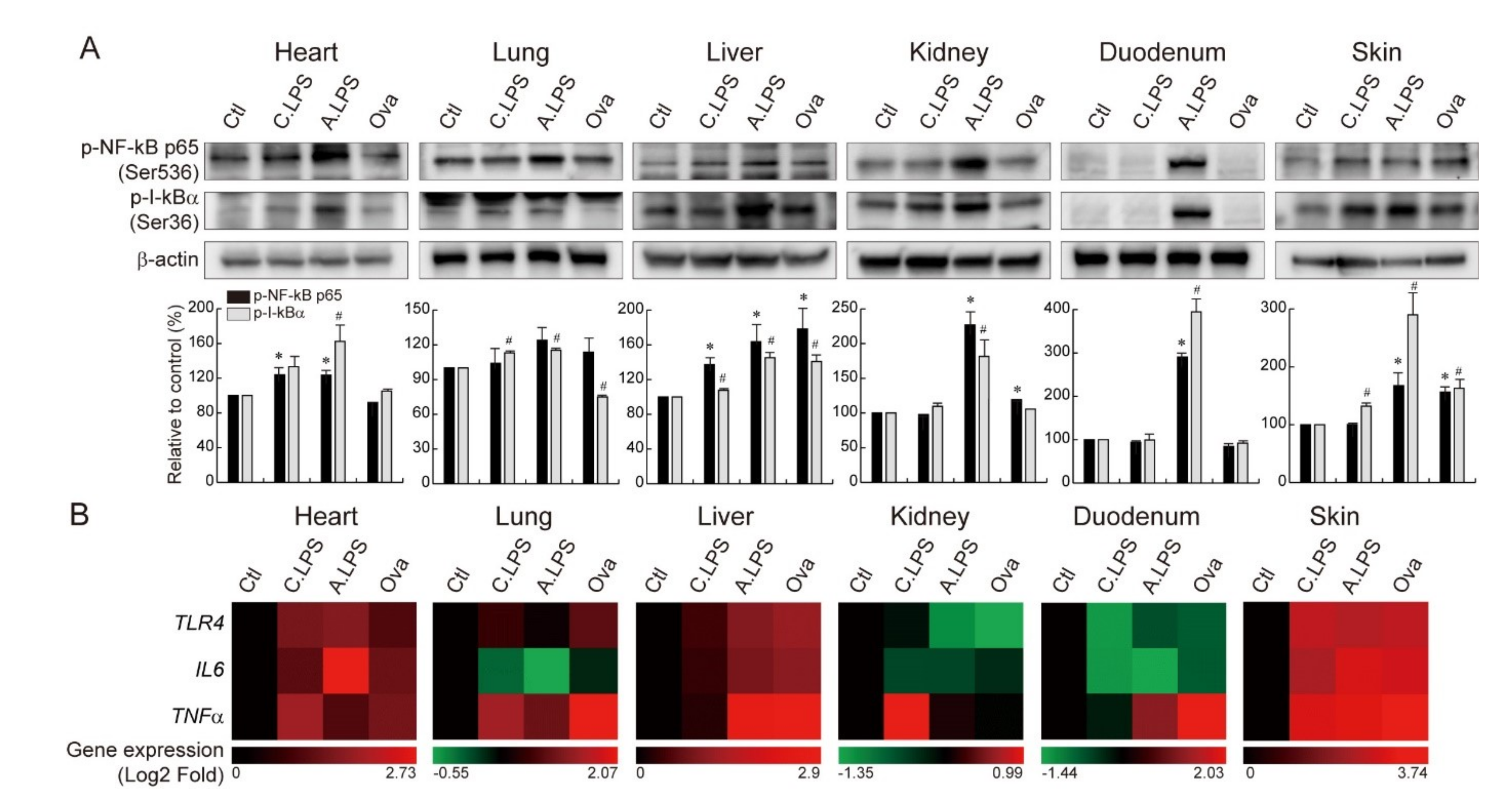
2.3. Increased NF-κB Activation and NF-κB Target Gene Expression under Inflammation
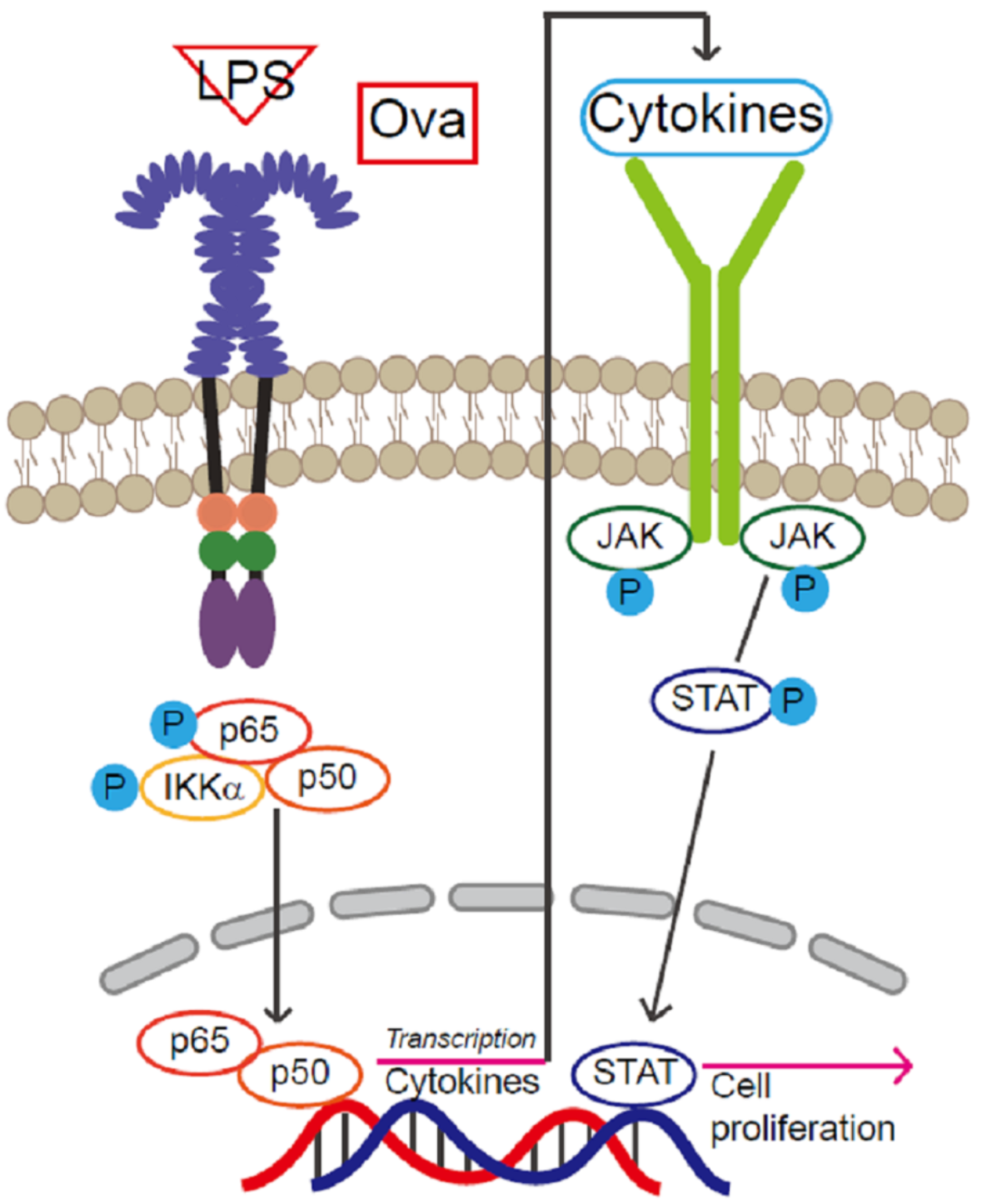
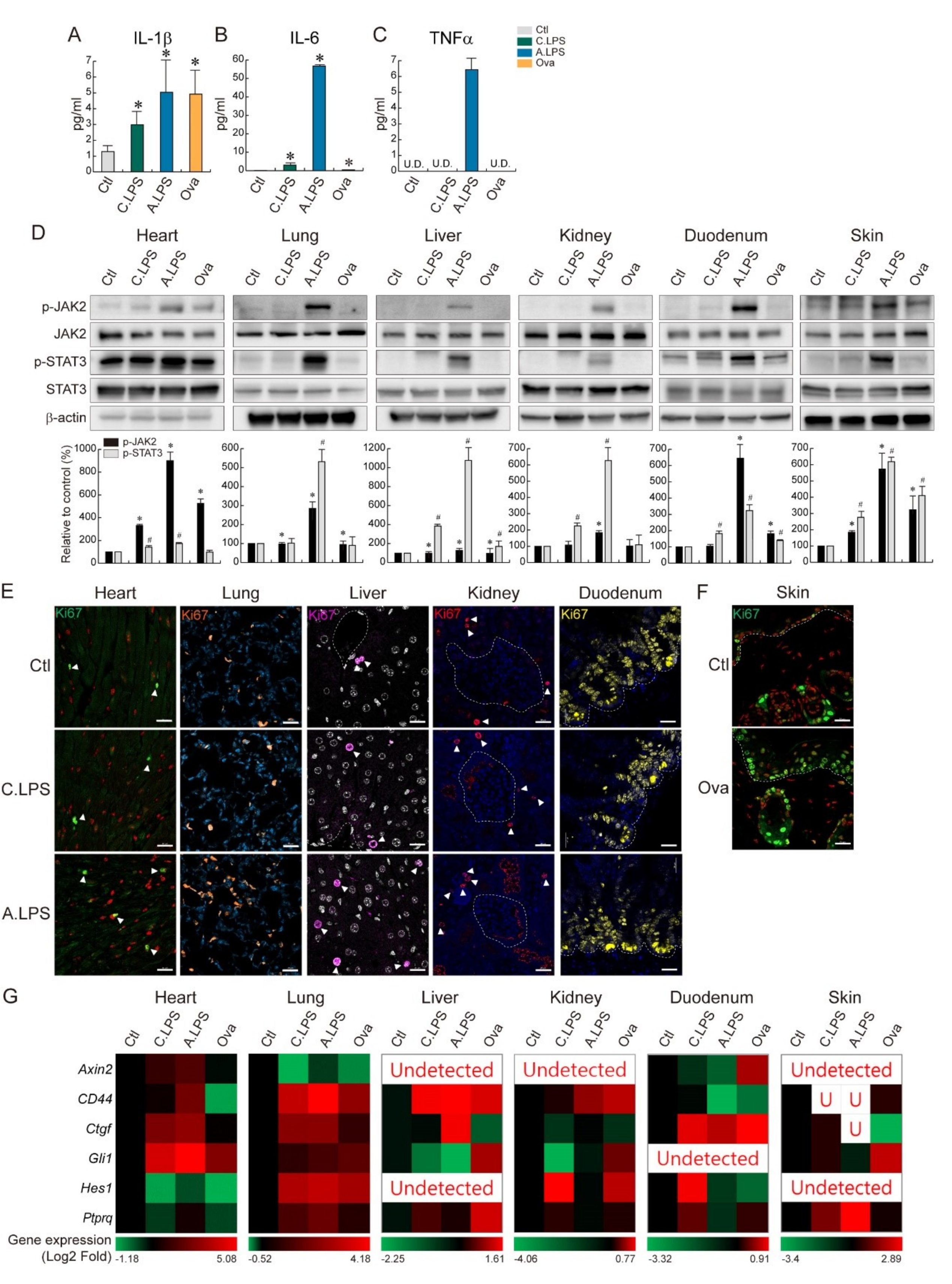
2.4. JAK2- STAT3-Induced Tissue Regeneration after Inflammatory Damage
3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.2. Histological Analysis
Histopathology
4.3. Immunohistochemistry
4.4. Immunoblot
4.5. Gene Analysis
4.6. Enzyme-Linked Immunosorbent Assay (ELISA)
4.7. Statistical Analyses
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Van Dyke, T.E.; van Winkelhoff, A.J. Infection and inflammatory mechanisms. J. Clin. Periodontol. 2013, 40 (Suppl. 14), S1–S7. [Google Scholar] [CrossRef]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [Green Version]
- Stortz, J.A.; Raymond, S.L.; Mira, J.C.; Moldawer, L.L.; Mohr, A.M.; Efron, P.A. Murine Models of Sepsis and Trauma: Can We Bridge the Gap? ILAR J. 2017, 58, 90–105. [Google Scholar] [CrossRef] [Green Version]
- Rossol, M.; Heine, H.; Meusch, U.; Quandt, D.; Klein, C.; Sweet, M.J.; Hauschildt, S. LPS-induced cytokine production in human monocytes and macrophages. Crit. Rev. Immunol. 2011, 31, 379–446. [Google Scholar] [CrossRef]
- Kim, W.H.; An, H.J.; Kim, J.Y.; Gwon, M.G.; Gu, H.; Jeon, M.; Sung, W.J.; Han, S.M.; Pak, S.C.; Kim, M.K.; et al. Beneficial effects of melittin on ovalbumin-induced atopic dermatitis in mouse. Sci. Rep. 2017, 7, 17679. [Google Scholar] [CrossRef] [Green Version]
- Casaro, M.; Souza, V.R.; Oliveira, F.A.; Ferreira, C.M. OVA-Induced Allergic Airway Inflammation Mouse Model. Methods Mol. Biol. 2019, 1916, 297–301. [Google Scholar] [CrossRef]
- Batmaz, S.B. Simple Markers for Systemic Inflammation in Pediatric Atopic Dermatitis Patients. Indian J. Dermatol. 2018, 63, 305–310. [Google Scholar] [CrossRef]
- Leung, D.Y.; Boguniewicz, M.; Howell, M.D.; Nomura, I.; Hamid, Q.A. New insights into atopic dermatitis. J. Clin. Investig. 2004, 113, 651–657. [Google Scholar] [CrossRef]
- Morris, R.; Kershaw, N.J.; Babon, J.J. The molecular details of cytokine signaling via the JAK/STAT pathway. Protein Sci. 2018, 27, 1984–2009. [Google Scholar] [CrossRef] [Green Version]
- Oshima, H.; Kok, S.Y.; Nakayama, M.; Murakami, K.; Voon, D.C.; Kimura, T.; Oshima, M. Stat3 is indispensable for damage-induced crypt regeneration but not for Wnt-driven intestinal tumorigenesis. FASEB J. 2019, 33, 1873–1886. [Google Scholar] [CrossRef] [Green Version]
- Dos Anjos Cassado, A. F4/80 as a Major Macrophage Marker: The Case of the Peritoneum and Spleen. Results Probl. Cell Differ. 2017, 62, 161–179. [Google Scholar] [CrossRef]
- Fink, M.P. Animal models of sepsis. Virulence 2014, 5, 143–153. [Google Scholar] [CrossRef]
- Malliaras, K.; Zhang, Y.; Seinfeld, J.; Galang, G.; Tseliou, E.; Cheng, K.; Sun, B.; Aminzadeh, M.; Marban, E. Cardiomyocyte proliferation and progenitor cell recruitment underlie therapeutic regeneration after myocardial infarction in the adult mouse heart. EMBO Mol. Med. 2013, 5, 191–209. [Google Scholar] [CrossRef]
- van der Flier, L.G.; Clevers, H. Stem cells, self-renewal, and differentiation in the intestinal epithelium. Annu. Rev. Physiol. 2009, 71, 241–260. [Google Scholar] [CrossRef]
- Suh, H.N.; Kim, Y.K.; Lee, J.Y.; Kang, G.H.; Hwang, J.H. Dissect the immunity using cytokine profiling and NF-kB target gene analysis in systemic inflammatory minipig model. PLoS ONE 2021, 16, e0252947. [Google Scholar] [CrossRef]
- Helal, M.G.; Megahed, N.A.; Abd Elhameed, A.G. Saxagliptin mitigates airway inflammation in a mouse model of acute asthma via modulation of NF-kB and TLR4. Life Sci. 2019, 239, 117017. [Google Scholar] [CrossRef]
- Li, H.Y.; Meng, J.X.; Liu, Z.; Liu, X.W.; Huang, Y.G.; Zhao, J. Propofol Attenuates Airway Inflammation in a Mast Cell-Dependent Mouse Model of Allergic Asthma by Inhibiting the Toll-like Receptor 4/Reactive Oxygen Species/Nuclear Factor kappaB Signaling Pathway. Inflammation 2018, 41, 914–923. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yang, N.; Wang, T.; Dai, B.; Shang, Y. Vitamin D reduces inflammatory response in asthmatic mice through HMGB1/TLR4/NFkappaB signaling pathway. Mol. Med. Rep. 2018, 17, 2915–2920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Wang, L.; Chen, B.; Zhuo, Q.; Bao, C.; Lin, L. Propofol inhibits NF-kappaB activation to ameliorate airway inflammation in ovalbumin (OVA)-induced allergic asthma mice. Int. Immunopharmacol. 2017, 51, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Ahmetaj-Shala, B.; Kirkby, N.S.; Knowles, R.; Al’Yamani, M.; Mazi, S.; Wang, Z.; Tucker, A.T.; Mackenzie, L.; Armstrong, P.C.; Nusing, R.M.; et al. Evidence that links loss of cyclooxygenase-2 with increased asymmetric dimethylarginine: Novel explanation of cardiovascular side effects associated with anti-inflammatory drugs. Circulation 2015, 131, 633–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinrich, P.C.; Behrmann, I.; Haan, S.; Hermanns, H.M.; Muller-Newen, G.; Schaper, F. Principles of interleukin (IL)-6-type cytokine signalling and its regulation. Biochem. J. 2003, 374, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miscia, S.; Marchisio, M.; Grilli, A.; Di Valerio, V.; Centurione, L.; Sabatino, G.; Garaci, F.; Zauli, G.; Bonvini, E.; Di Baldassarre, A. Tumor necrosis factor alpha (TNF-alpha) activates Jak1/Stat3-Stat5B signaling through TNFR-1 in human B cells. Cell Growth Differ. 2002, 13, 13–18. [Google Scholar] [PubMed]
- Jiang, H.; Patel, P.H.; Kohlmaier, A.; Grenley, M.O.; McEwen, D.G.; Edgar, B.A. Cytokine/Jak/Stat signaling mediates regeneration and homeostasis in the Drosophila midgut. Cell 2009, 137, 1343–1355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Liang, X.; Kellendonk, C.; Poli, V.; Taub, R. STAT3 contributes to the mitogenic response of hepatocytes during liver regeneration. J. Biol. Chem. 2002, 277, 28411–28417. [Google Scholar] [CrossRef] [Green Version]
- Beumer, J.; Clevers, H. Cell fate specification and differentiation in the adult mammalian intestine. Nat. Rev. Mol. Cell Biol. 2021, 22, 39–53. [Google Scholar] [CrossRef]
- Li, L.; Clevers, H. Coexistence of quiescent and active adult stem cells in mammals. Science 2010, 327, 542–545. [Google Scholar] [CrossRef] [Green Version]
- Post, Y.; Clevers, H. Defining Adult Stem Cell Function at Its Simplest: The Ability to Replace Lost Cells through Mitosis. Cell Stem Cell 2019, 25, 174–183. [Google Scholar] [CrossRef]
- Suh, H.N.; Kim, M.J.; Jung, Y.S.; Lien, E.M.; Jun, S.; Park, J.I. Quiescence Exit of Tert(+) Stem Cells by Wnt/beta-Catenin Is Indispensable for Intestinal Regeneration. Cell Rep. 2017, 21, 2571–2584. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Company & Cat# | Dilution |
|---|---|---|
| p-NF-KB p65 (Ser536) | Cell Signaling #3033 | 1:1000 |
| p-I-KBα (Ser36) | Abcam ab133462 | 1:1000 |
| p-JAK2 | Cell Signaling #3776 | 1:1000 |
| JAK2 | Cell Signaling #3230 | 1:1000 |
| p-STAT3 | Cell Signaling #9145 | 1:2000 |
| STAT3 | Cell Signaling #4904 | 1:2000 |
| β-actin | Santa Cruz sc-47778 | 1:5000 |
| Gene Symbol | Primer Sequences (from 5′ to 3′) | Length | Gene Bank ID |
|---|---|---|---|
| TNFα | F: CCC TCA CAC TCA GAT CAT CTT CT | 61 | NM 013693 |
| R: GCT ACG ACG TGG GCT ACA G | |||
| IL-6 | F: AGT CCT TCC TAC CCC AAT TTC C | 78 | NM 031168 |
| R: GTC TTG GTC CTT AGC CAC TCC | |||
| TLR4 | F: GTG CCA GTC AGG GTC ATT CA | 119 | NM 021297 |
| R: ACT CCC CAG CCC TTT ATG GA | |||
| Axin2 | F: TGCATCTCTCTCTGGAGGTG | 149 | NM 015732.4 |
| R: TATGTCTTTGCACCAGCCAC | |||
| CD44 | F: AGCGGCAGGTTACATTCAAA | 95 | NM 001177787.1 |
| R: CAAGTTTTGGTGGCACACAG | |||
| Ctgf | F: AGCCTCAAACTCCAAACACC | 181 | NM 010217.2 |
| R: CAACAGGGATTTGACCAC | |||
| Gli1 | F: ACCACCCTACCTCTGTCTATT | 121 | NM 010296.2 |
| R: TTCAGACCATTGCCCATCAC | |||
| Hes1 | F: GGTATTTCCCCAACACGGT | 101 | NM 008235.2 |
| R: GGCAGACATTCTGGAAATGA | |||
| Ptprq | F: CGGAGGTTACTGGAACCGTG | 111 | NM 008981.3 |
| R: CAGGGTCCCCACATAGCCT | |||
| 18s | F: AAG TCC CTG CCC TTT GTA CAC A | 70 | NR 003278.3 |
| R: GAT CCG AGG GCC TCA CTA AAC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.K.; Lee, J.Y.; Suh, H.N. Cytokine-Induced JAK2-STAT3 Activates Tissue Regeneration under Systemic or Local Inflammation. Int. J. Mol. Sci. 2022, 23, 2262. https://doi.org/10.3390/ijms23042262
Kim YK, Lee JY, Suh HN. Cytokine-Induced JAK2-STAT3 Activates Tissue Regeneration under Systemic or Local Inflammation. International Journal of Molecular Sciences. 2022; 23(4):2262. https://doi.org/10.3390/ijms23042262
Chicago/Turabian StyleKim, Young Kyu, Ju Young Lee, and Han Na Suh. 2022. "Cytokine-Induced JAK2-STAT3 Activates Tissue Regeneration under Systemic or Local Inflammation" International Journal of Molecular Sciences 23, no. 4: 2262. https://doi.org/10.3390/ijms23042262
APA StyleKim, Y. K., Lee, J. Y., & Suh, H. N. (2022). Cytokine-Induced JAK2-STAT3 Activates Tissue Regeneration under Systemic or Local Inflammation. International Journal of Molecular Sciences, 23(4), 2262. https://doi.org/10.3390/ijms23042262