
PACAP-38 Induces Transcriptomic Changes in Rat Trigeminal Ganglion Cells Related to Neuroinflammation and Altered Mitochondrial Function Presumably via PAC1/VPAC2 Receptor-Independent Mechanism

 ,
, 
Abstract
:1. Introduction
2. Results
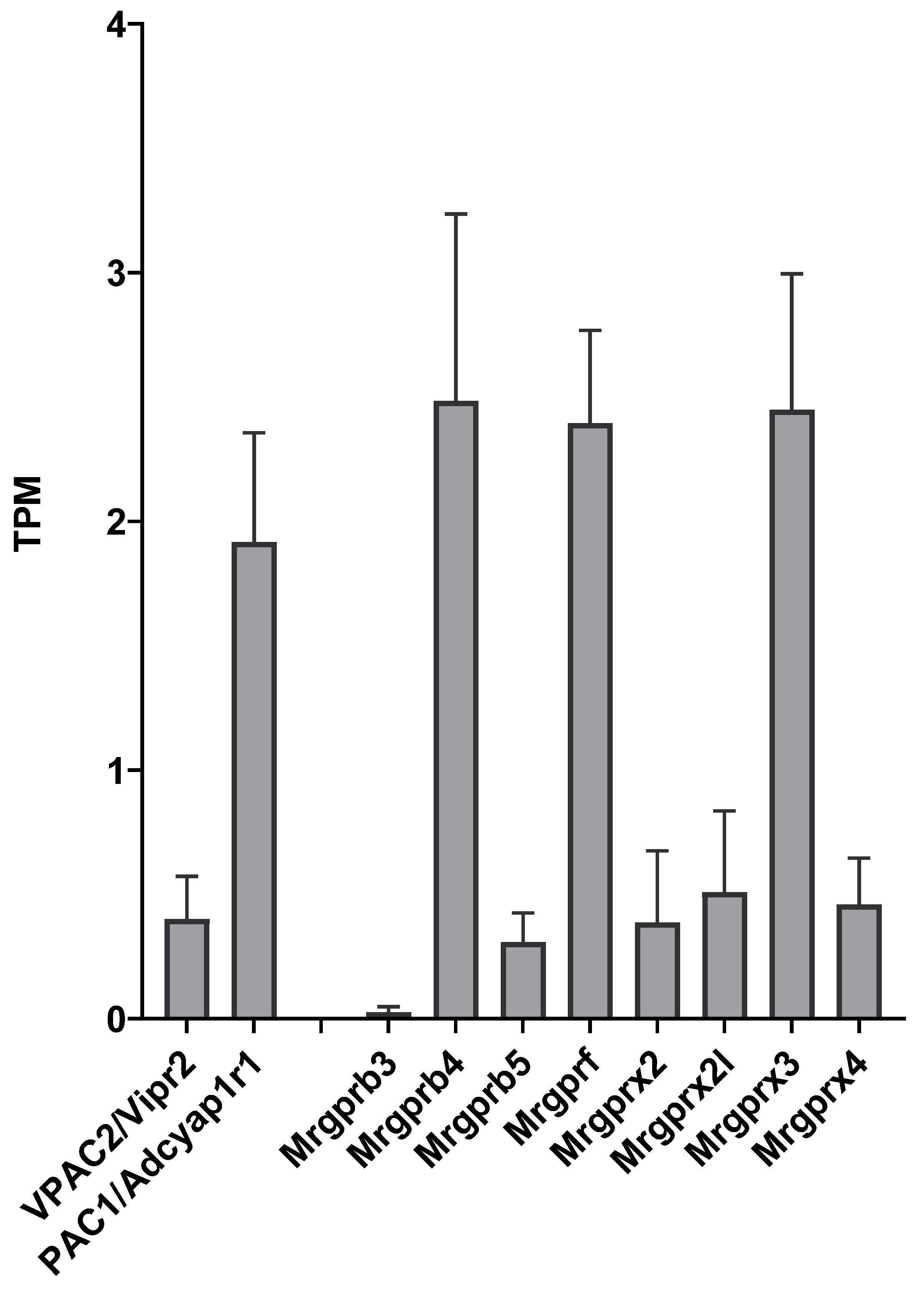
2.1. Expression of PAC1, VPAC2 and Mrgpr Receptor Transcripts in TG Cultures
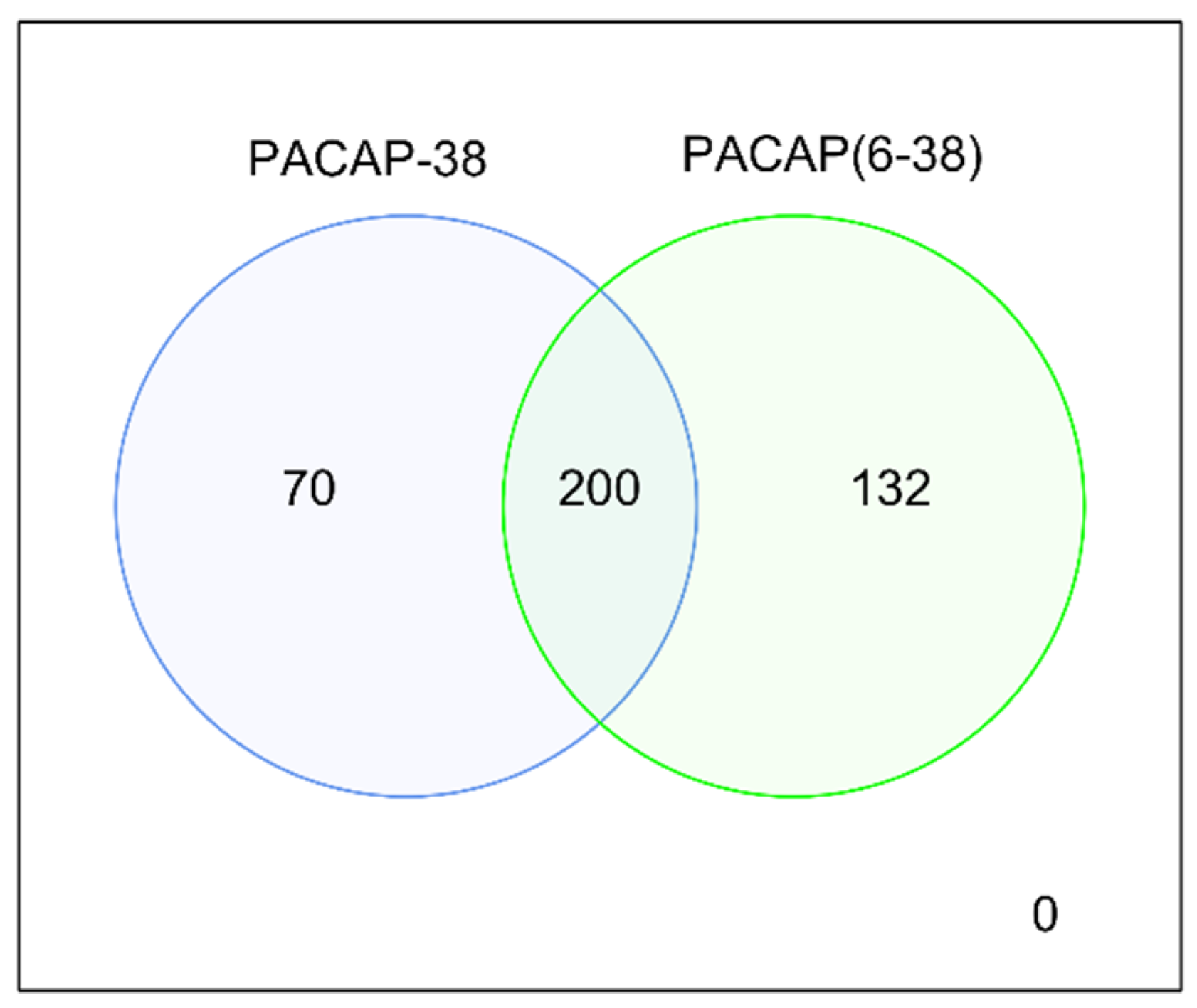
2.2. Differentially-Expressed (DE) Genes in Both PACAP-38- and PACAP(6-38)-Treated Trigeminal Ganglion Cultures
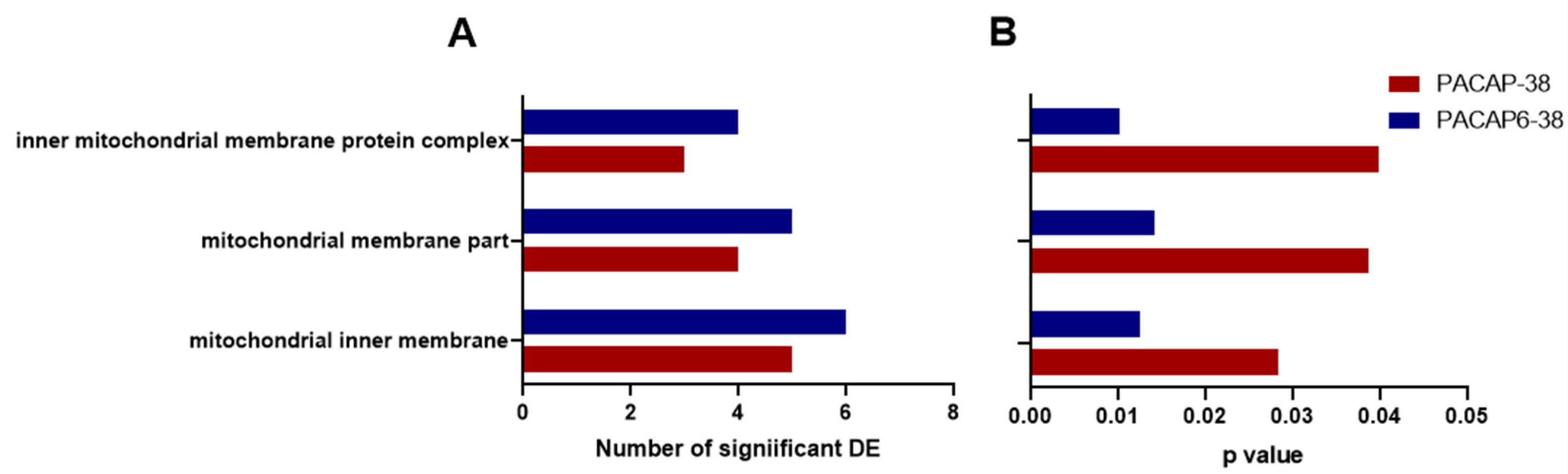
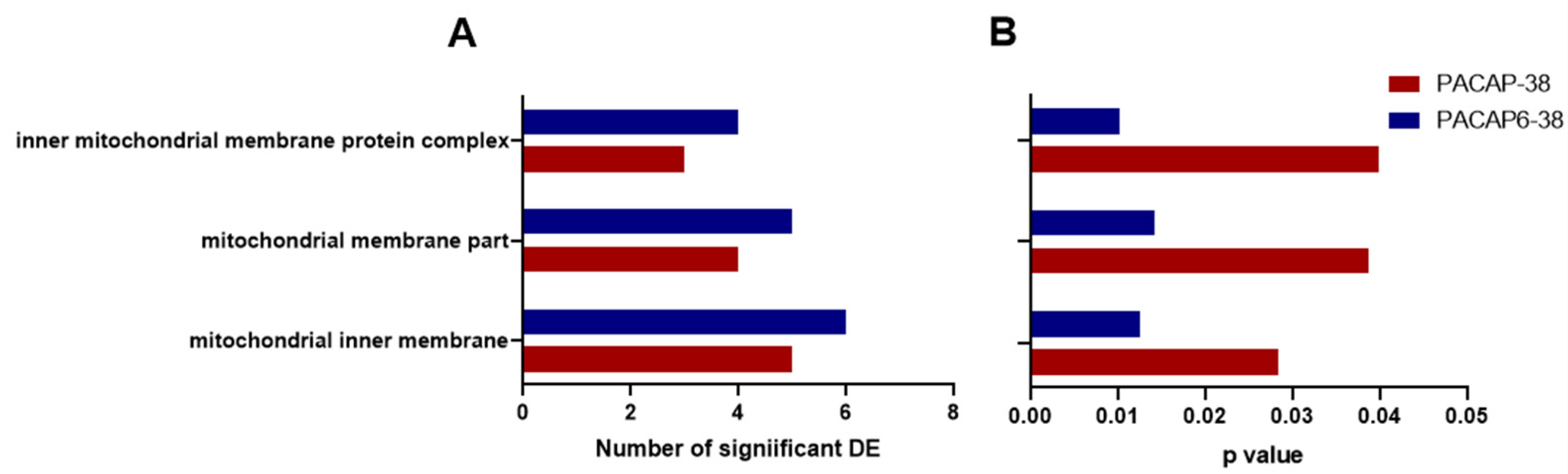
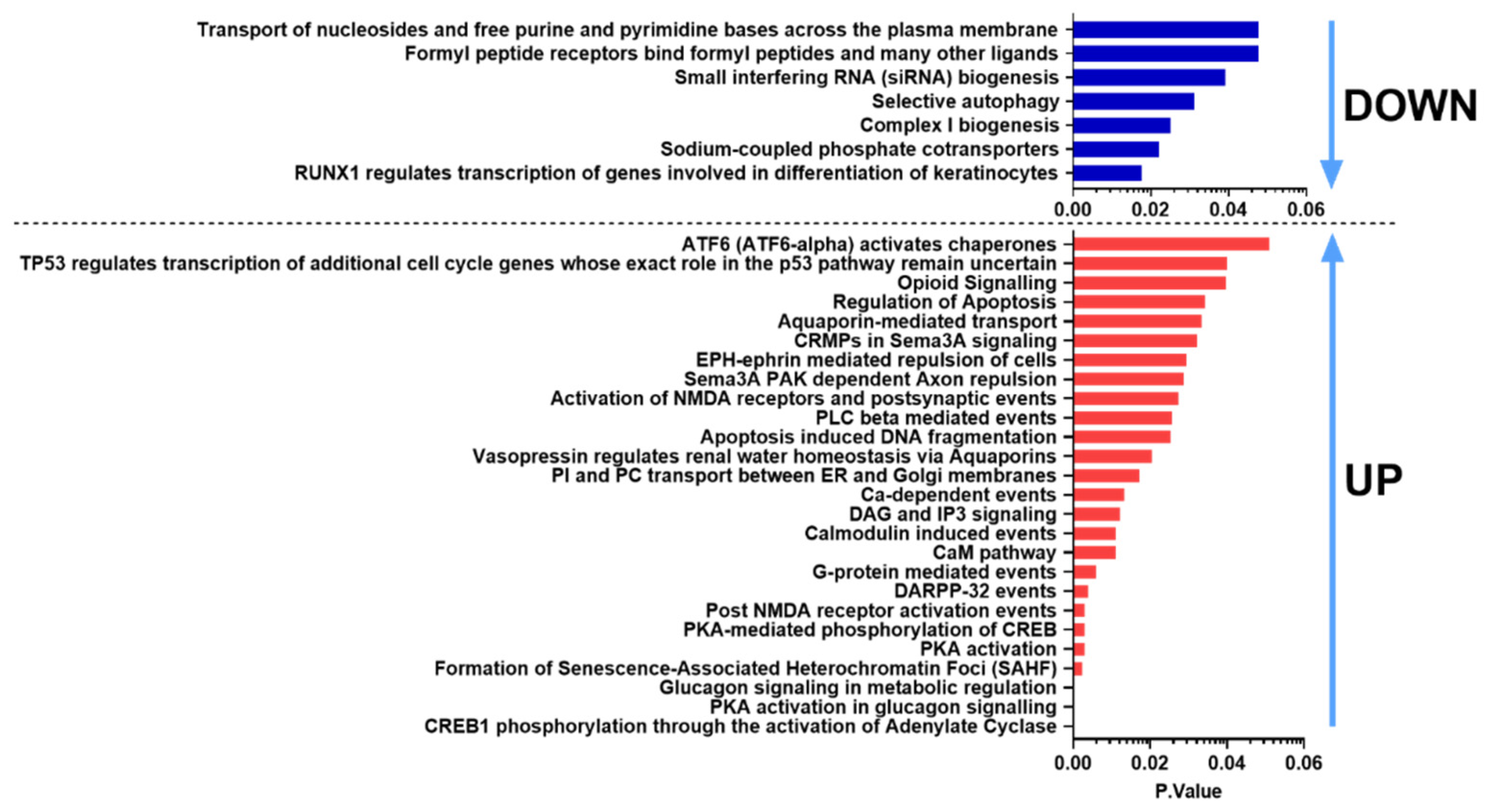
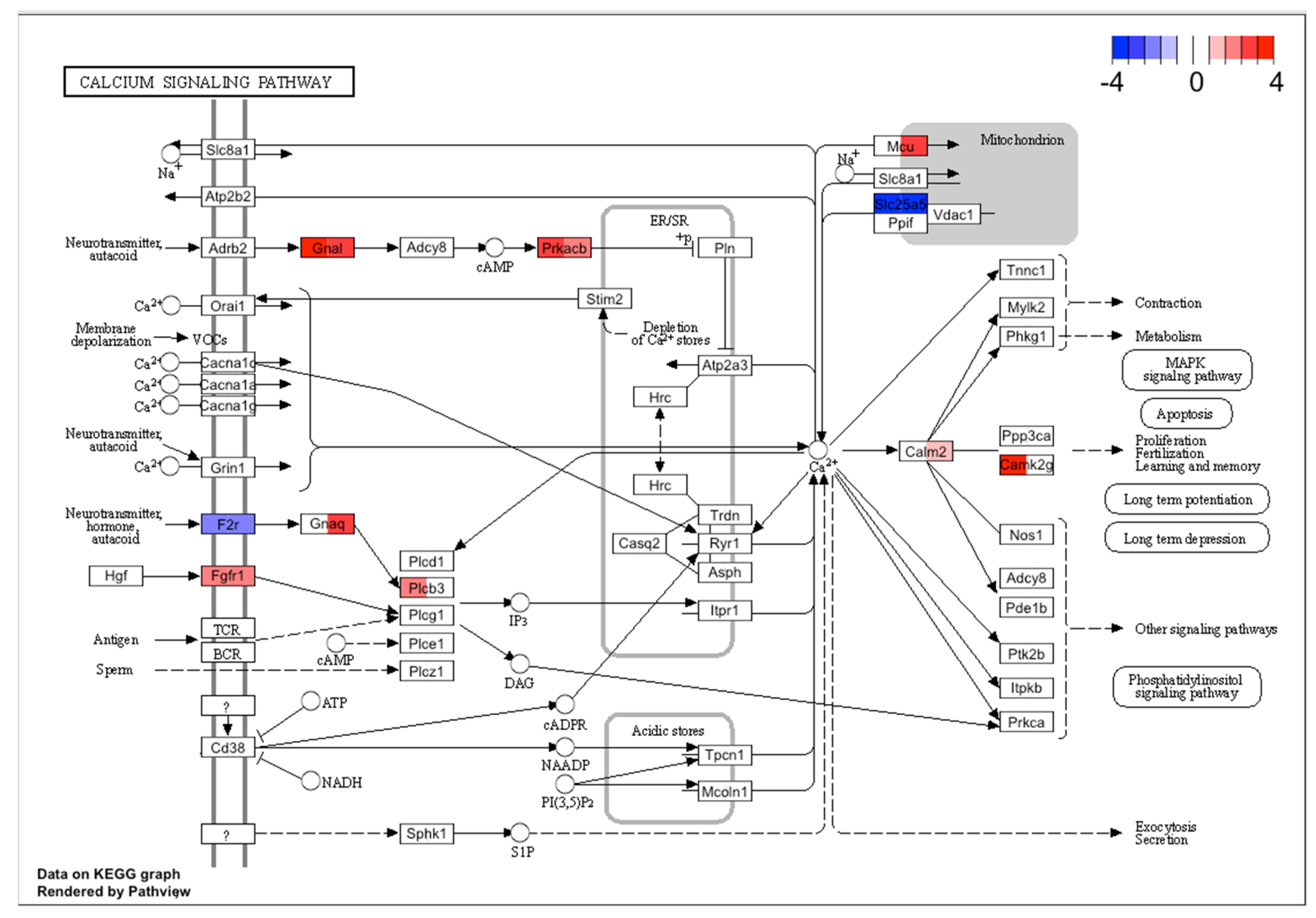
2.3. Pathway Analysis with Common DE Genes
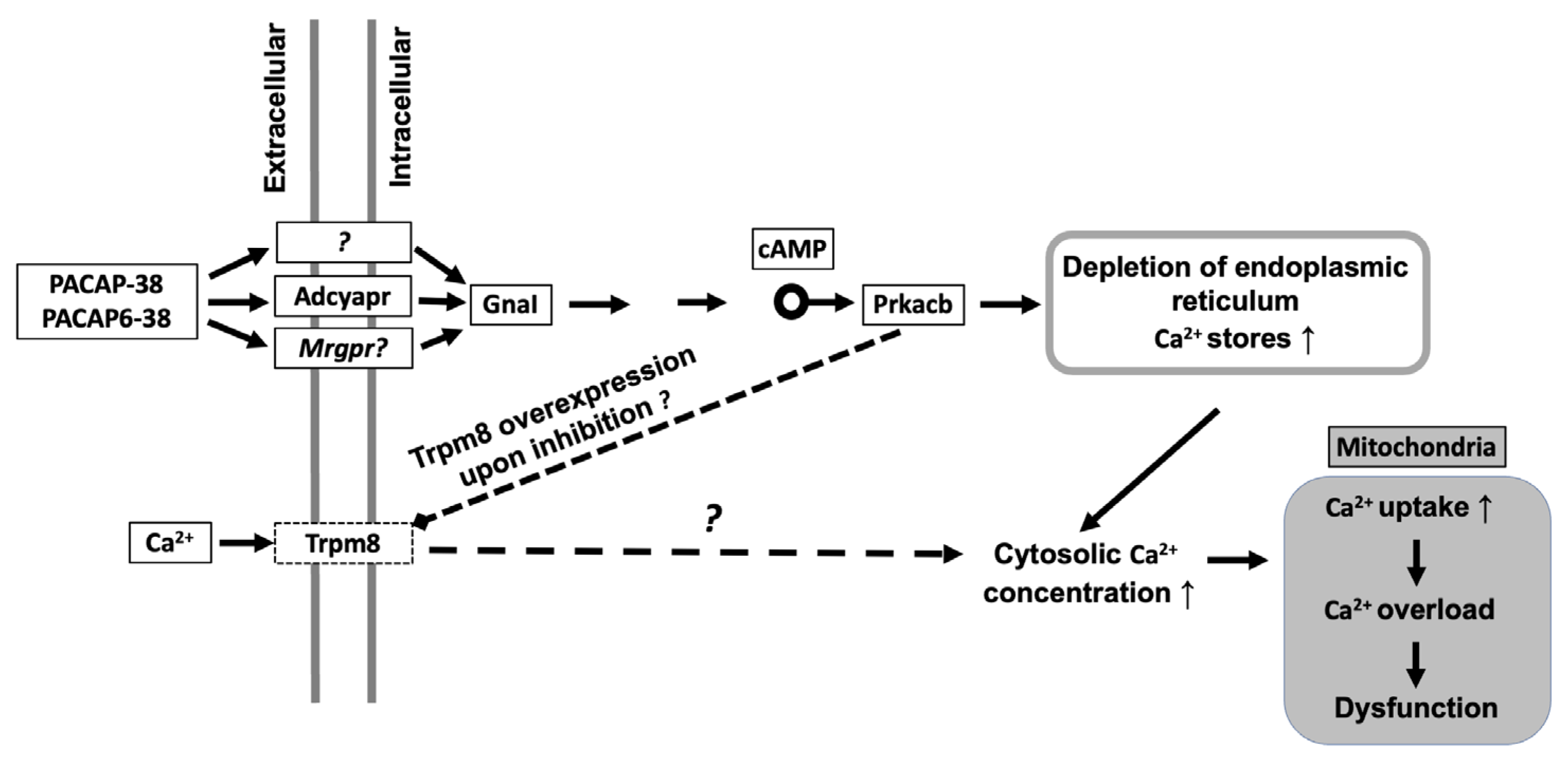
3. Discussion
4. Materials and Methods
4.1. Primary Cultures of TG Neurons
4.2. RNA Isolation and Quality Control
4.3. Illumina Library Preparation and Sequencing
4.4. Bioinformatics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Description |
| Abca3 | ATP binding cassette subfamily A member 3 |
| Adam23 | ADAM metallopeptidase domain 23 |
| Akap7 | A-kinase anchoring protein 7 |
| Ankh | ANKH inorganic pyrophosphate transport regulator |
| Antxr2 | ANTXR cell adhesion molecule 2 |
| Arhgap21 | Rho GTPase activating protein 21 |
| Arhgap23 | Rho GTPase activating protein 23 |
| Arhgap42 | Rho GTPase activating protein 42 |
| Art3 | ADP-ribosyltransferase 3 |
| Atic | 5-aminoimidazole-4-carboxamide ribonucleotide formyltransferase/IMP cyclohydrolase |
| Atp2c1 | ATPase secretory pathway Ca2+ transporting 1 |
| Bmp2k | BMP-2 inducible kinase |
| Cask | calcium/calmodulin dependent serine protein kinase |
| Ccl2 | C-C motif chemokine ligand 2 |
| Cenpb | centromere protein B |
| Clic4 | chloride intracellular channel 4 |
| Cluh | clustered mitochondria homolog |
| Col4a2 | collagen type IV alpha 2 chain |
| Ctsl | cathepsin L |
| Dgkd | diacylglycerol kinase, delta |
| Dnajc13 | DnaJ heat shock protein family (Hsp40) member C13 |
| Dync1h1 | dynein cytoplasmic 1 heavy chain 1 |
| Dync1li1 | dynein cytoplasmic 1 light intermediate chain 1 |
| Epb41l2 | erythrocyte membrane protein band 4.1-like 2 |
| Esyt2 | extended synaptotagmin 2 |
| F2r | coagulation factor II (thrombin) receptor |
| Fbl | fibrillarin |
| Fgfr1 | Fibroblast growth factor receptor 1 |
| Fhl2 | four and a half LIM domains 2 |
| Fpgs | folylpolyglutamate synthase |
| Gas7 | growth arrest specific 7 |
| Gnai1 | G protein subunit alpha i1 |
| Gnal | G protein subunit alpha L |
| Hbb | hemoglobin subunit beta |
| Hmga1 | high mobility group AT-hook 1 |
| Hsp90aa1 | heat shock protein 90 alpha family class A member 1 |
| Hspa9 | heat shock protein family A (Hsp70) member 9 |
| Lonp1 | lon peptidase 1, mitochondrial |
| Map2k4 | mitogen activated protein kinase kinase 4 |
| Map3k20 | mitogen-activated protein kinase kinase kinase 20 |
| Mapk8ip3 | mitogen-activated protein kinase 8 interacting protein 3 |
| Myef2 | myelin expression factor 2 |
| Nav1 | neuron navigator 1 |
| Ncs1 | neuronal calcium sensor 1 |
| Ndfip2 | Nedd4 family interacting protein 2 |
| Ndufab1 | NADH:ubiquinone oxidoreductase subunit AB1 |
| Ndufb6 | NADH:ubiquinone oxidoreductase subunit B6 |
| Opa1 | OPA1, mitochondrial dynamin-like GTPase |
| Osmr | oncostatin M receptor |
| Parp14 | poly (ADP-ribose) polymerase family, member 14 |
| Pcdhgc3 | protocadherin gamma subfamily C, 3 |
| Pitpnb | phosphatidylinositol transfer protein, beta |
| Plin2 | perilipin 2 |
| Plxna4 | plexin A4 |
| Ppp1cb | protein phosphatase 1 catalytic subunit beta |
| Praf2 | PRA1 domain family, member 2 |
| Prkacb | protein kinase cAMP-activated catalytic subunit beta |
| Prkar1a | protein kinase cAMP-dependent type I regulatory subunit alpha |
| Prkar2a | protein kinase cAMP-dependent type II regulatory subunit alpha |
| Prrc2b | proline-rich coiled-coil 2B |
| Psen1 | presenilin 1 |
| Ptprg | protein tyrosine phosphatase, receptor type, G |
| Rcn1 | reticulocalbin 1 |
| Slc20a2 | solute carrier family 20 member 2 |
| Slc25a5 | solute carrier family 25 member 5 |
| Slc30a9 | solute carrier family 30 member 9 |
| Slc3a2 | solute carrier family 3 member 2 |
| Snca | synuclein alpha |
| Tiam1 | TIAM Rac1 associated GEF 1 |
| Tmem128 | transmembrane protein 128 |
| Tmem131 | transmembrane protein 131 |
| Tollip | toll interacting protein |
| Tomm34 | translocase of outer mitochondrial membrane 34 |
| Tomm6 | translocase of outer mitochondrial membrane 6 |
| Tomm70 | translocase of outer mitochondrial membrane 70 |
| Tpm4 | tropomyosin 4 |
| Trio | trio Rho guanine nucleotide exchange factor |
| Trpm8 | transient receptor potential cation channel, subfamily M, member 8 |
| Txlna | taxilin alpha |
| Txn1 | thioredoxin 1 |
| Txndc17 | thioredoxin domain containing 17 |
References
- Somogyvári-Vigh, A.; Reglődi, D. Pituitary Adenylate Cyclase Activating Polypeptide: A Potential Neuroprotective Peptide. Curr. Pharm. Des. 2004, 10, 2861–2889. [Google Scholar] [CrossRef] [PubMed]
- Reglődi, D.; Kiss, P.; Lubics, A.; Tamás, A. Review on the Protective Effects of PACAP in Models of Neurodegenerative Diseases in Vitro and in Vivo. Curr. Pharm. Des. 2011, 17, 962–972. [Google Scholar] [CrossRef] [PubMed]
- Dickson, L.; Finlayson, K. VPAC and PAC Receptors: From Ligands to Function. Pharmacol. Ther. 2009, 121, 294–316. [Google Scholar] [CrossRef] [PubMed]
- Vaudry, D.; Falluel-Morel, A.; Bourgault, S.; Basille, M.; Burel, D.; Wurtz, O.; Fournier, A.; Chow, B.K.C.; Hashimoto, H.; Galas, L.; et al. Pituitary Adenylate Cyclase-Activating Polypeptide and Its Receptors: 20 Years after the Discovery. Pharm. Rev. 2009, 61, 283–357. [Google Scholar] [CrossRef]
- Harmar, A.J.; Fahrenkrug, J.; Gozes, I.; Laburthe, M.; May, V.; Pisegna, J.R.; Vaudry, D.; Vaudry, H.; Waschek, J.A.; Said, S.I. Pharmacology and Functions of Receptors for Vasoactive Intestinal Peptide and Pituitary Adenylate Cyclase-Activating Polypeptide: IUPHAR Review 1. Br. J. Pharmacol. 2012, 166, 4–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatemoto, K.; Nozaki, Y.; Tsuda, R.; Konno, S.; Tomura, K.; Furuno, M.; Ogasawara, H.; Edamura, K.; Takagi, H.; Iwamura, H.; et al. Immunoglobulin E-Independent Activation of Mast Cell Is Mediated by Mrg Receptors. Biochem. Biophys. Res. Commun. 2006, 349, 1322–1328. [Google Scholar] [CrossRef]
- Pedersen, S.H.; la Cour, S.H.; Calloe, K.; Hauser, F.; Olesen, J.; Klaerke, D.A.; Jansen-Olesen, I. PACAP-38 and PACAP(6-38) Degranulate Rat Meningeal Mast Cells via the Orphan MrgB3-Receptor. Front. Cell. Neurosci. 2019, 13, 114. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Han, S.; Zylka, M.J.; Simon, M.I.; Anderson, D.J. A Diverse Family of GPCRs Expressed in Specific Subsets of Nociceptive Sensory Neurons. Cell 2001, 106, 619–632. [Google Scholar] [CrossRef] [Green Version]
- Narita, M.; Dun, S.L.; Dun, N.J.; Tseng, L.F. Hyperalgesia Induced by Pituitary Adenylate Cyclase-Activating Polypeptide in the Mouse Spinal Cord. Eur. J. Pharmacol. 1996, 311, 121–126. [Google Scholar] [CrossRef]
- Shimizu, T.; Katahira, M.; Sugawara, H.; Inoue, K.; Miyata, A. Diverse Effects of Intrathecal Pituitary Adenylate Cyclase-Activating Polypeptide on Nociceptive Transmission in Mice Spinal Cord. Regul. Pept. 2004, 123, 117–122. [Google Scholar] [CrossRef]
- Yamamoto, T.; Tatsuno, I. Antinociceptive Effect of Intrathecally Administered Pituitary Adenylate Cyclase Activating Polypeptide (PACAP) on the Rat Formalin Test. Neurosci. Lett. 1995, 184, 32–35. [Google Scholar] [CrossRef]
- Zhang, Y.; Malmberg, A.B.; Sjölund, B.; Yaksh, T.L. The Effect of Pituitary Adenylate Cyclase Activating Peptide (PACAP) on the Nociceptive Formalin Test. Neurosci. Lett. 1996, 207, 187–190. [Google Scholar] [CrossRef]
- Sándor, K.; Bölcskei, K.; McDougall, J.J.; Schuelert, N.; Reglődi, D.; Elekes, K.; Pethő, G.; Pintér, E.; Szolcsányi, J.; Helyes, Z. Divergent Peripheral Effects of Pituitary Adenylate Cyclase-Activating Polypeptide-38 on Nociception in Rats and Mice. Pain 2009, 141, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Mabuchi, T.; Shintani, N.; Matsumura, S.; Okuda-Ashitaka, E.; Hashimoto, H.; Muratani, T.; Minami, T.; Baba, A.; Ito, S. Pituitary Adenylate Cyclase-Activating Polypeptide Is Required for the Development of Spinal Sensitization and Induction of Neuropathic Pain. J. Neurosci. 2004, 24, 7283–7291. [Google Scholar] [CrossRef]
- Sándor, K.; Kormos, V.; Botz, B.; Imreh, A.; Bölcskei, K.; Gaszner, B.; Markovics, A.; Szolcsányi, J.; Shintani, N.; Hashimoto, H.; et al. Impaired Nocifensive Behaviours and Mechanical Hyperalgesia, but Enhanced Thermal Allodynia in Pituitary Adenylate Cyclase-Activating Polypeptide Deficient Mice. Neuropeptides 2010, 44, 363–371. [Google Scholar] [CrossRef]
- Botz, B.; Imreh, A.; Sándor, K.; Elekes, K.; Szolcsányi, J.; Reglődi, D.; Quinn, J.P.; Stewart, J.; Zimmer, A.; Hashimoto, H.; et al. Role of Pituitary Adenylate-Cyclase Activating Polypeptide and Tac1 Gene Derived Tachykinins in Sensory, Motor and Vascular Functions under Normal and Neuropathic Conditions. Peptides 2013, 43, 105–112. [Google Scholar] [CrossRef]
- Botz, B.; Bölcskei, K.; Kereskai, L.; Kovács, M.; Németh, T.; Szigeti, K.; Horváth, I.; Máthé, D.; Kovács, N.; Hashimoto, H.; et al. Differential Regulatory Role of Pituitary Adenylate Cyclase-Activating Polypeptide in the Serum-Transfer Arthritis Model: Complex Regulatory Role of PACAP in Arthritis. Arthritis Rheumatol. 2014, 66, 2739–2750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuka, B.; Helyes, Z.; Markovics, A.; Bagoly, T.; Szolcsányi, J.; Szabó, N.; Tóth, E.; Kincses, Z.T.; Vécsei, L.; Tajti, J. Alterations in PACAP-38-like Immunoreactivity in the Plasma during Ictal and Interictal Periods of Migraine Patients. Cephalalgia 2013, 33, 1085–1095. [Google Scholar] [CrossRef] [Green Version]
- Schytz, H.W.; Birk, S.; Wienecke, T.; Kruuse, C.; Olesen, J.; Ashina, M. PACAP38 Induces Migraine-like Attacks in Patients with Migraine without Aura. Brain 2009, 132, 16–25. [Google Scholar] [CrossRef] [Green Version]
- Amin, F.M.; Asghar, M.S.; Guo, S.; Hougaard, A.; Hansen, A.E.; Schytz, H.W.; van der Geest, R.J.; de Koning, P.J.; Larsson, H.B.; Olesen, J.; et al. Headache and Prolonged Dilatation of the Middle Meningeal Artery by PACAP38 in Healthy Volunteers. Cephalalgia 2012, 32, 140–149. [Google Scholar] [CrossRef]
- Lukacs, M.; Tajti, J.; Fulop, F.; Toldi, J.; Edvinsson, L.; Vecsei, L. Migraine, Neurogenic Inflammation, Drug Development—Pharmacochemical Aspects. Curr. Med. Chem. 2017, 24, 3649–3665. [Google Scholar] [CrossRef] [PubMed]
- Baun, M.; Pedersen, M.H.F.; Olesen, J.; Jansen-Olesen, I. Dural Mast Cell Degranulation Is a Putative Mechanism for Headache Induced by PACAP-38. Cephalalgia 2012, 32, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Okragly, A.J.; Morin, S.M.; DeRosa, D.; Martin, A.P.; Johnson, K.W.; Johnson, M.P.; Benschop, R.J. Human Mast Cells Release the Migraine-Inducing Factor Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP). Cephalalgia 2018, 38, 1564–1574. [Google Scholar] [CrossRef] [PubMed]
- Tajti, J.; Tuka, B.; Botz, B.; Helyes, Z.; Vecsei, L. Role of Pituitary Adenylate Cyclase-Activating Polypeptide in Nociception and Migraine. CNS Neurol. Disord. Drug Targets 2015, 14, 540–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koyuncu Irmak, D.; Kilinc, E.; Tore, F. Shared Fate of Meningeal Mast Cells and Sensory Neurons in Migraine. Front. Cell. Neurosci. 2019, 13, 136. [Google Scholar] [CrossRef] [Green Version]
- Moller, K.; Zhang, Y.-Z.; Håkanson, R.; Luts, A.; Sjölund, B.; Uddman, R.; Sundler, F. Pituitary Adenylate Cyclase Activating Peptide Is a Sensory Neuropeptide: Immunocytochemical and Immunochemical Evidence. Neuroscience 1993, 57, 725–732. [Google Scholar] [CrossRef]
- Chaudhary, P.; Baumann, T.K. Expression of VPAC2 Receptor and PAC1 Receptor Splice Variants in the Trigeminal Ganglion of the Adult Rat. Brain Res. Mol. Brain Res. 2002, 104, 137–142. [Google Scholar] [CrossRef]
- Uddman, R.; Tajti, J.; Hou, M.; Sundler, F.; Edvinsson, L. Neuropeptide Expression in the Human Trigeminal Nucleus Caudalis and in the Cervical Spinal Cord C1 and C2. Cephalalgia 2002, 22, 112–116. [Google Scholar] [CrossRef] [Green Version]
- Markovics, A.; Kormos, V.; Gaszner, B.; Lashgarara, A.; Szoke, E.; Sandor, K.; Szabadfi, K.; Tuka, B.; Tajti, J.; Szolcsanyi, J.; et al. Pituitary Adenylate Cyclase-Activating Polypeptide Plays a Key Role in Nitroglycerol-Induced Trigeminovascular Activation in Mice. Neurobiol. Dis. 2012, 45, 633–644. [Google Scholar] [CrossRef]
- Tuka, B.; Helyes, Z.; Markovics, A.; Bagoly, T.; Németh, J.; Márk, L.; Brubel, R.; Reglődi, D.; Párdutz, Á.; Szolcsányi, J.; et al. Peripheral and Central Alterations of Pituitary Adenylate Cyclase Activating Polypeptide-like Immunoreactivity in the Rat in Response to Activation of the Trigeminovascular System. Peptides 2012, 33, 307–316. [Google Scholar] [CrossRef]
- Rahmann, A.; Wienecke, T.; Hansen, J.M.; Fahrenkrug, J.; Olesen, J.; Ashina, M. Vasoactive Intestinal Peptide Causes Marked Cephalic Vasodilation, but Does Not Induce Migraine. Cephalalgia 2008, 28, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Jansen-Olesen, I.; Baun, M.; Amrutkar, D.V.; Ramachandran, R.; Christophersen, D.V.; Olesen, J. PACAP-38 but Not VIP Induces Release of CGRP from Trigeminal Nucleus Caudalis via a Receptor Distinct from the PAC1 Receptor. Neuropeptides 2014, 48, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Solinski, H.J.; Gudermann, T.; Breit, A. Pharmacology and Signaling of MAS-Related G Protein-Coupled Receptors. Pharm. Rev. 2014, 66, 570–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bader, M.; Alenina, N.; Andrade-Navarro, M.A.; Santos, R.A. Mas and Its Related G Protein–Coupled Receptors, Mrgprs. Pharm. Rev. 2014, 66, 1080–1105. [Google Scholar] [CrossRef] [PubMed]
- Németh, J.; Reglődi, D.; Pozsgai, G.; Szabó, Á.; Elekes, K.; Pintér, E.; Szolcsányi, J.; Helyes, Z. Effect of Pituitary Adenylate Cyclase Activating Polypeptide-38 on Sensory Neuropeptide Release and Neurogenic Inflammation in Rats and Mice. Neuroscience 2006, 143, 223–230. [Google Scholar] [CrossRef]
- Sághy, É.; Payrits, M.; Helyes, Z.; Reglődi, D.; Bánki, E.; Tóth, G.; Couvineau, A.; Szőke, É. Stimulatory Effect of Pituitary Adenylate Cyclase-Activating Polypeptide 6-38, M65 and Vasoactive Intestinal Polypeptide 6–28 on Trigeminal Sensory Neurons. Neuroscience 2015, 308, 144–156. [Google Scholar] [CrossRef]
- Walker, C.S.; Sundrum, T.; Hay, D.L. PACAP Receptor Pharmacology and Agonist Bias: Analysis in Primary Neurons and Glia from the Trigeminal Ganglia and Transfected Cells: PACAP Pharmacology in the Trigeminal Ganglia. Br. J. Pharmacol. 2014, 171, 1521–1533. [Google Scholar] [CrossRef] [Green Version]
- Reglődi, D.; Börzsei, R.; Bagoly, T.; Boronkai, A.; Rácz, B.; Tamás, A.; Kiss, P.; Horváth, G.; Brubel, R.; Németh, J.; et al. Agonistic Behavior of PACAP6-38 on Sensory Nerve Terminals and Cytotrophoblast Cells. J. Mol. Neurosci. 2008, 36, 270–278. [Google Scholar] [CrossRef]
- Ichinose, M.; Asai, M.; Imai, K.; Sawada, M. Enhancement of Phagocytosis in Mouse Macrophages by Pituitary Adenylate Cyclase Activating Polypeptide (PACAP) and Related Peptides. Immunopharmacology 1995, 30, 217–224. [Google Scholar] [CrossRef]
- Juhász, T.; Matta, C.; Katona, É.; Somogyi, C.; Takács, R.; Gergely, P.; Csernoch, L.; Panyi, G.; Tóth, G.; Reglődi, D.; et al. Pituitary Adenylate Cyclase Activating Polypeptide (PACAP) Signalling Exerts Chondrogenesis Promoting and Protecting Effects: Implication of Calcineurin as a Downstream Target. PLoS ONE 2014, 9, e91541. [Google Scholar] [CrossRef] [Green Version]
- McNeil, B.D.; Pundir, P.; Meeker, S.; Han, L.; Undem, B.J.; Kulka, M.; Dong, X. Identification of a Mast-Cell-Specific Receptor Crucial for Pseudo-Allergic Drug Reactions. Nature 2015, 519, 237–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, D.P. The Role of Mrgprs in Pain. Neurosci. Lett. 2021, 744, 135544. [Google Scholar] [CrossRef] [PubMed]
- Vrontou, S.; Wong, A.M.; Rau, K.K.; Koerber, H.R.; Anderson, D.J. Genetic Identification of C Fibres That Detect Massage-like Stroking of Hairy Skin in Vivo. Nature 2013, 493, 669–673. [Google Scholar] [CrossRef] [PubMed]
- Hucho, T.; Levine, J.D. Signaling Pathways in Sensitization: Toward a Nociceptor Cell Biology. Neuron 2007, 55, 365–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.-Q.; Liu, D.-Q.; Chen, S.-P.; Sun, J.; Zhou, X.-R.; Luo, F.; Tian, Y.-K.; Ye, D.-W. Cellular and Molecular Mechanisms of Calcium/Calmodulin-Dependent Protein Kinase II in Chronic Pain. J. Pharm. Exp. Ther. 2017, 363, 176–183. [Google Scholar] [CrossRef] [Green Version]
- Bhave, G.; Zhu, W.; Wang, H.; Brasier, D.J.; Oxford, G.S.; Gereau, R.W. CAMP-Dependent Protein Kinase Regulates Desensitization of the Capsaicin Receptor (VR1) by Direct Phosphorylation. Neuron 2002, 35, 721–731. [Google Scholar] [CrossRef] [Green Version]
- Varga, A.; Bölcskei, K.; Szöke, é.; Almási, R.; Czéh, G.; Szolcsányi, J.; Pethö, G. Relative Roles of Protein Kinase A and Protein Kinase C in Modulation of Transient Receptor Potential Vanilloid Type 1 Receptor Responsiveness in Rat Sensory Neurons in Vitro and Peripheral Nociceptors in Vivo. Neuroscience 2006, 140, 645–657. [Google Scholar] [CrossRef]
- Gold, M.S.; Levine, J.D.; Correa, A.M. Modulation of TTX-R INa by PKC and PKA and Their Role in PGE2-Induced Sensitization of Rat Sensory Neurons In Vitro. J. Neurosci. 1998, 18, 10345–10355. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, E.M.; Okuse, K.; Wood, J.N.; Dolphin, A.C.; Moss, S.J. CAMP-Dependent Phosphorylation of the Tetrodotoxin-Resistant Voltage-Dependent Sodium Channel SNS. J. Physiol. 1999, 516, 433–446. [Google Scholar] [CrossRef]
- Fuentes, E.; Araya-Maturana, R.; Urra, F.A. Regulation of Mitochondrial Function as a Promising Target in Platelet Activation-Related Diseases. Free Radic. Biol. Med. 2019, 136, 172–182. [Google Scholar] [CrossRef]
- Gross, E.C.; Lisicki, M.; Fischer, D.; Sándor, P.S.; Schoenen, J. The Metabolic Face of Migraine—From Pathophysiology to Treatment. Nat. Rev. Neurol. 2019, 15, 627–643. [Google Scholar] [CrossRef] [PubMed]
- Aczél, T.; Körtési, T.; Kun, J.; Urbán, P.; Bauer, W.; Herczeg, R.; Farkas, R.; Kovács, K.; Vásárhelyi, B.; Karvaly, G.B.; et al. Identification of Disease- and Headache-Specific Mediators and Pathways in Migraine Using Blood Transcriptomic and Metabolomic Analysis. J. Headache Pain 2021, 22, 117. [Google Scholar] [CrossRef] [PubMed]
- Meeus, M.; Nijs, J.; Hermans, L.; Goubert, D.; Calders, P. The Role of Mitochondrial Dysfunctions Due to Oxidative and Nitrosative Stress in the Chronic Pain or Chronic Fatigue Syndromes and Fibromyalgia Patients: Peripheral and Central Mechanisms as Therapeutic Targets? Expert Opin. Ther. Targets 2013, 17, 1081–1089. [Google Scholar] [CrossRef] [PubMed]
- van den Ameele, J.; Fuge, J.; Pitceathly, R.D.S.; Berry, S.; McIntyre, Z.; Hanna, M.G.; Lee, M.; Chinnery, P.F. Chronic Pain Is Common in Mitochondrial Disease. Neuromuscul. Disord. 2020, 30, 413–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yousuf, M.S.; Maguire, A.D.; Simmen, T.; Kerr, B.J. Endoplasmic Reticulum–Mitochondria Interplay in Chronic Pain: The Calcium Connection. Mol. Pain 2020, 16, 1744806920946889. [Google Scholar] [CrossRef] [PubMed]
- Salvemini, D.; Little, J.W.; Doyle, T.; Neumann, W.L. Roles of Reactive Oxygen and Nitrogen Species in Pain. Free Radic. Biol. Med. 2011, 51, 951. [Google Scholar] [CrossRef] [Green Version]
- Nesuashvili, L.; Hadley, S.H.; Bahia, P.K.; Taylor-Clark, T.E. Sensory Nerve Terminal Mitochondrial Dysfunction Activates Airway Sensory Nerves via Transient Receptor Potential (TRP) Channels. Mol. Pharmacol. 2013, 83, 1007–1019. [Google Scholar] [CrossRef] [Green Version]
- Hadley, S.H.; Bahia, P.K.; Taylor-Clark, T.E. Sensory Nerve Terminal Mitochondrial Dysfunction Induces Hyperexcitability in Airway Nociceptors via Protein Kinase C. Mol. Pharm. 2014, 85, 839–848. [Google Scholar] [CrossRef] [Green Version]
- McKemy, D.D.; Neuhausser, W.M.; Julius, D. Identification of a Cold Receptor Reveals a General Role for TRP Channels in Thermosensation. Nature 2002, 416, 52–58. [Google Scholar] [CrossRef]
- Peier, A.M.; Moqrich, A.; Hergarden, A.C.; Reeve, A.J.; Andersson, D.A.; Story, G.M.; Earley, T.J.; Dragoni, I.; McIntyre, P.; Bevan, S.; et al. A TRP Channel That Senses Cold Stimuli and Menthol. Cell 2002, 108, 705–715. [Google Scholar] [CrossRef] [Green Version]
- Okazawa, M.; Inoue, W.; Hori, A.; Hosokawa, H.; Matsumura, K.; Kobayashi, S. Noxious Heat Receptors Present in Cold-Sensory Cells in Rats. Neurosci. Lett. 2004, 359, 33–36. [Google Scholar] [CrossRef] [PubMed]
- Abe, J.; Hosokawa, H.; Okazawa, M.; Kandachi, M.; Sawada, Y.; Yamanaka, K.; Matsumura, K.; Kobayashi, S. TRPM8 Protein Localization in Trigeminal Ganglion and Taste Papillae. Brain Res. Mol. Brain Res. 2005, 136, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Sarria, I.; Gu, J. Menthol Response and Adaptation in Nociceptive-like and Nonnociceptive-like Neurons: Role of Protein Kinases. Mol. Pain 2010, 6, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bautista, D.M.; Siemens, J.; Glazer, J.M.; Tsuruda, P.R.; Basbaum, A.I.; Stucky, C.L.; Jordt, S.-E.; Julius, D. The Menthol Receptor TRPM8 Is the Principal Detector of Environmental Cold. Nature 2007, 448, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Dhaka, A.; Murray, A.N.; Mathur, J.; Earley, T.J.; Petrus, M.J.; Patapoutian, A. TRPM8 Is Required for Cold Sensation in Mice. Neuron 2007, 54, 371–378. [Google Scholar] [CrossRef] [Green Version]
- Knowlton, W.M.; Bifolck-Fisher, A.; Bautista, D.M.; McKemy, D.D. TRPM8, but Not TRPA1, Is Required for Neural and Behavioral Responses to Acute Noxious Cold Temperatures and Cold-Mimetics in Vivo. Pain 2010, 150, 340–350. [Google Scholar] [CrossRef] [Green Version]
- Colburn, R.W.; Lubin, M.L.; Stone, D.J.; Wang, Y.; Lawrence, D.; D’Andrea, M.R.; Brandt, M.R.; Liu, Y.; Flores, C.M.; Qin, N. Attenuated Cold Sensitivity in TRPM8 Null Mice. Neuron 2007, 54, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Chasman, D.I.; Schürks, M.; Anttila, V.; de Vries, B.; Schminke, U.; Launer, L.J.; Terwindt, G.M.; van den Maagdenberg, A.M.J.M.; Fendrich, K.; Völzke, H.; et al. Genome-Wide Association Study Reveals Three Susceptibility Loci for Common Migraine in the General Population. Nat. Genet. 2011, 43, 695–698. [Google Scholar] [CrossRef] [Green Version]
- Freilinger, T.; Anttila, V.; de Vries, B.; Malik, R.; Kallela, M.; Terwindt, G.M.; Pozo-Rosich, P.; Winsvold, B.; Nyholt, D.R.; van Oosterhout, W.P.J.; et al. Genome-Wide Association Analysis Identifies Susceptibility Loci for Migraine without Aura. Nat. Genet. 2012, 44, 777–782. [Google Scholar] [CrossRef]
- Gormley, P.; Anttila, V.; Winsvold, B.S.; Palta, P.; Esko, T.; Pers, T.H.; Farh, K.-H.; Cuenca-Leon, E.; Muona, M.; Furlotte, N.A.; et al. Meta-Analysis of 375,000 Individuals Identifies 38 Susceptibility Loci for Migraine. Nat. Genet. 2016, 48, 856–866. [Google Scholar] [CrossRef] [Green Version]
- Borhani Haghighi, A.; Motazedian, S.; Rezaii, R.; Mohammadi, F.; Salarian, L.; Pourmokhtari, M.; Khodaei, S.; Vossoughi, M.; Miri, R. Cutaneous Application of Menthol 10% Solution as an Abortive Treatment of Migraine without Aura: A Randomised, Double-Blind, Placebo-Controlled, Crossed-over Study. Int. J. Clin. Pract. 2010, 64, 451–456. [Google Scholar] [CrossRef]
- Kayama, Y.; Shibata, M.; Takizawa, T.; Ibata, K.; Shimizu, T.; Ebine, T.; Toriumi, H.; Yuzaki, M.; Suzuki, N. Functional Interactions between Transient Receptor Potential M8 and Transient Receptor Potential V1 in the Trigeminal System: Relevance to Migraine Pathophysiology. Cephalalgia 2018, 38, 833–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bidaux, G.; Borowiec, A.; Gordienko, D.; Beck, B.; Shapovalov, G.G.; Lemonnier, L.; Flourakis, M.; Vandenberghe, M.; Slomianny, C.; Dewailly, E.; et al. Epidermal TRPM8 Channel Isoform Controls the Balance between Keratinocyte Proliferation and Differentiation in a Cold-Dependent Manner. Proc. Natl. Acad. Sci. USA 2015, 112, E3345–E3354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabnis, A.S.; Shadid, M.; Yost, G.S.; Reilly, C.A. Human Lung Epithelial Cells Express a Functional Cold-Sensing TRPM8 Variant. Am. J. Respir. Cell Mol. Biol. 2008, 39, 466. [Google Scholar] [CrossRef] [Green Version]
- Thebault, S.; Lemonnier, L.; Bidaux, G.; Flourakis, M.; Bavencoffe, A.; Gordienko, D.; Roudbaraki, M.; Delcourt, P.; Panchin, Y.; Shuba, Y.; et al. Novel Role of Cold/Menthol-Sensitive Transient Receptor Potential Melastatine Family Member 8 (TRPM8) in the Activation of Store-Operated Channels in LNCaP Human Prostate Cancer Epithelial Cells. J. Biol. Chem. 2005, 280, 39423–39435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, S.; Wang, B.; Lin, S.; Zhang, H.; Li, Y.; Wei, X.; Cui, Y.; Wei, X.; Lu, Z.; Gao, P.; et al. Activation of Transient Receptor Potential Melastatin Subtype 8 Attenuates Cold-Induced Hypertension Through Ameliorating Vascular Mitochondrial Dysfunction. JAHA 2017, 6, e005495. [Google Scholar] [CrossRef]
- Kárai, L.J.; Russell, J.T.; Iadarola, M.J.; Oláh, Z. Vanilloid Receptor 1 Regulates Multiple Calcium Compartments and Contributes to Ca2+-Induced Ca2+ Release in Sensory Neurons. J. Biol. Chem. 2004, 279, 16377–16387. [Google Scholar] [CrossRef] [Green Version]
- Gallego-Sandín, S.; Rodríguez-García, A.; Alonso, M.T.; García-Sancho, J. The Endoplasmic Reticulum of Dorsal Root Ganglion Neurons Contains Functional TRPV1 Channels. J. Biol. Chem. 2009, 284, 32591. [Google Scholar] [CrossRef] [Green Version]
- Szőke, É.; Balla, Z.; Csernoch, L.; Czéh, G.; Szolcsányi, J. Interacting Effects of Capsaicin and Anandamide on Intracellular Calcium in Sensory Neurones. NeuroReport 2000, 11, 1949–1952. [Google Scholar] [CrossRef]
- Aczél, T.; Kun, J.; Szőke, É.; Rauch, T.; Junttila, S.; Gyenesei, A.; Bölcskei, K.; Helyes, Z. Transcriptional Alterations in the Trigeminal Ganglia, Nucleus and Peripheral Blood Mononuclear Cells in a Rat Orofacial Pain Model. Front. Mol. Neurosci. 2018, 11, 219. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python Framework to Work with High-Throughput Sequencing Data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Law, C.W.; Chen, Y.; Shi, W.; Smyth, G.K. Voom: Precision Weights Unlock Linear Model Analysis Tools for RNA-Seq Read Counts. Genome Biol. 2014, 15, R29. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma Powers Differential Expression Analyses for RNA-Sequencing and Microarray Studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Luo, W.; Brouwer, C. Pathview: An R/Bioconductor Package for Pathway-Based Data Integration and Visualization. Bioinformatics 2013, 29, 1830–1831. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takács-Lovász, K.; Kun, J.; Aczél, T.; Urbán, P.; Gyenesei, A.; Bölcskei, K.; Szőke, É.; Helyes, Z. PACAP-38 Induces Transcriptomic Changes in Rat Trigeminal Ganglion Cells Related to Neuroinflammation and Altered Mitochondrial Function Presumably via PAC1/VPAC2 Receptor-Independent Mechanism. Int. J. Mol. Sci. 2022, 23, 2120. https://doi.org/10.3390/ijms23042120
Takács-Lovász K, Kun J, Aczél T, Urbán P, Gyenesei A, Bölcskei K, Szőke É, Helyes Z. PACAP-38 Induces Transcriptomic Changes in Rat Trigeminal Ganglion Cells Related to Neuroinflammation and Altered Mitochondrial Function Presumably via PAC1/VPAC2 Receptor-Independent Mechanism. International Journal of Molecular Sciences. 2022; 23(4):2120. https://doi.org/10.3390/ijms23042120
Chicago/Turabian StyleTakács-Lovász, Krisztina, József Kun, Timea Aczél, Péter Urbán, Attila Gyenesei, Kata Bölcskei, Éva Szőke, and Zsuzsanna Helyes. 2022. "PACAP-38 Induces Transcriptomic Changes in Rat Trigeminal Ganglion Cells Related to Neuroinflammation and Altered Mitochondrial Function Presumably via PAC1/VPAC2 Receptor-Independent Mechanism" International Journal of Molecular Sciences 23, no. 4: 2120. https://doi.org/10.3390/ijms23042120
APA StyleTakács-Lovász, K., Kun, J., Aczél, T., Urbán, P., Gyenesei, A., Bölcskei, K., Szőke, É., & Helyes, Z. (2022). PACAP-38 Induces Transcriptomic Changes in Rat Trigeminal Ganglion Cells Related to Neuroinflammation and Altered Mitochondrial Function Presumably via PAC1/VPAC2 Receptor-Independent Mechanism. International Journal of Molecular Sciences, 23(4), 2120. https://doi.org/10.3390/ijms23042120