
Omega-3 Polyunsaturated Fatty Acids—Vascular and Cardiac Effects on the Cellular and Molecular Level (Narrative Review)


Abstract
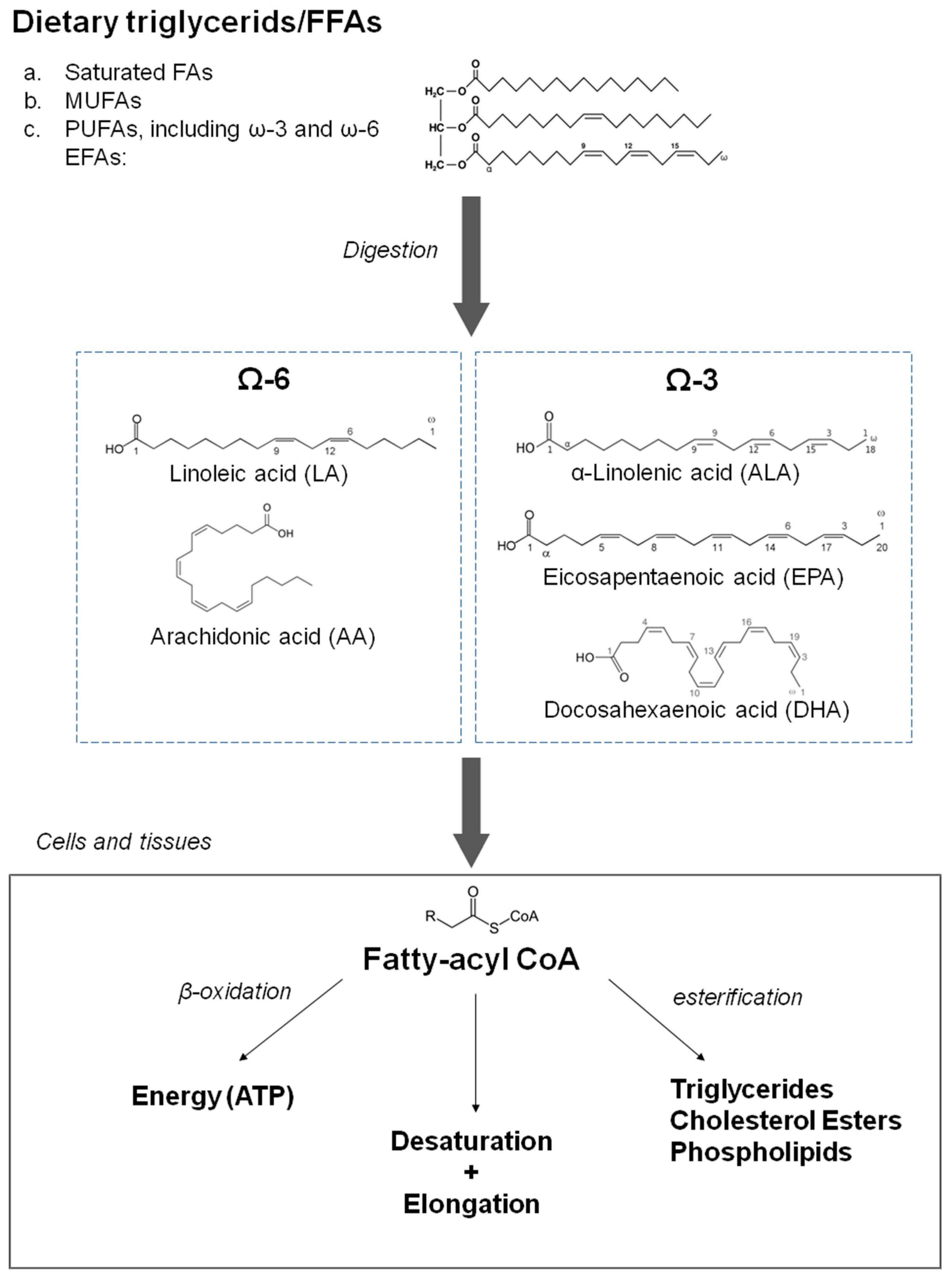
:1. Introduction
2. The Effects of n-3 PUFAs on Lipid Metabolism
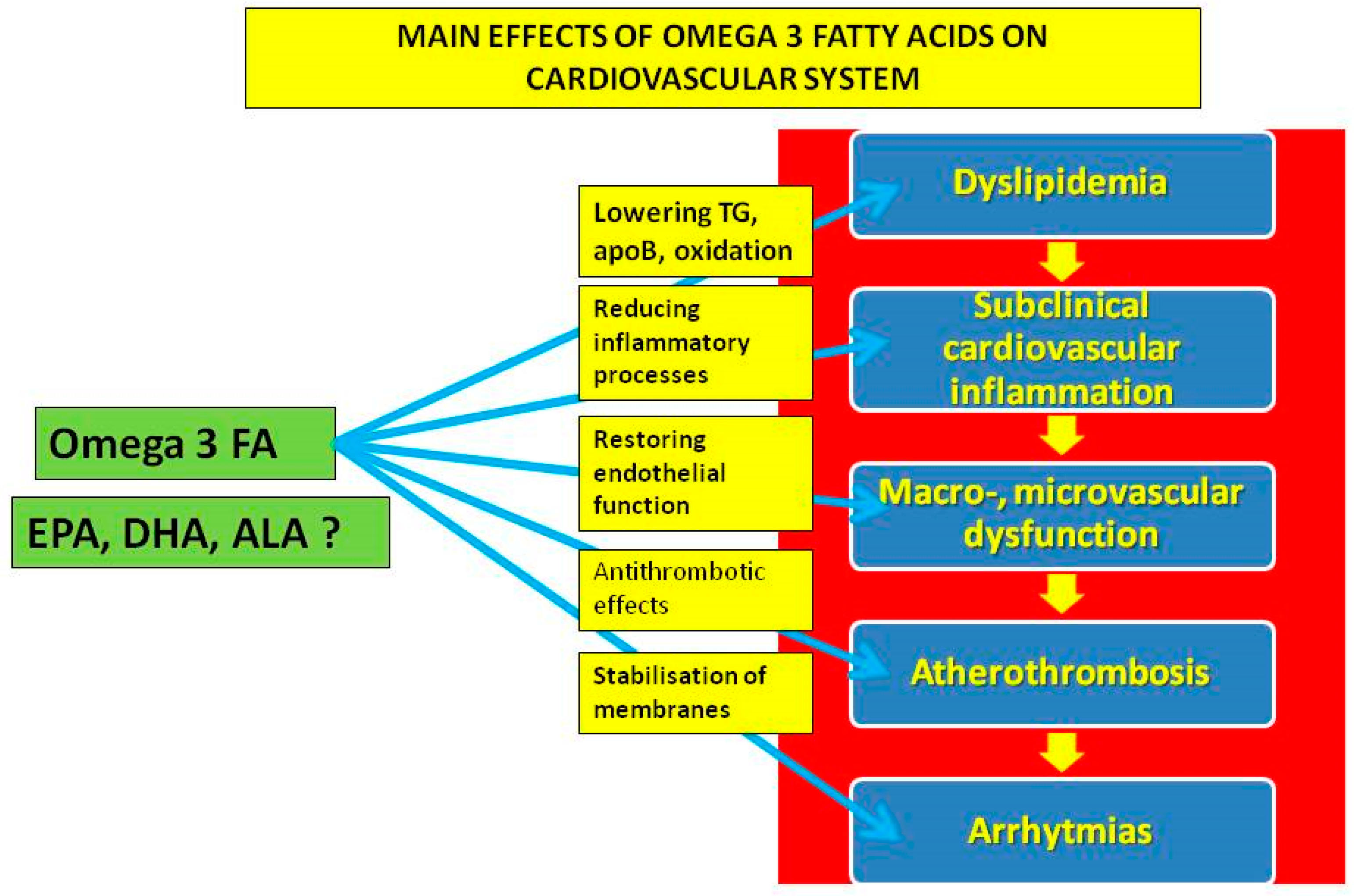
3. The Effect of n-3 PUFAs on Inflammatory and Prothrombogenic Processes
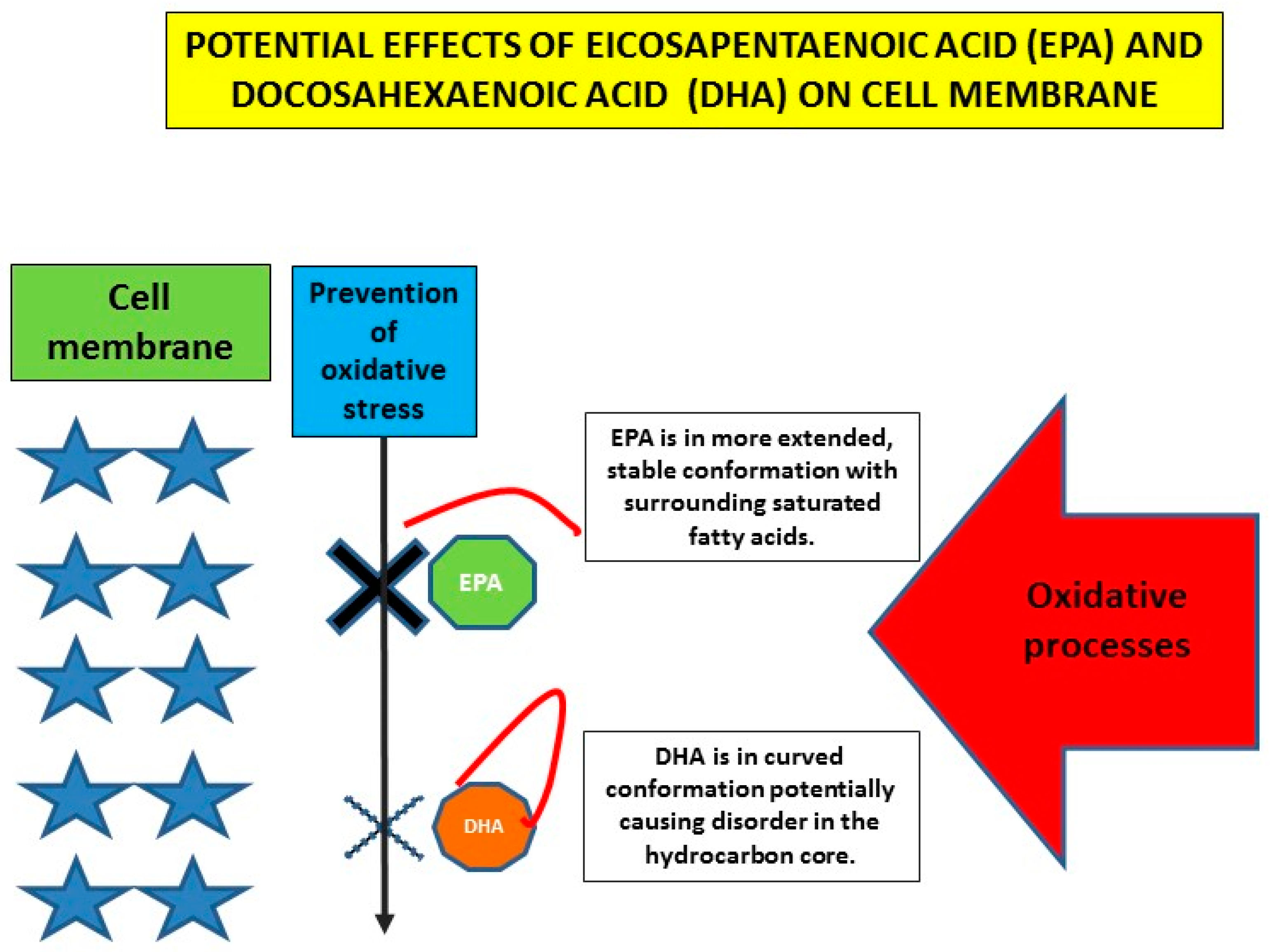
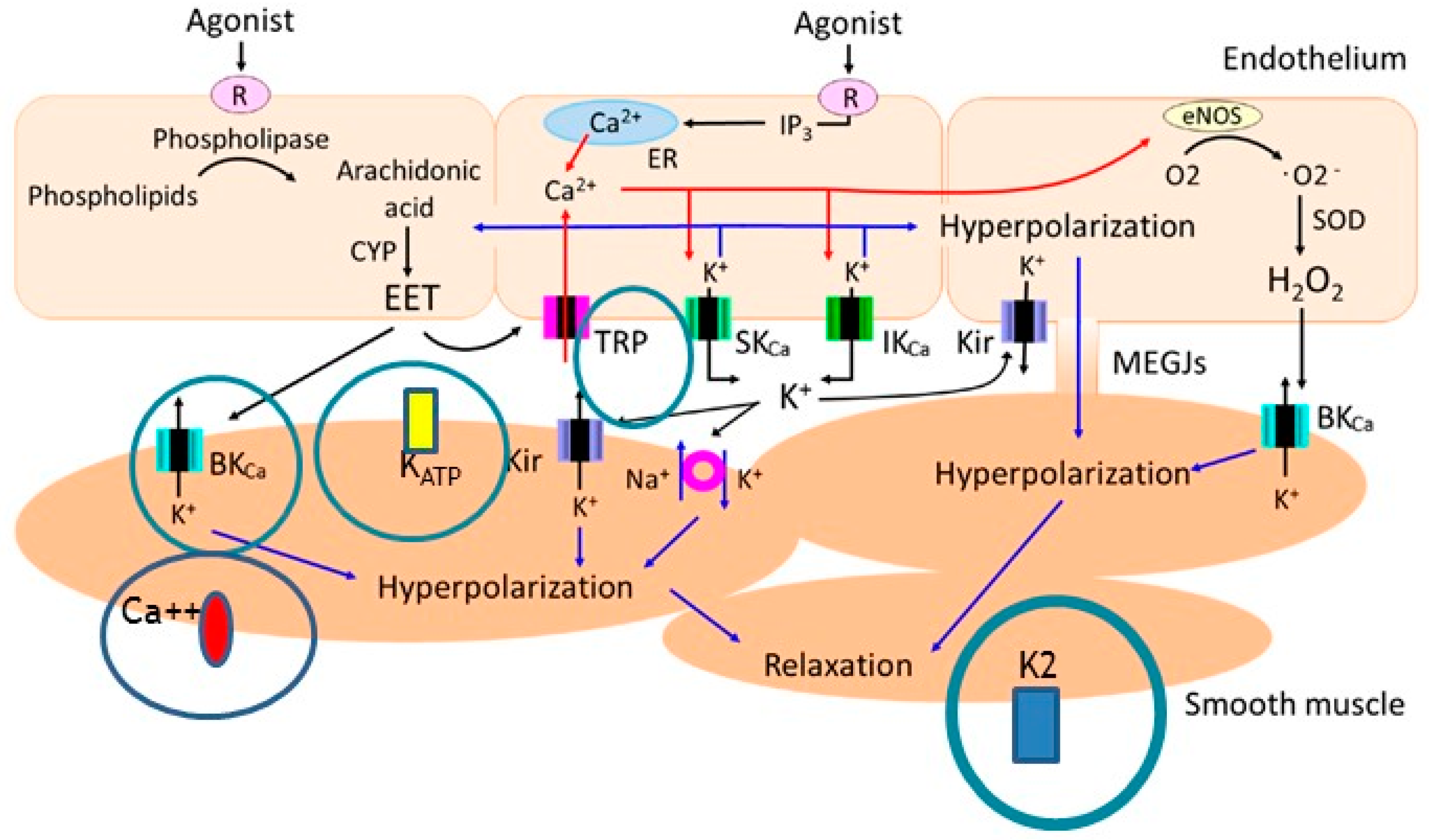
4. Cardiovascular Effects of n-3 PUFAs on (Sub)Cellular Level
5. Cardiovascular Effects of n-3 PUFAs in Studies of Subclinical Vascular Impairment
6. Summary
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| ApoB | Apolipoprotein B |
| ApoE | Apolipoprotein E |
| CVSMCs | Cerebrovascular smooth muscle cells |
| CCL2 | C–C motif chemokine ligand 2 |
| DHA | Docosahexaenoic acid |
| eNOS | Endothelial nitric oxide synthase |
| EPA | Eicosapentaenoic acid |
| Ffar4 | Free fatty acid receptor 4 |
| HFpEF | Heart failure with preserved ejection fraction |
| hsCRP | C-reactive protein measured by highly sensitive method |
| IL | Interleukin |
| MCP-1 | Macrophage chemoattractant protein 1 |
| mRNA | Messenger ribonucleic acid |
| n-3 PUFAs | Omega-3 polyunsaturated fatty acids |
| RvE1 | Resolvin E1 |
| TGFβ-1 | Transforming growth factor β-1 |
| TNF-α | Tumor necrosis factor-α |
| TRPV4 | Transient receptor potential vanilloid 4 |
| VCAM1 | Vascular cells adhesion molecules 1 |
| 11,12-EET | 11,12-Epoxyeicosatrienoic acid |
| 17,18-EEQ | Epoxyeicosatetraenoic acid |
References
- Patel, D.; Busch, R. Omega-3 Fatty Acids and Cardiovascular Disease: A Narrative Review for Pharmacists. J. Cardiovasc. Pharmacol. Ther. 2021, 26, 524–532. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, K.; Alfaddagh, A.; Stone, N.J.; Blumenthal, R.S. Update on the omega-3 fatty acid trial landscape: A narrative review with implications for primary prevention. J. Clin. Lipidol. 2021, 15, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Kotlega, D.; Zembron-Lacny, A.; Golab-Janowska, M.; Nowacki, P.; Szczuko, M. The Association of Free Fatty Acids and Eicosanoids with the Severity of Depressive Symptoms in Stroke Patients. Int. J. Mol. Sci. 2020, 21, 5220. [Google Scholar] [CrossRef] [PubMed]
- Ciappolino, V.; Mazzocchi, A.; Enrico, P.; Syrén, M.L.; Delvecchio, G.; Agostoni, C.; Brambilla, P. N-3 Polyunsatured Fatty Acids in Menopausal Transition: A Systematic Review of Depressive and Cognitive Disorders with Accompanying Vasomotor Symptoms. Int. J. Mol. Sci. 2018, 19, 1849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Hedström, D.; de la Fuente-Fernández, M.; Priego, T.; Martín, A.I.; Amor, S.; López-Calderón, A.; Inarejos-García, A.M.; García-Villalón, Á.L.; Granado, M. Addition of Olive Leaf Extract to a Mixture of Algae and Extra Virgin Olive Oils Decreases Fatty Acid Oxidation and Synergically Attenuates Age-Induced Hypertension, Sarcopenia and Insulin Resistance in Rats. Antioxidants 2021, 10, 1066. [Google Scholar] [CrossRef]
- Aung, T.; Halsey, J.; Kromhout, D.; Gerstein, H.C.; Marchioli, R.; Tavazzi, L.; Geleijnse, J.M.; Rauch, B.; Ness, A.; Galan, P.; et al. Omega-3 Treatment Trialists’ Collaboration. Associations of Omega-3 Fatty Acid Supplement Use with Cardiovascular Disease Risks: Meta-analysis of 10 Trials Involving 77 917 Individuals. JAMA Cardiol. 2018, 3, 225–234. [Google Scholar] [CrossRef] [Green Version]
- Abdelhamid, A.S.; Brown, T.J.; Brainard, J.S.; Biswas, P.; Thorpe, G.C.; Moore, H.J.; Deane, K.H.; Summerbell, C.D.; Worthington, H.V.; Song, F.; et al. Omega-3 fatty acids for the primary and secondary prevention of cardiovascular disease. Cochrane Database Syst. Rev. 2020, 3, CD003177. [Google Scholar] [CrossRef]
- Bhatt, D.L.; Steg, P.G.; Miller, M.; Brinton, E.A.; Jacobson, T.A.; Ketchum, S.B.; Doyle, R.T., Jr.; Juliano, R.A.; Jiao, L.; Granowitz, C.; et al. Cardiovascular Risk Reduction with Icosapent Ethyl for Hypertriglyceridemia. N. Engl. J. Med. 2019, 380, 11–22. [Google Scholar] [CrossRef]
- Budoff, M.J.; Bhatt, D.L.; Kinninger, A.; Lakshmanan, S.; Muhlestein, J.B.; Le, V.T.; May, H.T.; Shaikh, K.; Shekar, C.; Roy, S.K.; et al. Effect of icosapent ethyl on progression of coronary atherosclerosis in patients with elevated triglycerides on statin therapy: Final results of the EVAPORATE trial. Eur. Heart J. 2020, 41, 3925–3932. [Google Scholar] [CrossRef]
- Yokoyama, M.; Origasa, H.; Matsuzaki, M.; Matsuzawa, Y.; Saito, Y.; Ishikawa, Y.; Oikawa, S.; Sasaki, J.; Hishida, H.; Itakura, H.; et al. Effects of eicosapentaenoic acid on major coronary events in hypercholesterolaemic patients (JELIS): A randomised open-label, blinded endpoint analysis. Lancet 2007, 369, 1090–1098. [Google Scholar] [CrossRef]
- ASCEND Study Collaborative Group; Bowman, L.; Mafham, M.; Wallendszus, K.; Stevens, W.; Buck, G.; Barton, J.; Murphy, K.; Aung, T.; Haynes, R.; et al. Effects of n-3 Fatty Acid Supplements in Diabetes Mellitus. N. Engl. J. Med. 2018, 379, 1540–1550. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, S.J.; Lincoff, A.M.; Garcia, M.; Bash, D.; Ballantyne, C.M.; Barter, P.J.; Davidson, M.H.; Kastelein, J.J.P.; Koenig, W.; McGuire, D.K.; et al. Effect of High-Dose Omega-3 Fatty Acids vs Corn Oil on Major Adverse Cardiovascular Events in Patients at High Cardiovascular Risk: The STRENGTH Randomized Clinical Trial. JAMA 2020, 324, 2268–2280. [Google Scholar] [CrossRef] [PubMed]
- Unique Identifier: NCT02104817. Available online: https://www.clinicaltrials.gov (accessed on 12 October 2021).
- Kalstad, A.A.; Myhre, P.L.; Laake, K.; Tveit, S.H.; Schmidt, E.B.; Smith, P.; Nilsen, D.W.T.; Tveit, A.; Fagerland, M.W.; Solheim, S.; et al. OMEMI Investigators. Effects of n-3 Fatty Acid Supplements in Elderly Patients after Myocardial Infarction: A Randomized, Controlled Trial. Circulation 2021, 143, 528–539. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.C.; So, J.; Lamon-Fava, S. Differential and shared effects of eicosapentaenoic acid and docosahexaenoic acid on serum metabolome in subjects with chronic inflammation. Sci. Rep. 2021, 11, 16324. [Google Scholar] [CrossRef]
- Pareek, M.; Mason, R.P.; Bhatt, D.L. Icosapent ethyl: Safely reducing cardiovascular risk in adults with elevated triglycerides. Expert Opin. Drug Saf. 2022, 21, 31–42. [Google Scholar] [CrossRef]
- Lombardi, M.; Carbone, S.; Del Buono, M.G.; Chiabrando, J.G.; Vescovo, G.M.; Camilli, M.; Montone, R.A.; Vergallo, R.; Abbate, A.; Biondi-Zoccai, G.; et al. Omega-3 fatty acids supplementation and risk of atrial fibrillation: An updated meta-analysis of randomized controlled trials. Eur. Heart J. Cardiovasc. Pharmacother. 2021, 7, e69–e70. [Google Scholar] [CrossRef]
- Doi, T.; Langsted, A.; Nordestgaard, B.G. A possible explanation for the contrasting results of REDUCE-IT vs. STRENGTH: Cohort study mimicking trial designs. Eur. Heart J. 2021, 42, 4807–4817. [Google Scholar] [CrossRef]
- Lakshmanan, S.; Shekar, C.; Kinninger, A.; Dahal, S.; Onuegbu, A.; Cai, A.N.; Hamal, S.; Birudaraju, D.; Roy, S.K.; Nelson, J.R.; et al. Comparison of mineral oil and non-mineral oil placebo on coronary plaque progression by coronary computed tomography angiography. Cardiovasc. Res. 2020, 116, 479–482. [Google Scholar] [CrossRef] [Green Version]
- Tadic, M.; Sala, C.; Grassi, G.; Mancia, G.; Taddei, S.; Rottbauer, W.; Cuspidi, C. Omega-3 Fatty Acids and Coronary Artery Disease: More Questions Than Answers. J. Clin. Med. 2021, 10, 2495. [Google Scholar] [CrossRef]
- Sarajlic, P.; Artiach, G.; Larsson, S.C.; Back, M. Dose-dependent risk reduction for myocardial infarction with eicosapentaenoic acid: A meta-analysis and metaregression including the STRENGTH trial. Cardiovasc. Drugs Ther. 2021, 35, 1079–1081. [Google Scholar] [CrossRef]
- Pirillo, A.; Catapano, A.L. Omega-3 for Cardiovascular Diseases: Where Do We Stand after REDUCE-IT and STRENGTH? Circulation 2021, 144, 183–185. [Google Scholar] [CrossRef] [PubMed]
- Margină, D.; Ungurianu, A.; Purdel, C.; Nițulescu, G.M.; Tsoukalas, D.; Sarandi, E.; Thanasoula, M.; Burykina, T.I.; Tekos, F.; Buha, A.; et al. Analysis of the intricate effects of polyunsaturated fatty acids and polyphenols on inflammatory pathways in health and disease. Food Chem. Toxicol. 2020, 143, 111558. [Google Scholar] [CrossRef] [PubMed]
- Elagizi, A.; Lavie, C.J.; O’Keefe, E.; Marshall, K.; O’Keefe, J.H.; Milani, R.V. An Update on Omega-3 Polyunsaturated Fatty Acids and Cardiovascular Health. Nutrients 2021, 13, 204. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, T.; Miller, M. A Fishy Topic: VITAL, REDUCE-IT, STRENGTH, and Beyond: Putting Omega-3 Fatty Acids into Practice in 2021. Curr. Cardiol. Rep. 2021, 23, 111. [Google Scholar] [CrossRef]
- Watanabe, T.; Ando, K.; Daidoji, H.; Otaki, Y.; Sugawara, S.; Matsui, M.; Ikeno, E.; Hirono, O.; Miyawaki, H.; Yashiro, Y.; et al. CHERRY study investigators. A randomized controlled trial of eicosapentaenoic acid in patients with coronary heart disease on statins. J. Cardiol. 2017, 70, 537–544. [Google Scholar] [CrossRef] [Green Version]
- ORIGIN Trial Investigators; Bosch, J.; Gerstein, H.C.; Dagenais, G.R.; Díaz, R.; Dyal, L.; Jung, H.; Maggiono, A.P.; Probstfield, J.; Ramachandran, A.; et al. N-3 fatty acids and cardiovascular outcomes in patients with dysglycemia. N. Engl. J. Med. 2012, 367, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Manson, J.E.; Cook, N.R.; Lee, I.M.; Christen, W.; Bassuk, S.S.; Mora, S.; Gibson, H.; Albert, C.M.; Gordon, D.; Copeland, T.; et al. Marine n-3 Fatty Acids and Prevention of Cardiovascular Disease and Cancer. N. Engl. J. Med. 2019, 380, 23–32. [Google Scholar] [CrossRef]
- Thomsen, M.; Varbo, A.; Tybjærg-Hansen, A.; Nordestgaard, B.G. Low nonfasting triglycerides and reduced all-cause mortality: A mendelian randomization study. Clin. Chem. 2014, 60, 737–746. [Google Scholar] [CrossRef] [Green Version]
- Varbo, A.; Nordestgaard, B.G. Nonfasting Triglycerides, Low-Density Lipoprotein Cholesterol, and Heart Failure Risk: Two Cohort Studies of 113,554 Individuals. Arter. Thromb. Vasc. Biol. 2018, 38, 464–472. [Google Scholar] [CrossRef] [Green Version]
- Annuzzi, G.; Rivellese, A.A.; Wang, H.; Patti, L.; Vaccaro, O.; Riccardi, G.; Ebbesson, S.O.; Comuzzie, A.G.; Umans, J.G.; Howard, B.V. Lipoprotein subfractions and dietary intake of n-3 fatty acid: The Genetics of Coronary Artery Disease in Alaska Natives study. Am. J. Clin. Nutr. 2012, 95, 1315–1322. [Google Scholar] [CrossRef]
- Amigó, N.; Akinkuolie, A.O.; Chiuve, S.E.; Correig, X.; Cook, N.R.; Mora, S. Habitual Fish Consumption, ω-3 Fatty Acids, and Nuclear Magnetic Resonance Lipoprotein Subfractions in Women. J. Am. Heart Assoc. 2020, 9, e014963. [Google Scholar] [CrossRef] [PubMed]
- Oelrich, B.; Dewell, A.; Gardner, C.D. Effect of fish oil supplementation on serum triglycerides, LDL cholesterol and LDL subfractions in hypertriglyceridemic adults. Nutr. Metab. Cardiovasc. Dis. 2013, 23, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Harris, W.S.; Connor, W.E.; Alam, N.; Illingworth, D.R. Reduction of postprandial triglyceridemia in humans by dietary n-3 fatty acids. J. Lipid Res. 1988, 29, 1451–1460. [Google Scholar] [CrossRef]
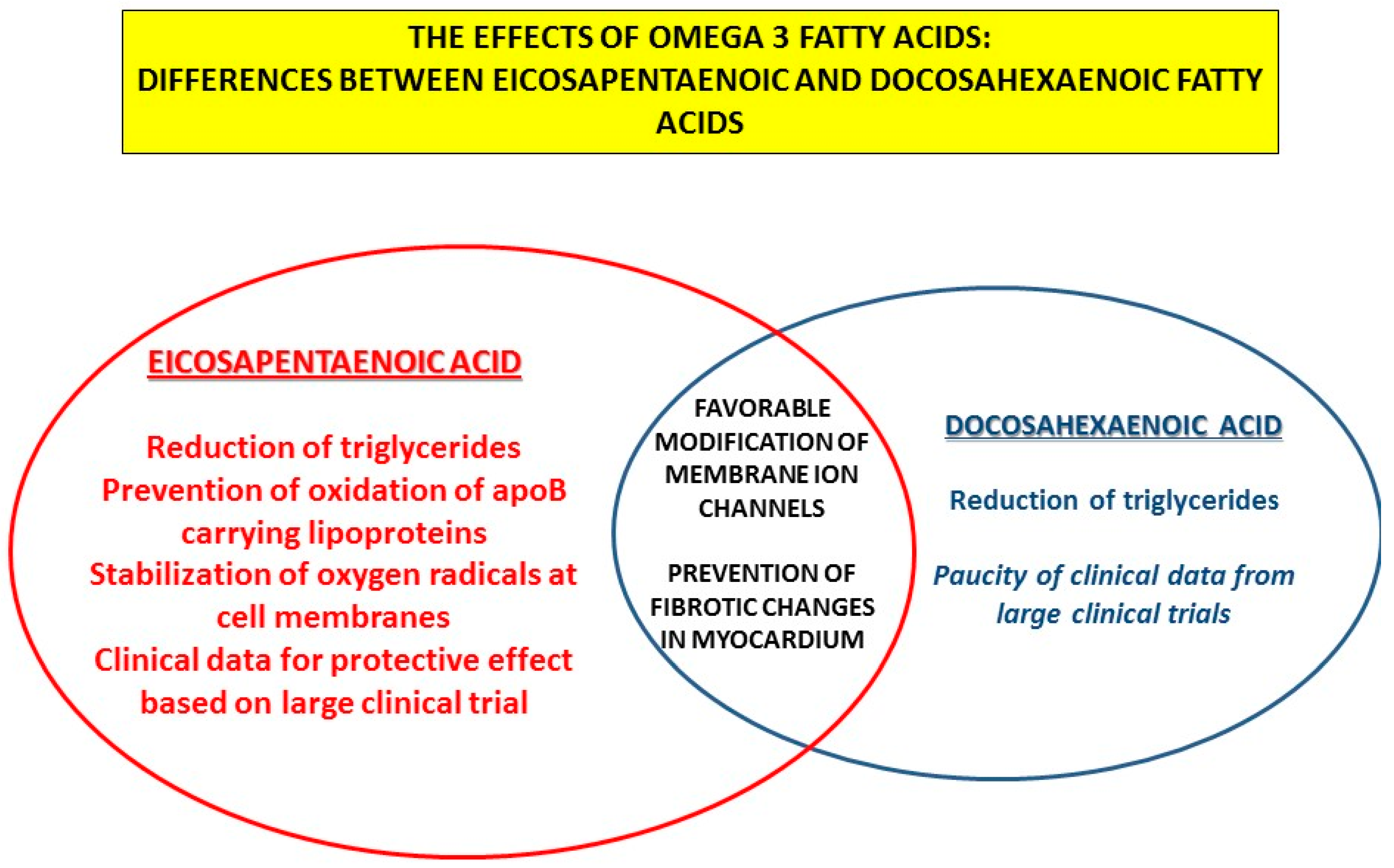
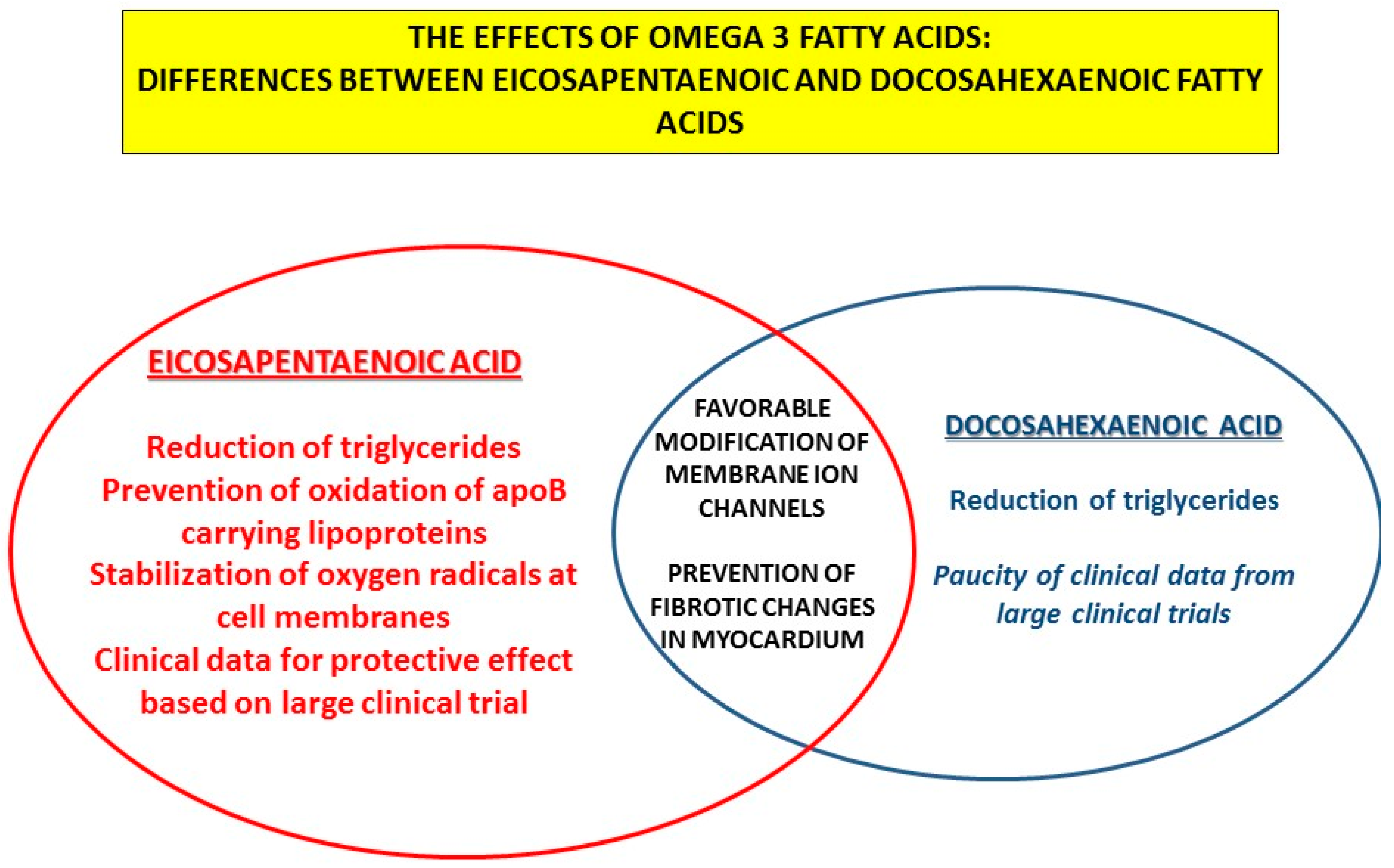
- Innes, J.K.; Calder, P.C. The Differential Effects of Eicosapentaenoic Acid and Docosahexaenoic Acid on Cardiometabolic Risk Factors: A Systematic Review. Int. J. Mol. Sci. 2018, 19, 532. [Google Scholar] [CrossRef] [Green Version]
- Mason, R.P.; Jacob, R.F.; Shrivastava, S.; Sherratt, S.C.R.; Chattopadhyay, A. Eicosapentaenoic acid reduces membrane fluidity, inhibits cholesterol domain formation, and normalizes bilayer width in atherosclerotic-like model membranes. Biochim. Biophys Acta 2016, 1858, 3131–3140. [Google Scholar] [CrossRef] [Green Version]
- Mason, R.P.; Sherratt, S.C.; Jacob, R.F. Eicosapentaenoic acid inhibits oxidation of ApoB-containing lipoprotein particles of different size in vitro when administered alone or in combination with atorvastatin active metabolite compared with other triglyceride-lowering agents. J. Cardiovasc. Pharmacol. 2016, 68, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Mason, R.P.; Jacob, R.F. Eicosapentaenoic acid inhibits glucose-induced membrane cholesterol crystalline domain formation through a potent antioxidant mechanism. Biochim. Biophys. Acta 2015, 1848, 502–509. [Google Scholar] [CrossRef] [Green Version]
- Mason, R.P.; Libby, P.; Bhatt, D.L. Emerging Mechanisms of Cardiovascular Protection for the Omega-3 Fatty Acid Eicosapentaenoic Acid. Arter. Thromb. Vasc. Biol. 2020, 40, 1135–1147. [Google Scholar] [CrossRef]
- Masuda, D.; Miyata, Y.; Matsui, S.; Yamashita, S. Omega-3 fatty acid ethyl esters improve low-density lipoprotein subclasses without increasing low-density lipoprotein-cholesterol levels: A phase 4, randomized study. Atherosclerosis 2020, 292, 163–170. [Google Scholar] [CrossRef] [Green Version]
- Pitha, J.; Poledne, R. The Truth About Fish (Oil) in the Treatment of Dyslipidemia. Curr Atheroscler. Rep. 2021, 23, 10. [Google Scholar] [CrossRef]
- Spencer, M.; Finlin, B.S.; Unal, R.; Zhu, B.; Morris, A.J.; Shipp, L.R.; Lee, J.; Walton, R.G.; Adu, A.; Erfani, R.; et al. Omega-3 fatty acids reduce adipose tissue macrophages in human subjects with insulin resistance. Diabetes 2013, 62, 1709–1717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.R.; Midgette, Y.; Shah, R. Fish Oil Derived Omega 3 Fatty Acids Suppress Adipose NLRP3 Inflammasome Signaling in Human Obesity. J. Endocr. Soc. 2018, 3, 504–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pisaniello, A.D.; Psaltis, P.J.; King, P.M.; Liu, G.; Gibson, R.A.; Tan, J.T.; Duong, M.; Nguyen, T.; Bursill, C.A.; Worthley, M.I.; et al. Omega-3 fatty acids ameliorate vascular inflammation: A rationale for their atheroprotective effects. Atherosclerosis 2021, 324, 27–37. [Google Scholar] [CrossRef]
- Mihalj, M.; Stupin, A.; Kolobarić, N.; Bujak, I.T.; Matić, A.; Kralik, Z.; Jukić, I.; Stupin, M.; Drenjančević, I. Leukocyte Activation and Antioxidative Defense Are Interrelated and Moderately Modified by n-3 Polyunsaturated Fatty Acid-Enriched Eggs Consumption-Double-Blind Controlled Randomized Clinical Study. Nutrients 2020, 12, 3122. [Google Scholar] [CrossRef] [PubMed]
- Kolobarić, N.; Drenjančević, I.; Matić, A.; Šušnjara, P.; Mihaljević, Z.; Mihalj, M. Dietary Intake of n-3 PUFA-Enriched Hen Eggs Changes Inflammatory Markers’ Concentration and Treg/Th17 Cells Distribution in Blood of Young Healthy Adults—A Randomised Study. Nutrients 2021, 13, 1851. [Google Scholar] [CrossRef] [PubMed]
- Stupin, A.; Rasic, L.; Matic, A.; Stupin, M.; Kralik, Z.; Kralik, G.; Grcevic, M.; Drenjancevic, I. Omega-3 polyunsaturated fatty acids-enriched hen eggs consumption enhances microvascular reactivity in young healthy individuals. Appl. Physiol. Nutr. Metab. 2018, 43, 988–995. [Google Scholar] [CrossRef]
- Stupin, A.; Mihalj, M.; Kolobarić, N.; Šušnjara, P.; Kolar, L.; Mihaljević, Z.; Matić, A.; Stupin, M.; Jukić, I.; Kralik, Z.; et al. Anti-Inflammatory Potential of n-3 Polyunsaturated Fatty Acids Enriched Hen Eggs Consumption in Improving Microvascular Endothelial Function of Healthy Individuals-Clinical Trial. Int. J. Mol. Sci. 2020, 21, 4149. [Google Scholar] [CrossRef]
- Allam-Ndoul, B.; Guénard, F.; Barbier, O.; Vohl, M.C. Effect of different concentrations of omega-3 fatty acids on stimulated THP-1 macrophages. Genes Nutr. 2017, 12, 7. [Google Scholar] [CrossRef] [Green Version]
- Koppelmann, T.; Pollak, Y.; Ben-Shahar, Y.; Gorelik, G.; Sukhotnik, I. The Mechanisms of the Anti-Inflammatory and Anti-Apoptotic Effects of Omega-3 Polyunsaturated Fatty Acids during Methotrexate-Induced Intestinal Damage in Cell Line and in a Rat Model. Nutrients 2021, 13, 888. [Google Scholar] [CrossRef]
- Endo, J.; Arita, M. Cardioprotective mechanism of omega-3 polyunsaturated fatty acids. J. Cardiol. 2016, 67, 22–27. [Google Scholar] [CrossRef] [Green Version]
- Soni, N.; Ross, A.B.; Scheers, N.; Nookaew, I.; Gabrielsson, B.G.; Sandberg, A.S. The Omega-3 Fatty Acids EPA and DHA, as a Part of a Murine High-Fat Diet, Reduced Lipid Accumulation in Brown and White Adipose Tissues. Int. J. Mol. Sci. 2019, 20, 5895. [Google Scholar] [CrossRef] [Green Version]
- Doaei, S.; Gholami, S.; Rastgoo, S.; Gholamalizadeh, M.; Bourbour, F.; Bagheri, S.E.; Samipoor, F.; Akbari, M.E.; Shadnoush, M.; Ghorat, F.; et al. The effect of omega-3 fatty acid supplementation on clinical and biochemical parameters of critically ill patients with COVID-19: A randomized clinical trial. J. Transl. Med. 2021, 19, 128. [Google Scholar] [CrossRef]
- Darwesh, A.M.; Bassiouni, W.; Sosnowski, D.K.; Seubert, J.M. Can N-3 polyunsaturated fatty acids be considered a potential adjuvant therapy for COVID-19-associated cardiovascular complications? Pharmacol. Ther. 2021, 219, 107703. [Google Scholar] [CrossRef]
- Díaz, R. PREPARE-IT 2: A pragmatic trial evaluating icosapent ethyl in nonhospitalized patients with a positive diagnosis of COVID-19 to reduce hospitalization rates and complications. In Proceedings of the American Heart Association’s Scientific Sessions 2021, Boston, MA, USA, 13–15 November 2021. [Google Scholar]
- Kosmopoulos, A.; Bhatt, D.L.; Meglis, G.; Verma, R.; Pan, Y.; Quan, A.; Teoh, H.; Verma, M.; Jiao, L.; Wang, R.; et al. A randomized trial of icosapent ethyl in ambulatory patients with COVID-19. iScience 2021, 24, 103040. [Google Scholar] [CrossRef]
- Dyerberg, J.; Bang, H.O.; Stoffersen, E.; Moncada, S.; Vane, J.R. Eicosapentaenoic acid and prevention of thrombosis and atherosclerosis? Lancet 1978, 2, 117–119. [Google Scholar] [CrossRef]
- Dona, M.; Fredman, G.; Schwab, J.M.; Chiang, N.; Arita, M.; Goodarzi, A.; Cheng, G.; von Andrian, U.H.; Serhan, C.N. Resolvin E1, an EPA-derived mediator in whole blood, selectively counterregulates leukocytes and platelets. Blood 2008, 112, 848–855. [Google Scholar] [CrossRef] [Green Version]
- Stupin, M.; Kibel, A.; Stupin, A.; Selthofer-Relatić, K.; Matić, A.; Mihalj, M.; Mihaljević, Z.; Jukić, I.; Drenjančević, I. The Physiological Effect of n-3 Polyunsaturated Fatty Acids (n-3 PUFAs) Intake and Exercise on Hemorheology, Microvascular Function, and Physical Performance in Health and Cardiovascular Diseases; Is There an Interaction of Exercise and Dietary n-3 PUFA Intake? Front. Physiol. 2019, 10, 1129. [Google Scholar] [CrossRef] [Green Version]
- Shahar, E.; Folsom, A.R.; Wu, K.K.; Dennis, B.H.; Shimakawa, T.; Conlan, M.G.; Davis, C.E.; Williams, O.D. Associations of fish intake and dietary n-3 polyunsaturated fatty acids with a hypocoagulable profile. The Atherosclerosis Risk in Communities (ARIC) Study. Arter. Thromb. 1993, 13, 1205–1212. [Google Scholar] [CrossRef] [Green Version]
- Cawood, A.L.; Ding, R.; Napper, F.L.; Young, R.H.; Williams, J.A.; Ward, M.J.; Gudmundsen, O.; Vige, R.; Payne, S.P.; Ye, S.; et al. Eicosapentaenoic acid (EPA) from highly concentrated n-3 fatty acid ethyl esters is incorporated into advanced atherosclerotic plaques and higher plaque EPA is associated with decreased plaque inflammation and increased stability. Atherosclerosis 2010, 212, 252–259. [Google Scholar] [CrossRef]
- Sherratt, S.C.R.; Juliano, R.A.; Mason, R.P. Eicosapentaenoic acid (EPA) has optimal chain length and degree of unsaturation to inhibit oxidation of small dense LDL and membrane cholesterol domains as compared to related fatty acids in vitro. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183254. [Google Scholar] [CrossRef]
- Earley, S.; Pauyo, T.; Drapp, R.; Tavares, M.J.; Liedtke, W.; Brayden, J.E. TRPV4-dependent dilation of peripheral resistance arteries influences arterial pressure. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H1096–H1102. [Google Scholar] [CrossRef] [Green Version]
- Namiranian, K.; Lloyd, E.E.; Crossland, R.F.; Marrelli, S.P.; Taffet, G.E.; Reddy, A.K.; Hartley, C.J.; Bryan, R.M., Jr. Cerebrovascular responses in mice deficient in the potassium channel, TREK-1. AJP Integr. Comp. Physiol. 2010, 299, R461–R469. [Google Scholar] [CrossRef] [Green Version]
- Goto, K.; Ohtsubo, T.; Kitazono, T. Endothelium-Dependent Hyperpolarization (EDH) in Hypertension: The Role of Endothelial Ion Channels. Int. J. Mol. Sci. 2018, 19, 315. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Hong, S.; Li, P.L.; Zhang, Y. Docosahexanoic acid-induced coronary arterial dilation: Actions of 17S-hydroxy docosahexanoic acid on K+ channel activity. J. Pharmacol. Exp. Ther. 2011, 336, 891–899. [Google Scholar] [CrossRef] [Green Version]
- Caires, R.; Sierra-Valdez, F.J.; Millet, J.R.M.; Herwig, J.D.; Roan, E.; Vásquez, V.; Cordero-Morales, J.F. Omega-3 Fatty Acids Modulate TRPV4 Function through Plasma Membrane Remodeling. Cell Rep. 2017, 21, 246–258. [Google Scholar] [CrossRef] [Green Version]
- Engler, M.B.; Ma, Y.H.; Engler, M.M. Calcium-mediated mechanisms of eicosapentaenoic acid-induced relaxation in hypertensive rat aorta. Am. J. Hypertens. 1999, 12, 1225–1235. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Zhang, C.; Dong, Y.; Wang, S.; Song, P.; Viollet, B.; Zou, M.-H. Activation of the AMP-activated protein kinase by eicosapentaenoic acid (EPA, 20:5 n-3) improves endothelial function in vivo. PLoS ONE 2012, 7, e35508. [Google Scholar] [CrossRef] [Green Version]
- Van den Elsen, L.W.J.; Spijkers, L.J.A.; van den Akker, R.F.P.; van Winssen, A.M.H.; Balvers, M.; Wijesinghe, D.S.; Chalfant, C.E.; Garssen, J.; Willemsen, L.E.M.; Alewijnse, A.E.; et al. Dietary fish oil improves endothelial function and lowers blood pressure via suppression of sphingolipid-mediated contractions in spontaneously hypertensive rats. J. Hypertens. 2014, 32, 1050–1058. [Google Scholar] [CrossRef] [Green Version]
- Gwon, D.H.; Hwang, T.W.; Ro, J.Y.; Kang, Y.J.; Jeong, J.Y.; Kim, D.K.; Lim, K.; Kim, D.W.; Choi, D.E.; Kim, J.J. High Endogenous Accumulation of ω-3 Polyunsaturated Fatty Acids Protect against Ischemia-Reperfusion Renal Injury through AMPK-Mediated Autophagy in Fat-1 Mice. Int. J. Mol. Sci. 2017, 18, 2081. [Google Scholar] [CrossRef] [Green Version]
- Demaison, L.; Sergiel, J.P.; Moreau, D.; Grynberg, A. Influence of the phospholipid n-6/n-3 polyunsaturated fatty acid ratio on the mitochondrial oxidative metabolism before and after myocardial ischemia. Biochim. Biophys. Acta 1994, 1227, 53–59. [Google Scholar] [CrossRef]
- O’Connell, T.D.; Block, R.C.; Huang, S.P.; Shearer, G.C. ω3-Polyunsaturated fatty acids for heart failure: Effects of dose on efficacy and novel signaling through free fatty acid receptor 4. J. Mol. Cell Cardiol. 2017, 103, 74–92. [Google Scholar] [CrossRef] [Green Version]
- Eclov, J.A.; Qian, Q.; Redetzke, R.; Chen, Q.; Wu, S.C.; Healy, C.L.; Ortmeier, S.B.; Harmon, E.; Shearer, G.C.; O’Connell, T.D. EPA, not DHA, prevents fibrosis in pressure overload-induced heart failure: Potential role of free fatty acid receptor 4. J. Lipid Res. 2015, 56, 2297–2308. [Google Scholar] [CrossRef] [Green Version]
- Matsuo, N.; Miyoshi, T.; Takaishi, A.; Kishinoue, T.; Yasuhara, K.; Tanimoto, M.; Nakano, Y.; Onishi, N.; Ueeda, M.; Ito, H. High Plasma Docosahexaenoic Acid Associated to Better Prognoses of Patients with Acute Decompensated Heart Failure with Preserved Ejection Fraction. Nutrients 2021, 13, 371. [Google Scholar] [CrossRef]
- Białek, M.; Białek, A.; Czauderna, M. Maternal and Early Postnatal Diet Supplemented with Conjugated Linoleic Acid Isomers Affect Lipid Profile in Hearts of Offspring Rats with Mammary Tumors. Animals 2020, 10, 464. [Google Scholar] [CrossRef] [Green Version]
- Białek, A.; Białek, M.; Lepionka, T.; Ruszczyńska, A.; Bulska, E.; Czauderna, M. Cancer Influences the Elemental Composition of the Myocardium More Strongly than Conjugated Linoleic Acids-Chemometric Approach to Cardio-Oncological Studies. Molecules 2021, 26, 7127. [Google Scholar] [CrossRef]
- Ramirez, J.L.; Gasper, W.J.; Khetani, S.A.; Zahner, G.J.; Hills, N.K.; Mitchell, P.T.; Sansbury, B.E.; Conte, M.S.; Spite, M.; Grenon, S.M. Fish Oil Increases Specialized Pro-resolving Lipid Mediators in PAD (The OMEGA-PAD II Trial) Randomized Controlled Trial. J. Surg. Res. 2019, 238, 164–174. [Google Scholar] [CrossRef]
- Sawada, T.; Tsubata, H.; Hashimoto, N.; Takabe, M.; Miyata, T.; Aoki, K.; Yamashita, S.; Oishi, S.; Osue, T.; Yokoi, K.; et al. Effects of 6-month eicosapentaenoic acid treatment on postprandial hyperglycemia, hyperlipidemia, insulin secretion ability, and concomitant endothelial dysfunction among newly-diagnosed impaired glucose metabolism patients with coronary artery disease. An open label, single blinded, prospective randomized controlled trial. Cardiovasc. Diabetol. 2016, 15, 121. [Google Scholar] [CrossRef] [Green Version]
- Filipovic, M.G.; Aeschbacher, S.; Reiner, M.F.; Stivala, S.; Gobbato, S.; Bonetti, N.; Risch, M.; Risch, L.; Camici, G.G.; Luescher, T.F.; et al. Whole blood omega-3 fatty acid concentrations are inversely associated with blood pressure in young, healthy adults. J. Hypertens. 2018, 36, 548–1554. [Google Scholar] [CrossRef] [Green Version]
- Colussi, G.L.; Catena, C.; Dialti, V.; Mos, L.; Sechi, L.A. The vascular response to vasodilators is related to the membrane content of polyunsaturated fatty acids in hypertensive patients. J. Hypertens. 2015, 33, 993–1000. [Google Scholar] [CrossRef]
- Strand, E.; Pedersen, E.R.; Svingen, G.F.T.; Schartum-Hansen, H.; Rebnord, E.W.; Bjørndal, B.; Seifert, R.; Bohov, P.; Meyer, K.; Hiltunen, J.K.; et al. Dietary intake of n-3 long-chain polyunsaturated fatty acids and risk of myocardial infarction in coronary artery disease patients with or without diabetes mellitus: A prospective cohort study. BMC Med. 2013, 11, 216. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Intervention | Main Characteristics | Clinical Effect Including Adverse Events | |
|---|---|---|---|
| Successful—primary endpoint reached | |||
| REDUCE-IT [8] | Icosapent ethyl 4 g/day, mineral oil as control | Patients with established CVD or DM on statin therapy with increased TG (n = 8179; 29% women, FU 4.9 years), median TG 216 mg/dL (2.44 mmol/L) | The risk of CV death, MI, revascularization, unstable angina was significantly lower in treated patients (26% reduction). The incidence of atrial fibrillation and peripheral edema were significantly higher in the treatment group. The overall rates of serious adverse bleeding were similar. No fatal bleedings and no between-group differences in the rates of hemorrhagic stroke, serious central nervous system bleeding, or gastrointestinal bleeding. |
| EVAPORATE [9] | Icosapent ethyl 4 g/day, mineral oil as control | Patients with confirmed coronary artery stenosis, on statin therapy with increased TG (n = 80; 44 women, FU 1.5 year), median TG 191–200 mg/dL (2.48–2.60 mmol/L) | Low-attenuation plaque volume and thickening of fibrotic cap were significantly reduced in the treatment group. No serious adverse effects were described. |
| JELIS [10] | EPA 1.8 g/day (+pravastatin or simvastatin), no placebo | Patients with previous MI or PCI or with confirmed angina pectoris or without CVD (n = 18,645; 69% women, FU 4.6 years), TG 150.4 mg/dL (1.7 mmol/L) | All-cause mortality was reduced in secondary, but not in primary prevention. Adverse experiences and discontinuation rate due to treatment was more common in the treatment group, including laboratory data, gastrointestinal disturbances, skin abnormalities, and cerebral and fundal bleedings, epistaxis and subcutaneous bleeding (all mild). Incidence of new cancers was not different between groups. |
| CHERRY [26] | Pitavastatin and EPA therapy (4 mg/day and 1800 mg/day) vs. pitavastatin (4 mg/day), | Patients after PCI, pitavastatin/EPA group (n = 97) vs. pitavastatin group (n = 96), 16–20% women, FU 6–8 months, median TG 105–111 mg/dL (1.2–1.25 mmol/L) | Statin + EPA therapy significantly reduced coronary plaque volume vs. statin therapy alone. Plaque stabilization was reinforced by statin/EPA in patients with stable angina pectoris. No significant difference in adverse events, including atrial fibrillation. |
| Not successful | |||
| ORIGIN [27] | EPA (465 mg) + DHA (375 mg) vs. placebo (approx. 1 g of olive oil). | High risk of CVD + impaired Fasting glucose/glucose intolerance/DM (n = 12,536; 35% women; FU 6.2 years). Median TG 140–142 mg/dL (1.58–1.60 mmol/L) | No reduction in non-fatal MI or stroke, death from CV cause or arrhythmia was observed. No differences in major bleeding. |
| ASCEND [11] | EPA + DHA combined (1 g/day) vs. olive oil capsule | Persons older than 40 years + DM, without CVD (n = 15,480, FU 7.4 years, 37% female), TG not measured | No effect on non-fatal MI or stroke, vascular death in treated patients. No significant between-group differences in the rates of non-fatal serious adverse events, including new cancers. |
| STRENGTH [12] | EPA + DHA combined (4 g/day) vs. corn oil. | Patients with CVD or at high risk for CVD (n = 13,078; FU 3.5 years, 35% women), median TG 240 mg (2.72 mmol/L) | No significant effect of treatment on CV death, non-fatal MI, stroke, coronary revascularization or unstable angina observed. The incidence of gastrointestinal adverse events was higher in the treatment group. New-onset atrial fibrillation was more common in the treatment group. Major and minor bleeding events were not different between groups. |
| OMEMI [14] | EPA 930 mg + DHA 660 mg vs. corn oil | Age 70 to 82 year + recent (2–8 weeks) acute MI (n = 1014, FU 2 years; 29% females), mean TG 111.4 mg/dL (1.26 mmol/L) | No reduction in clinical events. No differences in major bleeding (10.7% vs. 11.0% in controls). No patients withdrew because of bleeding problems. Reasons for discontinuing treatment not different between the groups (gastrointestinal symptoms, difficulty swallowing capsules, other disease burdens not related to the study intervention). |
| VITAL [28] | Vitamin D3 (2000 IU per day) and 1 g fish-oil capsule (EPA-460 mg, DHA-380 mg) vs. placebo (not specified) | Healthy men > 50 and women > 55 years of age (n = 25,871; FU 5.3 years, 51% women), TG not specified | No effect on MI, stroke, CV death in treated patients was detected; benefit was observed in African Americans (HR: 0.23); PCI (HR: 0.78); fatal MI (HR: 0.50). No excess risks of bleeding or other serious adverse events, including gastrointestinal symptoms, major bleeding episodes observed |
| Main Targets | Mechanisms | Evidence | Clinical Effect |
|---|---|---|---|
| Indirect effects: Circulating free fatty acids Circulating triglycerides Thrombotic factors Inflammatory factors | Free fatty acid availability Reduction of subclinical inflammation, detectable by measurement of the wide spectrum of inflammatory markers, anti-fibrotic effects | One successful randomized trial with icosapent ethyl (EPA derivative) in higher dose, parallel trial with reduction of coronary atherosclerotic plaques in DHA data less convincing, potential role in HFpEF (evidence from observational study based on DHA plasma levels). | Lowering plasma triglycerides Lowering blood pressure Improving endothelial function measured in peripheral vessels (mostly experimental settings in humans) |
| Direct effects: Cell membranes Ion channels | Membrane stabilization, including their protection against free radicals Modification of different types of ion channels | In human, mostly indirect evidence from experiments, in vitro studies. Observational studies indicating decrease of sudden death with intake of fish oil. | Experimental data In vitro studies Observational/epidemiological studies in humans |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drenjančević, I.; Pitha, J. Omega-3 Polyunsaturated Fatty Acids—Vascular and Cardiac Effects on the Cellular and Molecular Level (Narrative Review). Int. J. Mol. Sci. 2022, 23, 2104. https://doi.org/10.3390/ijms23042104
Drenjančević I, Pitha J. Omega-3 Polyunsaturated Fatty Acids—Vascular and Cardiac Effects on the Cellular and Molecular Level (Narrative Review). International Journal of Molecular Sciences. 2022; 23(4):2104. https://doi.org/10.3390/ijms23042104
Chicago/Turabian StyleDrenjančević, Ines, and Jan Pitha. 2022. "Omega-3 Polyunsaturated Fatty Acids—Vascular and Cardiac Effects on the Cellular and Molecular Level (Narrative Review)" International Journal of Molecular Sciences 23, no. 4: 2104. https://doi.org/10.3390/ijms23042104
APA StyleDrenjančević, I., & Pitha, J. (2022). Omega-3 Polyunsaturated Fatty Acids—Vascular and Cardiac Effects on the Cellular and Molecular Level (Narrative Review). International Journal of Molecular Sciences, 23(4), 2104. https://doi.org/10.3390/ijms23042104