
Genome Editing Using CRISPR-Cas9 and Autoimmune Diseases: A Comprehensive Review

 ,
,  , , , , , and
, , , , , and 
Abstract
:1. Introduction
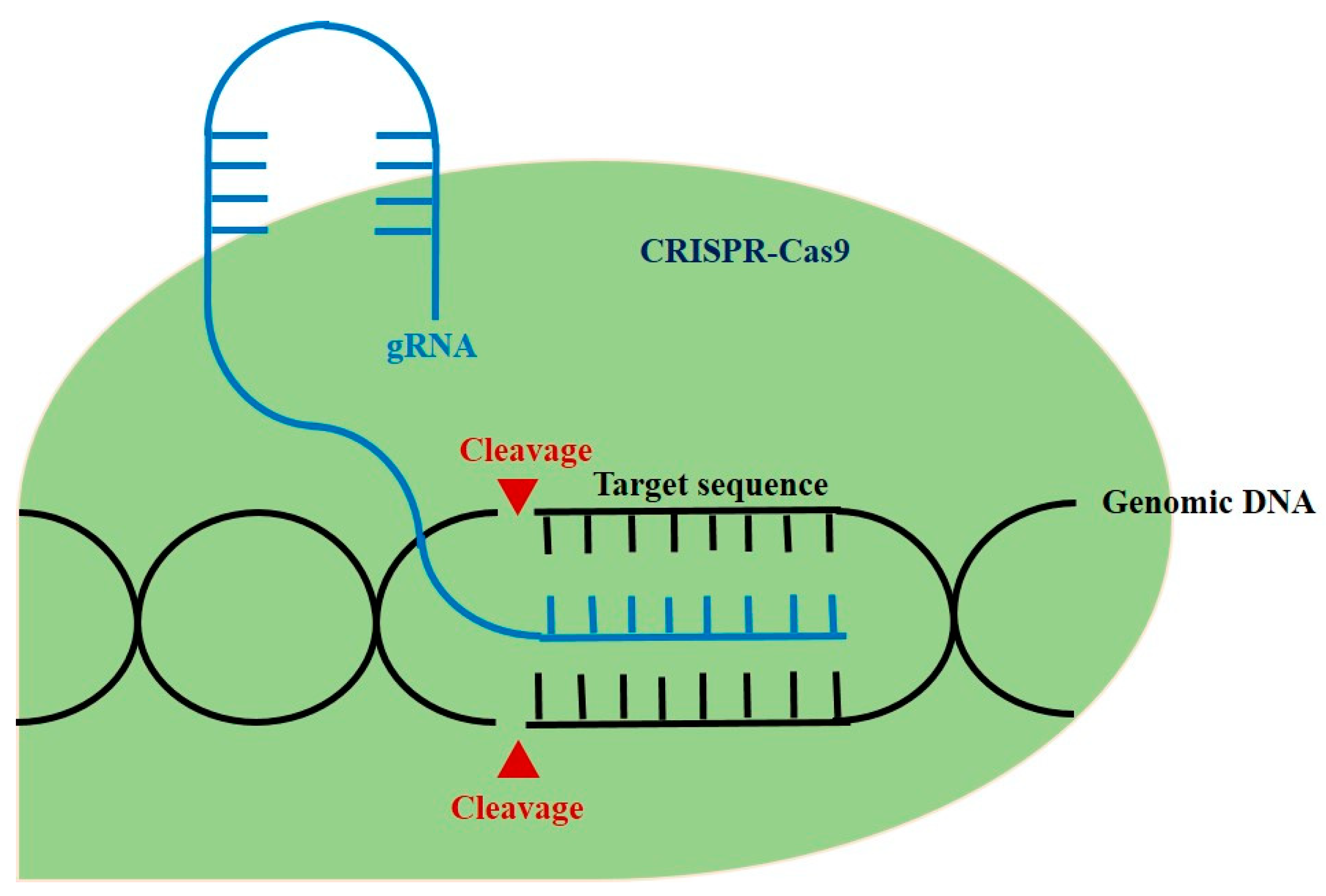
2. CRISPR-Cas9 and Autoimmune Diseases
2.1. In Vivo Therapeutic Trials Using CRISPR-Cas9
2.2. Inflammatory Molecules, Immunogenetics and CRISPR-Cas9
2.3. CRISPR-Cas9 and iPSC
3. CRISPR-Cas9 and Rheumatoid Arthritis
4. CRISPR-Cas9 and Inflammatory Bowel Disease
4.1. Studies on CRISPR-Cas9 and IBD Using Human Immune Cells
4.2. Studies on CRISPR-Cas9 and IBD Using Non-Human Cells
5. CRISPR-Cas9 and Systemic Lupus Erythematosus
6. CRISPR-Cas9 and Multiple Sclerosis
7. CRISPR-Cas9 and Type 1 Diabetes Mellitus
8. CRISPR-Cas9 and Psoriasis
9. CRISPR-Cas9 and Type 1 Coeliac Disease
10. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Marrack, P.; Kappler, J.; Kotzin, B.L. Autoimmune disease: Why and where it occurs. Nat. Med. 2001, 7, 899–905. [Google Scholar] [CrossRef] [PubMed]
- Benros, M.E.; Waltoft, B.L.; Nordentoft, M.; Ostergaard, S.D.; Eaton, W.W.; Krogh, J.; Mortensen, P.B. Autoimmune diseases and severe infections as risk factors for mood disorders a nationwide study. JAMA Psychiatry 2013, 70, 812–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, G.S.; Miller, F.W.; Pandey, J.P. The role of genetic factors in autoimmune disease: Implications for environmental research. Environ. Health Perspect. 1999, 107 (Suppl. S5), 693–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, S.-A.; Wang, J.; Tao, M.-H.; Leung, P.S.C. Gene therapy for autoimmune disease. Clin. Rev. Allergy Immunol. 2015, 49, 163–176. [Google Scholar] [CrossRef]
- Yin, H.; Kauffman, K.J.; Anderson, D.G. Delivery technologies for genome editing. Nat. Rev. Drug Discov. 2017, 16, 387–399. [Google Scholar] [CrossRef]
- Gilbert, L.A.; Larson, M.H.; Morsut, L.; Liu, Z.; Brar, G.A.; Torres, S.E.; Stern-Ginossar, N.; Brandman, O.; Whitehead, E.H.; Doudna, J.A.; et al. Crispr-mediated modular rna-guided regulation of transcription in eukaryotes. Cell 2013, 154, 442–451. [Google Scholar] [CrossRef] [Green Version]
- Doudna, J.A.; Charpentier, E. Genome editing. The new frontier of genome engineering with crispr-cas9. Science 2014, 346, 1258096. [Google Scholar] [CrossRef]
- Hsu, P.D.; Lander, E.S.; Zhang, F. Development and applications of crispr-cas9 for genome engineering. Cell 2014, 157, 1262–1278. [Google Scholar] [CrossRef] [Green Version]
- Ding, Q.; Strong, A.; Patel, K.M.; Ng, S.L.; Gosis, B.S.; Regan, S.N.; Cowan, C.A.; Rader, D.J.; Musunuru, K. Permanent alteration of pcsk9 with in vivo crispr-cas9 genome editing. Circ. Res. 2014, 115, 488–492. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Smith, J.; Breton, C.; Clark, P.; Zhang, J.; Ying, L.; Che, Y.; Lape, J.; Bell, P.; Calcedo, R.; et al. Meganuclease targeting of pcsk9 in macaque liver leads to stable reduction in serum cholesterol. Nat. Biotechnol. 2018, 36, 717–725. [Google Scholar] [CrossRef]
- Pankowicz, F.P.; Barzi, M.; Legras, X.; Hubert, L.; Mi, T.; Tomolonis, J.A.; Ravishankar, M.; Sun, Q.; Yang, D.; Borowiak, M.; et al. Reprogramming metabolic pathways in vivo with crispr/cas9 genome editing to treat hereditary tyrosinaemia. Nat. Commun. 2016, 7, 12642. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, L.; Bell, P.; McMenamin, D.; He, Z.; White, J.; Yu, H.; Xu, C.; Morizono, H.; Musunuru, K.; et al. A dual aav system enables the cas9-mediated correction of a metabolic liver disease in newborn mice. Nat. Biotechnol. 2016, 34, 334–338. [Google Scholar] [CrossRef] [PubMed]
- Bengtsson, N.E.; Hall, J.K.; Odom, G.L.; Phelps, M.P.; Andrus, C.R.; Hawkins, R.D.; Hauschka, S.D.; Chamberlain, J.R.; Chamberlain, J.S. Muscle-specific crispr/cas9 dystrophin gene editing ameliorates pathophysiology in a mouse model for duchenne muscular dystrophy. Nat. Commun. 2017, 8, 14454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendell, J.R.; Rodino-Klapac, L.R. Duchenne muscular dystrophy: Crispr/cas9 treatment. Cell Res. 2016, 26, 513–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, C.E.; Hakim, C.H.; Ousterout, D.G.; Thakore, P.I.; Moreb, E.A.; Castellanos Rivera, R.M.; Madhavan, S.; Pan, X.; Ran, F.A.; Yan, W.X.; et al. In vivo genome editing improves muscle function in a mouse model of duchenne muscular dystrophy. Science 2016, 351, 403–407. [Google Scholar] [CrossRef] [Green Version]
- Daya, S.; Berns, K.I. Gene therapy using adeno-associated virus vectors. Clin. Microbiol. Rev. 2008, 21, 583–593. [Google Scholar] [CrossRef] [Green Version]
- Tycko, J.; Myer, V.E.; Hsu, P.D. Methods for optimizing crispr-cas9 genome editing specificity. Mol. Cell 2016, 63, 355–370. [Google Scholar] [CrossRef] [Green Version]
- Ran, F.A.; Cong, L.; Yan, W.X.; Scott, D.A.; Gootenberg, J.S.; Kriz, A.J.; Zetsche, B.; Shalem, O.; Wu, X.; Makarova, K.S.; et al. In vivo genome editing using staphylococcus aureus cas9. Nature 2015, 520, 186–191. [Google Scholar] [CrossRef]
- Kim, E.; Koo, T.; Park, S.W.; Kim, D.; Kim, K.; Cho, H.Y.; Song, D.W.; Lee, K.J.; Jung, M.H.; Kim, S.; et al. In vivo genome editing with a small cas9 orthologue derived from campylobacter jejuni. Nat Commun. 2017, 8, 14500. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Su, J.; Liu, Y.; Jin, X.; Zhong, X.M.; Mo, L.; Wang, Q.N.; Deng, H.X.; Yang, Y. In vivo pcsk9 gene editing using an all-in-one self-cleavage aav-crispr system. Mol. Ther.-Methods Clin. Dev. 2021, 20, 652–659. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. Interleukin-1 and tumor necrosis factor: Effector cytokines in autoimmune diseases. Semin. Immunol. 1992, 4, 133–145. [Google Scholar] [PubMed]
- Garlanda, C.; Dinarello, C.A.; Mantovani, A. The interleukin-1 family: Back to the future. Immunity 2013, 39, 1003–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishida, A.; Hidaka, K.; Kanda, T.; Imaeda, H.; Shioya, M.; Inatomi, O.; Bamba, S.; Kitoh, K.; Sugimoto, M.; Andoh, A. Increased expression of interleukin-36, a member of the interleukin-1 cytokine family, in inflammatory bowel disease. Inflamm. Bowel Dis. 2016, 22, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Ciccia, F.; Accardo-Palumbo, A.; Alessandro, R.; Alessandri, C.; Priori, R.; Guggino, G.; Raimondo, S.; Carubbi, F.; Valesini, G.; Giacomelli, R.; et al. Interleukin-36alpha axis is modulated in patients with primary sjogren’s syndrome. Clin. Exp. Immunol. 2015, 181, 230–238. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Wang, B.; Ma, Z.; Sun, X.; Tang, Y.; Li, X.; Wu, X. Detection of the novel il-1 family cytokines by qah-il1f-1 assay in rheumatoid arthritis. Cell Mol. Biol. 2016, 62, 31–34. [Google Scholar]
- Swindell, W.R.; Beamer, M.A.; Sarkar, M.K.; Loftus, S.; Fullmer, J.; Xing, X.; Ward, N.L.; Tsoi, L.C.; Kahlenberg, M.J.; Liang, Y.; et al. Rna-seq analysis of il-1b and il-36 responses in epidermal keratinocytes identifies a shared myd88-dependent gene signature. Front. Immunol. 2018, 9, 80. [Google Scholar] [CrossRef] [Green Version]
- Shembade, N.; Ma, A.; Harhaj, E.W. Inhibition of nf-kappab signaling by a20 through disruption of ubiquitin enzyme complexes. Science 2010, 327, 1135–1139. [Google Scholar] [CrossRef] [Green Version]
- Stahl, E.A.; Raychaudhuri, S.; Remmers, E.F.; Xie, G.; Eyre, S.; Thomson, B.P.; Li, Y.; Kurreeman, F.A.; Zhernakova, A.; Hinks, A.; et al. Genome-wide association study meta-analysis identifies seven new rheumatoid arthritis risk loci. Nat. Genet. 2010, 42, 508–514. [Google Scholar] [CrossRef]
- Zhang, M.; Peng, L.L.; Wang, Y.; Wang, J.S.; Liu, J.; Liu, M.M.; Hu, J.; Song, B.; Yang, H.B. Roles of a20 in autoimmune diseases. Immunol. Res. 2016, 64, 337–344. [Google Scholar] [CrossRef]
- Bulatov, E.; Khaiboullina, S.; dos Reis, H.J.; Palotás, A.; Venkataraman, K.; Vijayalakshmi, M.; Rizvanov, A. Mookambeswaran Vijayalakshmi & Albert Rizvanov Ubiquitin-proteasome system: Promising therapeutic targets in autoimmune and neurodegenerative diseases. BioNanoScience 2016, 6, 341–344. [Google Scholar] [CrossRef]
- Zhou, Q.; Wang, H.; Schwartz, D.M.; Stoffels, M.; Park, Y.H.; Zhang, Y.; Yang, D.; Demirkaya, E.; Takeuchi, M.; Tsai, W.L.; et al. Loss-of-function mutations in tnfaip3 leading to a20 haploinsufficiency cause an early-onset autoinflammatory disease. Nat. Genet. 2016, 48, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Huang, Y.; Ye, Z.; Wang, Y.; Tang, Z.; Lu, J.; Wu, J.; Zhou, Y.; Wang, L.; Huang, Z.; et al. Infantile onset intractable inflammatory bowel disease due to novel heterozygous mutations in tnfaip3 (a20). Inflamm. Bowel Dis. 2018, 24, 2613–2620. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wen, F.; Tessneer, K.L.; Gaffney, P.M. Talen-mediated enhancer knockout influences tnfaip3 gene expression and mimics a molecular phenotype associated with systemic lupus erythematosus. Genes Immun. 2016, 17, 165–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Yang, S.; Yu, D.; Gao, W.; Liu, X.; Zhang, K.; Fu, X.; Bao, W.; Zhang, K.; Yu, J.; et al. Crispr/cas9 mediated knockout of an intergenic variant rs6927172 identified il-20ra as a new risk gene for multiple autoimmune diseases. Genes Immun. 2018, 20, 103–111. [Google Scholar] [CrossRef]
- Fontenot, J.D.; Rasmussen, J.P.; Williams, L.M.; Dooley, J.L.; Farr, A.G.; Rudensky, A.Y. Regulatory t cell lineage specification by the forkhead transcription factor foxp3. Immunity 2005, 22, 329–341. [Google Scholar] [CrossRef]
- Bennett, C.L.; Christie, J.; Ramsdell, F.; Brunkow, M.E.; Ferguson, P.J.; Whitesell, L.; Kelly, T.E.; Saulsbury, F.T.; Chance, P.F.; Ochs, H.D. The immune dysregulation, polyendocrinopathy, enteropathy, x-linked syndrome (ipex) is caused by mutations of foxp3. Nat. Genet. 2001, 27, 20–21. [Google Scholar] [CrossRef]
- Jeffries, M.A. Epigenetic editing: How cutting-edge targeted epigenetic modification might provide novel avenues for autoimmune disease therapy. Clin. Immunol. 2018, 196, 49–58. [Google Scholar] [CrossRef]
- Brunger, J.M.; Zutshi, A.; Willard, V.P.; Gersbach, C.A.; Guilak, F. Genome engineering of stem cells for autonomously regulated, closed-loop delivery of biologic drugs. Stem Cell Rep. 2017, 8, 1202–1213. [Google Scholar] [CrossRef] [Green Version]
- Brunger, J.M.; Zutshi, A.; Willard, V.P.; Gersbach, C.A.; Guilak, F. Crispr/cas9 editing of murine induced pluripotent stem cells for engineering inflammation-resistant tissues. Arthritis Rheumatol. 2017, 69, 1111–1121. [Google Scholar] [CrossRef]
- Gerace, D.; Martiniello-Wilks, R.; Nassif, N.T.; Lal, S.; Steptoe, R.; Simpson, A.M. Crispr-targeted genome editing of mesenchymal stem cell-derived therapies for type 1 diabetes: A path to clinical success? Stem Cell Res. 2017, 8, 62. [Google Scholar] [CrossRef] [Green Version]
- Gimenez, C.A.; Ielpi, M.; Mutto, A.; Grosembacher, L.; Argibay, P.; Pereyra-Bonnet, F. Crispr-on system for the activation of the endogenous human ins gene. Gene 2016, 23, 543–547. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; McGovern, A.; Martin, P.; Duffus, K.; Ge, X.; Zarrineh, P.; Morris, A.P.; Adamson, A.; Fraser, P.; Rattray, M.; et al. Analysis of chromatin organization and gene expression in t cells identifies functional genes for rheumatoid arthritis. Nat. Commun. 2020, 11, 4402. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Jin, Z.; Liu, X.; Gao, W.; Gao, X.; Zhang, S.; Ma, S.; Yu, J.; Wang, S. Manipulation of an ra-associated intergenic snp by crispr/cas9 system alters expression of multiple genes at the tnfaip3 locus, poster presentation abstracts. Int. J. Rheum. Dis. 2016, 19 (Suppl. S2), 89. [Google Scholar] [CrossRef] [Green Version]
- Jing, W.; Zhang, X.; Sun, W.; Hou, X.; Yao, Z.; Zhu, Y. Crispr/cas9-mediated genome editing of mirna-155 inhibits proinflammatory cytokine production by raw264.7 cells. Biomed. Res. Int. 2015, 2015, 326042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardinale, C.J.; March, M.E.; Lin, X.; Liu, Y.; Spruce, L.A.; Bradfield, J.P.; Wei, Z.; Seeholzer, S.H.; Grant, S.F.A.; Hakonarson, H. Regulation of janus kinase 2 by an inflammatory bowel disease causal non-coding single nucleotide polymorphism. J. Crohns Colitis 2020, 14, 646–653. [Google Scholar] [CrossRef] [PubMed]
- Mokhtar, N.; Den Low, E.N.; Yahaya, A.; Ali, R.A.R. Characterisation of sgk2 gene using crispr/cas9 gene editing tool in ulcerative colitis. J. Gastroenterol. Hepatol. 2019, 34 (Suppl. S3), 274. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Kuemmerle, J. P119 the role of ptpn2 snp in the pathogenesis of fibrosis in crohn’s disease. Inflamm. Bowel Dis. 2019, 25, S57–S58. [Google Scholar] [CrossRef]
- Matthews, S.M.; Eshelman, M.A.; Berg, A.S.; Koltun, W.A.; Yochum, G.S. The crohn’s disease associated snp rs6651252 impacts myc gene expression in human colonic epithelial cells. PLoS ONE 2019, 14, e0212850. [Google Scholar] [CrossRef]
- Pai, Y.-C.; Shih, D.Q.; Targan, S.R.; Turner, J.; Wei, S.C.; Yu, L.C.H. Su1097 tl1a-mediated bacterial transcytosis by a mlck splicing variant contributes to inflammatory bowel disease. Gastroenterology 2020, 158, S-508. [Google Scholar] [CrossRef]
- Friedrich, M.; Ganther, J.; Breiderhoff, T.; Rosenthal, R.; Glauben, R.; Siegmund, B. P063 hdac as versatile regulators of the intestinal epithelial barrier in inflammatory bowel disease. J. Crohns Colitis 2017, 11, S109. [Google Scholar] [CrossRef] [Green Version]
- Eftychi, C.; Schwarzer, R.; Vlantis, K.; Wachsmuth, L.; Basic, M.; Wagle, P.; Neurath, M.F.; Becker, C.; Bleich, A.; Pasparakis, M. Temporally distinct functions of the cytokines il-12 and il-23 drive chronic colon inflammation in response to intestinal barrier impairment. Immunity 2019, 51, 367–380.e4. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.D.; Sun, M.M.; Wu, W.; Ma, C.Y.; Zhang, C.; He, C.; Li, J.X.; Cong, Y.Z.; Zhang, D.K.; Liu, Z.J. Microrna-125a suppresses intestinal mucosal inflammation through targeting ets-1 in patients with inflammatory bowel diseases. J. Autoimmun. 2019, 101, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Odqvist, L.; Jevnikar, Z.; Riise, R.; Oberg, L.; Rhedin, M.; Leonard, D.; Yrlid, L.; Jackson, S.; Mattsson, J.; Nanda, S.; et al. Genetic variations in a20 dub domain provide a genetic link to citrullination and neutrophil extracellular traps in systemic lupus erythematosus. Ann. Rheum. Dis. 2019, 78, 1363–1370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, V.M.; Koelsch, K.A.; Kurien, B.T.; Harley, I.T.W.; Wren, J.D.; Harley, J.B.; Scofield, R.H. Characterization of cxorf21 provides molecular insight into female-bias immune response in sle pathogenesis. Front. Immunol. 2019, 10, 2160. [Google Scholar] [CrossRef]
- Gregory, S.G.; Schmidt, S.; Seth, P.; Oksenberg, J.R.; Hart, J.; Prokop, A.; Caillier, S.J.; Ban, M.; Goris, A.; Barcellos, L.F.; et al. Interleukin 7 receptor alpha chain (il7r) shows allelic and functional association with multiple sclerosis. Nat. Genet. 2007, 39, 1083–1091. [Google Scholar] [CrossRef]
- Galarza-Munoz, G.; Briggs, F.B.S.; Evsyukova, I.; Schott-Lerner, G.; Kennedy, E.M.; Nyanhete, T.; Wang, L.; Bergamaschi, L.; Widen, S.G.; Tomaras, G.D.; et al. Human epistatic interaction controls il7r splicing and increases multiple sclerosis risk. Cell 2017, 169, 72–84.e13. [Google Scholar] [CrossRef] [Green Version]
- Maier, L.M.; Lowe, C.E.; Cooper, J.; Downes, K.; Anderson, D.E.; Severson, C.; Clark, P.M.; Healy, B.; Walker, N.; Aubin, C.; et al. Il2ra genetic heterogeneity in multiple sclerosis and type 1 diabetes susceptibility and soluble interleukin-2 receptor production. PLoS Genet. 2009, 5, e1000322. [Google Scholar] [CrossRef]
- Gregory, A.P.; Dendrou, C.A.; Attfield, K.E.; Haghikia, A.; Xifara, D.K.; Butter, F.; Poschmann, G.; Kaur, G.; Lambert, L.; Leach, O.A.; et al. Tnf receptor 1 genetic risk mirrors outcome of anti-tnf therapy in multiple sclerosis. Nature 2012, 488, 508–511. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Q.; Wang, J.; Zhang, L.; Bian, W.; Lin, M.; Xu, X.; Zhou, X. Lck rs10914542-g allele associates with type 1 diabetes in children via t cell hyporesponsiveness. Pediatr. Res. 2019, 86, 311–315. [Google Scholar] [CrossRef]
- Ratiu, J.J.; Racine, J.J.; Hasham, M.G.; Wang, Q.; Branca, J.A.; Chapman, H.D.; Zhu, J.; Donghia, N.; Philip, V.; Schott, W.H.; et al. Genetic and small molecule disruption of the aid/rad51 axis similarly protects nonobese diabetic mice from type 1 diabetes through expansion of regulatory b lymphocytes. J. Immunol. 2017, 198, 4255–4267. [Google Scholar] [CrossRef] [Green Version]
- Arakawa, A.; Vollmer, S.; Arakawa, Y.; He, M.; Reeves, E.; James, E.; Prinz, J. Erap1 Regulates the Risk for Psoriasis through Affecting Immunogenicity of Melanocytes and the Psoriatic Autoantigen Adamts-Like Protein 5. In Proceedings of the 46th Annual Meeting of the Arbeitsgemeinschaft Dermatologische Forschung (ADF), Munich, Germany, 13–16 March 2019; Wiley: Munich, Germany, 2019; pp. E66–E67. [Google Scholar]
- Roth-Carter, Q.R.; Godsel, L.; Koetsier, J.L.; Broussard, J.A.; Burks, H.E.; Fitz, G.; Huffine, A.L.; Amagai, S.; Lloyd, S.; Kweon, J.; et al. 225 desmoglein 1 deficiency in knockout mice impairs epidermal barrier formation and results in a psoriasis-like gene signature in e18. 5 embryos. J. Investig. Dermatol. 2020, 140, S26. [Google Scholar] [CrossRef]
- Jouanin, A.; Schaart, J.G.; Boyd, L.A.; Cockram, J.; Leigh, F.J.; Bates, R.; Wallington, E.J.; Visser, R.G.F.; Smulders, M.J.M. Outlook for coeliac disease patients: Towards bread wheat with hypoimmunogenic gluten by gene editing of alpha- and gamma-gliadin gene families. BMC Plant Biol. 2019, 19, 333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Xu, J.; Ge, S.; Lai, L. Crispr/cas: Advances, limitations, and applications for precision cancer research. Front. Med. 2021, 8. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Study | Autoimmune Disease | Target Gene | SNP, miRNA | Results | Ref |
|---|---|---|---|---|---|
| Yang et al., 2020 | RA | MYC, FOXO1 | - | By collecting ATAC-seq, Hi-C, Capture Hi-C, and nuclear RNA-seq data in stimulated CD4+ T cells over 24 hours, the study provides evidence that MYC and FOXO1 genes may be causal factors of RA. | [42] |
| Yu et al., 2016 | RA | IL20RA, IL22RA2, IFNGR1, OLIG3, TNFAIP3 | SNP rs6927172 | SNP rs6927172, TNFAIP3, and OLIG3 are importantly associated with the RA disease course. | [43] |
| Jing et al., 2015 | RA | - | microRNA 155 (miR-155) | Genome editing of miR-155 can be a potential therapeutic strategy for RA. | [44] |
| Cardinale et al., 2020 | IBD | - | SNP rs1887428 | rs1887428 does not have very large impacts on JAK2 expression, but it impacts STAT5B downwards. | [45] |
| Mokhtar et al., 2019 | IBD | SGK2 | - | SGK2 protein was localized in cytoplasm of colonic epithelial cells both in long and short duration ulcerative colitis. | [46] |
| Li et al., 2018 | IBD | PTPN2 | SNP rs7234029 | PTPN2 expression was increased in affected ileum compared to normal ileum and, in contrast, CRISPR-Cas9 mediated PTPN2 deletion resulted in higher levels of STAT3 and Erk1/2 phosphorylation and proliferation. | [47] |
| Matthews et al., 2019 | IBD | - | SNP rs6651252 | Using CRISPR-Cas9, they found that rs6651252 enhancer regulates expression of the c-MYC, and they also revealed that MYC expression levels are elevated in the Crohn’s disease patients. | [48] |
| Odqvist et al., 2019 | SLE | TNFAIP3 | SNP rs2230926 | The result showed that A20 C103A cells or cells with rs2230926 polymorphism draw increased neutrophil extracellular trap and increased frequency of auto antibodies to citrullinated epitopes. | [53] |
| Harris et al., 2019 | SLE | CXorf21 | - | They have done in vitro CRISPR-Cas9 knockdown experiments and found that CXorf21 knockdown resulted a decreased expression of TNF-alpha and IL-6. | [54] |
| Gregory et al., 2007 | MS | IL7R | SNP rs6897932 | There is a direct relationship between alternative rs6897932 alleles and MS risk. | [55] |
| Galarza-Munoz et al., 2017 | MS | DDX39B | SNP rs2523506 | It was shown that epistatic interaction between rs2523506 in DDX39B and rs6897932 in IL7R regulates IL7R exon 6 splicing and increases risk of MS. | [56] |
| Maier et al., 2009 | MS | IL-2RA | - | IL2RA variants contributed to the risk of MS and T1D respectively. | [57] |
| Gregory et al., 2012 | MS | TNFRSF1A | SNP rs1800693 | TNF blocking agents have side effects that promote the onset of MS, when it is used in other autoimmune diseases such as RA, but this study found that a signal of disease association for rs1800693 is predictive of the side effect. | [58] |
| Zhu et al., 2019 | T1DM | LCK | SNP rs10914542 | They demonstrated that G allele of SNP rs10914542 of LCK increases the risk of Type 1 DM. | [59] |
| Arakawa et al., 2019 | Psoriasis | ERAP1 | - | They generated ERAP1 knockout melanoma cell line and revealed that epistasis between HLA-C*06:02 and ERAP1 variants will affect psoriasis. | [61] |
| Jouanin et al., 2019 | Coeliac disease | α- or γ-gliadin | - | The result showed that it is possible to use CRISPR-Cas9 to edit α- or γ-gliadins and make safe forms of grains. | [63] |
| Study | Autoimmune Disease | Target Gene | SNP, miRNA | Results | Ref |
|---|---|---|---|---|---|
| Pai et al., 2020 | IBD | TL1A | - | The flare-up of TL1A, IFNγ, and MLCK is associated with IBD diseases course. | [49] |
| Friedrich et al., 2017 | IBD | HDAC7 | - | The result showed that HDAC function was reduced in IBD patients, and according to the knockout mice, HDAC were found to play a crucial role in maintenance of the intestinal barrier. | [50] |
| Eftychi et al., 2019 | IBD | IFN-γ | - | They found out that many pro-inflammatory cytokines such as TNF, IL1b, and IL6 showed decreased expressivity in the KO mice and suggested that IFN-γ is essential for colon inflammation. | [51] |
| Ge et al., 2019 | IBD | - | miRNA-125a | MicroRNA-125a suppresses Th1/Th17 cell differentiation and TNF-α production. Additionally, miR-125a knockout mice showed more severe forms of colitis. | [52] |
| Ratiu et al., 2017 | T1DM | AID/RAD51 | - | AID/RAD51 can be a target for treatment of Type 1 DM patients. | [60] |
| Roth-Carter et al., 2020 | Psoriasis | Desmoglein 1 | - | They supported a role of Dsg1 for differentiation of epidermis, formation of barriers, and regulation of inflammatory responses. | [62] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, M.H.; Shin, J.I.; Yang, J.W.; Lee, K.H.; Cha, D.H.; Hong, J.B.; Park, Y.; Choi, E.; Tizaoui, K.; Koyanagi, A.; et al. Genome Editing Using CRISPR-Cas9 and Autoimmune Diseases: A Comprehensive Review. Int. J. Mol. Sci. 2022, 23, 1337. https://doi.org/10.3390/ijms23031337
Lee MH, Shin JI, Yang JW, Lee KH, Cha DH, Hong JB, Park Y, Choi E, Tizaoui K, Koyanagi A, et al. Genome Editing Using CRISPR-Cas9 and Autoimmune Diseases: A Comprehensive Review. International Journal of Molecular Sciences. 2022; 23(3):1337. https://doi.org/10.3390/ijms23031337
Chicago/Turabian StyleLee, Min Ho, Jae Il Shin, Jae Won Yang, Keum Hwa Lee, Do Hyeon Cha, Jun Beom Hong, Yeoeun Park, Eugene Choi, Kalthoum Tizaoui, Ai Koyanagi, and et al. 2022. "Genome Editing Using CRISPR-Cas9 and Autoimmune Diseases: A Comprehensive Review" International Journal of Molecular Sciences 23, no. 3: 1337. https://doi.org/10.3390/ijms23031337
APA StyleLee, M. H., Shin, J. I., Yang, J. W., Lee, K. H., Cha, D. H., Hong, J. B., Park, Y., Choi, E., Tizaoui, K., Koyanagi, A., Jacob, L., Park, S., Kim, J. H., & Smith, L. (2022). Genome Editing Using CRISPR-Cas9 and Autoimmune Diseases: A Comprehensive Review. International Journal of Molecular Sciences, 23(3), 1337. https://doi.org/10.3390/ijms23031337