
Induced Neurodifferentiation of hBM-MSCs through Activation of the ERK/CREB Pathway via Pulsed Electromagnetic Fields and Physical Stimulation Promotes Neurogenesis in Cerebral Ischemic Models


Abstract
:1. Introduction
2. Results
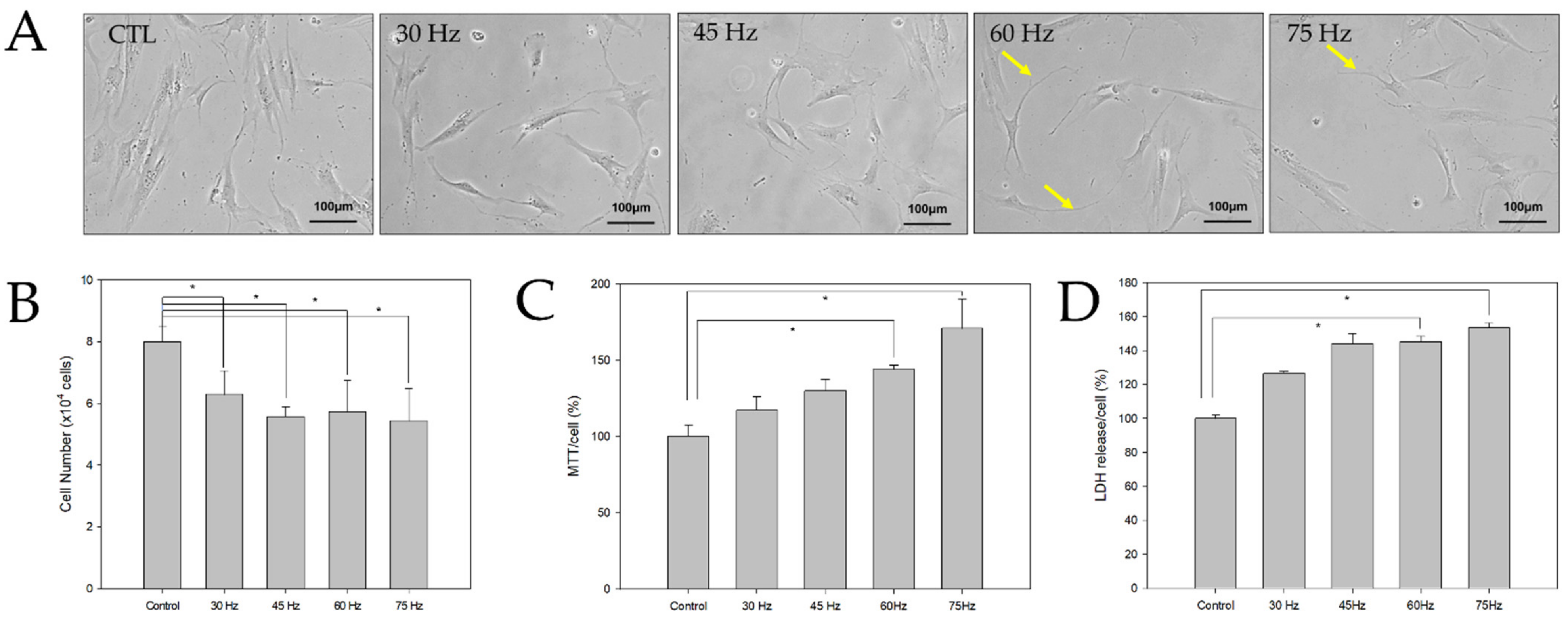
2.1. Morphological and Toxic Effects of PEMF Exposure on hBM-MSCs
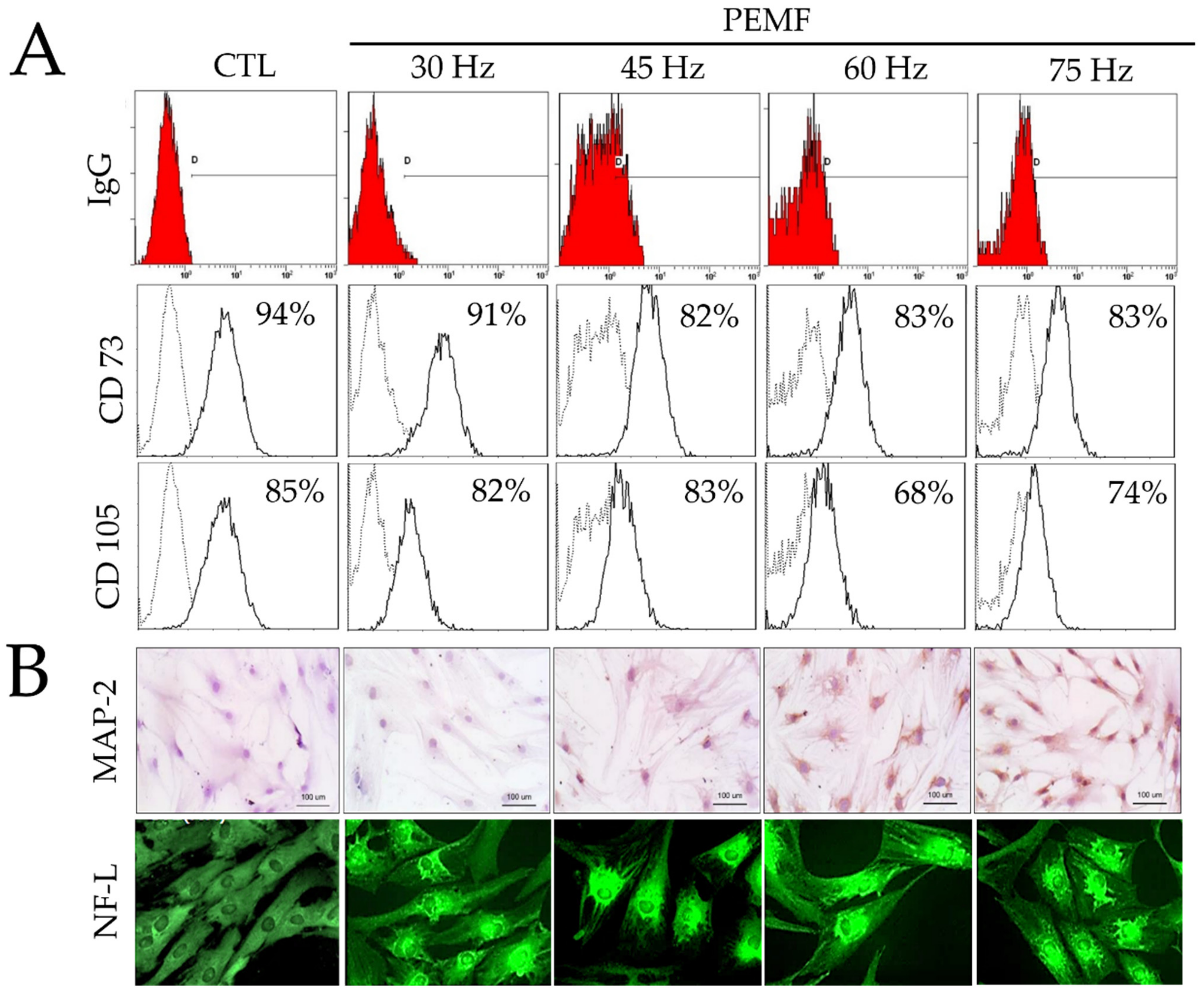
2.2. FACS Analysis
2.3. Immunohistochemical Staining
2.4. RT-PCR and Western Blotting
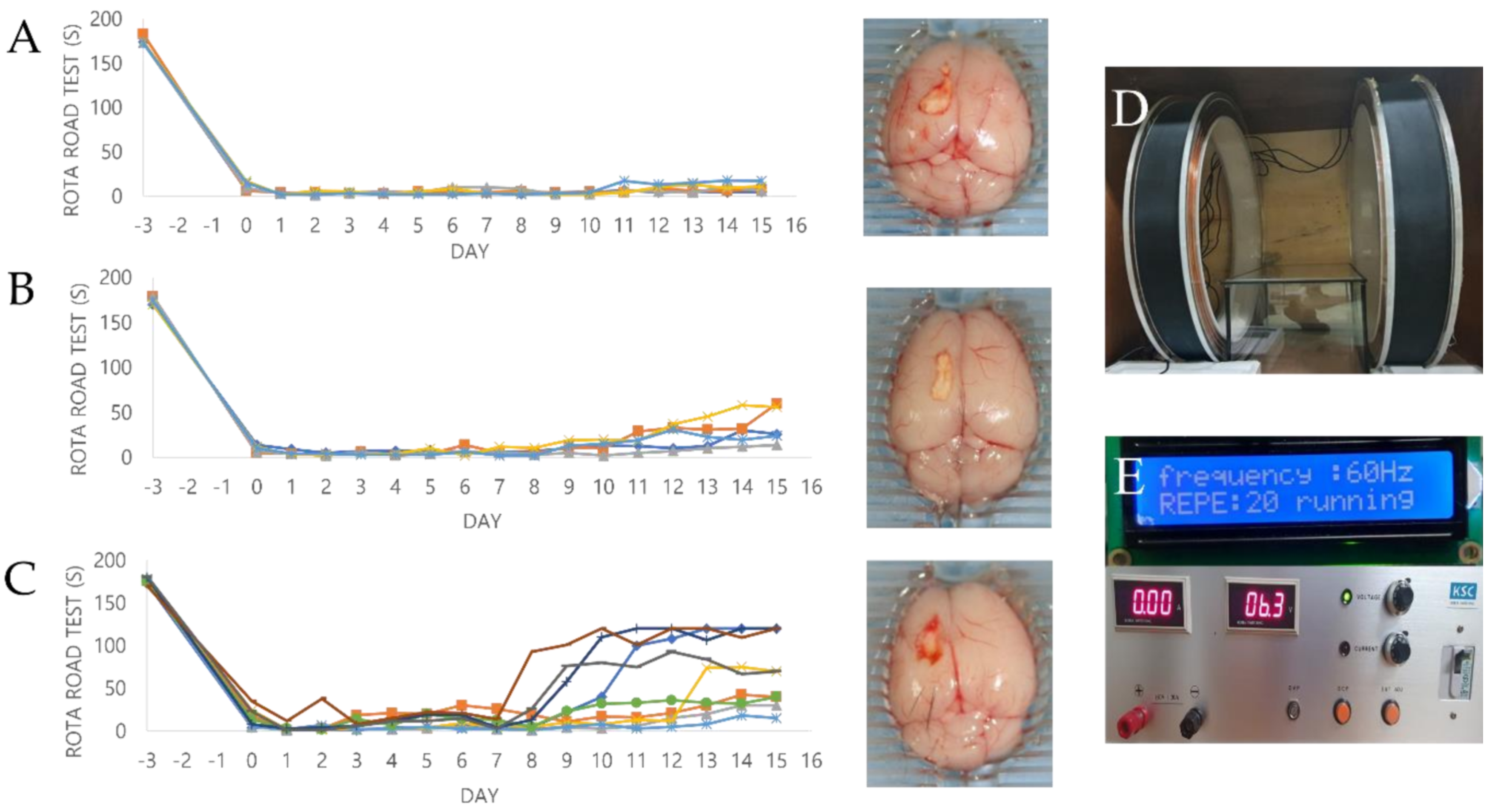
2.5. Effect of PEMF Exposure on Motor Function in a Mouse Ischemia Model
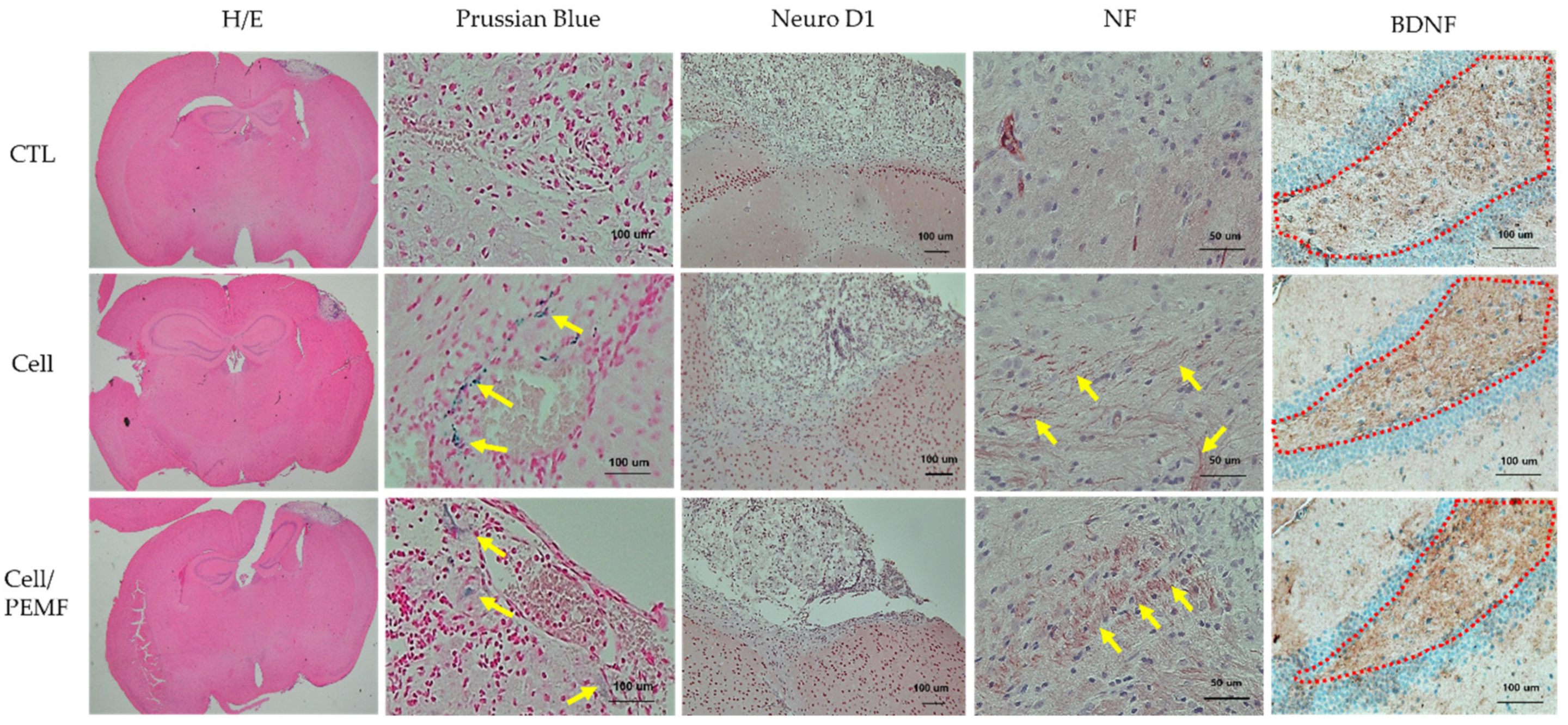
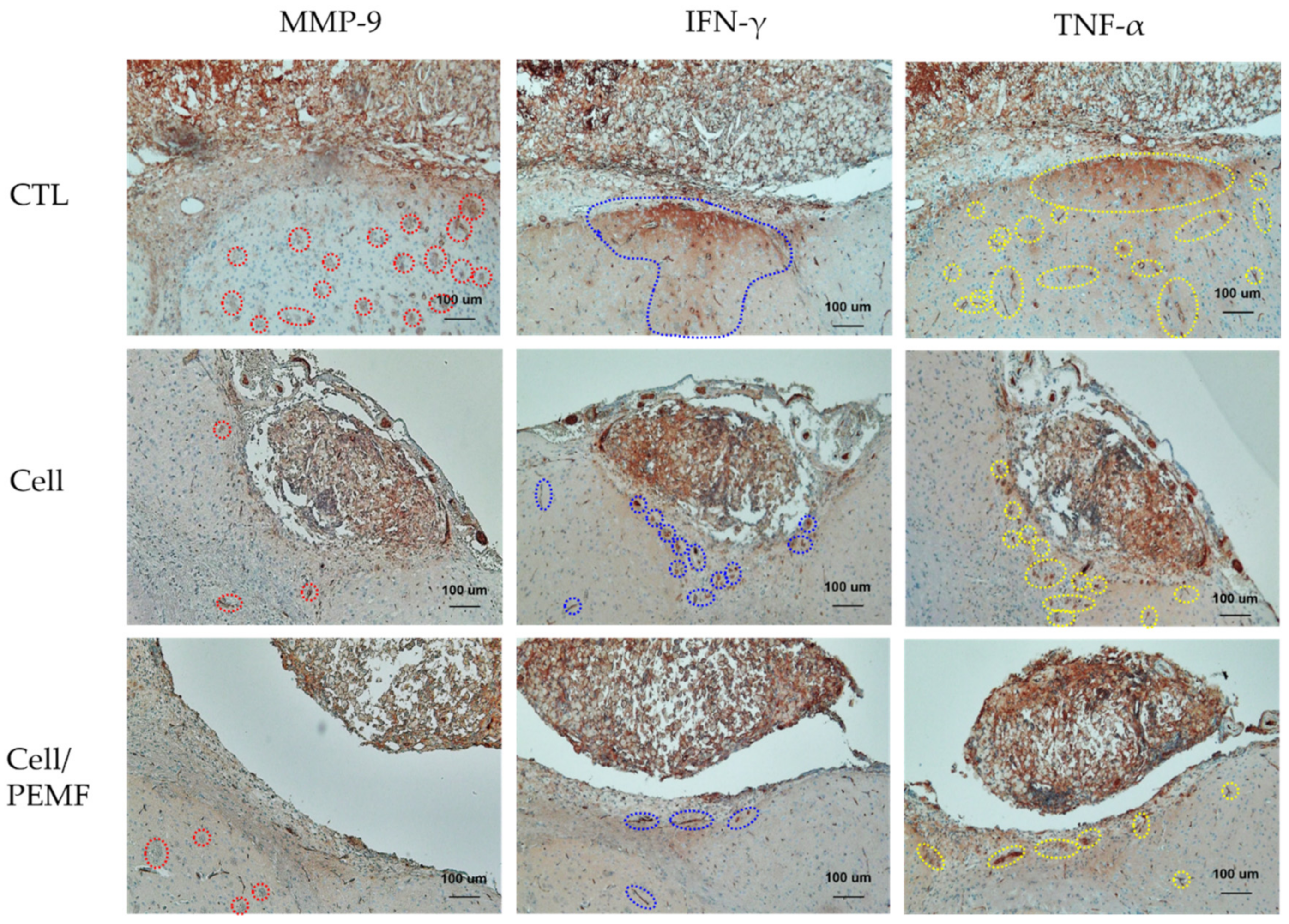
2.6. Evaluation of Neural Protein Expression and Histological Analysis after PEMF Exposure in the Mouse Stroke Model
3. Discussion
4. Materials and Methods
4.1. Culture and Neural Induction of hBM-MSCs
4.2. Design of the Magnetic Generator
4.3. Exposure of hBM-MSCs to Pulsed Electromagnetic Fields
4.4. hBM-MSC Growth Assay
4.5. Fluorescence-Activated Cell Sorting (FACS) Analysis
4.6. Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
4.7. Preparation of Experimental Animals
4.8. Western Blot Analysis
4.9. Histological and Immunohistological Analysis
4.10. Rotarod Test
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Avan, A.; Digaleh, H.; Di Napoli, M.; Stranges, S.; Behrouz, R.; Shojaeianbabaei, G.; Amiri, A.; Tabrizi, R.; Mokhber, N.; Spence, J.D.; et al. Socioeconomic status and stroke incidence, prevalence, mortality, and worldwide burden: An ecological analysis from the Global Burden of Disease Study 2017. BMC Med. 2019, 17, 191. [Google Scholar] [CrossRef] [PubMed]
- Donnan, G.A.; Fisher, M.; Macleod, M.; Davis, S.M. Stroke. Lancet 2008, 371, 1612–1623. [Google Scholar] [CrossRef]
- Virani, S.S.; Alonso, A.; Benjamin, E.J.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Chang, A.R.; Cheng, S.; Delling, F.N.; et al. Heart Disease and Stroke Statistics-2020 Update: A Report From the American Heart Association. Circulation 2020, 141, e139–e596. [Google Scholar] [CrossRef]
- Zhang, J.; Chopp, M. Cell-based therapy for ischemic stroke. Expert Opin. Biol. Ther. 2013, 13, 1229–1240. [Google Scholar] [CrossRef] [Green Version]
- Salem, H.K.; Thiemermann, C. Mesenchymal stromal cells: Current understanding and clinical status. Stem Cells 2010, 28, 585–596. [Google Scholar] [CrossRef] [Green Version]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.J.; Li, X.L.; Xue, Y.; Zhang, C.X.; Wang, Y.; Hu, X.; Dai, Q. Bone marrow cells differentiation into organ cells using stem cell therapy. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 2899–2907. [Google Scholar]
- Black, I.B.; Woodbury, D. Adult rat and human bone marrow stromal stem cells differentiate into neurons. Blood Cells Mol. Dis. 2001, 27, 632–636. [Google Scholar] [CrossRef]
- Kim, B.J.; Seo, J.H.; Bubien, J.K.; Oh, Y.S. Differentiation of adult bone marrow stem cells into neuroprogenitor cells in vitro. Neuroreport 2002, 13, 1185–1188. [Google Scholar] [CrossRef]
- Long, X.; Olszewski, M.; Huang, W.; Kletzel, M. Neural cell differentiation in vitro from adult human bone marrow mesenchymal stem cells. Stem Cells Dev. 2005, 14, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Boese, A.C.; Le, Q.E.; Pham, D.; Hamblin, M.H.; Lee, J.P. Neural stem cell therapy for subacute and chronic ischemic stroke. Stem Cell Res. Ther. 2018, 9, 154. [Google Scholar] [CrossRef]
- Fernández-Susavila, H.; Bugallo-Casal, A.; Castillo, J.; Campos, F. Adult Stem Cells and Induced Pluripotent Stem Cells for Stroke Treatment. Front. Neurol. 2019, 10, 908. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Venkat, P.; Zacharek, A.; Chopp, M. Neurorestorative therapy for stroke. Front. Hum. Neurosci. 2014, 8, 382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hei, W.H.; Byun, S.H.; Kim, J.S.; Kim, S.; Seo, Y.K.; Park, J.C.; Kim, S.M.; Jahng, J.W.; Lee, J.H. Effects of electromagnetic field (PEMF) exposure at different frequency and duration on the peripheral nerve regeneration: In vitro and in vivo study. Int. J. Neurosci. 2016, 126, 739–748. [Google Scholar]
- Cheing, G.L.; Li, X.; Huang, L.; Kwan, R.L.; Cheung, K.K. Pulsed electromagnetic fields (PEMF) promote early wound healing and myofibroblast proliferation in diabetic rats. Bioelectromagnetics 2014, 35, 161–169. [Google Scholar] [CrossRef]
- Leone, L.; Fusco, S.; Mastrodonato, A.; Piacentini, R.; Barbati, S.A.; Zaffina, S.; Pani, G.; Podda, M.V.; Grassi, C. Epigenetic modulation of adult hippocampal neurogenesis by extremely low-frequency electromagnetic fields. Mol. Neurobiol. 2014, 49, 1472–1486. [Google Scholar] [CrossRef]
- Pena-Philippides, J.C.; Yang, Y.; Bragina, O.; Hagberg, S.; Nemoto, E.; Roitbak, T. Effect of pulsed electromagnetic field (PEMF) on infarct size and inflammation after cerebral ischemia in mice. Transl. Stroke Res. 2014, 5, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zuo, H.; Wang, D.; Peng, R.; Song, T.; Wang, S.; Xu, X.; Gao, Y.; Li, Y.; Wang, S.; et al. Improvement of spatial memory disorder and hippocampal damage by exposure to electromagnetic fields in an Alzheimer’s disease rat model. PLoS ONE 2015, 10, e0126963. [Google Scholar] [CrossRef] [Green Version]
- Jang, S.; Cho, H.H.; Cho, Y.B.; Park, J.S.; Jeong, H.S. Functional neural differentiation of human adipose tissue-derived stem cells using bFGF and forskolin. BMC Cell Biol. 2010, 11, 25. [Google Scholar] [CrossRef] [Green Version]
- Safford, K.M.; Hicok, K.C.; Safford, S.D.; Halvorsen, Y.D.; Wilkison, W.O.; Gimble, J.M.; Rice, H.E. Neurogenic differentiation of murine and human adipose-derived stromal cells. Biochem. Biophys. Res. Commun. 2002, 294, 371–379. [Google Scholar] [CrossRef]
- Qian, D.X.; Zhang, H.T.; Ma, X.; Jiang, X.D.; Xu, R.X. Comparison of the efficiencies of three neural induction protocols in human adipose stromal cells. Neurochem. Res. 2010, 35, 572–579. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, K.E.; Weiss, M.L.; Mitchell, B.M.; Martin, P.; Davis, D.; Morales, L.; Helwig, B.; Beerenstrauch, M.; Abou-Easa, K.; Hildreth, T.; et al. Matrix cells from Wharton’s jelly form neurons and glia. Stem Cells 2003, 21, 50–60. [Google Scholar] [CrossRef] [Green Version]
- Mabuchi, Y.; Houlihan, D.D.; Akazawa, C.; Okano, H.; Matsuzaki, Y. Prospective isolation of murine and human bone marrow mesenchymal stem cells based on surface markers. Stem Cells Int. 2013, 2013, 507301. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.P.; Tripathy, N.K.; Nityanand, S. Comparison of phenotypic markers and neural differentiation potential of multipotent adult progenitor cells and mesenchymal stem cells. World J. Stem Cells 2013, 5, 53–60. [Google Scholar] [CrossRef]
- Kléber, M.; Sommer, L. Wnt signaling and the regulation of stem cell function. Curr. Opin. Cell Biol. 2004, 16, 681–687. [Google Scholar] [CrossRef]
- Bräutigam, C.; Raggioli, A.; Winter, J. The Wnt/β-catenin pathway regulates the expression of the miR-302 cluster in mouse ESCs and P19 cells. PLoS ONE 2013, 8, e75315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, M.; Uyama, M.; Sugiyama, Y.; Sato, M. Canonical Wnt signaling activates miR-34 expression during osteoblastic differentiation. Mol. Med. Rep. 2013, 8, 1807–1811. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Li, Z.; Ding, R.; Li, H.; Zhang, L.; Yuan, W.; Wang, Y. Parathyroid hormone induces epithelial-to-mesenchymal transition via the Wnt/β-catenin signaling pathway in human renal proximal tubular cells. Int. J. Clin. Exp. Pathol. 2014, 7, 5978–5987. [Google Scholar] [PubMed]
- Piccin, D.; Morshead, C.M. Wnt signaling regulates symmetry of division of neural stem cells in the adult brain and in response to injury. Stem Cells 2011, 29, 528–538. [Google Scholar] [CrossRef]
- Shioda, N.; Han, F.; Fukunaga, K. Role of Akt and ERK signaling in the neurogenesis following brain ischemia. Int. Rev. Neurobiol. 2009, 85, 375–387. [Google Scholar] [PubMed]
- Huang, C.Y.; Liou, Y.F.; Chung, S.Y.; Lin, W.Y.; Jong, G.P.; Kuo, C.H.; Tsai, F.J.; Cheng, Y.C.; Cheng, F.C.; Lin, J.Y. Role of ERK signaling in the neuroprotective efficacy of magnesium sulfate treatment during focal cerebral ischemia in the gerbil cortex. Chin. J. Physiol. 2010, 53, 299–309. [Google Scholar] [CrossRef] [Green Version]
- Tzeng, H.H.; Hsu, C.H.; Chung, T.H.; Lee, W.C.; Lin, C.H.; Wang, W.C.; Hsiao, C.Y.; Leu, Y.W.; Wang, T.H. Cell Signaling and Differential Protein Expression in Neuronal Differentiation of Bone Marrow Mesenchymal Stem Cells with Hypermethylated Salvador/Warts/Hippo (SWH) Pathway Genes. PLoS ONE 2015, 10, e0145542. [Google Scholar] [CrossRef]
- Takahashi, J.; Palmer, T.D.; Gage, F.H. Retinoic acid and neurotrophins collaborate to regulate neurogenesis in adult-derived neural stem cell cultures. J. Neurobiol. 1999, 38, 65–81. [Google Scholar] [CrossRef]
- Palmer, T.D.; Takahashi, J.; Gage, F.H. The adult rat hippocampus contains primordial neural stem cells. Mol. Cell. Neurosci. 1997, 8, 389–404. [Google Scholar] [CrossRef]
- Lonze, B.E.; Ginty, D.D. Function and regulation of CREB family transcription factors in the nervous system. Neuron 2002, 35, 605–623. [Google Scholar] [CrossRef] [Green Version]
- Park, J.E.; Seo, Y.K.; Yoon, H.H.; Kim, C.W.; Park, J.K.; Jeon, S. Electromagnetic fields induce neural differentiation of human bone marrow derived mesenchymal stem cells via ROS mediated EGFR activation. Neurochem. Int. 2013, 62, 418–424. [Google Scholar] [CrossRef]
- Ułamek-Kozioł, M.; Czuczwar, S.J.; Januszewski, S.; Pluta, R. Proteomic and Genomic Changes in Tau Protein, Which Are Associated with Alzheimer’s Disease after Ischemia-Reperfusion Brain Injury. Int. J. Mol. Sci. 2020, 21, 892. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Jiang, H. Tau as a potential therapeutic target for ischemic stroke. Aging 2019, 11, 12827–12843. [Google Scholar] [CrossRef]
- Michalczyk, K.; Ziman, M. Nestin structure and predicted function in cellular cytoskeletal organisation. Histol. Histopathol. 2005, 20, 665–671. [Google Scholar] [PubMed]
- Gao, Z.; Ure, K.; Ables, J.L.; Lagace, D.C.; Nave, K.A.; Goebbels, S.; Eisch, A.J.; Hsieh, J. Neurod1 is essential for the survival and maturation of adult-born neurons. Nat. Neurosci. 2009, 12, 1090–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuwabara, T.; Hsieh, J.; Muotri, A.; Yeo, G.; Warashina, M.; Lie, D.C.; Moore, L.; Nakashima, K.; Asashima, M.; Gage, F.H. Wnt-mediated activation of NeuroD1 and retro-elements during adult neurogenesis. Nat. Neurosci. 2009, 12, 1097–1105. [Google Scholar] [CrossRef] [Green Version]
- Boutin, C.; Hardt, O.; de Chevigny, A.; Coré, N.; Goebbels, S.; Seidenfaden, R.; Bosio, A.; Cremer, H. NeuroD1 induces terminal neuronal differentiation in olfactory neurogenesis. Proc. Natl. Acad. Sci. USA 2010, 107, 1201–1206. [Google Scholar] [CrossRef] [Green Version]
- Abbasnia, K.; Ghanbari, A.; Abedian, M.; Ghanbari, A.; Sharififar, S.; Azari, H. The effects of repetitive transcranial magnetic stimulation on proliferation and differentiation of neural stem cells. Anat. Cell Biol. 2015, 48, 104–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Chen, J.; Zhang, C.L.; Wang, L.; Lu, D.; Katakowski, M.; Gao, Q.; Shen, L.H.; Zhang, J.; Lu, M.; et al. Gliosis and brain remodeling after treatment of stroke in rats with marrow stromal cells. Glia 2005, 49, 407–417. [Google Scholar] [CrossRef]
- Shen, L.H.; Li, Y.; Chen, J.; Zhang, J.; Vanguri, P.; Borneman, J.; Chopp, M. Intracarotid transplantation of bone marrow stromal cells increases axon-myelin remodeling after stroke. Neuroscience 2006, 137, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Li, Y.; Qu, R.; Shen, L.; Gao, Q.; Zhang, X.; Lu, M.; Savant-Bhonsale, S.; Borneman, J.; Chopp, M. Axonal sprouting into the denervated spinal cord and synaptic and postsynaptic protein expression in the spinal cord after transplantation of bone marrow stromal cell in stroke rats. Brain Res. 2007, 1149, 172–180. [Google Scholar] [CrossRef] [Green Version]
- Pesce, M.; Patruno, A.; Speranza, L.; Reale, M. Extremely low frequency electromagnetic field and wound healing: Implication of cytokines as biological mediators. Eur. Cytokine Netw. 2013, 24, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Cui, N.; Hu, M.; Khalil, R.A. Biochemical and Biological Attributes of Matrix Metalloproteinases. Prog. Mol. Biol. Transl. Sci. 2017, 147, 1–73. [Google Scholar] [PubMed] [Green Version]
- Cabral-Pacheco, G.A.; Garza-Veloz, I.; Castruita-De la Rosa, C.; Ramirez-Acuña, J.M.; Perez-Romero, B.A.; Guerrero-Rodriguez, J.F.; Martinez-Avila, N.; Martinez-Fierro, M.L. The Roles of Matrix Metalloproteinases and Their Inhibitors in Human Diseases. Int. J. Mol. Sci. 2020, 21, 9739. [Google Scholar] [CrossRef]
- Urnukhsaikhan, E.; Mishig-Ochir, T.; Kim, S.C.; Park, J.K.; Seo, Y.K. Neuroprotective Effect of Low Frequency-Pulsed Electromagnetic Fields in Ischemic Stroke. Appl. Biochem. Biotechnol. 2017, 181, 1360–1371. [Google Scholar] [CrossRef]
- Blais, V.; Rivest, S. Effects of TNF-alpha and IFN-gamma on nitric oxide-induced neurotoxicity in the mouse brain. J. Immunol. 2004, 172, 7043–7052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batsaikhan, B.; Wang, J.Y.; Scerba, M.T.; Tweedie, D.; Greig, N.H.; Miller, J.P.; Hoffer, B.J.; Lin, C.T.; Wang, J.Y. Post-Injury Neuroprotective Effects of the Thalidomide Analog 3,6′-Dithiothalidomide on Traumatic Brain Injury. Int. J. Mol. Sci. 2019, 20, 502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seifert, H.A.; Collier, L.A.; Chapman, C.B.; Benkovic, S.A.; Willing, A.E.; Pennypacker, K.R. Pro-inflammatory interferon gamma signaling is directly associated with stroke induced neurodegeneration. J. Neuroimmune Pharmacol. 2014, 9, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Watson, B.D.; Dietrich, W.D.; Busto, R.; Wachtel, M.S.; Ginsberg, M.D. Induction of reproducible brain infarction by photochemically initiated thrombosis. Ann. Neurol. 1985, 17, 497–504. [Google Scholar] [CrossRef]
- Liguz-Lecznar, M.; Ziemka-Nalecz, M.; Aleksy, M.; Kossut, M.; Skangiel-Kramska, J.; Nowicka, D. Comparison of matrix metalloproteinase activation after focal cortical ischemia in young adult and aged mice. J. Neurosci. Res. 2012, 90, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.K.; Urnukhsaikhan, E.; Yoon, H.H.; Seo, Y.K.; Park, J.K. Effect of human mesenchymal stem cell transplantation on cerebral ischemic volume-controlled photothrombotic mouse model. Biotechnol. J. 2016, 11, 1397–1404. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Days | ||||||
|---|---|---|---|---|---|---|---|
| 9 | 10 | 11 | 12 | 13 | 14 | 15 | |
| CTL | 3.0 ± 0.94 | 3.4 ± 1.27 | 7.6 ± 5.19 | 8.2 ± 3.49 | 8.6 ± 4.74 | 9.6 ± 4.91 | 9.4 ± 4.70 |
| Cell | 11.6 ± 4.79 | 12 ± 6.30 | 16.8 ± 8.31 | 23.6 ± 13.19 | 24.3 ± 13.57 | 30.3 ± 16.45 | 36.0 ± 19.46 |
| Cell/PEMF | 31.5 ± 34.51 | 42.6 ± 44.56 | 47.0 ± 46.80 | 53.5 ± 50.16 | 60.3 ± 45.65 | 63.3 ± 41.25 | 64.0 ± 42.93 |
| CTL vs. Cell CTL vs. Cell/PEMF Cell vs. Cell/PEMF | p < 0.005 p < 0.05 | p < 0.0005 p < 0.05 p < 0.05 | p < 0.005 p < 0.05 | p < 0.005 p < 0.05 | p < 0.005 p < 0.005 p < 0.005 | p < 0.005 p < 0.005 p < 0.05 | p < 0.005 p < 0.005 |
| Gene | Primer Sequences | Length of Amplicon (bp) |
|---|---|---|
| GAPDH | 5′-ACCACAGTCCATGCCATCAC-3′ 5′-TCCACCACCCTGTTGCTGTA-3′ | 452 |
| DCX | 5′-GGAAGGGGAAAGCTATGTCTG-3′ 5′-TTGCTGCTAGCCAAGGACTG-3′ | 138 |
| β-catenin | 5′-GACACCTCCCAAGTCCTTTAT-3′ 5′-GTACAACGGGCTGTTTCTACG-3′ | 470 |
| MAP2 | 5′-CTCAACAGTTCTATCTCTTCTTCA-3′ 5′-CTTCTTGTTTAAATCCTAACCT-3′ | 401 |
| MBP | 5′-GAGGAAGTGAATGAGCCGGTTA-3′ 5′-TTAGCTGAATTCGCGTGTGG-3′ | 379 |
| NeuroD1 | 5′-TGAGACGCATGAAGGCTAAC-3′ 5′-GAAATGGTGAAACTGGCGTG-3′ | 793 |
| NF-L | 5′-CAAGAACATGCAGAACGCTG-3′ 5′-GCCTTCCAAGAGTTTCCTGT-3′ | 376 |
| Tau | 5′-AAAGGTGGCAGTGGTTCG-3′ 5′-GGCTGGTGCTTCAGGTTC-3′ | 138 |
| Wnt3α | 5′-TGTTGGGCCACAGTATTCCT-3′ 5′-ATGAGCGTGTCACTGCAAAG-3′ | 302 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, H.-J.; Choi, J.-H.; Nam, M.-H.; Seo, Y.-K. Induced Neurodifferentiation of hBM-MSCs through Activation of the ERK/CREB Pathway via Pulsed Electromagnetic Fields and Physical Stimulation Promotes Neurogenesis in Cerebral Ischemic Models. Int. J. Mol. Sci. 2022, 23, 1177. https://doi.org/10.3390/ijms23031177
Park H-J, Choi J-H, Nam M-H, Seo Y-K. Induced Neurodifferentiation of hBM-MSCs through Activation of the ERK/CREB Pathway via Pulsed Electromagnetic Fields and Physical Stimulation Promotes Neurogenesis in Cerebral Ischemic Models. International Journal of Molecular Sciences. 2022; 23(3):1177. https://doi.org/10.3390/ijms23031177
Chicago/Turabian StylePark, Hee-Jung, Ju-Hye Choi, Myeong-Hyun Nam, and Young-Kwon Seo. 2022. "Induced Neurodifferentiation of hBM-MSCs through Activation of the ERK/CREB Pathway via Pulsed Electromagnetic Fields and Physical Stimulation Promotes Neurogenesis in Cerebral Ischemic Models" International Journal of Molecular Sciences 23, no. 3: 1177. https://doi.org/10.3390/ijms23031177
APA StylePark, H.-J., Choi, J.-H., Nam, M.-H., & Seo, Y.-K. (2022). Induced Neurodifferentiation of hBM-MSCs through Activation of the ERK/CREB Pathway via Pulsed Electromagnetic Fields and Physical Stimulation Promotes Neurogenesis in Cerebral Ischemic Models. International Journal of Molecular Sciences, 23(3), 1177. https://doi.org/10.3390/ijms23031177