
EN1 Regulates Cell Growth and Proliferation in Human Glioma Cells via Hedgehog Signaling

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
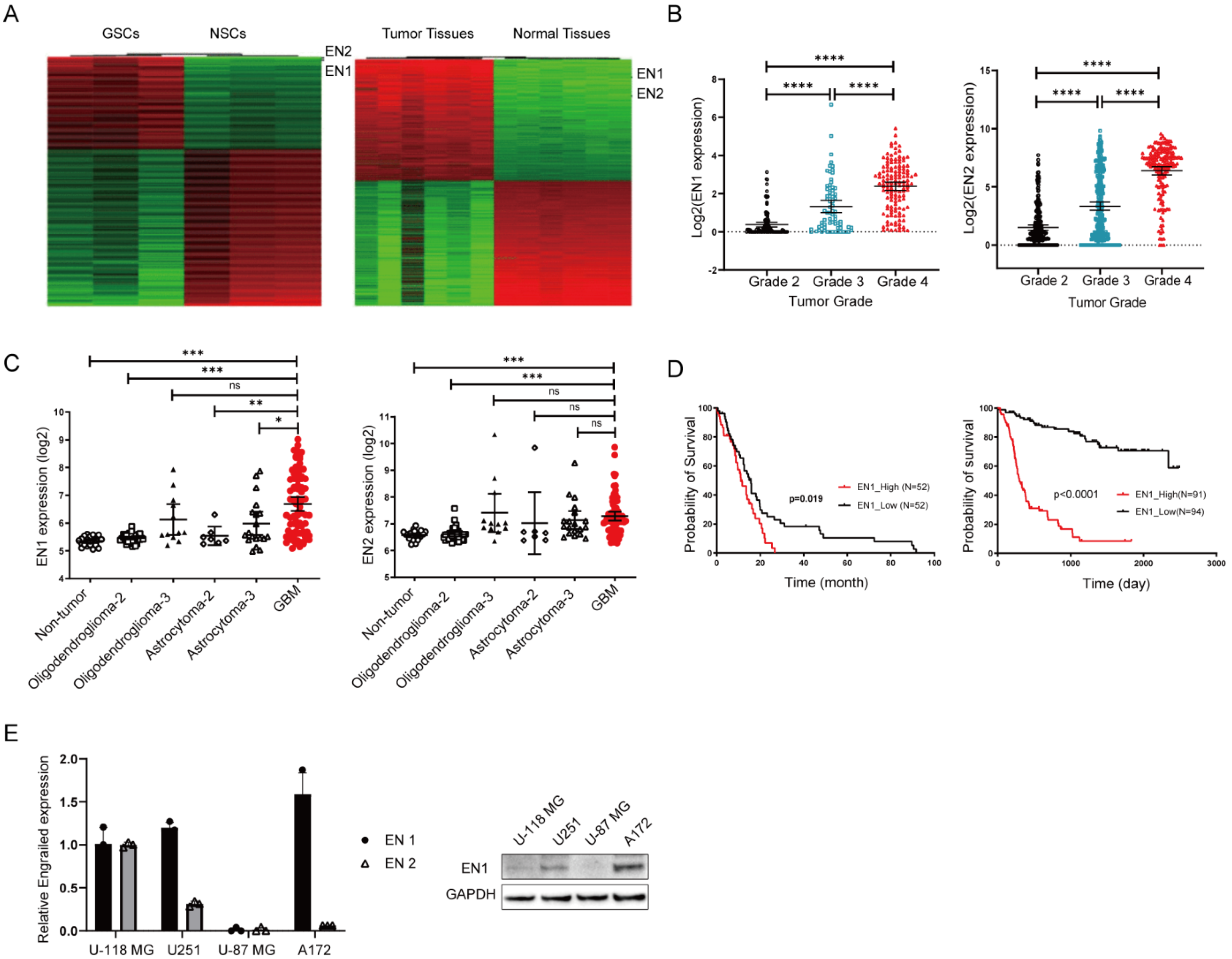
2.1. EN1 Is Upregulated in Glioblastoma and Is Related to Poor Patient Prognosis
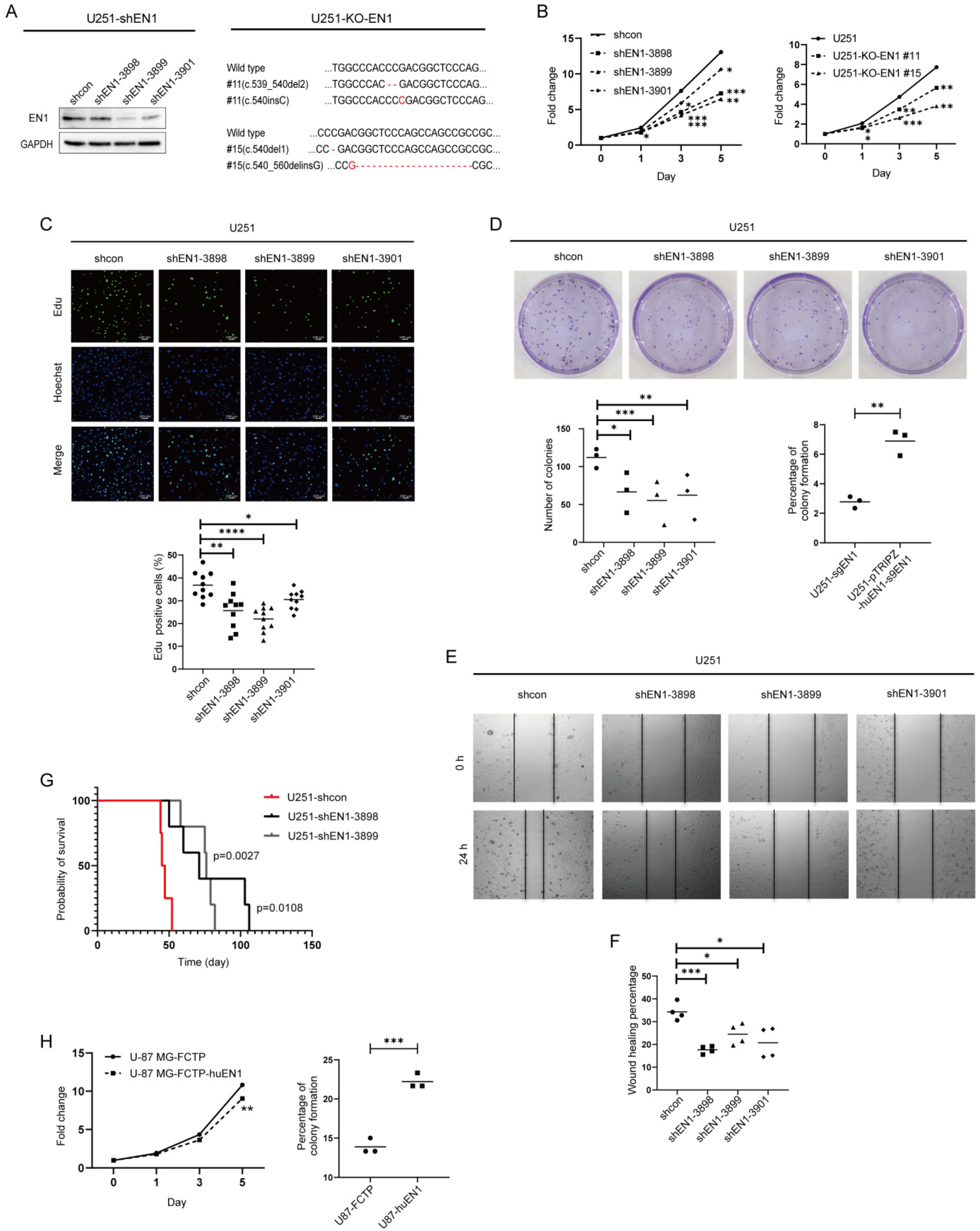
2.2. EN1 Knockdown Restrains the Proliferation and Migration of Glioblastoma Cells In Vitro and In Vivo
2.3. EN1 Knockdown Elevated Intracellular ROS Levels and Susceptibility to γ-ray Irradiation
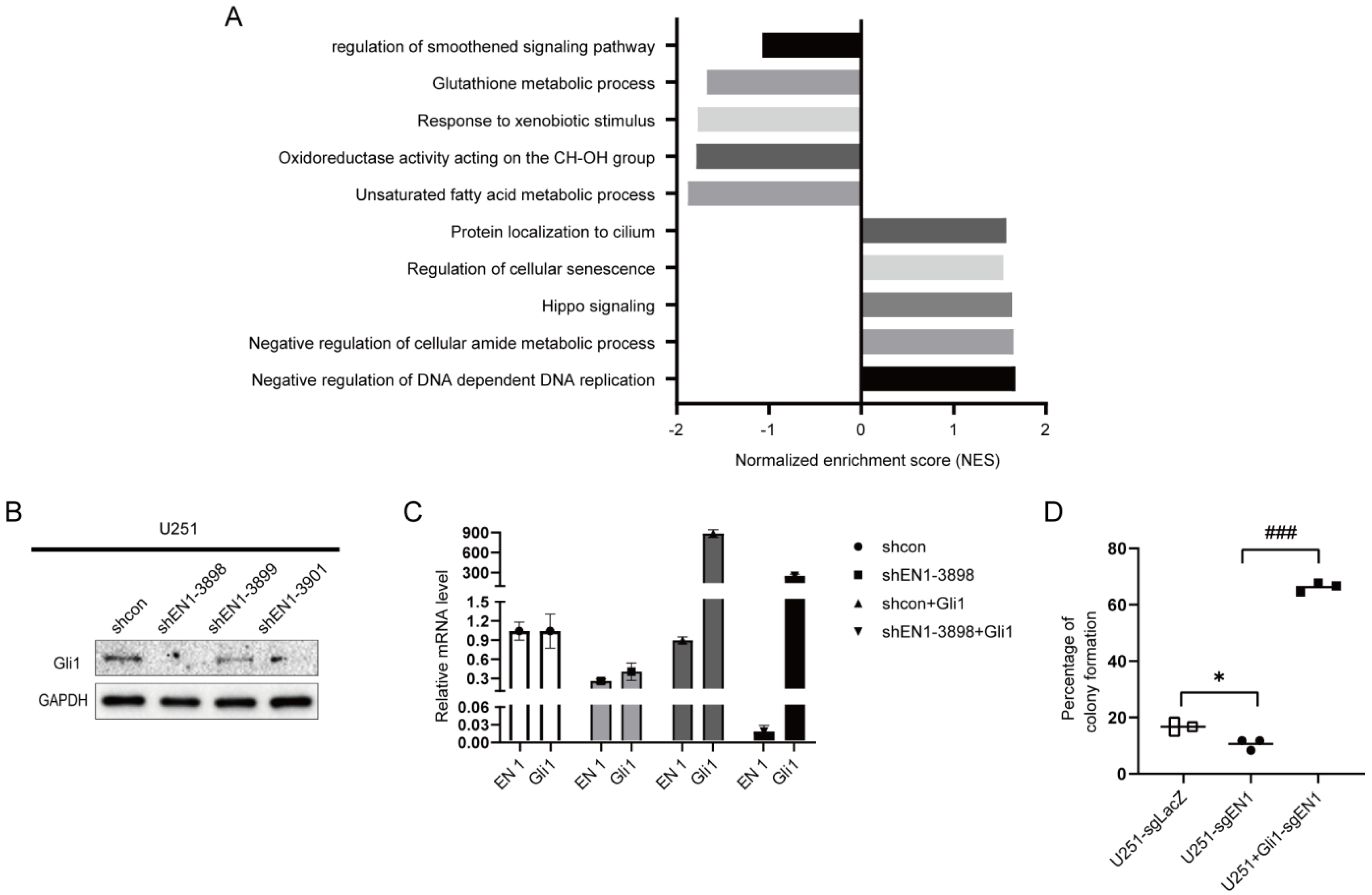
2.4. EN1 Increases Hedgehog Signaling Activity in Glioma Cells
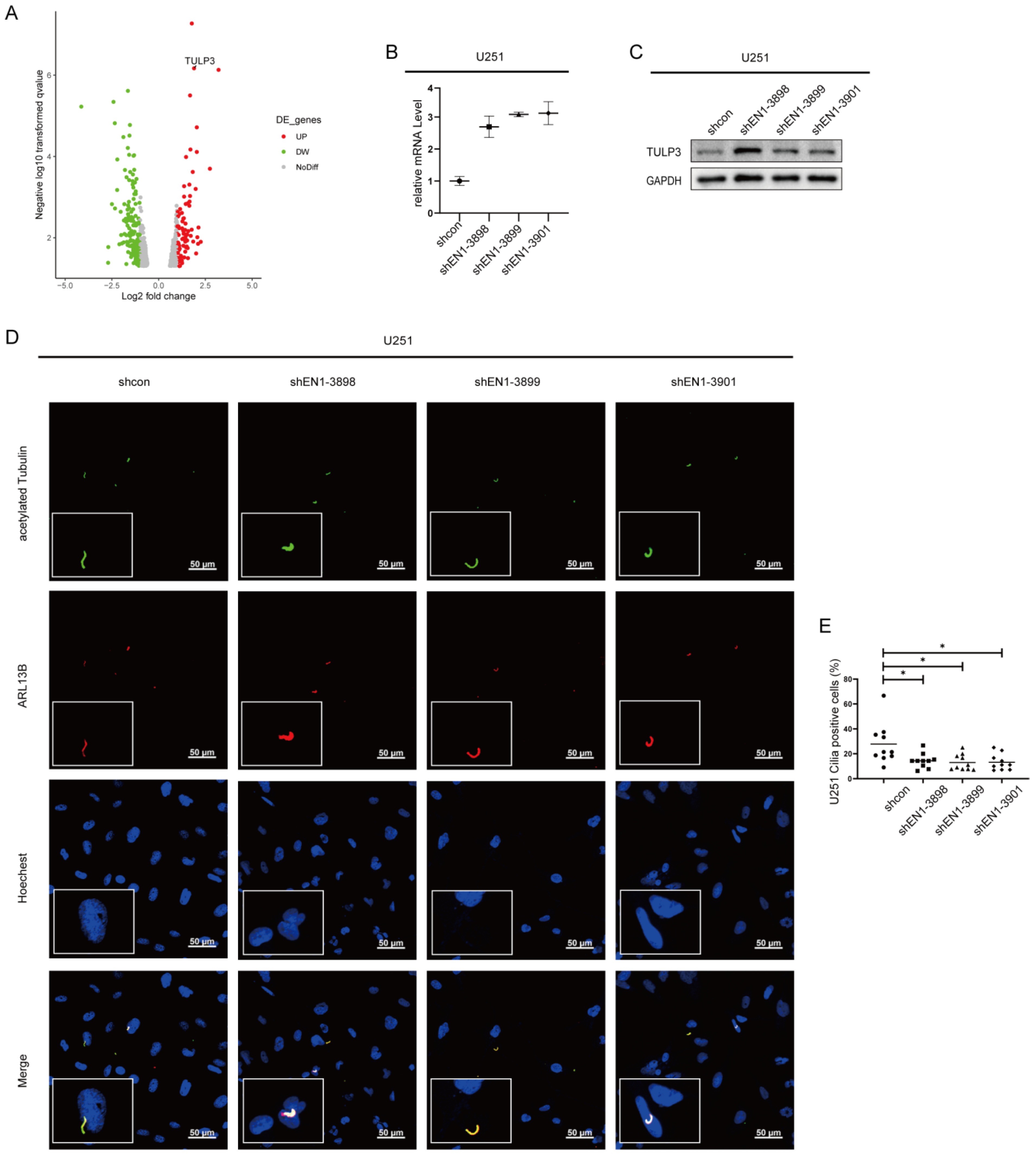
2.5. EN1 Knockdown Upregulates the Expression of TULP3
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Culture
4.2. Plasmids and Lentiviral Transduction
4.3. Quantitative PCR (qPCR)
4.4. Immunoblotting
4.5. Cell Proliferation Assays
4.6. Wound-Healing Assay
4.7. Colony Formation Assays
4.8. ROS Level
4.9. Immunofluorescence
4.10. RNA-Seq
4.11. Bioinformatics Analysis
4.12. Animal Studies
4.13. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Medikonda, R.; Dunn, G.; Rahman, M.; Fecci, P.; Lim, M. A review of glioblastoma immunotherapy. J. Neuro-Oncol. 2021, 151, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Hanif, F.; Muzaffar, K.; Perveen, K.; Malhi, S.M.; Simjee, S.U. Glioblastoma Multiforme: A Review of its Epidemiology and Pathogenesis through Clinical Presentation and Treatment. Asian Pac. J. Cancer Prev. 2017, 18, 3–9. [Google Scholar] [PubMed]
- Van Meir, E.G.; Hadjipanayis, C.G.; Norden, A.D.; Shu, H.K.; Wen, P.Y.; Olson, J.J. Exciting New Advances in Neuro-Oncology: The Avenue to a Cure for Malignant Glioma. CA A Cancer J. Clin. 2010, 60, 166–193. [Google Scholar] [CrossRef] [PubMed]
- Davis, M. Glioblastoma: Overview of Disease and Treatment. Clin. J. Oncol. Nurs. 2016, 20, S2–S8. [Google Scholar] [CrossRef] [Green Version]
- Osuka, S.; Van Meir, E.G. Overcoming therapeutic resistance in glioblastoma: The way forward. J. Clin. Investig. 2017, 127, 415–426. [Google Scholar] [CrossRef] [Green Version]
- García-Bellido, A.; Santamaría, P. Developmental analysis of the wing disc in the mutant engrailed of Drosophila melanogaster. Genetics 1972, 72, 87–104. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, M.T.A.; Smidt, M.P. En1 and Wnt signaling in midbrain dopaminergic neuronal development. Neural Dev. 2011, 6, 23. [Google Scholar] [CrossRef] [Green Version]
- Orvis, G.D.; Hartzell, A.L.; Smith, J.B.; Barraza, L.H.; Wilson, S.L.; Szulc, K.U.; Turnbull, D.H.; Joyner, A.L. The engrailed homeobox genes are required in multiple cell lineages to coordinate sequential formation of fissures and growth of the cerebellum. Dev. Biol. 2012, 367, 25–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willett, R.T.; Bayin, N.S.; Lee, A.S.; Krishnamurthy, A.; Wojcinski, A.; Lao, Z.; Stephen, D.; Rosello-Diez, A.; Dauber-Decker, K.L.; Orvis, G.D.; et al. Cerebellar nuclei excitatory neurons regulate developmental scaling of presynaptic Purkinje cell number and organ growth. eLife 2019, 8, e50617. [Google Scholar] [CrossRef]
- Hanks, M.; Wurst, W.; Anson-Cartwright, L.; Auerbach, A.B.; Joyner, A.L. Rescue of the En-1 mutant phenotype by replacement of En-1 with En-2. Science 1995, 269, 679–682. [Google Scholar] [CrossRef]
- Kim, Y.J.; Sung, M.; Oh, E.; Vrancken, M.V.; Song, J.Y.; Jung, K.; Choi, Y.L. Engrailed 1 overexpression as a potential prognostic marker in quintuple-negative breast cancer. Cancer Biol. Ther. 2018, 19, 335–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beltran, A.S.; Graves, L.M.; Blancafort, P. Novel role of Engrailed 1 as a prosurvival transcription factor in basal-like breast cancer and engineering of interference peptides block its oncogenic function. Oncogene 2013, 33, 4767–4777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Do Carmo Silva, J.; Vesely, S.; Novak, V.; Luksanova, H.; Prusa, R.; Babjuk, M. Engrailed 2 in urine fails as a marker in prostate cancer detection. Eur. Urol. Suppl. 2017, 16, e2646. [Google Scholar] [CrossRef]
- Punia, N.; Primon, M.; Simpson, G.R.; Pandha, H.S.; Morgan, R. Membrane insertion and secretion of the Engrailed-2 (EN2) transcription factor by prostate cancer cells may induce antiviral activity in the stroma. Sci. Rep. 2019, 9, 5138. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, J.; Xiao, Q.; Tian, R.; Zhou, Z.; Gan, Y.; Li, Y.; Shu, G.; Yin, G. EN2 as an oncogene promotes tumor progression via regulating CCL20 in colorectal cancer. Cell Death Dis. 2020, 11, 604. [Google Scholar] [CrossRef] [PubMed]
- Michael, A.; McGrath, S.; Annels, N.; Denyer, M.; Morgan, R.; Pandha, H. 438 Engrailed 2 protein (EN2) as a novel biomarker in epithelial ovarian cancer. Eur. J. Cancer 2015, 51, S91. [Google Scholar] [CrossRef]
- McGrath, S.E.; Annels, N.; Madhuri, T.K.; Tailor, A.; Butler-Manuel, S.A.; Morgan, R.; Pandha, H.; Michael, A. Engrailed-2 (EN2)—A novel biomarker in epithelial ovarian cancer. BMC Cancer 2018, 18, 943. [Google Scholar] [CrossRef] [Green Version]
- Martin, N.L.; Saba-El-Leil, M.K.; Sadekova, S.; Meloche, S.; Sauvageau, G. EN2 is a candidate oncogene in human breast cancer. Oncogene 2005, 24, 6890–6901. [Google Scholar] [CrossRef] [Green Version]
- Skoda, A.M.; Simovic, D.; Karin, V.; Kardum, V.; Vranic, S.; Serman, L. The role of the Hedgehog signaling pathway in cancer: A comprehensive review. Bosn. J. Basic Med. Sci. 2018, 18, 8–20. [Google Scholar] [CrossRef]
- Chen, M.H.; Wilson, C.W.; Li, Y.J.; Law, K.K.; Lu, C.S.; Gacayan, R.; Zhang, X.; Hui, C.C.; Chuang, P.T. Cilium-independent regulation of Gli protein function by Sufu in Hedgehog signaling is evolutionarily conserved. Genes Dev. 2009, 23, 1910–1928. [Google Scholar] [CrossRef] [Green Version]
- Jeng, K.S.; Chang, C.F.; Lin, S.S. Sonic Hedgehog Signaling in Organogenesis, Tumors, and Tumor Microenvironments. Int. J. Mol. Sci. 2020, 21, 758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, M.; Ventura, P.B.; Jiang, Y.; Rodriguez, F.J.; Wang, L.; Perry, J.S.A.; Yang, Y.; Wahl, K.; Crittenden, R.B.; Bennett, M.L.; et al. Astrocytic trans-Differentiation Completes a Multicellular Paracrine Feedback Loop Required for Medulloblastoma Tumor Growth. Cell 2020, 180, 502–520.e19. [Google Scholar] [CrossRef] [PubMed]
- Santoni, M.; Burattini, L.; Nabissi, M.; Beatrice Morelli, M.; Berardi, R.; Santoni, G.; Cascinu, S. Essential Role of Gli Proteins in Glioblastoma Multiforme. Curr. Protein Pept. Sci. 2013, 14, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Alcantara Llaguno, S.; Chen, J.; Kwon, C.H.; Jackson, E.L.; Li, Y.; Burns, D.K.; Alvarez-Buylla, A.; Parada, L.F. Malignant astrocytomas originate from neural stem/progenitor cells in a somatic tumor suppressor mouse model. Cancer Cell 2009, 15, 45–56. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Li, Y.; Yu, T.S.; McKay, R.M.; Burns, D.K.; Kernie, S.G.; Parada, L.F. A restricted cell population propagates glioblastoma growth after chemotherapy. Nature 2012, 488, 522–526. [Google Scholar] [CrossRef] [Green Version]
- Auffinger, B.; Spencer, D.; Pytel, P.; Ahmed, A.U.; Lesniak, M.S. The role of glioma stem cells in chemotherapy resistance and glioblastoma multiforme recurrence. Expert Rev. Neurother. 2015, 15, 741–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gusev, Y.; Bhuvaneshwar, K.; Song, L.; Zenklusen, J.C.; Fine, H.; Madhavan, S. The REMBRANDT study, a large collection of genomic data from brain cancer patients. Sci. Data 2018, 5, 180158. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Zhang, K.N.; Wang, Q.; Li, G.; Zeng, F.; Zhang, Y.; Wu, F.; Chai, R.; Wang, Z.; Zhang, C.; et al. Chinese Glioma Genome Atlas (CGGA): A Comprehensive Resource with Functional Genomic Data from Chinese Glioma Patients. Genom. Proteom. Bioinform. 2021, 19, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. ROS Are Good. Trends. Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindell Jonsson, E.; Erngren, I.; Engskog, M.; Haglof, J.; Arvidsson, T.; Hedeland, M.; Petterson, C.; Laurell, G.; Nestor, M. Exploring Radiation Response in Two Head and Neck Squamous Carcinoma Cell Lines Through Metabolic Profiling. Front. Oncol. 2019, 9, 825. [Google Scholar] [CrossRef] [Green Version]
- Azzam, E.I.; Jay-Gerin, J.P.; Pain, D. Ionizing radiation-induced metabolic oxidative stress and prolonged cell injury. Cancer Lett. 2012, 327, 48–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamori, T.; Yasui, H.; Yamazumi, M.; Wada, Y.; Nakamura, Y.; Nakamura, H.; Inanami, O. Ionizing radiation induces mitochondrial reactive oxygen species production accompanied by upregulation of mitochondrial electron transport chain function and mitochondrial content under control of the cell cycle checkpoint. Free Radic. Biol. Med. 2012, 53, 260–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singer, E.; Judkins, J.; Salomonis, N.; Matlaf, L.; Soteropoulos, P.; McAllister, S.; Soroceanu, L. Reactive oxygen species-mediated therapeutic response and resistance in glioblastoma. Cell Death Dis. 2015, 6, e1601–e1611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, S.; Takebe, N.; Lorusso, P. Targeting the Hedgehog pathway in cancer. Ther. Adv. Med. Oncol. 2010, 2, 237–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carpenter, R.L.; Lo, H.W. Hedgehog pathway and GLI1 isoforms in human cancer. Discov. Med. 2012, 13, 105. [Google Scholar] [PubMed]
- Badgandi, H.B.; Hwang, S.H.; Shimada, I.S.; Loriot, E.; Mukhopadhyay, S. Tubby family proteins are adapters for ciliary trafficking of integral membrane proteins. J. Cell Biol. 2017, 216, 743–760. [Google Scholar] [CrossRef] [Green Version]
- Bangs, F.; Anderson, K.V. Primary cilia and mammalian hedgehog signaling. Cold Spring Harb. Perspect. Biol. 2017, 9, a028175. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.; Wen, X.; Ratti, N.; Loktev, A.; Rangell, L.; Scales, S.J.; Jackson, P.K. The ciliary G-protein-coupled receptor Gpr161 negatively regulates the Sonic hedgehog pathway via cAMP signaling. Cell 2013, 152, 210–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norman, R.X.; Ko, H.W.; Huang, V.; Eun, C.M.; Abler, L.L.; Zhang, Z.; Sun, X.; Eggenschwiler, J.T. Tubby-like protein 3 (TULP3) regulates patterning in the mouse embryo through inhibition of Hedgehog signaling. Hum. Mol. Genet. 2009, 18, 1740–1754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, D.; Bell, A.; Roberts, D.; Weber, R.S.; El-Naggar, A.K. Developmental transcription factor EN1- novel biomarker in human salivary gland adenoid cystic carcinoma. Cancer 2012, 118, 1288–1292. [Google Scholar] [CrossRef] [Green Version]
- Bachar-Dahan, L.; Goltzmann, J.; Yaniv, A.; Gazit, A. Engrailed-1 negatively regulates beta-catenin transcriptional activity by destabilizing beta-catenin via a glycogen synthase kinase-3beta-independent pathway. Mol. Biol. Cell 2006, 17, 2572–2580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clement, V.; Sanchez, P.; de Tribolet, N.; Radovanovic, I.; Ruiz i Altaba, A. HEDGEHOG-GLI1 Signaling Regulates Human Glioma Growth, Cancer Stem Cell Self-Renewal, and Tumorigenicity. Curr. Biol. 2007, 17, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Lee, S.Y.; Oh, S.C. Hedgehog signaling pathway as a potential target in the treatment of advanced gastric cancer. Tumor Biol. 2017, 39, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilovocky, N.A.; Romito-DiGiacomo, R.R.; Murcia, C.L.; Maricich, S.M.; Herrup, K. Factors in the genetic background suppress the engrailed-1 cerebellar phenotype. J. Neurosci. 2003, 23, 5105–5112. [Google Scholar] [CrossRef]
- Tabata, T.; Eaton, S.; Kornberg, T.B. The Drosophila hedgehog gene is expressed specifically in posterior compartment cells and is a target of engrailed regulation. Genes Dev. 1992, 6, 2635–2645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jonkman, J.E.; Cathcart, J.A.; Xu, F.; Bartolini, M.E.; Amon, J.E.; Stevens, K.M.; Colarusso, P. An introduction to the wound healing assay using live-cell microscopy. Cell Adh. Migr. 2014, 8, 440–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franken, N.A.; Rodermond, H.M.; Stap, J.; Haveman, J.; van Bree, C. Clonogenic assay of cells in vitro. Nat. Protoc. 2006, 1, 2315–2319. [Google Scholar] [CrossRef]
- Darmanis, S.; Sloan, S.A.; Croote, D.; Mignardi, M.; Chernikova, S.; Samghababi, P.; Zhang, Y.; Neff, N.; Kowarsky, M.; Caneda, C. Single-cell RNA-seq analysis of infiltrating neoplastic cells at the migrating front of human glioblastoma. Cell Rep. 2017, 21, 1399–1410. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, J.; Guo, C.; Li, J.; Liang, Z.; Wang, Y.; Yu, A.; Liu, R.; Guo, Y.; Chen, J.; Huang, S. EN1 Regulates Cell Growth and Proliferation in Human Glioma Cells via Hedgehog Signaling. Int. J. Mol. Sci. 2022, 23, 1123. https://doi.org/10.3390/ijms23031123
Chang J, Guo C, Li J, Liang Z, Wang Y, Yu A, Liu R, Guo Y, Chen J, Huang S. EN1 Regulates Cell Growth and Proliferation in Human Glioma Cells via Hedgehog Signaling. International Journal of Molecular Sciences. 2022; 23(3):1123. https://doi.org/10.3390/ijms23031123
Chicago/Turabian StyleChang, Jinchun, Chenjia Guo, Jianyu Li, Zhangqian Liang, Yankai Wang, Anliang Yu, Runze Liu, Yuting Guo, Jian Chen, and Song Huang. 2022. "EN1 Regulates Cell Growth and Proliferation in Human Glioma Cells via Hedgehog Signaling" International Journal of Molecular Sciences 23, no. 3: 1123. https://doi.org/10.3390/ijms23031123
APA StyleChang, J., Guo, C., Li, J., Liang, Z., Wang, Y., Yu, A., Liu, R., Guo, Y., Chen, J., & Huang, S. (2022). EN1 Regulates Cell Growth and Proliferation in Human Glioma Cells via Hedgehog Signaling. International Journal of Molecular Sciences, 23(3), 1123. https://doi.org/10.3390/ijms23031123