

Reduced Elastin Fibers and Melanocyte Loss in Vitiliginous Skin Are Restored after Repigmentation by Phototherapy and/or Autologous Minigraft Transplantation

Abstract
1. Introduction
2. Results
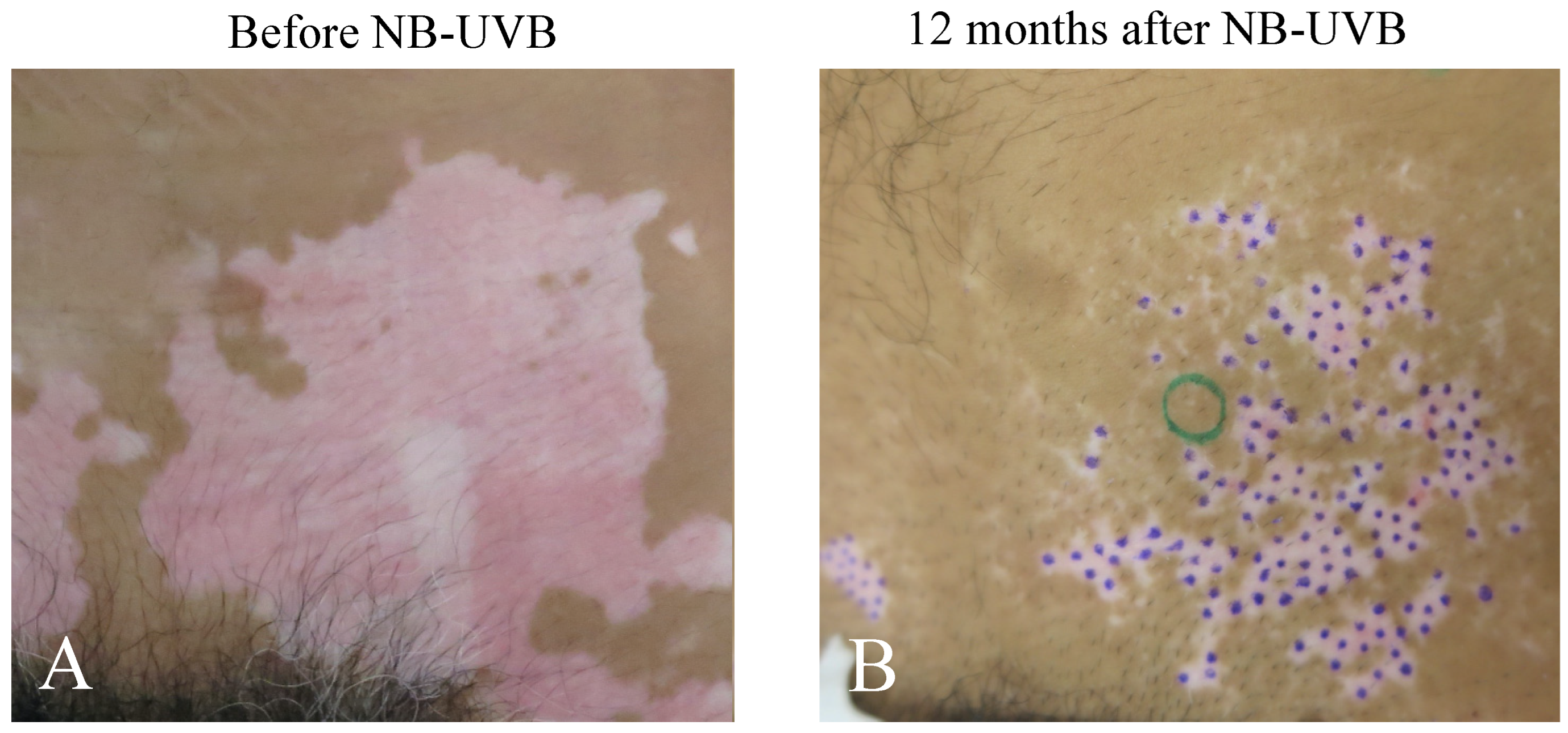
2.1. Repigmentation of Lesional Skin after Phototherapy and/or mMG
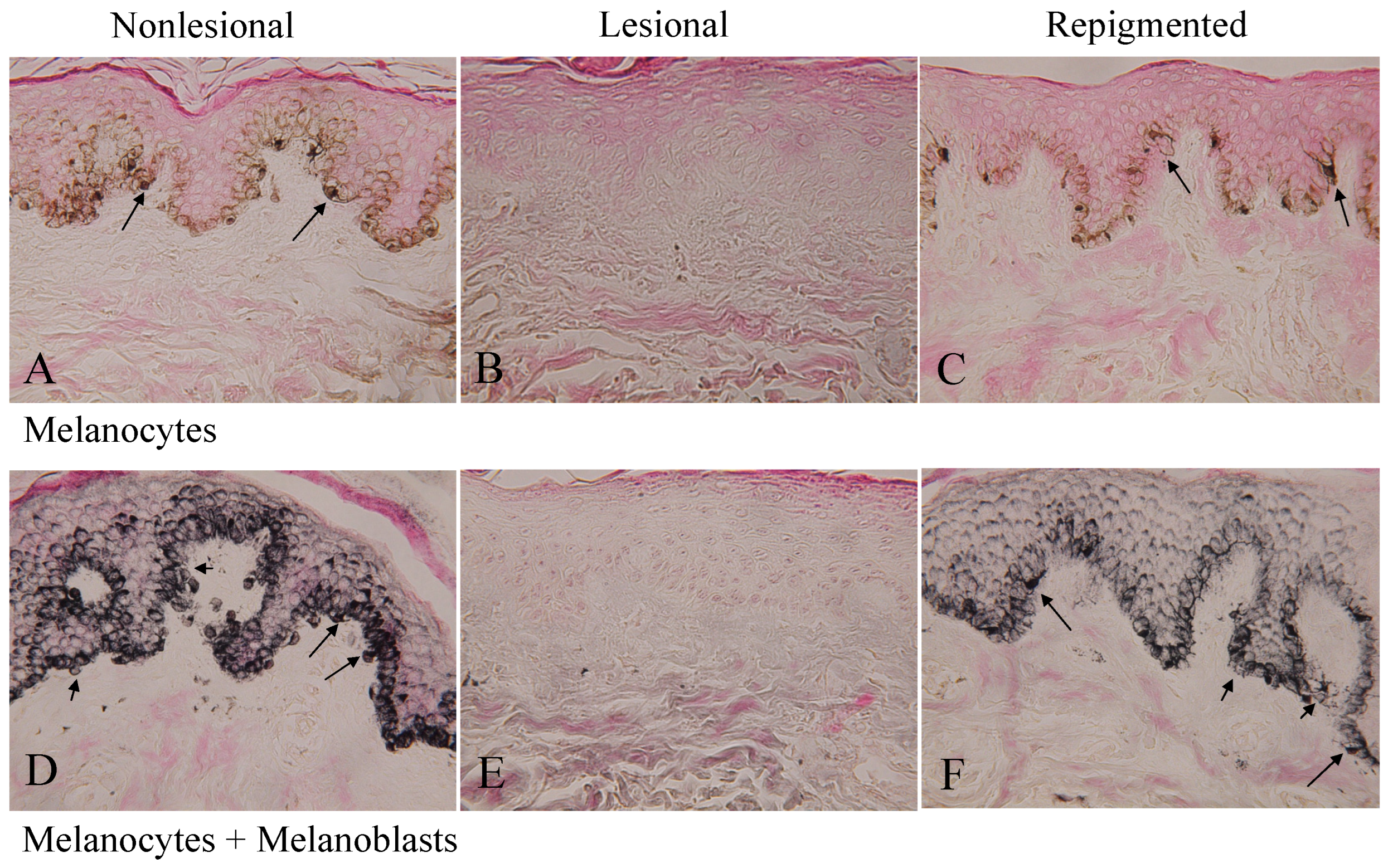
2.2. Melanocytes and Melanoblasts in Nonlesional, Lesional, and Repigmented Skin
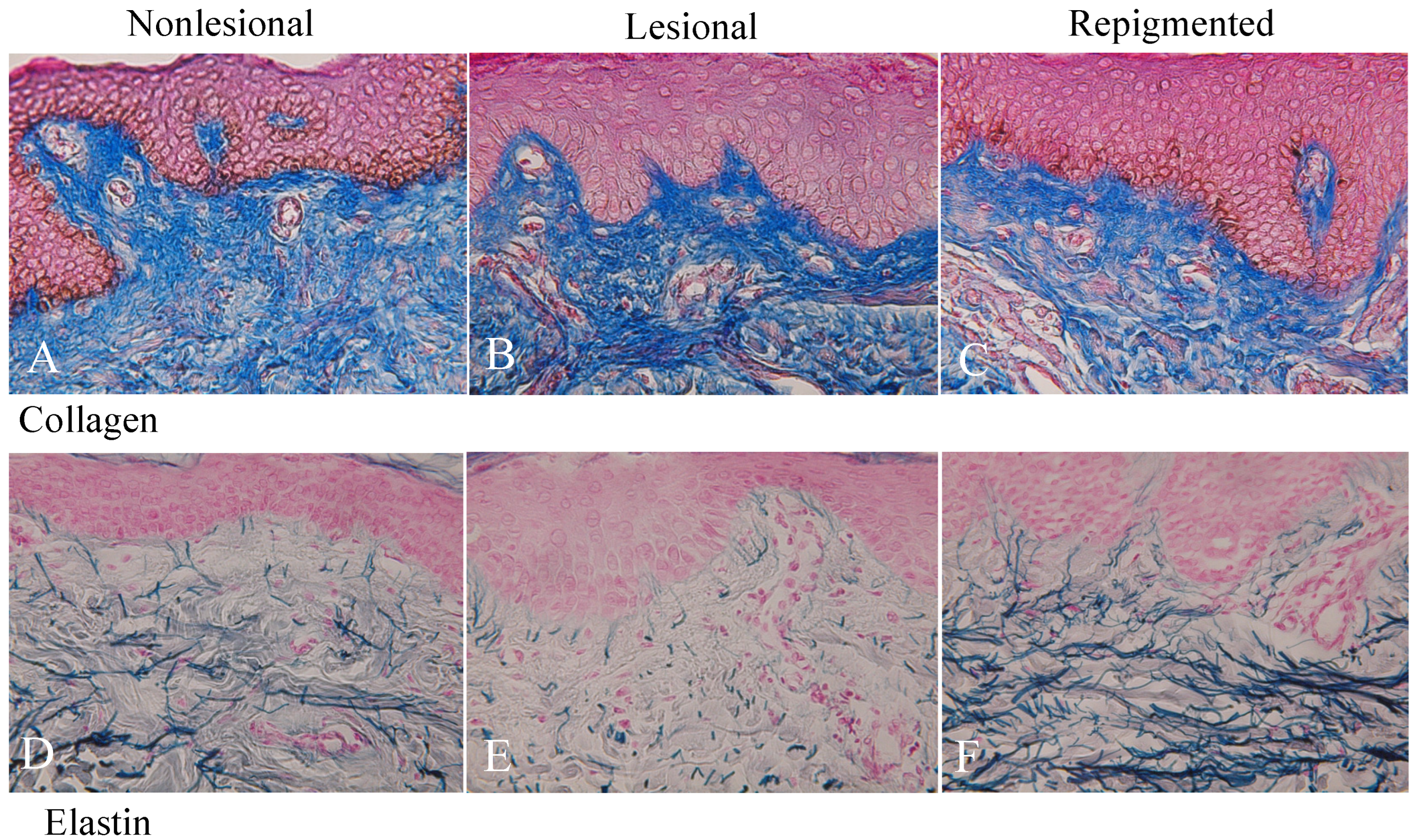
2.3. Collagen and Elastin Fibers in Nonlesional, Lesional, and Repigmented Skin
2.4. Differences between Nonlesional and Repigmented Skin in the Number of Epidermal Melanocytes and Melanoblasts
2.5. Differences between Single and Double Phototherapies in the Recovery of Epidermal Melanocytes/Melanocytes and Elastin Fibers in the Repigmented Skin
2.6. Differences between Phototherapy Alone and Phototherapy plus mMG in the Recovery of Epidermal Melanocytes/Melanocytes and Elastin Fibers in the Repigmented Skin
2.7. Gender Differences in the Recovery of Epidermal Melanocytes/Melanocytes and Elastin Fibers in the Repigmented Skin after Phototherapy and/or Phototherapy plus mMG
2.8. Age Differences in the Recovery of Epidermal Melanocytes/Melanocytes and Elastin Fibers in the Repigmented Skin after Phototherapy and Phototherapy plus mMG
3. Discussion
4. Materials and Methods
4.1. Exposures of MEL and NB-UVB
4.2. Skin Transplantation (mMG)
4.3. Collection of Skin Samples
4.4. Histochemistry
4.5. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hirobe, T. How are proliferation and differentiation of melanocytes regulated? Pigment. Cell Melanoma Res. 2011, 24, 462–478. [Google Scholar] [CrossRef]
- Hearing, V.J. The melanosome: The perfect model for cellular response to the environment. Pigment Cell Res. 2000, 13 (Suppl. S8), 23–34. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; Wakamatsu, K. Chemistry of mixed melanogenesis-pivotal roles of dopaquinone. Photochem. Photobiol. 2008, 84, 582–592. [Google Scholar] [CrossRef]
- Riding, R.L.; Harris, J.E. The role of memory CD8+ T cells in vitiligo. J. Immunol. 2019, 203, 11–19. [Google Scholar] [CrossRef]
- Seneschal, J.; Boniface, K.; D’Arino, A.; Picardo, M. An update on vitiligo pathogenesis. Pigment. Cell Melanoma Res. 2021, 34, 236–243. [Google Scholar] [CrossRef]
- Richmond, J.M.; Bangari, D.S.; Essien, K.I.; Currimbhoy, S.D.; Groom, J.; Pandya, A.; Youd, M.E.; Luster, A.D.; Harris, J.E. Keratinocyte-derived chemokines orchestrate T-cell positioning in the epidermis during vitiligo and may serve as biomarkers of disease. J. Investig. Dermatol. 2017, 137, 350–358. [Google Scholar] [CrossRef]
- Biswas, K.B.; Takahashi, A.; Mizutani, Y.; Takayama, S.; Ishitsuka, A.; Yang, L.; Yang, F.; Iddamalgoda, A.; Katayama, I.; Inoue, S. GPNMB is expressed in human epidermal keratinocytes but disappears in the vitiligo lesional skin. Sci. Rep. 2020, 10, 4930. [Google Scholar] [CrossRef] [PubMed]
- Rani, S.; Bhardwaj, S.; Srivastava, N.; Sharma, V.L.; Parsad, D.; Kumar, R. Senescence in the lesional fibroblasts of non-segmental vitiligo patients. Arch. Dermatol. Res. 2017, 309, 123–132. [Google Scholar] [CrossRef]
- Theocharis, A.D.; Manou, D.; Karamanos, N.K. The extracellular matrix as a multitasking player in disease. FASEB J. 2019, 286, 2830–2869. [Google Scholar] [CrossRef] [PubMed]
- Rosenbloom, J.; Abrams, W.R.; Mecham, R. Extracellular matrix 4: The elastic fiber. FASEB J. 1993, 7, 1208–1218. [Google Scholar] [CrossRef]
- Romero-Ortuno, R.; Kenny, R.A.; McManus, R. Collagens and elastin genetic variations and their potential role in aging-related diseases and longevity in humans. Exp. Gerontol. 2020, 129, 110781. [Google Scholar] [CrossRef] [PubMed]
- Hirobe, T.; Enami, H.; Nakayama, A. Elastin fiber but not collagen fiber is decreased dramatically in the dermis of vitiligo patients. Int. J. Dermatol. 2020, 59, e369–e372. [Google Scholar] [CrossRef]
- Parsad, D.; Pandhi, R.; Dogra, S.; Kumar, B. Clinical study of repigmentation patterns with different treatment modalities and their correlation with speed and stability of repigmentation in 352 vitiliginous patches. J. Am. Acad. Dermatol. 2004, 50, 63–67. [Google Scholar] [CrossRef]
- Park, K.K.; Liao, W.; Murase, J.E. A review of monochromatic excimer light in vitiligo. Br. J. Dermatol. 2012, 167, 468–478. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Qiao, M.; Wang, X.; Zhao, X.; Sun, Q. Effect of narrow band ultraviolet B phototherapy as monotherapy or combination therapy for vitiligo: A meta-analysis. Photodermatol. Photoimmunol. Photomed. 2017, 33, 22–31. [Google Scholar] [CrossRef]
- Hirobe, T.; Enami, H. Excellent color-matched repigmentation of human vitiligo can be obtained by mini-punch grafting using a machine in combination with ultraviolet therapy. Dermatol. Sin. 2018, 36, 203–206. [Google Scholar] [CrossRef]
- Bae, J.M.; Lee, J.H.; Kwon, H.S.; Kim, J.; Kim, D.S. Motorized 0.8-mm micropunch grafting for refractory vitiligo: A retrospective study of 230 cases. J. Am. Acad. Dermatol. 2018, 79, 720–727. [Google Scholar] [CrossRef]
- Razmi, T.M.; Afra, T.P.; Parsad, D. Vitiligo surgery: A journey from tissues via cells to the stems! Exp. Dermatol. 2019, 28, 690–694. [Google Scholar] [CrossRef]
- Nishimura, E.K.; Jordan, S.A.; Oshima, H.; Yoshida, H.; Osawa, M.; Moriyama, M.; Jackson, I.J.; Barrandon, Y.; Miyachi, Y.; Nishikawa, S.-I. Dominant role of the niche in melanocyte stem-cell fate determination. Nature 2002, 416, 854–860. [Google Scholar] [CrossRef]
- Hirobe, T.; Enami, H. Histochemical study of the distribution of epidermal melanoblasts and melanocytes in Asian human skin. Skin Res. Technol. 2019, 25, 299–304. [Google Scholar] [CrossRef]
- Goldstein, N.; Koster, M.I.; Hoaglin, L.G.; Spoelstra, N.S.; Kechris, K.J.; Robinson, S.E.; Robinson, W.; Roop, D.R.; Norris, D.A.; Birlea, S.A. Narrow band ultraviolet B treatment for human vitiligo is associated with proliferation, migration, and differentiation of melanocyte precursors. J. Investig. Dermatol. 2015, 135, 2068–2076. [Google Scholar] [CrossRef] [PubMed]
- Grichnik, J.M.; Ali, W.N.; Burch, J.A.; Byers, J.D.; Garcia, C.A.; Clark, R.E.; Shea, C.R. KIT expression reveals a population of precursor melanocytes in human skin. J. Investig. Dermatol. 1996, 106, 967–971. [Google Scholar] [CrossRef] [PubMed]
- Michalak-Mićka, K.; Büchler, V.L.; Zapiórkowska-Blumer, N.; Biedermann, T.; Klar, A.S. Characterization of a melanocyte progenitor population in human interfollicular epidermis. Cell Rep. 2022, 38, 110419. [Google Scholar] [CrossRef]
- Hirobe, T.; Enami, H. Elastin peptides with ferrous ferric chloride activate human melanocytes and elastin fibers. J. Skin Stem Cell 2022, 9, e127254. [Google Scholar] [CrossRef]
- Chang, C.-H.; Kawa, Y.; Tsai, R.-K.; Shieh, J.-H.; Lee, J.-W.; Watabe, H.; Kawakami, T.; Soma, Y.; Tajima, S.; Mizoguchi, M. Melanocyte precursors express elastin binding protein and elastin-derived peptide (VGVAPG) stimulates their melanogenesis and dendrite formation. J. Dermatol. Sci. 2008, 51, 158–170. [Google Scholar] [CrossRef]
- Ono, T.; Mah, K.; Hu, F. Dermal melanocytes and elastic fibers. J. Cutan. Pathol. 1985, 12, 468–475. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Itami, S.; Watabe, H.; Yasumoto, K.I.; Abdel-Malek, Z.A.; Kubo, T.; Rouzaud, F.; Tanemura, A.; Yoshikawa, K.; Hearing, V.J. Mesenchymal–epithelial interactions in the skin: Increased expression of dickkopf1 by palmoplantar fibroblasts inhibits melanocyte growth and differentiation. J. Cell Biol. 2004, 165, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Hirobe, T.; Hasegawa, K.; Furuya, R.; Fujiwara, R.; Sato, K. Effects of fibroblast-derived factors on the proliferation and differentiation of human melanocytes in culture. J. Dermatol. Sci. 2013, 71, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Hirobe, T. Role of keratinocyte-derived factors involved in regulating the proliferation and differentiation of mammalian epidermal melanocytes. Pigment Cell Res. 2005, 18, 2–12. [Google Scholar] [CrossRef]
- Mildner, M.; Mlitz, V.; Gruber, F.; Wojta, J.; Tschachler, E. Hepatocyte growth factor establishes autocrine and paracrine feedback loops for the protection of skin cells after UV irradiation. J. Investig. Dermatol. 2007, 127, 2637–2644. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| No. | Gender (Age) | Skin | MEL/NB (No. of Sess) | MEL/NB Month | Dose J/cm2 | mMG Month | No. of M | No. of Mb | Collagen | Elastin | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Non | Les | Repig | Non | Les | Repig | Les | Repig | Les | Repig | |||||||
| 1. | M (15) | Belly | NB (50) | 12 | 38.3 | No | 110.9 | 0 | 105.8 | 104.3 | 0 | 106.3 | → | → | ↓↓↓ | ↑↑↑ |
| 2. | M (31) | Chin | NB (70) | 10 | 51.3 | No | 134.8 | 0 | 120.4 | 117.4 | 0 | 82.9 | → | → | ↓↓ | ↑↑↑ |
| 3. | F (11) | Neck | MEL (8) + NB (8) | 4 | 4.1 | 4 | 127.5 | 0 | 177.9 | 103.8 | 0 | 103.9 | ↓ | ↑ | ↓↓↓ | ↑↑↑ |
| 4. | M (71) | Neck | MEL (1) + NB (6) | 1 | 10.1 | 1 | 53.7 | 0 | 82.3 | 95.0 | 0 | 94.0 | → | → | ↓↓↓ | ↑↑↑ |
| 5. | F (45) | Neck | MEL (5) + NB (6) | 0.7 | 4.5 | 0.7 | 87.5 | 0 | 91.6 | 90.0 | 0 | 71.1 | → | → | ↓↓↓ | ↑↑↑ |
| 6. | M (15) | Neck | MEL (3) + NB (1) | 2 | 1.1 | 2 | 111.3 | 0 | 167.8 | 98.9 | 0 | 150.4 | → | → | ↓↓ | ↑↑↑ |
| 7. | M (6) | Eyelid | MEL (27) + NB (3) | 5 | 8.0 | 5 | 107.8 | 0 | 108.3 | 99.9 | 0 | 126.9 | → | → | ↓ | ↑↑ |
| 8. | M (10) | Head | MEL (35) + NB (31) | 10 | 50.1 | 10 | 125.2 | 0 | 299.5 | 100.1 | 0 | 283.5 | → | → | ↓↓ | ↑↑ |
| 9. | F (14) | Eyelid | MEL (4) | 1 | 0.3 | No | 130.5 | 0 | 85.7 | 102.7 | 0 | 96.4 | → | → | ↓ | ↑↑ |
| 10. | F (13) | Face | MEL (12) + NB (13) | 3 | 7.6 | No | 93.4 | 0 | 126.7 | 101.2 | 0 | 128.6 | → | → | ↓ | ↑↑ |
| 11. | F (41) | Eyelid | MEL (2) + NB (38) | 7 | 25.5 | 7 | 109.9 | 0 | 174.1 | 113.8 | 0 | 108.1 | → | → | ↓↓↓ | ↑↑↑ |
| 12. | F (71) | Arm | NB (51) | 24 | 52.4 | No | 98.5 | 0 | 107.6 | 103.5 | 0 | 97.9 | ↓ | ↑ | ↓↓↓ | ↑↑↑ |
| 13. | F (77) | Hand | NB (151) | 12 | 64.1 | No | 150.5 | 0 | 138.6 | 107.3 | 0 | 99.5 | → | → | ↓↓↓ | ↑↑↑ |
| 14. | M (65) | Cheek | MEL (12) + NB (12) | 2 | 8.2 | 2 | 120.8 | 0 | 88.4 | 106.5 | 0 | 89.9 | → | → | ↓↓↓ | ↑↑↑ |
| Ave | 34.6 ± 6.8 | 39.2 ± 10.0 | 6.7 ± 1.7 | 23.3 ± 5.9 | 111.6 ± 6.1 | 133.9 ± 14.9 | 103.2 ± 1.8 | 117.1 ± 13.4 | ||||||||
| No. of Melanocytes/0.1 mm2 | No. of Melanoblasts/0.1 mm2 | |||
|---|---|---|---|---|
| (A) | Single MEL/NB vs Double MEL/NB | |||
| Single | Double | Single | Double | |
| (n = 5) | (n = 9) | (n = 5) | (n = 9) | |
| 111.6 ± 7.8 | 146.3 ± 21.7 | 96.6 ± 3.4 | 128.5 ± 19.7 | |
| (B) | MEL/NB alone vsMEL/NB + mMG | |||
| MEL/NB | MEL/NB + mMG | MEL/NB | MEL/NB + mMG | |
| (n = 6) | (n = 8) | (n = 6) | (n = 8) | |
| 114.1 ± 6.9 | 148.7 ± 24.2 | 101.9 ± 5.6 | 128.5 ± 22.2 | |
| (C) | Gender | |||
| Female | Male | Female | Male | |
| (n = 7) | (n = 7) | (n = 7) | (n = 7) | |
| 128.9 ± 13.0 | 138.9 ± 26.6 | 100.8 ± 6.0 | 133.4 ± 24.6 | |
| (D) | Age | |||
| Young | Old | Young | Old | |
| (n = 7) | (n = 7) | (n = 7) | (n = 7) | |
| 153.1 ± 25.5 | 114.7 ± 11.5 | 142.3 ± 22.7 * | 91.9 ± 4.2 * | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hirobe, T.; Enami, H. Reduced Elastin Fibers and Melanocyte Loss in Vitiliginous Skin Are Restored after Repigmentation by Phototherapy and/or Autologous Minigraft Transplantation. Int. J. Mol. Sci. 2022, 23, 15361. https://doi.org/10.3390/ijms232315361
Hirobe T, Enami H. Reduced Elastin Fibers and Melanocyte Loss in Vitiliginous Skin Are Restored after Repigmentation by Phototherapy and/or Autologous Minigraft Transplantation. International Journal of Molecular Sciences. 2022; 23(23):15361. https://doi.org/10.3390/ijms232315361
Chicago/Turabian StyleHirobe, Tomohisa, and Hisao Enami. 2022. "Reduced Elastin Fibers and Melanocyte Loss in Vitiliginous Skin Are Restored after Repigmentation by Phototherapy and/or Autologous Minigraft Transplantation" International Journal of Molecular Sciences 23, no. 23: 15361. https://doi.org/10.3390/ijms232315361
APA StyleHirobe, T., & Enami, H. (2022). Reduced Elastin Fibers and Melanocyte Loss in Vitiliginous Skin Are Restored after Repigmentation by Phototherapy and/or Autologous Minigraft Transplantation. International Journal of Molecular Sciences, 23(23), 15361. https://doi.org/10.3390/ijms232315361