
BDNF as a Mediator of Antidepressant Response: Recent Advances and Lifestyle Interactions


Abstract
1. The Paradigm Shift in Antidepressant Mechanism of Action
2. BDNF as a Critical Antidepressant Mediator
2.1. BDNF Exerts Distinct Effects in MDD-Linked Brain Regions
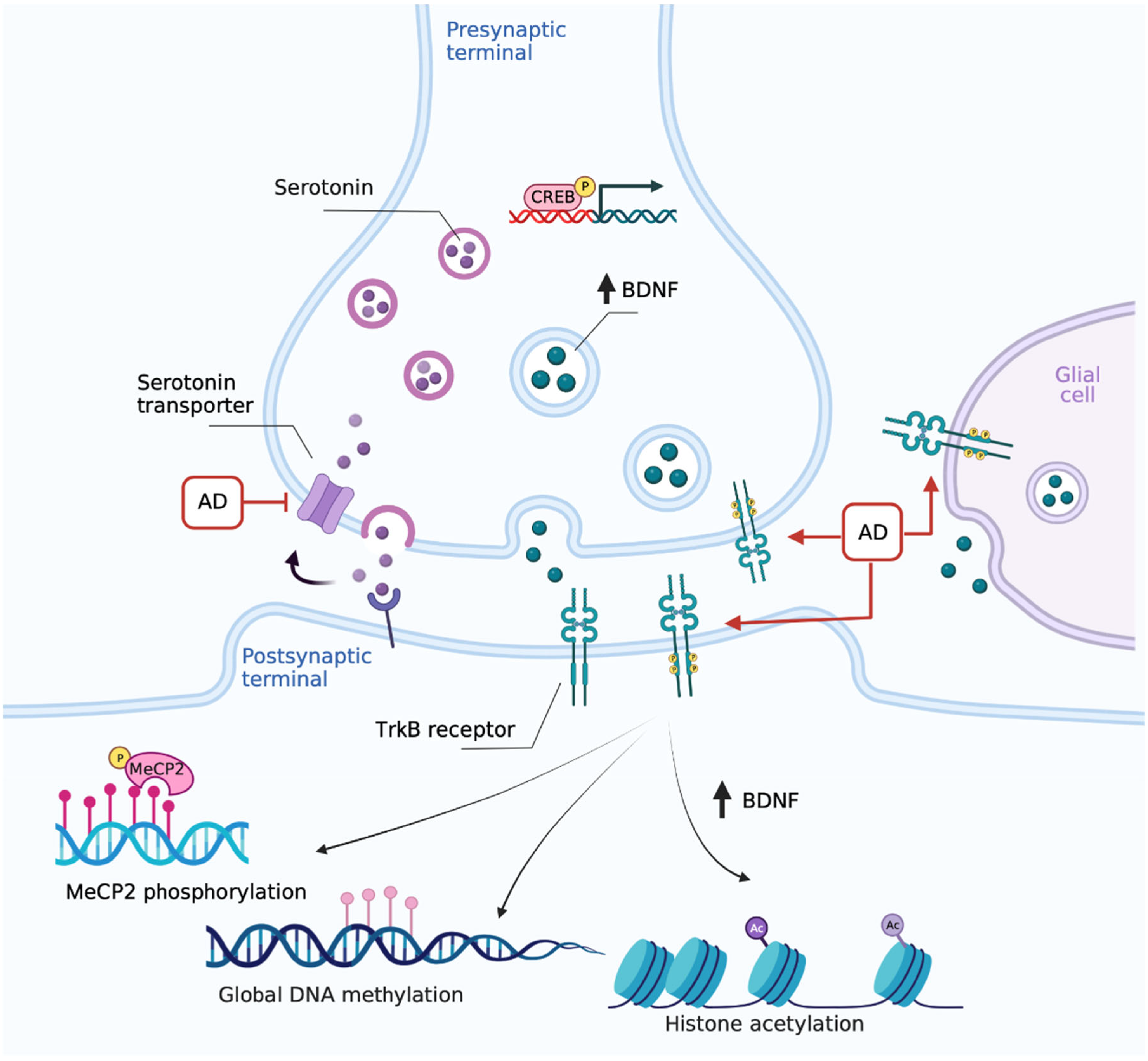
2.2. Multiple Cell Types Contribute to Increased BDNF Following Antidepressant Treatment
2.3. Epigenetic Regulation of the Bdnf Gene
3. Lifestyle Interactions with Antidepressants in Rodent Models and Humans
3.1. Nutrition
3.2. Environment
3.3. Aging

{kind=link}
| Adjunctive Nutrient | Antidepressant | Outcome | Reference |
|---|---|---|---|
| Omega-3 PUFA 1000 mg/d | Sertraline | MADRS: Omega-3 PUFA < placebo | [81] |
| Ethyl-EPA 1 g/d | Fluoxetine 20 mg/d | HDRS: Ethyl-EPA < placebo | [79] |
| EPA 1, 2 or 4 g/d | Unspecified antidepressants | HDRS EPA < placebo (best 1 g/d) | [82] |
| EPA 2 g/d | Unspecified antidepressants | HDRS EPA < placebo | [84] |
| DHA 260 mg/d or 520 mg/d | Unspecified antidepressants | HDRS < after treatment resistance | [87] |
| EPA 1.8 g/d + DHA 0.4 g/d | Citalopram 20–40 mg/d | HDRS: Fatty acids < placebo | [88] |
| EPA 0.93 g/d + DHA 0.75 g/d | Sertraline 50 mg/d | BDI-II or HDRS: Fatty acids = placebo | [80] |
| EPA 1 g/d, DHA 1 g/d | Unspecified antidepressants | HDRS: EPA < DHA or placebo | [86] |
| EPA + DHA 3 g/d (EPA 0.6 g; DHA 2.4 g) | Unspecified antidepressants | HDRS-SF: EPA + DHA = placebo | [85] |
| EPA + DHA 9.6 g/d | Unspecified antidepressants | HDRS EPA + DHA < placebo | [83] |
| Palmitoylethanolamide 1200 mg/d | Citalopram 20 mg/d | HDRS: Palmitoylethanolamide < placebo | [90] |
| Creatine 4 mg/d | Fluoxetine 20–40 mg/d | CDRS-R: Creatine < placebo | [104] |
| Creatine 5 g/d | Escitalopram 20 mg/d | HDRS: creatine < placebo | [103] |
| Tryptophan 4 g/d | Fluoxetine 20 mg/d | HDRS: Tryptophan < placebo | [110] |
| DL-Tryptophan 0.1 g/kg body weight | Chlorimipramine 150 mg/d | Cronholm-Ottosson Depression Scale: n.s. | [111] |
| DL-Tryptophan 12, 15, 18 g depending on body weight | Phenelzine 60 mg/d | HDRS: DL-Tryptophan < placebo | [112] |
| SAMe 800 mg/d | Unspecified antidepressants | HDRS: < after treatment resistance | [206,207] |
| SAMe 1600 mg/d | SSRIs | HDRS: SAMe < placebo (in treatment resistance) | [116] |
| SAMe 200 mg/d, NAC 200 mg/d and folate 200 ug/d | SSRIs | HADS-A and CGI: better with SAMe, NAC and folate | [114] |
| L-Methylfolate 15 mg/d | SSRIs | QIDS-SR and CGI: better with L-Methylfolate in SSRI resistance | [129] |
| L-Methylfolate | SSRIs/SNRIs | CGI: L-Methylfolate < placebo | [130] |
| Folic acid 0.5 mg/d | Fluoxetine 20 mg/d | HDRS: Folic acid < placebo | [126] |
| Folic acid 10 mg/d | Fluoxetine 20 mg/d | HDRS: Folic acid < placebo | [127] |
| Folic acid 5 mg/d | Unspecified antidepressants | BDI-II: Folic acid = placebo | [131] |
| Folic acid 2 mg/d + Vitamin B12 0.5 mg/d + Vitamin B6 25 mg/d | Citalopram 20–40 mg/d | MADRS: Folic acid + B12 + B6 = placebo | [135] |
| Folic acid 400 ug/d + Vitamin B12 100 ug/d | Unspecified antidepressants | PHQ-9: n.s. | [134] |
| Vitamin B12 1 mg/d | SSRIs 20–40 mg/d | HDRS: with Vitamin B12 < placebo | [132] |
| TCA 100–200 mg/d | HDRS: with Vitamin B12 < placebo | [132] | |
| Vitamin C 1 g/d | Fluoxetine 10–20 mg/d | CDRS: with Vitamin C < placebo | [136] |
| Citalopram 20 mg/d | HDRS: with Vitamin C = placebo | [138] | |
| Vitamin D3 1500IU/d | Fluoxetine 20 mg/d | HDRS: with Vitamin D3 < placebo | [137] |
| Vitamin D3 300.000 U | Unspecified antidepressants | HDRS with Vitamin D3 < placebo (after 4 weeks) | [139] |
| Zinc 25 mg/d | Fluoxetine 20–60 mg/d | BDI: Zinc < placebo | [148] |
| Citalopram 20–60 mg/d | BDI: Zinc < placebo | [148] | |
| Imipramine 100–200 mg/d | BDI, HDRS, CGI, MADRS: Zinc < placebo | [150,151] | |
| Unspecified antidepressants | HDRS, BDI: Zinc < placebo | [208] | |
| Magnesium 120 mg/d | Fluoxetine 20–40 mg/d | HDRS: Magnesium = placebo | [154] |
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Malhi, G.S.; Mann, J.J. Depression. Lancet 2018, 392, 2299–2312. [Google Scholar] [CrossRef]
- Nestler, E.J.; Barrot, M.; DiLeone, R.J.; Eisch, A.J.; Gold, S.J.; Monteggia, L.M. Neurobiology of depression. Neuron 2002, 34, 13–25. [Google Scholar] [CrossRef]
- Moncrieff, J.; Moncrieff, J.; Cooper, R.E.; Stockmann, T.; Amendola, S.; Hengartner, M.P.; Horowitz, M.A. The serotonin theory of depression: A systematic umbrella review of the evidence. Mol. Psychiatry 2022, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Nibuya, M.; Morinobu, S.; Duman, R.S. Regulation of BDNF and trkB mRNA in rat brain by chronic electroconvulsive seizure and antidepressant drug treatments. J. Neurosci. 1995, 15, 7539–7547. [Google Scholar] [CrossRef] [PubMed]
- Duman, R.S.; Heninger, G.R.; Nestler, E.J. A molecular and cellular theory of depression. Arch. Gen. Psychiatry 1997, 54, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Bjorkholm, C.; Monteggia, L.M. BDNF—A key transducer of antidepressant effects. Neuropharmacology 2016, 102, 72–79. [Google Scholar] [CrossRef]
- Ilchibaeva, T.; Tsybko, A.; Zeug, A.; Müller, F.E.; Guseva, D.; Bischoff, S.; Ponimaskin, E.; Naumenko, V. Serotonin Receptor 5-HT2A Regulates TrkB Receptor Function in Heteroreceptor Complexes. Cells 2022, 11, 2384. [Google Scholar] [CrossRef]
- Castrén, E.; Antila, H. Neuronal plasticity and neurotrophic factors in drug responses. Mol. Psychiatry 2017, 22, 1085–1095. [Google Scholar]
- Kishimoto, T.; Chawla, J.M.; Hagi, K.; Zarate, C.A.; Kane, J.M.; Bauer, M.; Correll, C.U. Single-dose infusion ketamine and non-ketamine N-methyl-d-aspartate receptor antagonists for unipolar and bipolar depression: A meta-analysis of efficacy, safety and time trajectories. Psychol. Med. 2016, 46, 1459–1472. [Google Scholar] [CrossRef]
- Jelen, L.A.; Young, A.H.; Stone, J.M. Ketamine: A tale of two enantiomers. J. Psychopharmacol. 2021, 35, 109–123. [Google Scholar] [CrossRef]
- Zanos, P.; Moaddel, R.; Morris, P.J.; Georgiou, P.; Fischell, J.; Elmer, G.I.; Alkondon, M.; Yuan, P.; Pribut, H.J.; Singh, N.S.; et al. NMDAR inhibition-independent antidepressant actions of ketamine metabolites. Nature 2016, 533, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Zanos, P.; Gould, T.D. Mechanisms of ketamine action as an antidepressant. Mol. Psychiatry 2018, 23, 801–811. [Google Scholar] [CrossRef] [PubMed]
- Soppet, D.; Escandon, E.; Maragos, J.; Middlemas, D.S.; Reid, S.W.; Blair, J.; Burton, L.E.; Stanton, B.R.; Kaplan, D.R.; Hunter, T.; et al. The neurotrophic factors brain-derived neurotrophic factor and neurotrophin-3 are ligands for the trkB tyrosine kinase receptor. Cell 1991, 65, 895–903. [Google Scholar] [CrossRef]
- Yoshii, A.; Constantine-Paton, M. Postsynaptic BDNF-TrkB signaling in synapse maturation, plasticity, and disease. Dev. Neurobiol. 2010, 70, 304–322. [Google Scholar] [CrossRef] [PubMed]
- Esvald, E.E.; Tuvikene, J.; Sirp, A.; Patil, S.; Bramham, C.R.; Timmusk, T. CREB Family Transcription Factors Are Major Mediators of BDNF Transcriptional Autoregulation in Cortical Neurons. J. Neurosci. 2020, 40, 1405–1426. [Google Scholar]
- Duman, R.S.; Monteggia, L.M. A Neurotrophic Model for Stress-Related Mood Disorders. Biol. Psychiatry 2006, 59, 1116–1127. [Google Scholar] [CrossRef]
- Saarelainen, T.; Hendolin, P.; Lucas, G.; Koponen, E.; Sairanen, M.; MacDonald, E.; Agerman, K.; Haapasalo, A.; Nawa, H.; Aloyz, R.; et al. Activation of the TrkB neurotrophin receptor is induced by antidepressant drugs and is required for antidepressant-induced behavioral effects. J. Neurosci. 2003, 23, 349–357. [Google Scholar] [CrossRef]
- Rantamäki, T.; Hendolin, P.; Kankaanpää, A.; Mijatovic, J.; Piepponen, P.; Domenici, E.; Chao, M.V.; Männistö, P.T.; Castrén, E. Pharmacologically diverse antidepressants rapidly activate brain-derived neurotrophic factor receptor TrkB and induce phospholipase-Cgamma signaling pathways in mouse brain. Neuropsychopharmacology 2007, 32, 2152–2162. [Google Scholar]
- Rantamäki, T.; Vesa, L.; Antila, H.; Di Lieto, A.; Tammela, P.; Schmitt, A.; Lesch, K.P.; Rios, M.; Castrén, E. Antidepressant drugs transactivate TrkB neurotrophin receptors in the adult rodent brain independently of BDNF and monoamine transporter blockade. PLoS ONE 2011, 6, e20567. [Google Scholar] [CrossRef]
- Park, H.; Poo, M.M. Neurotrophin regulation of neural circuit development and function. Nat. Rev. Neurosci. 2013, 14, 7–23. [Google Scholar] [CrossRef]
- Maya Vetencourt, J.F.; Sale, A.; Viegi, A.; Baroncelli, L.; De Pasquale, R.; O’Leary, O.F.; Castrén, E.; Maffei, L. The antidepressant fluoxetine restores plasticity in the adult visual cortex. Science 2008, 320, 385–388. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Luikart, B.W.; Birnbaum, S.; Chen, J.; Kwon, C.H.; Kernie, S.G.; Bassel-Duby, R.; Parada, L.F. TrkB Regulates Hippocampal Neurogenesis and Governs Sensitivity to Antidepressive Treatment. Neuron 2008, 59, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Zang, T.; Birnbaum, S.G.; Wang, Z.; Johnson, J.E.; Zhang, C.L.; Parada, L.F. TrkB dependent adult hippocampal progenitor differentiation mediates sustained ketamine antidepressant response. Nat. Commun. 2017, 8, 1668. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, Y.; Rizavi, H.S.; Conley, R.R.; Roberts, R.C.; Tamminga, C.A.; Pandey, G.N. Altered gene expression of brain-derived neurotrophic factor and receptor tyrosine kinase B in postmortem brain of suicide subjects. Arch. Gen. Psychiatry 2003, 60, 804–815. [Google Scholar] [CrossRef]
- Pandey, G.N.; Ren, X.; Rizavi, H.S.; Conley, R.R.; Roberts, R.C.; Dwivedi, Y. Brain-derived neurotrophic factor and tyrosine kinase B receptor signalling in post-mortem brain of teenage suicide victims. Int. J. Neuropsychopharmacol. 2008, 11, 1047–1061. [Google Scholar] [CrossRef]
- Dwivedi, Y.; Rizavi, H.S.; Zhang, H.; Mondal, A.C.; Roberts, R.C.; Conley, R.R.; Pandey, G.N. Neurotrophin receptor activation and expression in human postmortem brain: Effect of suicide. Biol. Psychiatry 2009, 65, 319–328. [Google Scholar] [CrossRef]
- Banerjee, R.; Ghosh, A.K.; Ghosh, B.; Bhattacharyya, S.; Mondal, A.C. Decreased mRNA and Protein Expression of BDNF, NGF, and their Receptors in the Hippocampus from Suicide: An Analysis in Human Postmortem Brain. Clin. Med. Insights Pathol. 2013, 6, 1–11. [Google Scholar] [CrossRef]
- Jang, S.W.; Liu, X.; Chan, C.B.; Weinshenker, D.; Hall, R.A.; Xiao, G.; Ye, K. Amitriptyline is a TrkA and TrkB receptor agonist that promotes TrkA/TrkB heterodimerization and has potent neurotrophic activity. Chem. Biol. 2009, 16, 644–656. [Google Scholar] [CrossRef]
- Casarotto, P.C.; Girych, M.; Fred, S.M.; Kovaleva, V.; Moliner, R.; Enkavi, G.; Biojone, C.; Cannarozzo, C.; Sahu, M.P.; Kaurinkoski, K.; et al. Antidepressant drugs act by directly binding to TRKB neurotrophin receptors. Cell 2021, 184, 1299–1313.e19. [Google Scholar] [CrossRef]
- Faron-Górecka, A.; Kuśmider, M.; Gruca, P.; Pabian, P.; Korlatowicz, A.; Solich, J.; Kolasa, M.; Dziedzicka-Wasylewska, M. Pro-cognitive effect of acute imipramine administration correlates with direct interaction of BDNF with its receptor, Trkβ. Brain Res. 2022, 1789, 147948. [Google Scholar] [CrossRef]
- McAllister, A.K.; Katz, L.C.; Lo, D.C. Neurotrophins and synaptic plasticity. Annu. Rev. Neurosci. 1999, 22, 295–318. [Google Scholar] [CrossRef] [PubMed]
- Mamounas, L.A.; Altar, C.A.; Blue, M.E.; Kaplan, D.R.; Tessarollo, L.; Lyons, W.E. BDNF promotes the regenerative sprouting, but not survival, of injured serotonergic axons in the adult rat brain. J. Neurosci. 2000, 20, 771–782. [Google Scholar] [CrossRef] [PubMed]
- Nibuya, M.; Nestler, E.J.; Duman, R.S. Chronic antidepressant administration increases the expression of cAMP response element binding protein (CREB) in rat hippocampus. J. Neurosci. 1996, 16, 2365–2372. [Google Scholar] [CrossRef]
- Monteggia, L.M.; Barrot, M.; Powell, C.M.; Berton, O.; Galanis, V.; Gemelli, T.; Meuth, S.; Nagy, A.; Greene, R.W.; Nestler, E.J. Essential role of brain-derived neurotrophic factor in adult hippocampal function. Proc. Natl. Acad. Sci. USA 2004, 101, 10827–10832. [Google Scholar] [CrossRef]
- Siuciak, J.A.; Lewis, D.R.; Wiegand, S.J.; Lindsay, R.M. Antidepressant-like effect of brain-derived neurotrophic factor (BDNF). Pharmacol. Biochem. Behav. 1997, 56, 131–137. [Google Scholar] [CrossRef]
- Shirayama, Y.; Chen, A.C.H.; Nakagawa, S.; Russell, D.S.; Duman, R.S. Brain-derived neurotrophic factor produces antidepressant effects in behavioral models of depression. J. Neurosci. 2002, 22, 3251–3261. [Google Scholar] [CrossRef]
- Adachi, M.; Barrot, M.; Autry, A.E.; Theobald, D.; Monteggia, L.M. Selective loss of brain-derived neurotrophic factor in the dentate gyrus attenuates antidepressant efficacy. Biol. Psychiatry 2008, 63, 642–649. [Google Scholar] [CrossRef] [PubMed]
- Aloyz, R.; Fawcett, J.; Kaplan, D.; Murphy, R.; Miller, F. Activity-dependent activation of TrkB neurotrophin receptors in the adult CNS. Learn. Mem. 1999, 6, 216–231. [Google Scholar] [CrossRef] [PubMed]
- Balkowiec, A.; Katz, D.M. Activity-dependent release of endogenous brain-derived neurotrophic factor from primary sensory neurons detected by ELISA in situ. J. Neurosci. 2000, 20, 7417–7423. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Jurič, D.M.; Lončar, D.; Čarman-Kržan, M. Noradrenergic stimulation of BDNF synthesis in astrocytes: Mediation via alpha1- and beta1/beta2-adrenergic receptors. Neurochem. Int. 2008, 52, 297–306. [Google Scholar] [CrossRef]
- Galter, D.; Unsicker, K. Sequential activation of the 5-HT1(A) serotonin receptor and TrkB induces the serotonergic neuronal phenotype. Mol. Cell. Neurosci. 2000, 15, 446–455. [Google Scholar] [CrossRef] [PubMed]
- Poon, C.H.; Heng, B.C.; Lim, L.W. New insights on brain-derived neurotrophic factor epigenetics: From depression to memory extinction. Ann. N. Y. Acad. Sci. 2021, 1484, 9–31. [Google Scholar] [CrossRef] [PubMed]
- Deyama, S.; Kaneda, K. The duration of the antidepressant-like effects of a single infusion of brain-derived neurotrophic factor into the medial prefrontal cortex in mice. Behav. Brain Res. 2020, 394, 112844. [Google Scholar] [CrossRef] [PubMed]
- Koo, J.W.; Labonté, B.; Engmann, O.; Calipari, E.S.; Juarez, B.; Lorsch, Z.; Walsh, J.J.; Friedman, A.K.; Yorgason, J.T.; Han, M.H.; et al. Essential Role of Mesolimbic Brain-Derived Neurotrophic Factor in Chronic Social Stress-Induced Depressive Behaviors. Biol. Psychiatry 2015, 80, 469–478. [Google Scholar] [CrossRef]
- Koo, J.W.; Chaudhury, D.; Han, M.H.; Nestler, E.J. Role of Mesolimbic Brain-Derived Neurotrophic Factor in Depression. Biol. Psychiatry 2019, 86, 738–748. [Google Scholar] [CrossRef]
- Molteni, R.; Calabrese, F.; Bedogni, F.; Tongiorgi, E.; Fumagalli, F.; Racagni, G.; Riva, M.A. Chronic treatment with fluoxetine up-regulates cellular BDNF mRNA expression in rat dopaminergic regions. Int. J. Neuropsychopharmacol. 2006, 9, 307–317. [Google Scholar] [CrossRef]
- Nestler, E.J.; Carlezon, W.A., Jr. The mesolimbic dopamine reward circuit in depression. Biol. Psychiatry 2006, 59, 1151–1159. [Google Scholar] [CrossRef]
- Eisch, A.J.; Bolaños, C.A.; de Wit, J.; Simonak, R.D.; Pudiak, C.M.; Barrot, M.; Verhaagen, J.; Nestler, E.J. Brain-derived neurotrophic factor in the ventral midbrain-nucleus accumbens pathway: A role in depression. Biol. Psychiatry 2003, 54, 994–1005. [Google Scholar] [CrossRef]
- Furuse, K.; Ukai, W.; Hashimoto, E.; Hashiguchi, H.; Kigawa, Y.; Ishii, T.; Tayama, M.; Deriha, K.; Shiraishi, M.; Kawanishi, C. Antidepressant activities of escitalopram and blonanserin on prenatal and adolescent combined stress-induced depression model: Possible role of neurotrophic mechanism change in serum and nucleus accumbens. J. Affect. Disord. 2019, 247, 97–104. [Google Scholar] [CrossRef]
- Brancato, A.; Castelli, V.; Cavallaro, A.; Lavanco, G.; Plescia, F.; Cannizzaro, C. Pre-conceptional and Peri-Gestational Maternal Binge Alcohol Drinking Produces Inheritance of Mood Disturbances and Alcohol Vulnerability in the Adolescent Offspring. Front. Psychiatry 2018, 9, 150. [Google Scholar] [CrossRef]
- Leßmann, V.; Brigadski, T. Mechanisms, locations, and kinetics of synaptic BDNF secretion: An update. Neurosci. Res. 2009, 65, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Barreda Tomás, F.J.; Turko, P.; Heilmann, H.; Trimbuch, T.; Yanagawa, Y.; Vida, I.; Münster-Wandowski, A. BDNF Expression in Cortical GABAergic Interneurons. Int. J. Mol. Sci. 2020, 21, 1567. [Google Scholar] [CrossRef] [PubMed]
- Rafa-Zabłocka, K.; Kreiner, G.; Baginska, M.; Nalepa, I. Selective Depletion of CREB in Serotonergic Neurons Affects the Upregulation of Brain-Derived Neurotrophic Factor Evoked by Chronic Fluoxetine Treatment. Front. Neurosci. 2018, 12, 637. [Google Scholar] [CrossRef] [PubMed]
- Leschik, J.; Gentile, A.; Cicek, C.; Péron, S.; Tevosian, M.; Beer, A.; Radyushkin, K.; Bludau, A.; Ebner, K.; Neumann, I.; et al. Brain-derived neurotrophic factor expression in serotonergic neurons improves stress resilience and promotes adult hippocampal neurogenesis. Prog. Neurobiol. 2022, 217, 102333. [Google Scholar] [CrossRef] [PubMed]
- Hisaoka-Nakashima, K.; Kajitani, N.; Kaneko, M.; Shigetou, T.; Kasai, M.; Matsumoto, C.; Yokoe, T.; Azuma, H.; Takebayashi, M.; Morioka, N.; et al. Amitriptyline induces brain-derived neurotrophic factor (BDNF) mRNA expression through ERK-dependent modulation of multiple BDNF mRNA variants in primary cultured rat cortical astrocytes and microglia. Brain Res. 2016, 1634, 57–67. [Google Scholar] [CrossRef]
- Musazzi, L.; Rimland, J.M.; Ieraci, A.; Racagni, G.; Domenici, E.; Popoli, M. Pharmacological characterization of BDNF promoters I, II and IV reveals that serotonin and norepinephrine input is sufficient for transcription activation. Int. J. Neuropsychopharmacol. 2014, 17, 779–791. [Google Scholar] [CrossRef]
- Tsankova, N.M.; Berton, O.; Renthal, W.; Kumar, A.; Neve, R.L.; Nestler, E.J. Sustained hippocampal chromatin regulation in a mouse model of depression and antidepressant action. Nat. Neurosci. 2006, 9, 519–525. [Google Scholar] [CrossRef]
- Jones, P.A. Functions of DNA methylation: Islands, start sites, gene bodies and beyond. Nat. Rev. Genet. 2012, 13, 484–492. [Google Scholar] [CrossRef]
- Xing, Y.; Sun, T.; Li, G.; Xu, G.; Cheng, J.; Gao, S. The role of BDNF exon I region methylation in the treatment of depression with sertraline and its clinical diagnostic value. J. Clin. Lab. Anal. 2021, 35, e23993. [Google Scholar] [CrossRef]
- Li, L.; Wang, T.; Chen, S.; Yue, Y.; Xu, Z.; Yuan, Y. DNA methylations of brain-derived neurotrophic factor exon VI are associated with major depressive disorder and antidepressant-induced remission in females. J. Affect. Disord. 2021, 295, 101–107. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, C.; Lv, Q.; Bao, C.; Sun, H.; Ma, G.; Fang, Y.; Yi, Z.; Cai, W. Association of DNA methylation in BDNF with escitalopram treatment response in depressed Chinese Han patients. Eur. J. Clin. Pharmacol. 2018, 74, 1011–1020. [Google Scholar] [CrossRef] [PubMed]
- Tadić, A.; Müller-Engling, L.; Schlicht, K.F.; Kotsiari, A.; Dreimüller, N.; Kleimann, A.; Bleich, S.; Lieb, K.; Frieling, H. Methylation of the promoter of brain-derived neurotrophic factor exon IV and antidepressant response in major depression. Mol. Psychiatry 2014, 19, 281–283. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Li, M.; Wang, X.; He, Y.; Xia, Y.; Sweeney, J.A.; Kopp, R.F.; Liu, C.; Chen, C. Drug Response-Related DNA Methylation Changes in Schizophrenia, Bipolar Disorder, and Major Depressive Disorder. Front. Neurosci. 2021, 15, 674273. [Google Scholar] [CrossRef] [PubMed]
- Grassi, D.; Franz, H.; Vezzali, R.; Bovio, P.; Heidrich, S.; Dehghanian, F.; Lagunas, N.; Belzung, C.; Krieglstein, K.; Vogel, T. Neuronal Activity, TGFβ-Signaling and Unpredictable Chronic Stress Modulate Transcription of Gadd45 Family Members and DNA Methylation in the Hippocampus. Cereb. Cortex 2017, 27, 4166–4181. [Google Scholar] [CrossRef] [PubMed]
- Sales, A.J.; Biojone, C.; Terceti, M.S.; Guimarães, F.S.; Gomes, M.V.; Joca, S.R. Antidepressant-like effect induced by systemic and intra-hippocampal administration of DNA methylation inhibitors. Br. J. Pharmacol. 2011, 164, 1711–1721. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.J.; Pei, L.; Li, Y.N.; Zheng, H.; Yang, S.; Wan, Y.; Mao, L.; Xia, Y.P.; He, Q.W.; Li, M.; et al. Alleviative effects of fluoxetine on depressive-like behaviors by epigenetic regulation of BDNF gene transcription in mouse model of post-stroke depression. Sci. Rep. 2017, 7, 14926. [Google Scholar] [CrossRef]
- Seo, M.K.; Ly, N.N.; Lee, C.H.; Cho, H.Y.; Choi, C.M.; Nhu, L.H.; Lee, J.G.; Lee, B.J.; Kim, G.M.; Yoon, B.J.; et al. Early life stress increases stress vulnerability through BDNF gene epigenetic changes in the rat hippocampus. Neuropharmacology 2016, 105, 388–397. [Google Scholar] [CrossRef]
- Hutchinson, A.N.; Deng, J.V.; Cohen, S.; West, A.E. Phosphorylation of MeCP2 at Ser421 contributes to chronic antidepressant action. J. Neurosci. 2012, 32, 14355–14363. [Google Scholar] [CrossRef][Green Version]
- Park, S.W.; Seo, M.K.; Lee, J.G.; Hien, L.T.; Kim, Y.H. Effects of maternal separation and antidepressant drug on epigenetic regulation of the brain-derived neurotrophic factor exon I promoter in the adult rat hippocampus. Psychiatry Clin. Neurosci. 2018, 72, 255–265. [Google Scholar] [CrossRef]
- Homan, P.; Grob, S.; Milos, G.; Schnyder, U.; Eckert, A.; Lang, U.; Hasler, G. The role of BDNF, leptin, and catecholamines in reward learning in bulimia nervosa. Int. J. Neuropsychopharmacol. 2014, 18, 1–8. [Google Scholar] [CrossRef][Green Version]
- Hoepner, C.T.; McIntyre, R.S.; Papakostas, G.I. Impact of supplementation and nutritional interventions on pathogenic processes of mood disorders: A review of the evidence. Nutrients 2021, 13, 767. [Google Scholar] [CrossRef]
- Manosso, L.M.; Moretti, M.; Rodrigues, A.L.S. Nutritional strategies for dealing with depression. Food Funct. 2013, 4, 1776–1793. [Google Scholar] [CrossRef] [PubMed]
- Sarris, J.; Murphy, J.; Mischoulon, D.; Papakostas, G.I.; Fava, M.; Berk, M.; Ng, C.H. Adjunctive nutraceuticals for depression: A systematic review and meta-analyses. Am. J. Psychiatry 2016, 173, 575–587. [Google Scholar] [CrossRef] [PubMed]
- Aly, J.; Engmann, O. The Way to a Human’s Brain Goes Through Their Stomach: Dietary Factors in Major Depressive Disorder. Front. Neurosci. 2020, 14, 582–853. [Google Scholar] [CrossRef] [PubMed]
- Vines, A.; Delattre, A.M.; Lima, M.M.; Rodrigues, L.S.; Suchecki, D.; Machado, R.B.; Tufik, S.; Pereira, S.I.; Zanata, S.M.; Ferraz, A.C. The role of 5-HT₁A receptors in fish oil-mediated increased BDNF expression in the rat hippocampus and cortex: A possible antidepressant mechanism. Neuropharmacology 2012, 62, 184–191. [Google Scholar] [CrossRef]
- Wu, A.; Ying, Z.; Gomez-Pinilla, F. Omega-3 fatty acids supplementation restores mechanisms that maintain brain homeostasis in traumatic brain injury. J. Neurotrauma 2007, 24, 1587–1595. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Moon, H.J.; Kim, S.H. N-3 polyunsaturated fatty acid consumption produces neurobiological effects associated with prevention of depression in rats after the forced swimming test. J. Nutr. Biochem. 2012, 23, 924–928. [Google Scholar] [CrossRef]
- Paduchová, Z.; Katrenčíková, B.; Vaváková, M.; Laubertová, L.; Nagyová, Z.; Garaiova, I.; Ďuračková, Z.; Trebatická, J. The Effect of Omega-3 Fatty Acids on Thromboxane, Brain-Derived Neurotrophic Factor, Homocysteine, and Vitamin D in Depressive Children and Adolescents: Randomized Controlled Trial. Nutrients 2021, 13, 1095. [Google Scholar] [CrossRef]
- Jazayeri, S.; Tehrani-Doost, M.; Keshavarz, S.A.; Hosseini, M.; Djazayery, A.; Amini, H.; Jalali, M.; Peet, M. Comparison of therapeutic effects of omega-3 fatty acid eicosapentaenoic acid and fluoxetine, separately and in combination, in major depressive disorder. Aust. N. Z. J. Psychiatry 2008, 42, 192–198. [Google Scholar] [CrossRef]
- Carney, R.M.; Freedland, K.E.; Rubin, E.H.; Rich, M.W.; Steinmeyer, B.C.; Harris, W.S. Omega-3 augmentation of sertraline in treatment of depression in patients with coronary heart disease: A randomized controlled trial. JAMA 2009, 302, 1651–1657. [Google Scholar] [CrossRef]
- Jahangard, L.; Sadeghi, A.; Ahmadpanah, M.; Holsboer-Trachsler, E.; Sadeghi Bahmani, D.; Haghighi, M.; Brand, S. Influence of adjuvant omega-3-polyunsaturated fatty acids on depression, sleep, and emotion regulation among outpatients with major depressive disorders—Results from a double-blind, randomized and placebo-controlled clinical trial. J. Psychiatr. Res. 2018, 107, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Peet, M.; Horrobin, D.F. A dose-ranging study of the effects of ethyl-eicosapentaenoate in patients with ongoing depression despite apparently adequate treatment with standard drugs. Arch. Gen. Psychiatry 2002, 59, 913–919. [Google Scholar] [CrossRef]
- Su, K.P.; Huang, S.Y.; Chiu, C.C.; Shen, W.W. Omega-3 fatty acids in major depressive disorder: A preliminary double-blind, placebo-controlled trial. Eur. Neuropsychopharmacol. 2003, 13, 267–271. [Google Scholar] [CrossRef]
- Nemets, B.; Stahl, Z.; Belmaker, R.H. Addition of omega-3 fatty acid to maintenance medication treatment for recurrent unipolar depressive disorder. Am. J. Psychiatry 2002, 159, 477–479. [Google Scholar] [CrossRef] [PubMed]
- Silvers, K.M.; Woolley, C.C.; Hamilton, F.C.; Watts, P.M.; Watson, R.A. Randomised double-blind placebo-controlled trial of fish oil in the treatment of depression. Prostaglandins. Leukot. Essent. Fat. Acids 2005, 72, 211–218. [Google Scholar] [CrossRef]
- Mozaffari-Khosravi, H.; Yassini-Ardakani, M.; Karamati, M.; Shariati-Bafghi, S.E. Eicosapentaenoic acid versus docosahexaenoic acid in mild-to-moderate depression: A randomized, double-blind, placebo-controlled trial. Eur. Neuropsychopharmacol. 2013, 23, 636–644. [Google Scholar] [CrossRef]
- Smith, D.J.; Sarris, J.; Dowling, N.; O’Connor, M.; Ng, C.H. Adjunctive low-dose docosahexaenoic acid (DHA) for major depression: An open-label pilot trial. Nutr. Neurosci. 2018, 21, 224–228. [Google Scholar] [CrossRef]
- Gertsik, L.; Poland, R.E.; Bresee, C.; Rapaport, M.H. Omega-3 fatty acid augmentation of citalopram treatment for patients with major depressive disorder. J. Clin. Psychopharmacol. 2012, 32, 61–64. [Google Scholar] [CrossRef]
- Mocking, R.J.; Verburg, H.F.; Westerink, A.M.; Assies, J.; Vaz, F.M.; Koeter, M.W.; Ruhé, H.G.; Schene, A.H. Fatty acid metabolism and its longitudinal relationship with the hypothalamic-pituitary-adrenal axis in major depression: Associations with prospective antidepressant response. Psychoneuroendocrinology 2015, 59, 1–13. [Google Scholar] [CrossRef]
- Ghazizadeh-Hashemi, M.; Ghajar, A.; Shalbafan, M.R.; Ghazizadeh-Hashemi, F.; Afarideh, M.; Malekpour, F.; Ghaleiha, A.; Ardebili, M.E.; Akhondzadeh, S. Palmitoylethanolamide as adjunctive therapy in major depressive disorder: A double-blind, randomized and placebo-controlled trial. J. Affect. Disord. 2018, 232, 127–133. [Google Scholar] [CrossRef]
- Able, J.A.; Liu, Y.; Jandacek, R.; Rider, T.; Tso, P.; McNamara, R.K. Omega-3 fatty acid deficient male rats exhibit abnormal behavioral activation in the forced swim test following chronic fluoxetine treatment: Association with altered 5-HT1A and alpha2A adrenergic receptor expression. J. Psychiatr. Res. 2014, 50, 42–50. [Google Scholar] [CrossRef]
- McNamara, R.K.; Able, J.A.; Liu, Y.; Jandacek, R.; Rider, T.; Tso, P.; Lipton, J.W. Omega-3 fatty acid deficiency does not alter the effects of chronic fluoxetine treatment on central serotonin turnover or behavior in the forced swim test in female rats. Pharmacol. Biochem. Behav. 2013, 114, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Papakostas, G.I.; Ionescu, D.F. Towards new mechanisms: An update on therapeutics for treatment-resistant major depressive disorder. Mol. Psychiatry 2015, 20, 1142–1150. [Google Scholar] [CrossRef] [PubMed]
- Micale, V.; Scapagnini, G.; Colombrita, C.; Mazzola, C.; Alkon, D.L.; Drago, F. Behavioral effects of dietary cholesterol in rats tested in experimental models of mild stress and cognition tasks. Eur. Neuropsychopharmacol. 2008, 18, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Trepanowski, J.F.; Canale, R.E.; Marshall, K.E.; Kabir, M.M.; Bloomer, R.J. Impact of caloric and dietary restriction regimens on markers of health and longevity in humans and animals: A summary of available findings. Nutr. J. 2011, 10, 107. [Google Scholar] [CrossRef] [PubMed]
- Elesawy, B.H.; Raafat, B.M.; Al Muqbali, A.; Abbas, A.M.; Sakr, H.F. The Impact of Intermittent Fasting on Brain-Derived Neurotrophic Factor, Neurotrophin 3, and Rat Behavior in a Rat Model of Type 2 Diabetes Mellitus. Brain Sci. 2021, 11, 242. [Google Scholar] [CrossRef]
- Stapel, B.; Fraccarollo, D.; Westhoff-Bleck, M.; Bauersachs, J.; Lichtinghagen, R.; Jahn, K.; Burkert, A.; Buchholz, V.; Bleich, S.; Frieling, H.; et al. Impact of fasting on stress systems and depressive symptoms in patients with major depressive disorder: A cross-sectional study. Sci. Rep. 2022, 12, 7642. [Google Scholar] [CrossRef]
- Li, B.; Zhao, J.; Lv, J.; Tang, F.; Liu, L.; Sun, Z.; Wang, L.; Siwela, S.P.; Wang, Y.; Song, Y.; et al. Additive antidepressant-like effects of fasting with imipramine via modulation of 5-HT2 receptors in the mice. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2014, 48, 199–206. [Google Scholar] [CrossRef]
- Liu, L.; Li, B.; Zhou, Y.; Wang, L.; Tang, F.; Shao, D.; Jiang, X.; Zhao, H.; Cui, R.; Li, Y. Antidepressant-like effect of Fuzi total alkaloid on ovariectomized mice. J. Pharmacol. Sci. 2012, 120, 280–287. [Google Scholar] [CrossRef]
- Sial, O.K.; Gnecco, T.; Cardona-Acosta, A.M.; Vieregg, E.; Cardoso, E.A.; Parise, L.F.; Bolaños-Guzmán, C.A. Exposure to Vicarious Social Defeat Stress and Western-Style Diets During Adolescence Leads to Physiological Dysregulation, Decreases in Reward Sensitivity, and Reduced Antidepressant Efficacy in Adulthood. Front. Neurosci. 2021, 15, 701–919. [Google Scholar] [CrossRef]
- Ågren, H.; Niklasson, F. Creatinine and creatine in CSF: Indices of brain energy metabolism in depression. J. Neural Transm. 1988, 74, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Cunha, M.P.; Pazini, F.L.; Lieberknecht, V.; Rodrigues, A.L.S. Subchronic administration of creatine produces antidepressant-like effect by modulating hippocampal signaling pathway mediated by FNDC5/BDNF/Akt in mice. J. Psychiatr. Res. 2018, 104, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Lyoo, I.K.; Yoon, S.; Kim, T.S.; Hwang, J.; Kim, J.E.; Won, W.; Bae, S.; Renshaw, P.F. A randomized, double-blind placebo-controlled trial of oral creatine monohydrate augmentation for enhanced response to a selective serotonin reuptake inhibitor in women with major depressive disorder. Am. J. Psychiatry 2012, 169, 937–945. [Google Scholar] [CrossRef] [PubMed]
- Kondo, D.G.; Sung, Y.H.; Hellem, T.L.; Fiedler, K.K.; Shi, X.; Jeong, E.K.; Renshaw, P.F. Open-label adjunctive creatine for female adolescents with SSRI-resistant major depressive disorder: A 31-phosphorus magnetic resonance spectroscopy study. J. Affect. Disord. 2011, 135, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Allen, P.J.; D’Anci, K.E.; Kanarek, R.B.; Renshaw, P.F. Sex-specific antidepressant effects of dietary creatine with and without sub-acute fluoxetine in rats. Pharmacol. Biochem. Behav. 2012, 101, 588–601. [Google Scholar] [CrossRef] [PubMed]
- Cowen, P.J. Psychopharmacology of 5-HT(1A) receptors. Nucl. Med. Biol. 2000, 27, 437–439. [Google Scholar] [CrossRef]
- Cowen, P.J.; Browning, M. What has serotonin to do with depression? World Psychiatry 2015, 14, 158–160. [Google Scholar] [CrossRef]
- Weber, L.J.; Horita, A. A study of 5-hydroxytryptamine formation from L-tryptophan in the brain and other tissues. Biochem. Pharmacol. 1965, 14, 1141–1149. [Google Scholar] [CrossRef]
- Van Der Stelt, H.M.; Broersen, L.M.; Olivier, B.; Westenberg, H.G.M. Effects of dietary tryptophan variations on extracellular serotonin in the dorsal hippocampus of rats. Psychopharmacology 2004, 172, 137–144. [Google Scholar] [CrossRef]
- Levitan, R.; Shen, J.H.; Jindal, R.; Driver, H.S.; Kennedy, S.H.; Shapiro, C.M. Preliminary randomized double-blind placebo-controlled trial of tryptophan combined with fluoxetine to treat major depressive disorder: Antidepressant and hypnotic effects. J. Psychiatry Neurosci. 2000, 25, 337–346. [Google Scholar]
- Wålinder, J.; Skott, A.; Carlsson, A.; Nagy, A.; Roos, B.E. Potentiation of the antidepressant action of clomipramine by tryptophan. Arch. Gen. Psychiatry 1976, 33, 1384–1389. [Google Scholar] [CrossRef]
- Glassman, A.H.; Platman, S.R. Potentiation of a monoamine oxidase inhibitor by tryptophan. J. Psychiatr. Res. 1969, 7, 83–88. [Google Scholar] [CrossRef]
- Arnold, O.; Saletu, B.; Anderer, P.; Assandri, A.; di Padova, C.; Corrado, M. Saletu-Zyhlarz, G.M. Double-blind, placebo-controlled pharmacodynamic studies with a nutraceutical and a pharmaceutical dose of ademetionine (SAMe) in elderly subjects, utilizing EEG mapping and psychometry. Eur. Neuropsychopharmacol. 2005, 15, 533–543. [Google Scholar] [CrossRef] [PubMed]
- Ielmini, M.; Caselli, I.; Ceccon, F.; Diurni, M.; Poloni, N.; Callegari, C. Selective Serotonin Reuptake Inhibitors and Nutraceutical Combination in Major Depression Disorder: A Case-Control Study. Psychopharmacol. Bull. 2021, 51, 31–39. [Google Scholar]
- De Berardis, D.; Orsolini, L.; Serroni, N.; Girinelli, G.; Iasevoli, F.; Tomasetti, C.; de Bartolomeis, A.; Mazza, M.; Valchera, A.; Fornaro, M.; et al. A comprehensive review on the efficacy of S-Adenosyl-L-methionine in Major Depressive Disorder. CNS Neurol. Disord. Drug Targets 2016, 15, 35–44. [Google Scholar] [CrossRef]
- Papakostas, G.I.; Mischoulon, D.; Shyu, I.; Alpert, J.E.; Fava, M. S-adenosyl methionine (SAMe) augmentation of serotonin reuptake inhibitors for antidepressant nonresponders with major depressive disorder: A double-blind, randomized clinical trial. Am. J. Psychiatry 2010, 167, 942–948. [Google Scholar] [CrossRef]
- Sarris, J.; Murphy, J.; Stough, C.; Mischoulon, D.; Bousman, C.; MacDonald, P.; Adams, L.; Nazareth, S.; Oliver, G.; Cribb, L.; et al. S-Adenosylmethionine (SAMe) monotherapy for depression: An 8-week double-blind, randomised, controlled trial. Psychopharmacology 2020, 237, 209–218. [Google Scholar] [CrossRef]
- Panizzutti, B.; Bortolasci, C.; Hasebe, K.; Kidnapillai, S.; Gray, L.; Walder, K.; Berk, M.; Mohebbi, M.; Dodd, S.; Gama, C.; et al. Mediator effects of parameters of inflammation and neurogenesis from a N-acetyl cysteine clinical-trial for bipolar depression. Acta Neuropsychiatr. 2018, 30, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.C.; Chan, M.H.; Lee, M.Y.; Chen, Y.C.; Chen, H.H. N,N-dimethylglycine differentially modulates psychotomimetic and antidepressant-like effects of ketamine in mice. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2016, 71, 7–13. [Google Scholar] [CrossRef]
- Lin, J.C.; Lee, M.Y.; Chan, M.H.; Chen, Y.C.; Chen, H.H. Betaine enhances antidepressant-like, but blocks psychotomimetic effects of ketamine in mice. Psychopharmacology 2016, 233, 3223–3235. [Google Scholar] [CrossRef]
- Lazarou, C.; Kapsou, M. The role of folic acid in prevention and treatment of depression: An overview of existing evidence and implications for practice. Complement. Ther. Clin. Pract. 2010, 16, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.J.; Carney, S.M.; Goodwin, G.M.; Geddes, J.R. Folate for depressive disorders: Systematic review and meta-analysis of randomized controlled trials. J. Psychopharmacol. 2004, 18, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Bottiglieri, T.; Laundy, M.; Crellin, R.; Toone, B.K.; Carney, M.W.; Reynolds, E.H. Homocysteine, folate, methylation, and monoamine metabolism in depression. J. Neurol. Neurosurg. Psychiatry 2000, 69, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Morris, D.W.; Trivedi, M.H.; Rush, A.J. Folate and unipolar depression. J. Altern. Complement. Med. 2008, 14, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Cong, Y.; Liu, H. Folic acid ameliorates depression-like behaviour in a rat model of chronic unpredictable mild stress. BMC Neurosci. 2020, 21, 1. [Google Scholar] [CrossRef]
- Coppen, A.; Bailey, J. Enhancement of the antidepressant action of fluoxetine by folic acid: A randomised, placebo controlled trial. J. Affect. Disord. 2000, 60, 121–130. [Google Scholar] [CrossRef]
- Resler, G.; Lavie, R.; Campos, J.; Mata, S.; Urbina, M.; García, A.; Apitz, R.; Lima, L. Effect of folic acid combined with fluoxetine in patients with major depression on plasma homocysteine and vitamin B12, and serotonin levels in lymphocytes. Neuroimmunomodulation 2008, 15, 145–152. [Google Scholar] [CrossRef]
- Papakostas, G.I.; Öngür, D.; Iosifescu, D.V.; Mischoulon, D.; Fava, M. Cholesterol in mood and anxiety disorders: Review of the literature and new hypotheses. Eur. Neuropsychopharmacol. 2004, 14, 135–142. [Google Scholar] [CrossRef]
- Papakostas, G.I.; Shelton, R.C.; Zajecka, J.M.; Etemad, B.; Rickels, K.; Clain, A.; Baer, L.; Dalton, E.D.; Sacco, G.R.; Schoenfeld, D.; et al. L-methylfolate as adjunctive therapy for SSRI-resistant major depression: Results of two randomized, double-blind, parallel-sequential trials. Am. J. Psychiatry 2012, 169, 1267–1274. [Google Scholar] [CrossRef]
- Ginsberg, L.; Oubre, A.; Daoud, Y. L-methylfolate Plus SSRI or SNRI from Treatment Initiation Compared to SSRI or SNRI Monotherapy in a Major Depressive Episode. Innov. Clin. Neurosci. 2011, 8, 19–28. [Google Scholar]
- Bedson, E.; Bell, D.; Carr, D.; Carter, B.; Hughes, D.; Jorgensen, A.; Lewis, H.; Lloyd, K.; McCaddon, A.; Moat, S.; et al. Folate Augmentation of Treatment—Evaluation for Depression (FolATED): Randomised trial and economic evaluation. Health Technol. Assess. 2014, 18, 1–159. [Google Scholar] [CrossRef] [PubMed]
- Syed, E.; Wasay, M.; Awan, S. Vitamin B12 supplementation in treating major depressive disorder: A randomized controlled trial. Open Neurol. J. 2013, 7, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Trautmann, C.; Bock, A.; Urbach, A.; Hübner, A.; Engmann, O. Acute vitamin B12 supplementation evokes antidepressant response and alters Ntrk-2. Neuropharmacology 2020, 171, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Christensen, H.; Aiken, A.; Batterham, P.J.; Walker, J.; Mackinnon, A.J.; Fenech, M.; Hickie, I.B. No clear potentiation of antidepressant medication effects by folic acid + vitamin B12 in a large community sample. J. Affect. Disord. 2011, 130, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Almeida, O.P.; Ford, A.H.; Hirani, V.; Singh, V.; van Bockxmeer, F.M.; McCaul, K.; Flicker, L. B vitamins to enhance treatment response to antidepressants in middle-aged and older adults: Results from the B-VITAGE randomised, double-blind, placebo-controlled trial. Br. J. Psychiatry 2014, 205, 450–457. [Google Scholar] [CrossRef]
- Amr, M.; El-Mogy, A.; Shams, T.; Vieira, K.; Lakhan, S.E. Efficacy of vitamin C as an adjunct to fluoxetine therapy in pediatric major depressive disorder: A randomized, double-blind, placebo-controlled pilot study. Nutr. J. 2013, 12, 31. [Google Scholar] [CrossRef]
- Khoraminya, N.; Tehrani-Doost, M.; Jazayeri, S.; Hosseini, A.; Djazayery, A. Therapeutic effects of vitamin D as adjunctive therapy to fluoxetine in patients with major depressive disorder. Aust. N. Z. J. Psychiatry 2013, 47, 271–275. [Google Scholar] [CrossRef]
- Sahraian, A.; Ghanizadeh, A.; Kazemeini, F. Vitamin C as an adjuvant for treating major depressive disorder and suicidal behavior, a randomized placebo-controlled clinical trial. Trials 2015, 16, 94. [Google Scholar] [CrossRef]
- Zanetidou, S.; Belvederi Murri, M.; Buffa, A.; Malavolta, N.; Anzivino, F.; Bertakis, K. Vitamin D supplements in geriatric major depression. Int. J. Geriatr. Psychiatry 2011, 26, 1209–1210. [Google Scholar] [CrossRef]
- Xu, Y.; Liang, L. Vitamin D3/vitamin D receptor signaling mitigates symptoms of post-stroke depression in mice by upregulating hippocampal BDNF expression. Neurosci. Res. 2021, 170, 306–313. [Google Scholar] [CrossRef]
- McCall, K.A.; Huang, C.C.; Fierke, C.A. Function and mechanism of zinc metalloenzymes. J. Nutr. 2000, 130, 1437S–1446S. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.S. Discovery of human zinc deficiency: 50 years later. J. Trace Elem. Med. Biol. 2012, 26, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Mlyniec, K. Interaction between Zinc, GPR39, BDNF and Neuropeptides in Depression. Curr. Neuropharmacol. 2021, 19, 2012–2019. [Google Scholar] [CrossRef] [PubMed]
- Młyniec, K.; Nowak, G. Zinc deficiency induces behavioral alterations in the tail suspension test in mice. Effect of antidepressants. Pharmacol. Rep. 2012, 64, 249–255. [Google Scholar] [CrossRef]
- Młyniec, K.; Budziszewska, B.; Reczyński, W.; Sowa-Kućma, M.; Nowak, G. The role of the GPR39 receptor in zinc deficient-animal model of depression. Behav. Brain Res. 2013, 238, 30–35. [Google Scholar] [CrossRef]
- Młyniec, K.; Budziszewska, B.; Reczyński, W.; Doboszewska, U.; Pilc, A.; Nowak, G. Zinc deficiency alters responsiveness to antidepressant drugs in mice. Pharmacol. Rep. 2013, 65, 579–592. [Google Scholar] [CrossRef]
- Doboszewska, U.; Szewczyk, B.; Sowa-Kućma, M.; Młyniec, K.; Rafało, A.; Ostachowicz, B.; Lankosz, M.; Nowak, G. Antidepressant activity of fluoxetine in the zinc deficiency model in rats involves the NMDA receptor complex. Behav. Brain Res. 2015, 287, 323–330. [Google Scholar] [CrossRef]
- Ranjbar, E.; Shams, J.; Sabetkasaei, M.; M-Shirazi, M.; Rashidkhani, B.; Mostafavi, A.; Bornak, E.; Nasrollahzadeh, J. Effects of zinc supplementation on efficacy of antidepressant therapy, inflammatory cytokines, and brain-derived neurotrophic factor in patients with major depression. Nutr. Neurosci. 2014, 17, 65–71. [Google Scholar] [CrossRef]
- Nowak, G.; Siwek, M.; Dudek, D.; Zieba, A.; Pilc, A. Effect of zinc supplementation on antidepressant therapy in unipolar depression: A preliminary placebo-controlled study. Pol. J. Pharmacol. 2003, 55, 1143–1147. [Google Scholar]
- Siwek, M.; Dudek, D.; Schlegel-Zawadzka, M.; Morawska, A.; Piekoszewski, W.; Opoka, W.; Zieba, A.; Pilc, A.; Popik, P.; Nowak, G. Serum zinc level in depressed patients during zinc supplementation of imipramine treatment. J. Affect. Disord. 2010, 126, 447–452. [Google Scholar] [CrossRef]
- Siwek, M.; Dudek, D.; Paul, I.A.; Sowa-Kućma, M.; Zieba, A.; Popik, P.; Pilc, A.; Nowak, G. Zinc supplementation augments efficacy of imipramine in treatment resistant patients: A double blind, placebo-controlled study. J. Affect. Disord. 2009, 118, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Swaminathan, R. Magnesium metabolism and its disorders. Clin. Biochem. Rev. 2003, 24, 47–66. [Google Scholar] [PubMed]
- Lang, U.E.; Beglinger, C.; Schweinfurth, N.; Walter, M.; Borgwardt, S. Nutritional aspects of depression. Cell. Physiol. Biochem. 2015, 37, 1029–1043. [Google Scholar] [CrossRef] [PubMed]
- Ryszewska-Pokraśniewicz, B.; Mach, A.; Skalski, M.; Januszko, P.; Wawrzyniak, Z.M.; Poleszak, E.; Nowak, G.; Pilc, A.; Radziwoń-Zaleska, M. Effects of magnesium supplementation on unipolar depression: A placebo-controlled study and review of the importance of dosing and magnesium status in the therapeutic response. Nutrients 2018, 10, 1014. [Google Scholar] [CrossRef]
- Duncan, L.E.; Keller, M.C. A critical review of the first 10 years of candidate gene-by-environment interaction research in psychiatry. Am. J. Psychiatry 2011, 168, 1041–1049. [Google Scholar] [CrossRef]
- Domschke, K.; Tidow, N.; Schwarte, K.; Deckert, J.; Lesch, K.P.; Arolt, V.; Zwanzger, P.; Baune, B.T. Serotonin transporter gene hypomethylation predicts impaired antidepressant treatment response. Int. J. Neuropsychopharmacol. 2014, 17, 1167–1176. [Google Scholar] [CrossRef]
- Kang, H.J.; Kim, J.M.; Stewart, R.; Kim, S.Y.; Bae, K.Y.; Kim, S.W.; Shin, I.S.; Shin, M.G.; Yoon, J.S. Association of SLC6A4 methylation with early adversity, characteristics and outcomes in depression. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2013, 44, 23–28. [Google Scholar] [CrossRef]
- Zimmermann, N.; Zschocke, J.; Perisic, T.; Yu, S.; Holsboer, F.; Rein, T. Antidepressants inhibit DNA methyltransferase 1 through reducing G9a levels. Biochem. J. 2012, 448, 93–102. [Google Scholar] [CrossRef]
- Detich, N.; Bovenzi, V.; Szyf, M. Valproate induces replication-independent active DNA demethylation. J. Biol. Chem. 2003, 278, 27586–27592. [Google Scholar] [CrossRef]
- Klengel, T.; Binder, E.B. Gene×environment interactions in the prediction of response to antidepressant treatment. Int. J. Neuropsychopharmacol. 2013, 16, 701–711. [Google Scholar] [CrossRef]
- Tomita, T.; Yasui-Furukori, N.; Nakagami, T.; Kaneda, A.; Kaneko, S. The association between sunshine duration and paroxetine response time in patients with major depressive disorder. J. Affect. Disord. 2012, 136, 1067–1071. [Google Scholar] [CrossRef] [PubMed]
- Kendler, K.S.; Kessler, R.C.; Walters, E.E.; MacLean, C.; Neale, M.C.; Heath, A.C.; Eaves, L.J. Stressful life events, genetic liability, and Onset of an episode of Major depression in Women. Depress. Sci. Ment. Heal. 2013, 152, 833–842. [Google Scholar] [CrossRef]
- Sullivan, P.F.; Neale, M.C.; Kendler, K.S. Genetic epidemiology of major depression: Review and meta-analysis. Am. J. Psychiatry 2000, 157, 1552–1562. [Google Scholar] [CrossRef] [PubMed]
- Ising, M.; Lucae, S.; Binder, E.B.; Bettecken, T.; Uhr, M.; Ripke, S.; Kohli, M.A.; Hennings, J.M.; Horstmann, S.; Kloiber, S.; et al. A genomewide association study points to multiple loci that predict antidepressant drug treatment outcome in depression. Arch. Gen. Psychiatry 2009, 66, 966–975. [Google Scholar] [CrossRef]
- Garriock, H.A.; Kraft, J.B.; Shyn, S.I.; Peters, E.J.; Yokoyama, J.S.; Jenkins, G.D.; Reinalda, M.S.; Slager, S.L.; McGrath, P.J.; Hamilton, S.P. A Genomewide Association Study of Citalopram Response in Major Depressive Disorder. Biol. Psychiatry 2010, 67, 133–138. [Google Scholar] [CrossRef]
- Uher, R.; Perroud, N.; Ng, M.Y.; Hauser, J.; Henigsberg, N.; Maier, W.; Mors, O.; Placentino, A.; Rietschel, M.; Souery, D.; et al. Genome-wide pharmacogenetics of antidepressant response in the GENDEP project. Am. J. Psychiatry 2010, 167, 555–564. [Google Scholar] [CrossRef]
- Horstmann, S.; Lucae, S.; Menke, A.; Hennings, J.M.; Ising, M.; Roeske, D.; Müller-Myhsok, B.; Holsboer, F.; Binder, E.B. Polymorphisms in GRIK4, HTR2A, and FKBP5 show interactive effects in predicting remission to antidepressant treatment. Neuropsychopharmacology 2010, 35, 727–740. [Google Scholar] [CrossRef]
- Poggini, S.; Matte Bon, G.; Golia, M.T.; Ciano Albanese, N.; Viglione, A.; Poleggi, A.; Limatola, C.; Maggi, L.; Branchi, I. Selecting antidepressants according to a drug-by-environment interaction: A comparison of fluoxetine and minocycline effects in mice living either in enriched or stressful conditions. Behav. Brain Res. 2021, 408, 113256. [Google Scholar] [CrossRef]
- Alboni, S.; van Dijk, R.M.; Poggini, S.; Milior, G.; Perrotta, M.; Drenth, T.; Brunello, N.; Wolfer, D.P.; Limatola, C.; Amrein, I.; et al. Fluoxetine effects on molecular, cellular and behavioral endophenotypes of depression are driven by the living environment. Mol. Psychiatry 2017, 22, 635. [Google Scholar] [CrossRef]
- Krishnan, V.; Nestler, E.J. The molecular neurobiology of depression. Nature 2008, 455, 894–902. [Google Scholar] [CrossRef]
- Fitzgerald, P.J.; Yen, J.Y.; Watson, B.O. Stress-sensitive antidepressant-like effects of ketamine in the mouse forced swim test. PLoS ONE 2019, 14, e0215554. [Google Scholar] [CrossRef] [PubMed]
- Berton, O.; McClung, C.A.; Dileone, R.J.; Krishnan, V.; Renthal, W.; Russo, S.J.; Graham, D.; Tsankova, N.M.; Bolanos, C.A.; Rios, M.; et al. Essential role of BDNF in the mesolimbic dopamine pathway in social defeat stress. Science 2006, 311, 864–868. [Google Scholar] [CrossRef] [PubMed]
- Holsboer, F. The corticosteroid receptor hypothesis of depression. Neuropsychopharmacology 2000, 23, 477–501. [Google Scholar] [CrossRef]
- Wolkowitz, O.M.; Reus, V.I.; Manfredi, F.; Ingbar, J.; Brizendine, L.; Weingartner, H. Ketoconazole administration in hypercortisolemic depression. Am. J. Psychiatry 1993, 150, 810–812. [Google Scholar] [PubMed]
- Chang, H.S.; Won, E.; Lee, H.Y.; Ham, B.J.; Lee, M.S. Association analysis for corticotropin releasing hormone polymorphisms with the risk of major depressive disorder and the response to antidepressants. Behav. Brain Res. 2015, 292, 116–124. [Google Scholar] [CrossRef]
- Duman, R.S.; Li, N. A neurotrophic hypothesis of depression: Role of synaptogenesis in the actions of NMDA receptor antagonists. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 2475–2484. [Google Scholar] [CrossRef]
- Nanni, V.; Uher, R.; Danese, A. Childhood maltreatment predicts unfavorable course of illness and treatment outcome in depression: A meta-analysis. Am. J. Psychiatry 2012, 169, 141–151. [Google Scholar] [CrossRef]
- Nemeroff, C.B.; Heim, C.M.; Thase, M.E.; Klein, D.N.; Rush, A.J.; Schatzberg, A.F.; Ninan, P.T.; McCullough, J.P., Jr.; Weiss, P.M.; Dunner, D.L.; et al. Differential responses to psychotherapy versus pharmacotherapy in patients with chronic forms of major depression and childhood trauma. Proc. Natl. Acad. Sci. USA 2003, 100, 14293–14296. [Google Scholar] [CrossRef]
- Navailles, S.; Hof, P.R.; Schmauss, C. Antidepressant drug-induced stimulation of mouse hippocampal neurogenesis is age-dependent and altered by early life stress. J. Comp. Neurol. 2008, 509, 372–381. [Google Scholar] [CrossRef]
- Wearick-Silva, L.E.; Orso, R.; Martins, L.A.; Creutzberg, K.C.; Centeno-Silva, A.; Xavier, L.L.; Grassi-Oliveira, R.; Mestriner, R.G. Dual influences of early life stress induced by limited bedding on walking adaptability and Bdnf/TrkB and Drd1/Drd2 gene expression in different mouse brain regions. Behav. Brain Res. 2019, 359, 66–72. [Google Scholar] [CrossRef]
- Zheng, Y.; Fan, W.; Zhang, X.; Dong, E. Gestational stress induces depressive-like and anxiety-like phenotypes through epigenetic regulation of BDNF expression in offspring hippocampus. Epigenetics 2016, 11, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Pallarés, M.E.; Monteleone, M.C.; Pastor, V.; Grillo Balboa, J.; Alzamendi, A.; Brocco, M.A.; Antonelli, M.C. Early-Life Stress Reprograms Stress-Coping Abilities in Male and Female Juvenile Rats. Mol. Neurobiol. 2021, 58, 5837–5856. [Google Scholar] [CrossRef] [PubMed]
- Coley, E.J.L.; Demaestri, C.; Ganguly, P.; Honeycutt, J.A.; Peterzell, S.; Rose, N.; Ahmed, N.; Holschbach, M.; Trivedi, M.; Brenhouse, H.C. Cross-Generational Transmission of Early Life Stress Effects on HPA Regulators and Bdnf Are Mediated by Sex, Lineage, and Upbringing. Front. Behav. Neurosci. 2019, 13, 101. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, E.; Erol, K.; Ulupinar, E. Effects of sertraline on behavioral alterations caused by environmental enrichment and social isolation. Pharmacol. Biochem. Behav. 2012, 101, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Nasca, C.; Barnhill, O.; DeAngelis, P.; Watson, K.; Lin, J.; Beasley, J.; Young, S.P.; Myoraku, A.; Dobbin, J.; Bigio, B.; et al. Multidimensional predictors of antidepressant responses: Integrating mitochondrial, genetic, metabolic and environmental factors with clinical outcomes. Neurobiol. Stress 2021, 15, 100–407. [Google Scholar] [CrossRef]
- Takeuchi, N.; Nonen, S.; Kato, M.; Wakeno, M.; Takekita, Y.; Kinoshita, T.; Kugawa, F. Therapeutic Response to Paroxetine in Major Depressive Disorder Predicted by DNA Methylation. Neuropsychobiology 2018, 75, 81–88. [Google Scholar] [CrossRef]
- Branchi, I.; D’Andrea, I.; Sietzema, J.; Fiore, M.; Di Fausto, V.; Aloe, L.; Alleva, E. Early social enrichment augments adult hippocampal BDNF levels and survival of BRDU-positive cells while increasing anxiety- and ‘depression’-like behavior. J. Neurosci. Res. 2006, 83, 965–973. [Google Scholar] [CrossRef]
- Carek, P.J.; Laibstain, S.E.; Carek, S.M. Exercise for the treatment of depression and anxiety. Int. J. Psychiatry Med. 2011, 41, 15–28. [Google Scholar] [CrossRef]
- Karpova, N.N.; Rantamäki, T.; Di Lieto, A.; Lindemann, L.; Hoener, M.C.; Castrén, E. Darkness reduces BDNF expression in the visual cortex and induces repressive chromatin remodeling at the BDNF gene in both hippocampus and visual cortex. Cell. Mol. Neurobiol. 2010, 30, 1117–1123. [Google Scholar] [CrossRef]
- Sadighparvar, S.; Darband, S.G.; Yousefi, B.; Kaviani, M.; Ghaderi-Pakdel, F.; Mihanfar, A.; Babaei, G.; Mobaraki, K.; Majidinia, M. Combination of quercetin and exercise training attenuates depression in rats with 1,2-dimethylhydrazine-induced colorectal cancer: Possible involvement of inflammation and BDNF signalling. Exp. Physiol. 2020, 105, 1598–1609. [Google Scholar] [CrossRef]
- Lu, J.; Xu, Y.; Hu, W.; Gao, Y.; Ni, X.; Sheng, H.; Liu, Y. Exercise ameliorates depression-like behavior and increases hippocampal BDNF level in ovariectomized rats. Neurosci. Lett. 2014, 573, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.W.; Jung, S.Y.; Lee, S.W.; Lee, S.J.; Seo, T.B.; Kim, Y.P.; Kim, D.Y. Treadmill exercise ameliorates social isolation-induced depression through neuronal generation in rat pups. J. Exerc. Rehabil. 2017, 13, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.-W.; Lim, B.-V.; Kim, K.; Seo, J.-H.; Kim, C.-J. Treadmill exercise alleviates stress-induced impairment of social interaction through 5-hydroxytryptamine 1A receptor activation in rats. J. Exerc. Rehabil. 2015, 11, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Sleiman, S.F.; Henry, J.; Al-Haddad, R.; El Hayek, L.; Abou Haidar, E.; Stringer, T.; Ulja, D.; Karuppagounder, S.S.; Holson, E.B.; Ratan, R.R.; et al. Exercise promotes the expression of brain derived neurotrophic factor (BDNF) through the action of the ketone body β-hydroxybutyrate. elife 2016, 5, e15092. [Google Scholar] [CrossRef]
- Voisey, J.; Lawford, B.; Bruenig, D.; Harvey, W.; Morris, C.P.; Young, R.M.; Mehta, D.; PTSD Initiative. Differential BDNF methylation in combat exposed veterans and the association with exercise. Gene 2019, 698, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.; Lewis, D.A.; Sibille, E. The Role of BDNF in Age-Dependent Changes of Excitatory and Inhibitory Synaptic Markers in the Human Prefrontal Cortex. Neuropsychopharmacology 2016, 41, 3080–3091. [Google Scholar] [CrossRef]
- von Bohlen und Halbach, O.; Krause, S.; Medina, D.; Sciarretta, C.; Minichiello, L.; Unsicker, K. Regional- and age-dependent reduction in trkB receptor expression in the hippocampus is associated with altered spine morphologies. Biol. Psychiatry 2006, 59, 793–800. [Google Scholar] [CrossRef]
- Qiao, H.; Li, M.X.; Xu, C.; Chen, H.B.; An, S.C.; Ma, X.M. Dendritic Spines in Depression: What We Learned from Animal Models. Neural Plast. 2016, 2016, 8056370. [Google Scholar] [CrossRef]
- Coppens, C.M.; Siripornmongcolchai, T.; Wibrand, K.; Alme, M.N.; Buwalda, B.; de Boer, S.F.; Koolhaas, J.M.; Bramham, C.R. Social Defeat during Adolescence and Adulthood Differentially Induce BDNF-Regulated Immediate Early Genes. Front. Behav. Neurosci. 2011, 5, 72. [Google Scholar] [CrossRef]
- Li, Y.; Ji, Y.J.; Jiang, H.; Liu, D.X.; Zhang, Q.; Fan, S.J.; Pan, F. Effects of unpredictable chronic stress on behavior and brain-derived neurotrophic factor expression in CA3 subfield and dentate gyrus of the hippocampus in different aged rats. Chin. Med. J. 2009, 122, 1564–1569. [Google Scholar]
- Dong, B.E.; Xue, Y.; Sakata, K. The effect of enriched environment across ages: A study of anhedonia and BDNF gene induction. Genes. Brain. Behav. 2018, 17, e12485. [Google Scholar] [CrossRef] [PubMed]
- Spina, E.; Scordo, M.G. Clinically significant drug interactions with antidepressants in the elderly. Drugs Aging 2002, 19, 299–320. [Google Scholar] [CrossRef] [PubMed]
- Calati, R.; Salvina Signorelli, M.; Balestri, M.; Marsano, A.; De Ronchi, D.; Aguglia, E.; Serretti, A. Antidepressants in elderly: Metaregression of double-blind, randomized clinical trials. J. Affect. Disord. 2013, 147, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bjørnebekk, A.; Mathé, A.A.; Gruber, S.H.M.; Brené, S. Housing conditions modulate escitalopram effects on antidepressive-like behaviour and brain neurochemistry. Int. J. Neuropsychopharmacol. 2008, 11, 1135–1147. [Google Scholar] [CrossRef] [PubMed]
- Kok, R.M.; Van Baarsen, C.; Nolen, W.A.; Heeren, T.J. Early response as predictor of final remission in elderly depressed patients. Int. J. Geriatr. Psychiatry 2009, 24, 1299–1303. [Google Scholar] [CrossRef]
- De Berardis, D.; Marini, S.; Serroni, N.; Rapini, G.; Iasevoli, F.; Valchera, A.; Signorelli, M.; Aguglia, E.; Perna, G.; Salone, A.; et al. S-Adenosyl-L-Methionine augmentation in patients with stage II treatment-resistant major depressive disorder: An open label, fixed dose, single-blind study. Sci. World J. 2013, 2013, 204649. [Google Scholar] [CrossRef]
- Sarris, J.; Byrne, G.J.; Bousman, C.; Stough, C.; Murphy, J.; MacDonald, P.; Adams, L.; Nazareth, S.; Oliver, G.; Cribb, L.; et al. Adjunctive S-adenosylmethionine (SAMe) in treating non-remittent major depressive disorder: An 8-week double-blind, randomized, controlled trial. Eur. Neuropsychopharmacol. 2018, 28, 1126–1136. [Google Scholar] [CrossRef]
- Nowak, G.; Szewczyk, B.; Wieronska, J.M.; Branski, P.; Palucha, A.; Pilc, A.; Sadlik, K.; Piekoszewski, W. Antidepressant-like effects of acute and chronic treatment with zinc in forced swim test and olfactory bulbectomy model in rats. Brain Res. Bull. 2003, 61, 159–164. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cubillos, S.; Engmann, O.; Brancato, A. BDNF as a Mediator of Antidepressant Response: Recent Advances and Lifestyle Interactions. Int. J. Mol. Sci. 2022, 23, 14445. https://doi.org/10.3390/ijms232214445
Cubillos S, Engmann O, Brancato A. BDNF as a Mediator of Antidepressant Response: Recent Advances and Lifestyle Interactions. International Journal of Molecular Sciences. 2022; 23(22):14445. https://doi.org/10.3390/ijms232214445
Chicago/Turabian StyleCubillos, Susana, Olivia Engmann, and Anna Brancato. 2022. "BDNF as a Mediator of Antidepressant Response: Recent Advances and Lifestyle Interactions" International Journal of Molecular Sciences 23, no. 22: 14445. https://doi.org/10.3390/ijms232214445
APA StyleCubillos, S., Engmann, O., & Brancato, A. (2022). BDNF as a Mediator of Antidepressant Response: Recent Advances and Lifestyle Interactions. International Journal of Molecular Sciences, 23(22), 14445. https://doi.org/10.3390/ijms232214445