


Evaluation of Cisplatin-Induced Pathology in the Larval Zebrafish Lateral Line


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
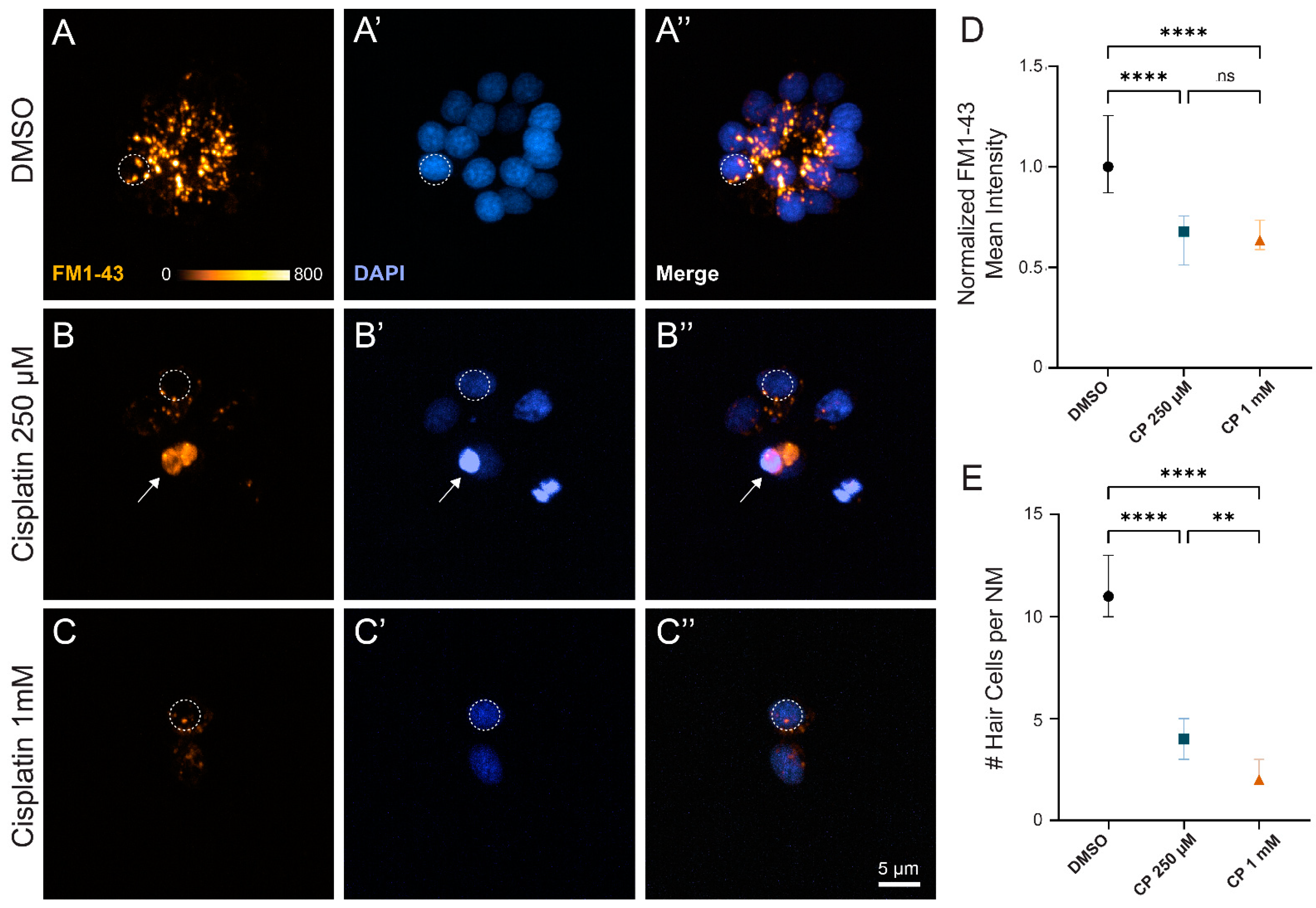
2. Results
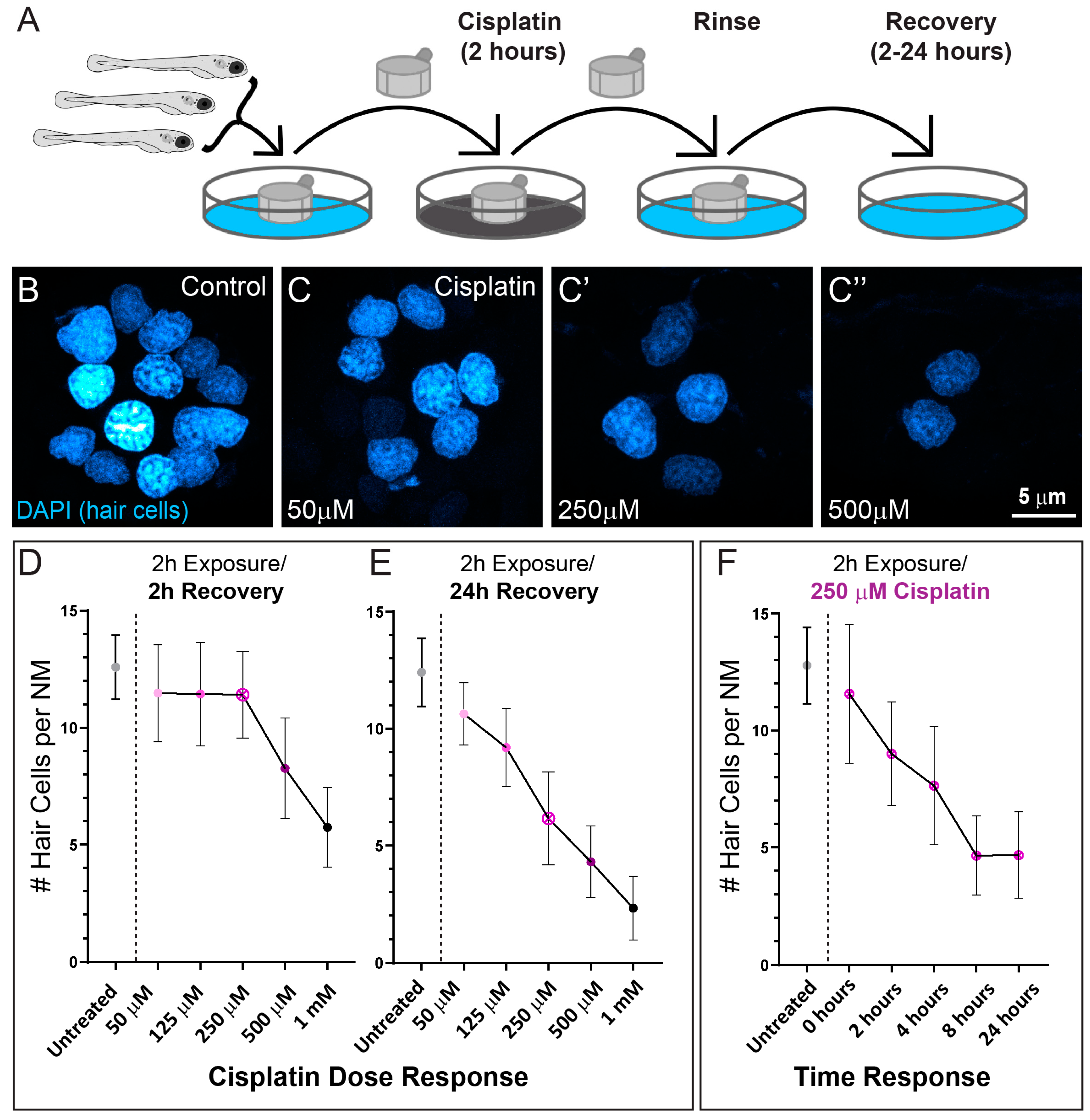
2.1. Cisplatin Exposure Initiates Progressive Lateral Line Hair Cell Loss
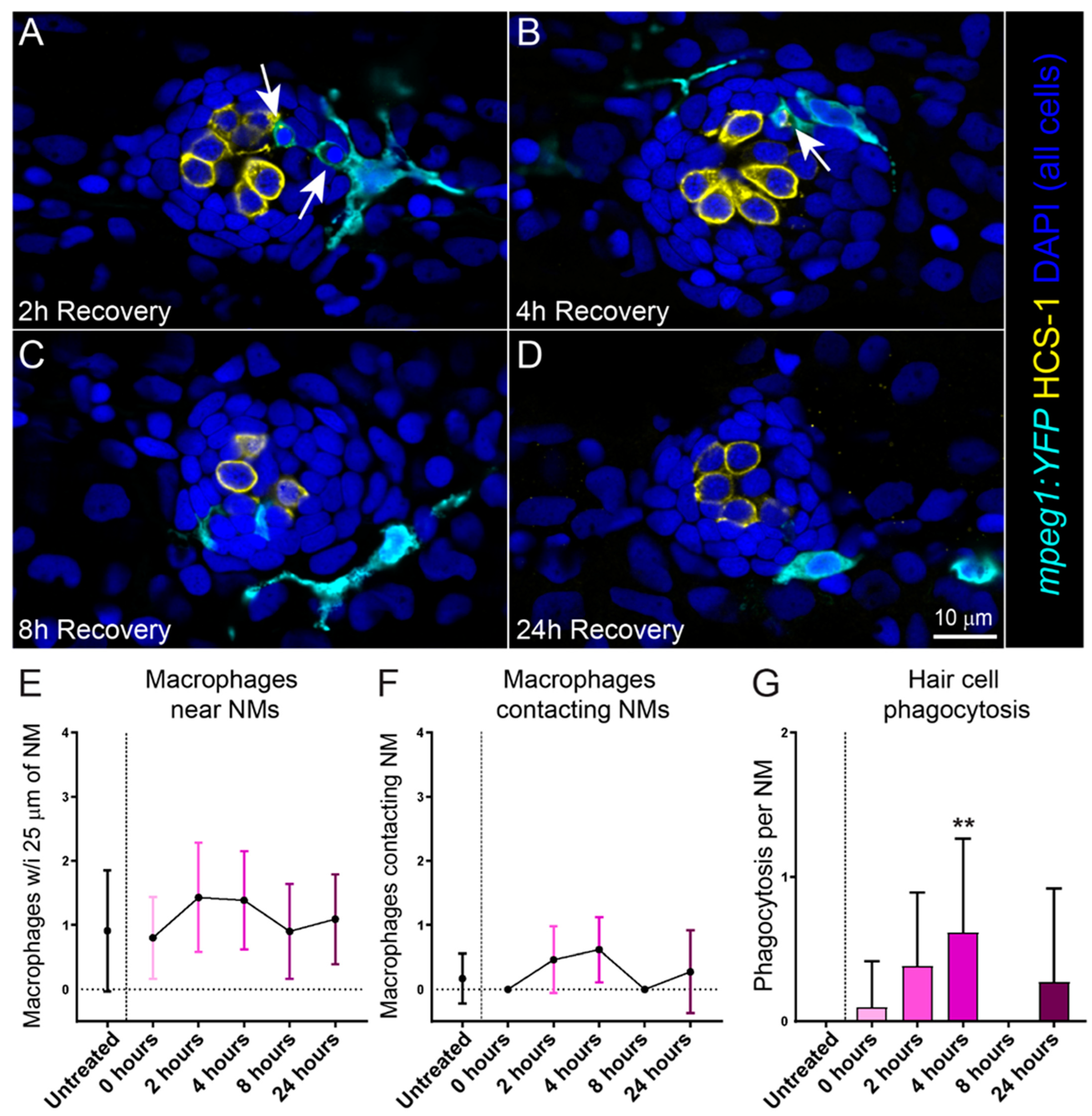
2.2. Cisplatin Treatment Induces an Inflammatory Response That Corresponds with the Timing of Neuromast Hair Cell Death
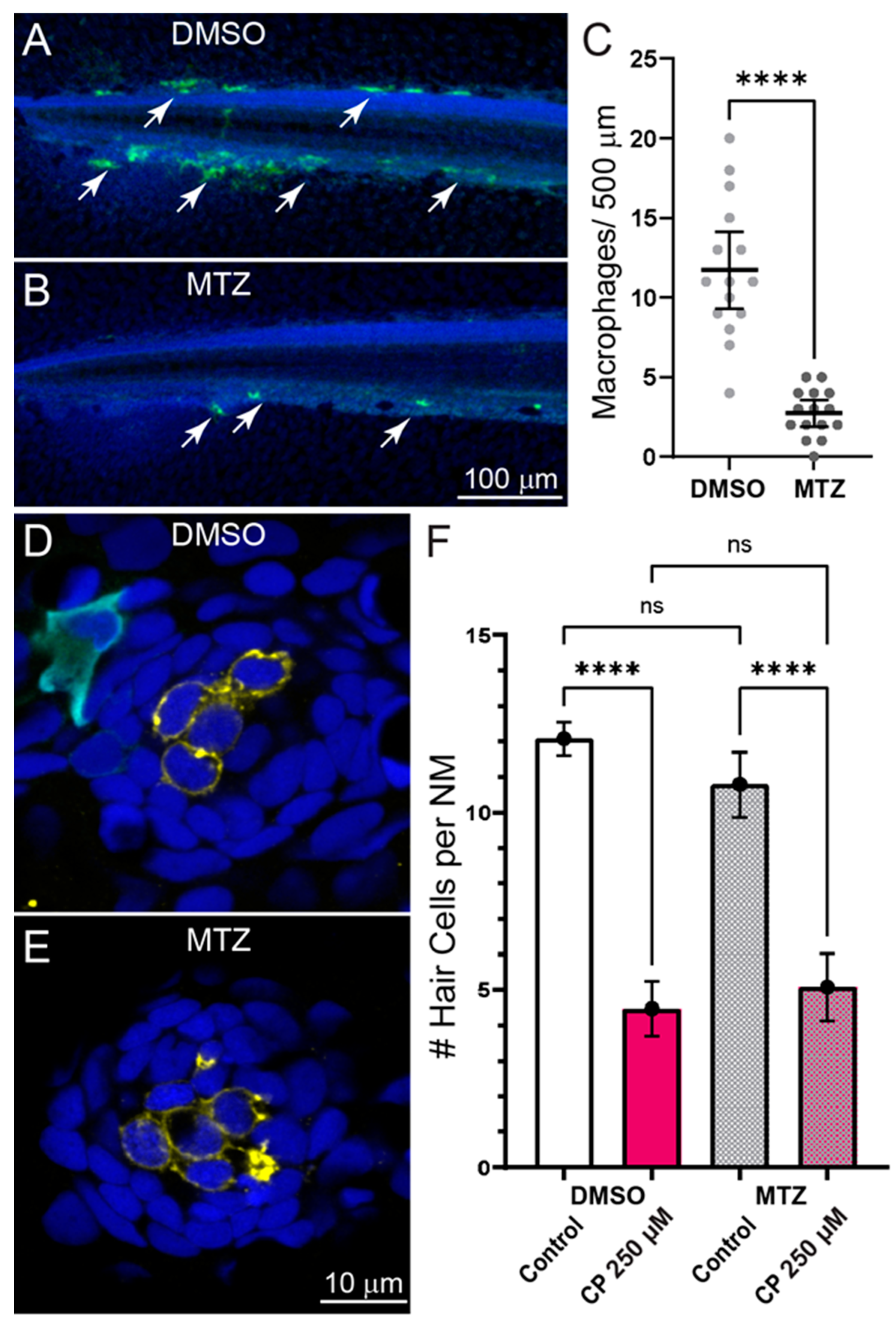
2.3. Inflammation Does Not Enhance Cisplatin Ototoxicity
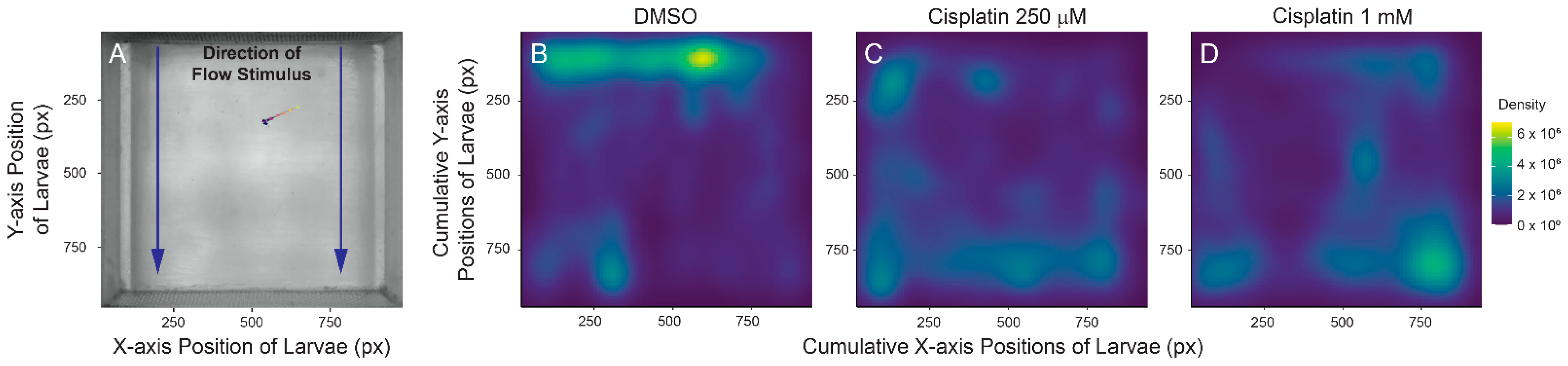
2.4. Cisplatin Treatment Impacts Rheotaxis Behavior and Residual Hair Cell Function
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Zebrafish Husbandry and Lines
4.3. Larval Zebrafish Exposure to Cisplatin
4.4. Selective Depletion of Macrophages
4.5. Immunohistochemical Labeling
4.6. Primary Antibodies
4.7. Confocal Imaging of Fixed Samples
4.8. Confocal Image Processing and Analysis
4.9. Rheotaxis Behavioral Assay and Analysis
4.10. Live Hair Cell Labeling, Imaging, and Analysis
4.11. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rosenberg, B.; Van Camp, L. The successful regression of large solid sarcoma 180 tumors by platinum compounds. Cancer Res. 1970, 30, 1799–1802. [Google Scholar] [PubMed]
- Kelland, L. The resurgence of platinum-based cancer chemotherapy. Nat. Rev. Cancer 2007, 7, 573–584. [Google Scholar] [CrossRef] [PubMed]
- Paken, J.; Govender, C.D.; Pillay, M.; Sewram, V. Cisplatin-Associated Ototoxicity: A Review for the Health Professional. J. Toxicol. 2016, 2016, 1809394. [Google Scholar] [CrossRef] [PubMed]
- Frisina, R.D.; Wheeler, H.; Fossa, S.D.; Kerns, S.L.; Fung, C.; Sesso, H.D.; Monahan, P.O.; Feldman, D.; Hamilton, R.; Vaughn, D.J.; et al. Comprehensive Audiometric Analysis of Hearing Impairment and Tinnitus After Cisplatin-Based Chemotherapy in Survivors of Adult-Onset Cancer. J. Clin. Oncol. 2016, 34, 2712–2720. [Google Scholar] [CrossRef] [PubMed]
- Brock, P.R.; Knight, K.R.; Freyer, D.R.; Campbell, K.C.; Steyger, P.S.; Blakley, B.W.; Rassekh, S.R.; Chang, K.W.; Fligor, B.J.; Rajput, K.; et al. Platinum-induced ototoxicity in children: A consensus review on mechanisms, predisposition, and protection, including a new International Society of Pediatric Oncology Boston ototoxicity scale. J. Clin. Oncol. 2012, 30, 2408–2417. [Google Scholar] [CrossRef]
- Rybak, L.P.; Whitworth, C.A.; Mukherjea, D.; Ramkumar, V. Mechanisms of cisplatin-induced ototoxicity and prevention. Hear. Res. 2007, 226, 157–167. [Google Scholar] [CrossRef]
- Casares, C.; Ramírez-Camacho, R.; Trinidad, A.; Roldán, A.; Jorge, E.; García-Berrocal, J.R. Reactive oxygen species in apoptosis induced by cisplatin: Review of physiopathological mechanisms in animal models. Eur. Arch. Otorhinolaryngol. 2012, 269, 2455–2459. [Google Scholar] [CrossRef]
- Corwin, J.T.; Warchol, M.E. Auditory hair cells: Structure, function, development, and regeneration. Annu. Rev. Neurosci. 1991, 14, 301–333. [Google Scholar] [CrossRef]
- Thomas, A.J.; Wu, P.; Raible, D.W.; Rubel, E.W.; Rubel, E.W.; Ou, H.C. Identification of small molecule inhibitors of cisplatin-induced hair cell death: Results of a 10,000 compound screen in the zebrafish lateral line. Otol. Neurotol. Off. Publ. Am. Otol. Soc. Am. Neurotol. Soc. Eur. Acad. Otol. Neurotol. 2015, 36, 519–525. [Google Scholar] [CrossRef]
- Wertman, J.N.; Melong, N.; Stoyek, M.R.; Piccolo, O.; Langley, S.; Orr, B.; Steele, S.L.; Razaghi, B.; Berman, J.N. The identification of dual protective agents against cisplatin-induced oto- and nephrotoxicity using the zebrafish model. eLife 2020, 9, e56235. [Google Scholar] [CrossRef]
- Ou, H.C.; Raible, D.W.; Rubel, E.W. Cisplatin-induced hair cell loss in zebrafish (Danio rerio) lateral line. Hear. Res. 2007, 233, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Zallocchi, M.; Hati, S.; Xu, Z.; Hausman, W.; Liu, H.; He, D.Z.; Zuo, J. Characterization of quinoxaline derivatives for protection against iatrogenically induced hearing loss. JCI Insight 2021, 6, e141561. [Google Scholar] [CrossRef] [PubMed]
- Coffin, A.B.; Williamson, K.L.; Mamiya, A.; Raible, D.W.; Rubel, E.W. Profiling drug-induced cell death pathways in the zebrafish lateral line. Apoptosis 2013, 18, 393–408. [Google Scholar] [CrossRef] [PubMed]
- Coffin, A.B.; Kelley, M.; Manley, G.A.; Popper, A.N. Evolution of sensory hair cells. In Evolution of the Auditory System; Manley, G.A., Fay, R.R., Popper, A.N., Eds.; Springer: New York, NY, USA, 2004. [Google Scholar]
- Ton, C.; Parng, C. The use of zebrafish for assessing ototoxic and otoprotective agents. Hear Res 2005, 208, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Nicolson, T. The genetics of hair-cell function in zebrafish. J. Neurogenet. 2017, 31, 102–112. [Google Scholar] [CrossRef]
- Cassar, S.; Adatto, I.; Freeman, J.L.; Gamse, J.T.; Iturria, I.; Lawrence, C.; Muriana, A.; Peterson, R.T.; Van Cruchten, S.; Zon, L.I. Use of Zebrafish in Drug Discovery Toxicology. Chem. Res. Toxicol. 2020, 33, 95–118. [Google Scholar] [CrossRef]
- Domarecka, E.; Skarzynska, M.; Szczepek, A.J.; Hatzopoulos, S. Use of zebrafish larvae lateral line to study protection against cisplatin-induced ototoxicity: A scoping review. Int. J. Immunopathol. Pharmacol. 2020, 34, 2058738420959554. [Google Scholar] [CrossRef]
- Warchol, M.E. Interactions between Macrophages and the Sensory Cells of the Inner Ear. Cold Spring Harb. Perspect. Med. 2019, 9, a033555. [Google Scholar] [CrossRef]
- Holmgren, M.; Ravicz, M.E.; Hancock, K.E.; Strelkova, O.; Kallogjeri, D.; Indzhykulian, A.A.; Warchol, M.E.; Sheets, L. Mechanical overstimulation causes acute injury and synapse loss followed by fast recovery in lateral-line neuromasts of larval zebrafish. eLife 2021, 10, e69264. [Google Scholar] [CrossRef]
- Warchol, M.E.; Schrader, A.; Sheets, L. Macrophages Respond Rapidly to Ototoxic Injury of Lateral Line Hair Cells but Are Not Required for Hair Cell Regeneration. Front. Cell Neurosci. 2021, 14, 613246. [Google Scholar] [CrossRef]
- Hirose, K.; Rutherford, M.A.; Warchol, M.E. Two cell populations participate in clearance of damaged hair cells from the sensory epithelia of the inner ear. Hear. Res. 2017, 352, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, Y.; Fu, X.; Wang, P.; Wang, Q.; Meng, W.; Wang, T.; Yang, J.; Chai, R. The Detrimental and Beneficial Functions of Macrophages After Cochlear Injury. Front. Cell Dev. Biol. 2021, 9, 631904. [Google Scholar] [CrossRef] [PubMed]
- Curado, S.; Stainier, D.Y.; Anderson, R.M. Nitroreductase-mediated cell/tissue ablation in zebrafish: A spatially and temporally controlled ablation method with applications in developmental and regeneration studies. Nat. Protoc. 2008, 3, 948–954. [Google Scholar] [CrossRef] [PubMed]
- Niihori, M.; Platto, T.; Igarashi, S.; Hurbon, A.; Dunn, A.M.; Tran, P.; Tran, H.; Mudery, J.A.; Slepian, M.J.; Jacob, A. Zebrafish swimming behavior as a biomarker for ototoxicity-induced hair cell damage: A high-throughput drug development platform targeting hearing loss. Transl. Res. 2015, 166, 440–450. [Google Scholar] [CrossRef] [PubMed]
- Newton, K.C.; Kacev, D.; Nilsson, S.R.; Golden, S.A.; Sheets, L. Lateral Line Ablation by Toxins Results in Distinct Rheotaxis Profiles in Fish. bioRxiv 2021. bioRxiv: 2021.11.15.468723. [Google Scholar]
- Gale, J.E.; Marcotti, W.; Kennedy, H.J.; Kros, C.J.; Richardson, G.P. FM1-43 dye behaves as a permeant blocker of the hair-cell mechanotransducer channel. J. Neurosci. Off. J. Soc. Neurosci. 2001, 21, 7013–7025. [Google Scholar] [CrossRef]
- Toro, C.; Trapani, J.G.; Pacentine, I.; Maeda, R.; Sheets, L.; Mo, W.; Nicolson, T. Dopamine Modulates the Activity of Sensory Hair Cells. J. Neurosci. Off. J. Soc. Neurosci. 2015, 35, 16494–16503. [Google Scholar] [CrossRef] [PubMed]
- Pickett, S.B.; Raible, D.W. Water Waves to Sound Waves: Using Zebrafish to Explore Hair Cell Biology. J. Assoc. Res. Otolaryngol. 2019, 20, 1–19. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, S.; Wong, H.-T.C.; He, X.J.; Beirl, A.; Petralia, R.S.; Wang, Y.-X.; Kindt, K.S. Synaptically silent sensory hair cells in zebrafish are recruited after damage. Nat. Commun. 2018, 9, 1388. [Google Scholar] [CrossRef]
- Lee, D.S.; Schrader, A.; Warchol, M.; Sheets, L. Cisplatin exposure acutely disrupts mitochondrial bioenergetics in the zebrafish lateral-line organ. Hear. Res. 2022, 108513. [Google Scholar] [CrossRef]
- Vargo, J.W.; Walker, S.N.; Gopal, S.R.; Deshmukh, A.R.; McDermott, B.M., Jr.; Alagramam, K.N.; Stepanyan, R. Inhibition of Mitochondrial Division Attenuates Cisplatin-Induced Toxicity in the Neuromast Hair Cells. Front. Cell Neurosci. 2017, 11, 393. [Google Scholar] [CrossRef]
- Holmgren, M.; Sheets, L. Influence of Mpv17 on Hair-Cell Mitochondrial Homeostasis, Synapse Integrity, and Vulnerability to Damage in the Zebrafish Lateral Line. Front. Cell Neurosci. 2021, 15, 693375. [Google Scholar] [CrossRef] [PubMed]
- Roca, F.J.; Ramakrishnan, L. TNF dually mediates resistance and susceptibility to mycobacteria via mitochondrial reactive oxygen species. Cell 2013, 153, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Ellett, F.; Pase, L.; Hayman, J.W.; Andrianopoulos, A.; Lieschke, G.J. mpeg1 promoter transgenes direct macrophage-lineage expression in zebrafish. Blood 2011, 117, e49–e56. [Google Scholar] [CrossRef] [PubMed]
- Uribe, P.M.; Mueller, M.A.; Gleichman, J.S.; Kramer, M.D.; Wang, Q.; Sibrian-Vazquez, M.; Strongin, R.M.; Steyger, P.S.; Cotanche, U.A.; Matsui, J.I. Dimethyl sulfoxide (DMSO) exacerbates cisplatin-induced sensory hair cell death in zebrafish (Danio rerio). PLoS ONE 2013, 8, e55359. [Google Scholar] [CrossRef]
- Saettele, A.L.; Wong, H.-T.C.; Kindt, K.S.; Warchol, M.E.; Sheets, L. Prolonged Dexamethasone Exposure Enhances Zebrafish Lateral-Line Regeneration but Disrupts Mitochondrial Homeostasis and Hair Cell Function. J. Assoc. Res. Otolaryngol. 2022. [Google Scholar] [CrossRef]
- Kindt, K.S.; Sheets, L. Transmission Disrupted: Modeling Auditory Synaptopathy in Zebrafish. Front. Cell Dev. Biol. 2018, 6, 114. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, D.S.; Schrader, A.; Bell, E.; Warchol, M.E.; Sheets, L. Evaluation of Cisplatin-Induced Pathology in the Larval Zebrafish Lateral Line. Int. J. Mol. Sci. 2022, 23, 14302. https://doi.org/10.3390/ijms232214302
Lee DS, Schrader A, Bell E, Warchol ME, Sheets L. Evaluation of Cisplatin-Induced Pathology in the Larval Zebrafish Lateral Line. International Journal of Molecular Sciences. 2022; 23(22):14302. https://doi.org/10.3390/ijms232214302
Chicago/Turabian StyleLee, David S., Angela Schrader, Emily Bell, Mark E. Warchol, and Lavinia Sheets. 2022. "Evaluation of Cisplatin-Induced Pathology in the Larval Zebrafish Lateral Line" International Journal of Molecular Sciences 23, no. 22: 14302. https://doi.org/10.3390/ijms232214302
APA StyleLee, D. S., Schrader, A., Bell, E., Warchol, M. E., & Sheets, L. (2022). Evaluation of Cisplatin-Induced Pathology in the Larval Zebrafish Lateral Line. International Journal of Molecular Sciences, 23(22), 14302. https://doi.org/10.3390/ijms232214302