
Nitro Dihydrocapsaicin, a Non-Pungent Capsaicin Analogue, Inhibits Cellular Senescence of Lens Epithelial Cells via Upregulation of SIRT1

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
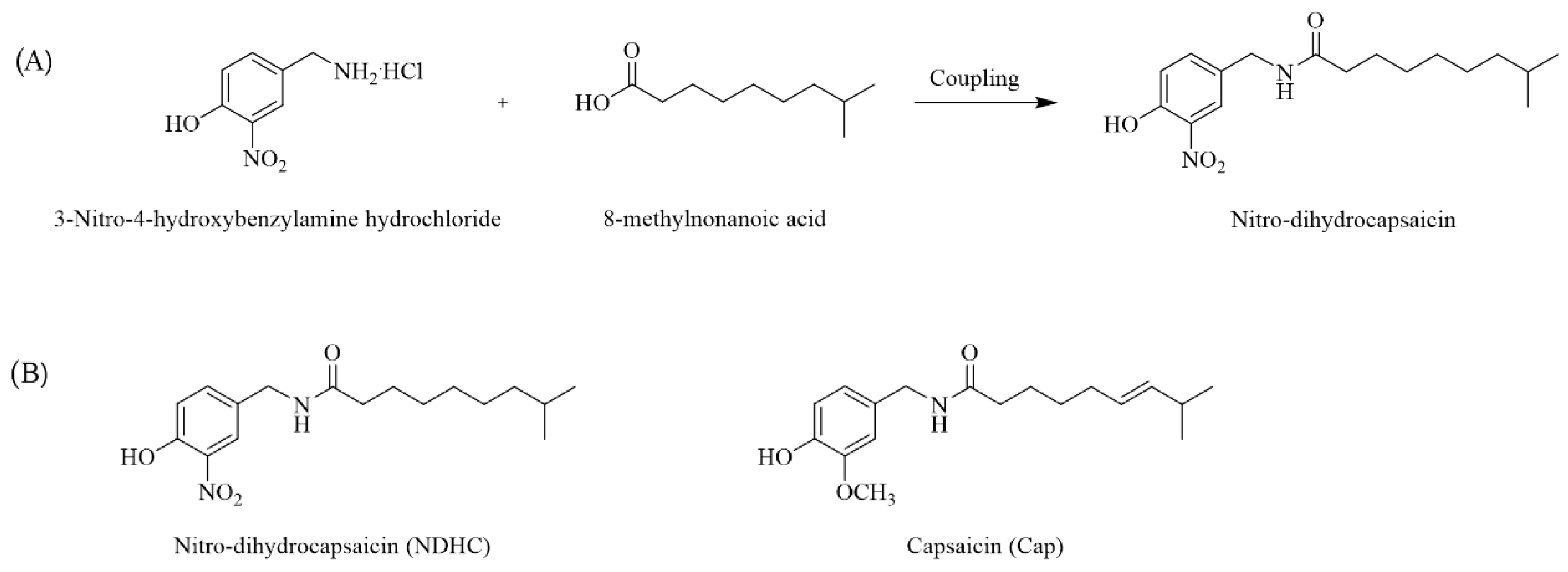
2.1. Nitro Dihydrocapsaicin (NDHC) Synthesis
2.2. Cell Viability Assay
2.3. Long-Term HG Exposure Did Not Impair the Viability of HLE Cells
2.4. NDHC Sustained the Viability of HLE by Alleviating the Oxidative Stress Response under High-Glucose Conditions
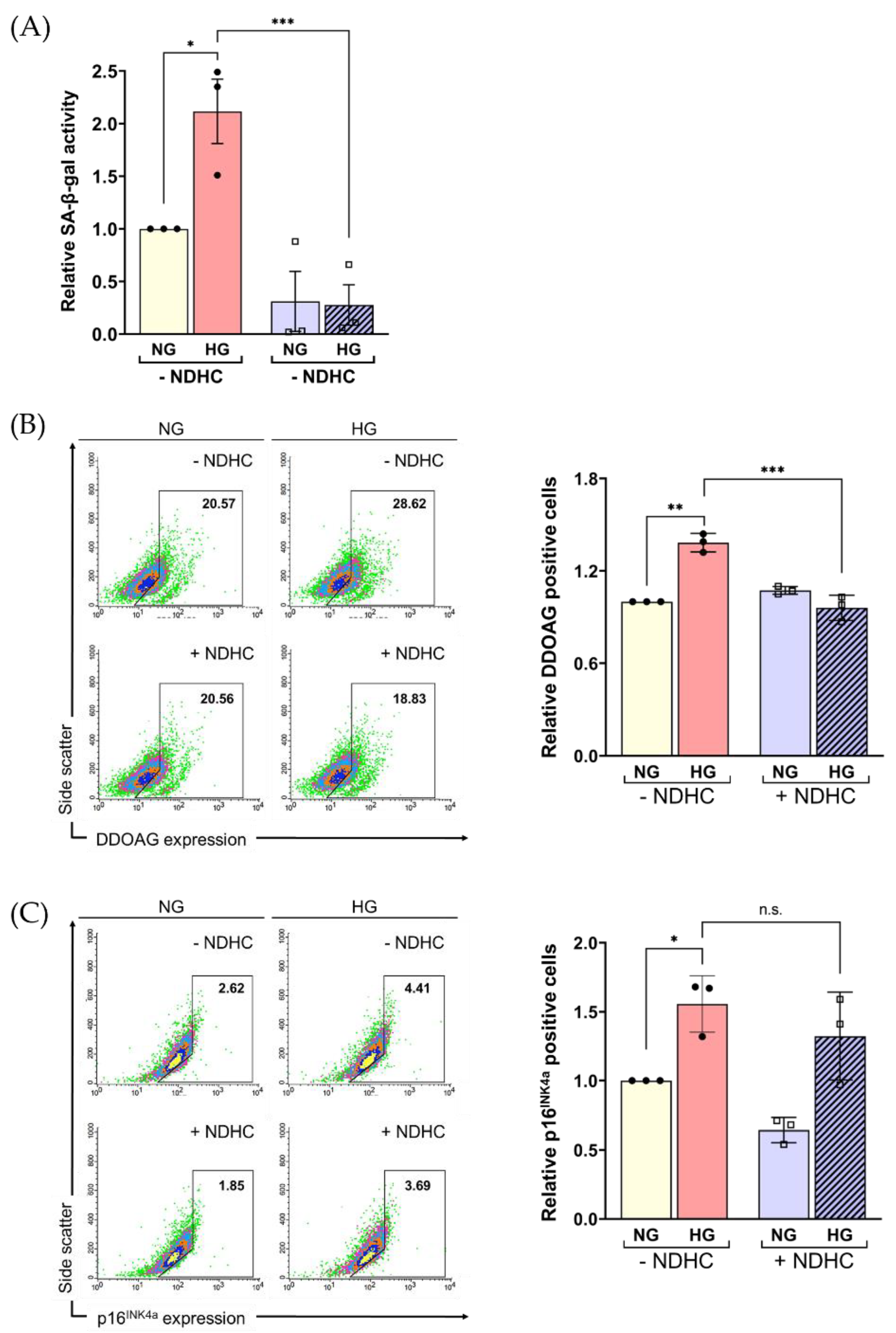
2.5. Enhancement of Cellular ROS Production and Senescence under HG Condition
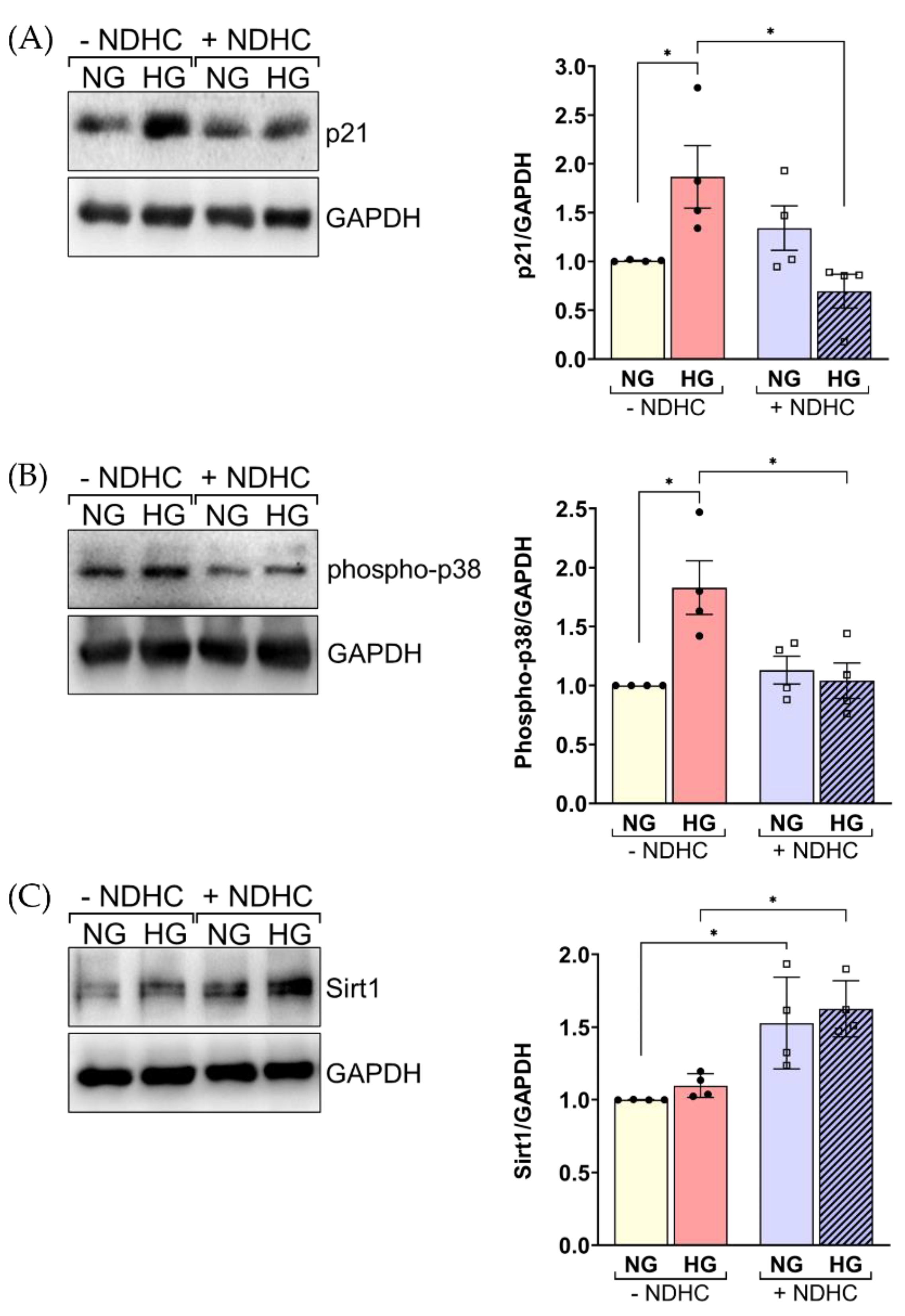
2.6. NDHC Reverses HLE Senescence by Upregulating SIRT1 and Inhibiting p21 and Phospho-p38 Expression
3. Materials and Methods
3.1. Reagents and Antibodies
3.2. A Nitro Dihydrocapsaicin Preparation
3.3. Cell Culture
3.4. Cell Viability Assay
3.5. ROS Production
3.6. SA-β gal Activity Assay
3.7. Western Blot
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haddad, N.M.N.; Sun, J.K.; Abujaber, S.; Schlossman, D.K.; Silva, P.S. Cataract Surgery and its Complications in Diabetic Patients. Semin. Ophthalmol. 2014, 29, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-Y.; Wu, T.-T.; Ho, C.-Y.; Yeh, T.-C.; Sun, G.-C.; Tseng, C.-J.; Cheng, P.-W. Blocking of SGLT2 to Eliminate NADPH-Induced Oxidative Stress in Lenses of Animals with Fructose-Induced Diabetes Mellitus. Int. J. Mol. Sci. 2022, 23, 7142. [Google Scholar] [CrossRef] [PubMed]
- Kiziltoprak, H.; Tekin, K.; Inanc, M.; Goker, Y.S. Cataract in diabetes mellitus. World J. Diabetes 2019, 10, 140–153. [Google Scholar] [CrossRef]
- Swarup, A.; Bell, B.A.; Du, J.; Han, J.Y.S.; Soto, J.; Abel, E.D.; Bravo-Nuevo, A.; FitzGerald, P.G.; Peachey, N.S.; Philp, N.J. Deletion of GLUT1 in mouse lens epithelium leads to cataract formation. Exp. Eye Res. 2018, 172, 45–53. [Google Scholar] [CrossRef]
- Jain, A.K.; Lim, G.; Langford, M.; Jain, S.K. Effect of high-glucose levels on protein oxidation in cultured lens cells, and in crystalline and albumin solution and its inhibition by vitamin B6 and N-acetylcysteine: Its possible relevance to cataract formation in diabetes. Free Radic. Biol. Med. 2002, 33, 1615–1621. [Google Scholar] [CrossRef]
- Albert-Garay, J.S.; Riesgo-Escovar, J.R.; Salceda, R. High glucose concentrations induce oxidative stress by inhibiting Nrf2 expression in rat Müller retinal cells in vitro. Sci. Rep. 2022, 12, 1261. [Google Scholar] [CrossRef]
- Chen, P.; Yao, Z.; He, Z. Resveratrol protects against high glucose-induced oxidative damage in human lens epithelial cells by activating autophagy. Exp. Med. 2021, 21, 440. [Google Scholar] [CrossRef]
- Truscott, R.J.W.; Friedrich, M.G. Molecular Processes Implicated in Human Age-Related Nuclear Cataract. Investig. Ophthalmol. Vis. Sci. 2019, 60, 5007–5021. [Google Scholar] [CrossRef]
- Hashim, Z.; Zarina, S. Advanced glycation end products in diabetic and non-diabetic human subjects suffering from cataract. Age 2011, 33, 377–384. [Google Scholar] [CrossRef]
- Bron, A.J.; Sparrow, J.; Brown, N.A.P.; Harding, J.J.; Blakytny, R. The lens in diabetes. Eye 1993, 7, 260–275. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Fu, Y.; Wang, X.; Wu, R.; Su, D.; Zhou, N.; Qi, Y. Metformin protects lens epithelial cells against senescence in a naturally aged mouse model. Cell Death Discov. 2022, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Kumari, R.; Jat, P. Mechanisms of Cellular Senescence: Cell Cycle Arrest and Senescence Associated Secretory Phenotype. Front. Cell Dev. Biol. 2021, 9, 645593. [Google Scholar] [CrossRef]
- Wiley, C.D.; Campisi, J. The metabolic roots of senescence: Mechanisms and opportunities for intervention. Nat. Metab. 2021, 3, 1290–1301. [Google Scholar] [CrossRef]
- Bertelli, P.M.; Pedrini, E.; Hughes, D.; McDonnell, S.; Pathak, V.; Peixoto, E.; Guduric-Fuchs, J.; Stitt, A.W.; Medina, R.J. Long term high glucose exposure induces premature senescence in retinal endothelial cells. Front. Physiol. 2022, 13, 1661. [Google Scholar] [CrossRef]
- Piwkowska, A.; Rogacka, D.; Audzeyenka, I.; Jankowski, M.; Angielski, S. High glucose concentration affects the oxidant-antioxidant balance in cultured mouse podocytes. J. Cell. Biochem. 2011, 112, 1661–1672. [Google Scholar] [CrossRef]
- Yao, Q.; Zhou, Y.; Yang, Y.; Cai, L.; Xu, L.; Han, X.; Guo, Y.; Li, P.A. Activation of Sirtuin1 by lyceum barbarum polysaccharides in protection against diabetic cataract. J. Ethnopharmacol. 2020, 261, 113165. [Google Scholar] [CrossRef] [PubMed]
- Mimura, T.; Kaji, Y.; Noma, H.; Funatsu, H.; Okamoto, S. The role of SIRT1 in ocular aging. Exp. Eye Res. 2013, 116, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Adetunji, T.L.; Olawale, F.; Olisah, C.; Adetunji, A.E.; Aremu, A.O. Capsaicin: A Two-Decade Systematic Review of Global Research Output and Recent Advances Against Human Cancer. Front. Oncol. 2022, 12, 908487. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Escogido Mde, L.; Gonzalez-Mondragon, E.G.; Vazquez-Tzompantzi, E. Chemical and pharmacological aspects of capsaicin. Molecules 2011, 16, 1253–1270. [Google Scholar] [CrossRef] [PubMed]
- Delamere, N.A.; Shahidullah, M. Ion Transport Regulation by TRPV4 and TRPV1 in Lens and Ciliary Epithelium. Front. Physiol. 2021, 12, 834916. [Google Scholar] [CrossRef]
- Nakazawa, Y.; Donaldson, P.J.; Petrova, R.S. Verification and spatial mapping of TRPV1 and TRPV4 expression in the embryonic and adult mouse lens. Exp. Eye Res. 2019, 186, 107707. [Google Scholar] [CrossRef] [PubMed]
- Basith, S.; Cui, M.; Hong, S.; Choi, S. Harnessing the therapeutic potential of capsaicin and its analogues in pain and other diseases. Molecules. 2016, 21, 966. [Google Scholar] [CrossRef] [PubMed]
- Chapa-Oliver, A.M.; Mejía-Teniente, L. Capsaicin: From Plants to a Cancer-Suppressing Agent. Molecules 2016, 21, 931. [Google Scholar] [CrossRef] [PubMed]
- Krishnatreyya, H.; Hazarika, H.; Saha, A.; Chattopadhyay, P. Fundamental pharmacological expressions on ocular exposure to capsaicin, the principal constituent in pepper sprays. Sci. Rep. 2018, 8, 12153. [Google Scholar] [CrossRef]
- Baron, R. Capsaicin and nociception: From basic mechanisms to novel drugs. Lancet 2000, 356, 785–787. [Google Scholar] [CrossRef]
- Laorob, T.; Bunyapraphatsara, N.; Waranuch, N.; Pongcharoen, S.; Punyain, W.; Chancharunee, S.; Sakchaisri, K.; Pratuangdejkul, J.; Chongruchiroj, S.; Kielar, F.; et al. Enhancement of Lipolysis in 3T3-L1 Adipocytes by Nitroarene Capsaicinoid Analogs. Nat. Prod. Commun. 2021, 16, 1934578X2098794. [Google Scholar] [CrossRef]
- Malewicz, N.M.; Rattray, Z.; Oeck, S.; Jung, S.; Escamilla-Rivera, V.; Chen, Z.; Tang, X.; Zhou, J.; LaMotte, R.H. Topical Capsaicin in Poly(lactic-co-glycolic)acid (PLGA) Nanoparticles Decreases Acute Itch and Heat Pain. Int. J. Mol. Sci. 2022, 23, 5275. [Google Scholar] [CrossRef]
- Moya, C.; Marquez-Aguirre, A. The Prospective Antiobesity Effect of Capsaicin Synthetic Analogs: A Matter of Weight. Med. Chem. 2016, 6, 365–371. [Google Scholar] [CrossRef]
- Wang, J.-L. A Study of Pungency of Capsaicinoid as Affected by Their Molecular Structure Alteration. Pharmacol. Pharm. 2011, 2, 109–115. [Google Scholar] [CrossRef]
- Tsukura, Y.; Mori, M.; Hirotani, Y.; Ikeda, K.; Amano, F.; Kato, R.; Ijiri, Y.; Tanaka, K. Effects of Capsaicin on Cellular Damage and Monolayer Permeability in Human Intestinal Caco-2 Cells. Biol. Pharm. Bull. 2007, 30, 1982–1986. [Google Scholar] [CrossRef]
- Du, S.; Shao, J.; Xie, D.; Zhang, F. Decorin inhibits glucose-induced lens epithelial cell apoptosis via suppressing p22phox-p38 MAPK signaling pathway. PLoS ONE 2020, 15, e0224251. [Google Scholar] [CrossRef]
- Fu, J.; Hu, X. Simvastatin alleviates epithelial-mesenchymal transition and oxidative stress of high glucose-induced lens epithelial cells in vitro by inhibiting RhoA/ROCK signaling. Exp. Med. 2022, 23, 420. [Google Scholar] [CrossRef]
- Weil, B.R.; Abarbanell, A.M.; Herrmann, J.L.; Wang, Y.; Meldrum, D.R. High glucose concentration in cell culture medium does not acutely affect human mesenchymal stem cell growth factor production or proliferation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R1735–R1743. [Google Scholar] [CrossRef] [PubMed]
- Aswamenakul, K.; Klabklai, P.; Pannengpetch, S.; Tawonsawatruk, T.; Isarankura-Na-Ayudhya, C.; Roytrakul, S.; Nantasenamat, C.; Supokawej, A. Proteomic study of in vitro osteogenic differentiation of mesenchymal stem cells in high glucose condition. Mol. Biol. Rep. 2020, 47, 7505–7516. [Google Scholar] [CrossRef]
- Chen, M.; Zheng, H.; Wei, T.; Wang, D.; Xia, H.; Zhao, L.; Ji, J.; Gao, H. High Glucose-Induced PC12 Cell Death by Increasing Glutamate Production and Decreasing Methyl Group Metabolism. BioMed Res. Int. 2016, 2016, 4125731. [Google Scholar] [CrossRef]
- Thongin, S.; Den-udom, T.; Uppakara, K.; Sriwantana, T.; Sibmooh, N.; Laorob, T.; Boonthip, C.; Wichai, U.; Muta, K.; Ketsawatsomkron, P. Beneficial effects of capsaicin and dihydrocapsaicin on endothelial inflammation, nitric oxide production and antioxidant activity. Biomed. Pharmacother. 2022, 154, 113521. [Google Scholar] [CrossRef] [PubMed]
- Debacq-Chainiaux, F.; Erusalimsky, J.D.; Campisi, J.; Toussaint, O. Protocols to detect senescence-associated beta-galactosidase (SA-βgal) activity, a biomarker of senescent cells in culture and in vivo. Nat. Protoc. 2009, 4, 1798–1806. [Google Scholar] [CrossRef] [PubMed]
- de Mera-Rodríguez, J.A.; Álvarez-Hernán, G.; Gañán, Y.; Martín-Partido, G.; Rodríguez-León, J.; Francisco-Morcillo, J. Is Senescence-Associated β-Galactosidase a Reliable in vivo Marker of Cellular Senescence during Embryonic Development? Front. Cell Dev. Biol. 2021, 9, 623175. [Google Scholar] [CrossRef]
- Harney, A.; Meade, T. Molecular imaging of in vivo gene expression. Future Med. Chem. 2010, 2, 503–519. [Google Scholar] [CrossRef]
- Liu, J.-Y.; Souroullas, G.P.; Diekman, B.O.; Krishnamurthy, J.; Hall, B.M.; Sorrentino, J.A.; Parker, J.S.; Sessions, G.A.; Gudkov, A.V.; Sharpless, N.E. Cells exhibiting strong p16INK4a promoter activation in vivo display features of senescence. Proc. Natl. Acad. Sci. USA 2019, 116, 2603–2611. [Google Scholar] [CrossRef]
- Bras, I.D.; Colitz, C.M.; Kusewitt, D.F.; Chandler, H.; Lu, P.; Gemensky-Metzler, A.J.; Wilkie, D.A. Evaluation of advanced glycation end-products in diabetic and inherited canine cataracts. Graefe’s Arch. Clin. Exp. Ophthalmol. 2007, 245, 249–257. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paensuwan, P.; Laorob, T.; Ngoenkam, J.; Wichai, U.; Pongcharoen, S. Nitro Dihydrocapsaicin, a Non-Pungent Capsaicin Analogue, Inhibits Cellular Senescence of Lens Epithelial Cells via Upregulation of SIRT1. Int. J. Mol. Sci. 2022, 23, 13960. https://doi.org/10.3390/ijms232213960
Paensuwan P, Laorob T, Ngoenkam J, Wichai U, Pongcharoen S. Nitro Dihydrocapsaicin, a Non-Pungent Capsaicin Analogue, Inhibits Cellular Senescence of Lens Epithelial Cells via Upregulation of SIRT1. International Journal of Molecular Sciences. 2022; 23(22):13960. https://doi.org/10.3390/ijms232213960
Chicago/Turabian StylePaensuwan, Pussadee, Thanet Laorob, Jatuporn Ngoenkam, Uthai Wichai, and Sutatip Pongcharoen. 2022. "Nitro Dihydrocapsaicin, a Non-Pungent Capsaicin Analogue, Inhibits Cellular Senescence of Lens Epithelial Cells via Upregulation of SIRT1" International Journal of Molecular Sciences 23, no. 22: 13960. https://doi.org/10.3390/ijms232213960
APA StylePaensuwan, P., Laorob, T., Ngoenkam, J., Wichai, U., & Pongcharoen, S. (2022). Nitro Dihydrocapsaicin, a Non-Pungent Capsaicin Analogue, Inhibits Cellular Senescence of Lens Epithelial Cells via Upregulation of SIRT1. International Journal of Molecular Sciences, 23(22), 13960. https://doi.org/10.3390/ijms232213960