
TREML2 Gene Expression and Its Missense Variant rs3747742 Associate with White Matter Hyperintensity Volume and Alzheimer’s Disease-Related Brain Atrophy in the General Population
 , , , , , , , , , ,
, , , , , , , , , ,
Abstract
1. Introduction
2. Results
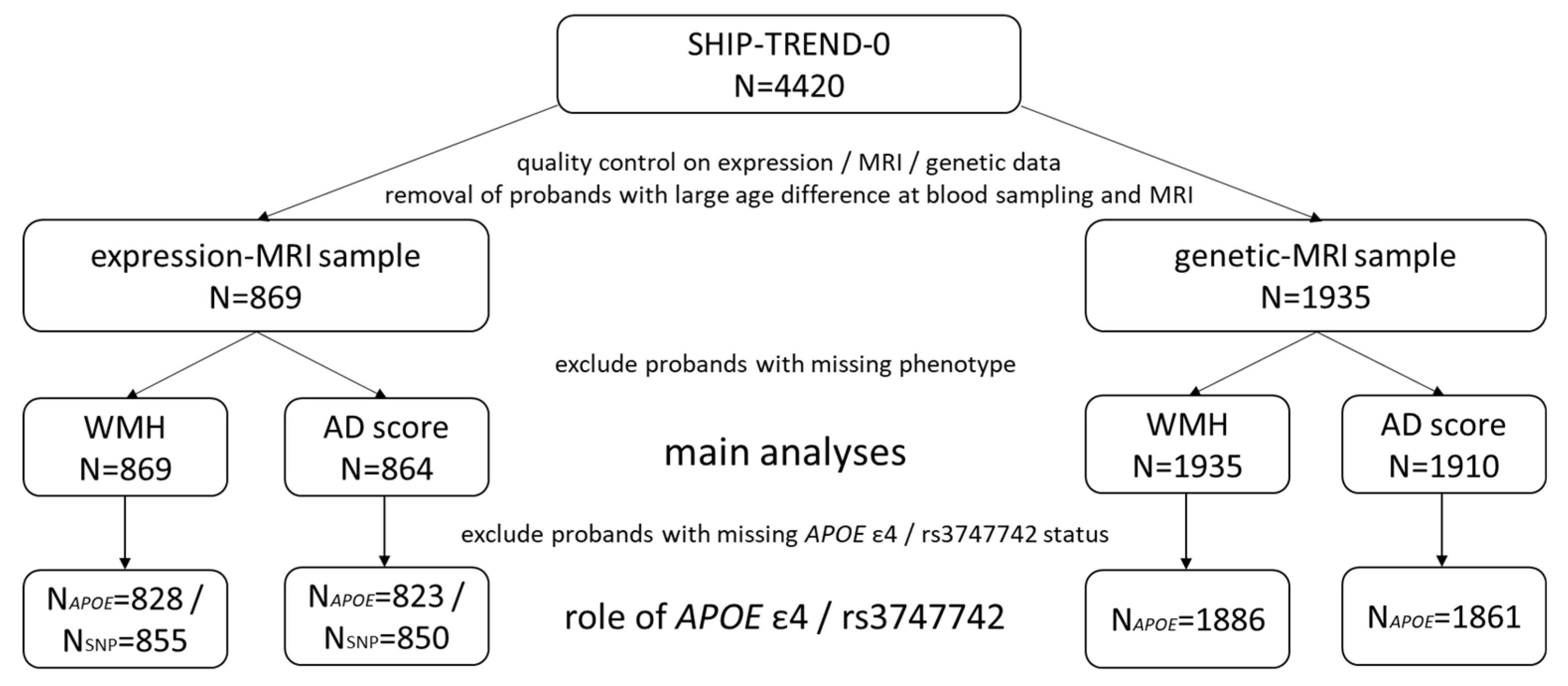
2.1. Study Population
2.2. TREML2 Expression Associates with White Matter Hyperintensity Volume Irrespective of rs3747742 and APOE ε4 Status
2.3. TREML2 Expression Does Not Associate with the Alzheimer’s Disease Score, Neither Directly nor in Interaction with rs3747742 or APOE ε4 Status
2.4. rs3747742 Associates with the Alzheimer’s Disease Score Irrespective of the APOE ε4 Status
2.5. rs3747742 Does Not Associate with White Matter Hyperintensity Volume, Neither Directly nor in Interaction with APOE ε4 Status
2.6. APOE ε4 Status Associates with the Alzheimer’s Disease Score but Not with White Matter Hyperintensity Volume
2.7. rs3747742 and TREML2 Expression
2.8. SNPs in Linkage Disequilibrium with rs3747742
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. MRI Measurements, White Matter Hyperintensity Volume and Alzheimer’s Disease Score
4.3. Genetic Measurements
4.4. Whole-Blood Transcriptome Measurements
4.5. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gauthier, S.; Rosa-Neto, P.; Morais, J.; Webster, C. World Alzheimer Report 2021: Journey through the Diagnosis of Dementia; Alzheimer’s Disease International: London, UK, 2021; Available online: https://www.alzint.org/u/World-Alzheimer-Report-2021.pdf (accessed on 22 July 2022).
- 2022 Alzheimer’s disease facts and figures. Alzheimers Dement. 2022, 18, 700–789. [CrossRef]
- Sattler, C.; Toro, P.; Schönknecht, P.; Schröder, J. Cognitive activity, education and socioeconomic status as preventive factors for mild cognitive impairment and Alzheimer’s disease. Psychiatry Res. 2012, 196, 90–95. [Google Scholar] [CrossRef]
- Uddin, M.S.; Kabir, M.T.; al Mamun, A.; Abdel-Daim, M.M.; Barreto, G.E.; Ashraf, G.M. APOE and Alzheimer’s disease: Evidence mounts that targeting APOE4 may combat Alzheimer’s pathogenesis. Mol. Neurobiol. 2019, 56, 2450–2465. [Google Scholar] [CrossRef]
- Bettcher, B.M.; Tansey, M.G.; Dorothée, G.; Heneka, M.T. Peripheral and central immune system crosstalk in Alzheimer disease—A research prospectus. Nat. Rev. Neurol. 2021, 17, 689–701. [Google Scholar] [CrossRef]
- Prins, N.D.; Scheltens, P. White matter hyperintensities, cognitive impairment and dementia: An update. Nat. Rev. Neurol. 2015, 11, 157–165. [Google Scholar] [CrossRef]
- Habes, M.; Pomponio, R.; Shou, H.; Doshi, J.; Mamourian, E.; Erus, G.; Nasrallah, I.; Launer, L.J.; Rashid, T.; Bilgel, M.; et al. The Brain Chart of Aging: Machine-learning analytics reveals links between brain aging, white matter disease, amyloid burden, and cognition in the iSTAGING consortium of 10,216 harmonized MR scans. Alzheimers Dement. J. Alzheimers Assoc. 2021, 17, 89–102. [Google Scholar] [CrossRef]
- Habes, M.; Sotiras, A.; Erus, G.; Toledo, J.B.; Janowitz, D.; Wolk, D.A.; Shou, H.; Bryan, N.R.; Doshi, J.; Völzke, H.; et al. White matter lesions: Spatial heterogeneity, links to risk factors, cognition, genetics, and atrophy. Neurology 2018, 91, e964–e975. [Google Scholar] [CrossRef]
- Habes, M.; Erus, G.; Toledo, J.B.; Zhang, T.; Bryan, N.; Launer, L.J.; Rosseel, Y.; Janowitz, D.; Doshi, J.; Van der Auwera, S.; et al. White matter hyperintensities and imaging patterns of brain ageing in the general population. Brain 2016, 139, 1164–1179. [Google Scholar] [CrossRef]
- Lin, H.; Satizabal, C.; Xie, Z.; Yang, Q.; Huan, T.; Joehanes, R.; Wen, C.; Munson, P.J.; Beiser, A.; Levy, D.; et al. Whole blood gene expression and white matter hyperintensities. Mol. Neurodegener. 2017, 12, 67. [Google Scholar] [CrossRef]
- Alber, J.; Alladi, S.; Bae, H.; Barton, D.A.; Beckett, L.A.; Bell, J.M.; Berman, S.E.; Biessels, G.J.; Black, S.E.; Bos, I.; et al. White matter hyperintensities in vascular contributions to cognitive impairment and dementia (VCID): Knowledge gaps and opportunities. Alzheimers Dement. Transl. Res. Clin. Interv. 2019, 5, 107–117. [Google Scholar] [CrossRef]
- Moroni, F.; Ammirati, E.; Rocca, M.A.; Filippi, M.; Magnoni, M.; Camici, P.G. Cardiovascular disease and brain health: Focus on white matter hyperintensities. IJC Heart Vasc. 2018, 19, 63–69. [Google Scholar] [CrossRef]
- Hu, H.-Y.; Ou, Y.-N.; Shen, X.-N.; Qu, Y.; Ma, Y.-H.; Wang, Z.-T.; Dong, Q.; Tan, L.; Yu, J.-T. White matter hyperintensities and risks of cognitive impairment and dementia: A systematic review and meta-analysis of 36 prospective studies. Neurosci. Biobehav. Rev. 2021, 120, 16–27. [Google Scholar] [CrossRef]
- Wang, Y.-L.; Chen, W.; Cai, W.-J.; Hu, H.; Xu, W.; Wang, Z.-T.; Cao, X.-P.; Tan, L.; Yu, J.-T. Alzheimer’s Disease Neuroimaging Initiative Associations of white matter hyperintensities with cognitive decline: A longitudinal study. J. Alzheimers Dis. 2020, 73, 759–768. [Google Scholar] [CrossRef]
- d’Arbeloff, T.; Elliott, M.L.; Knodt, A.R.; Melzer, T.R.; Keenan, R.; Ireland, D.; Ramrakha, S.; Poulton, R.; Anderson, T.; Caspi, A.; et al. White matter hyperintensities are common in midlife and already associated with cognitive decline. Brain Commun. 2019, 1, fcz041. [Google Scholar] [CrossRef]
- Moroni, F.; Ammirati, E.; Hainsworth, A.H.; Camici, P.G. Association of white matter hyperintensities and cardiovascular disease: The importance of microcirculatory disease. Circ. Cardiovasc. Imaging 2020, 13, e010460. [Google Scholar] [CrossRef]
- Debette, S.; Beiser, A.; DeCarli, C.; Au, R.; Himali, J.J.; Kelly-Hayes, M.; Romero, J.R.; Kase, C.S.; Wolf, P.A.; Seshadri, S. Association of MRI markers of vascular brain injury with incident stroke, mild cognitive impairment, dementia, and mortality: The Framingham Offspring Study. Stroke 2010, 41, 600–606. [Google Scholar] [CrossRef]
- Jeerakathil, T.; Wolf, P.A.; Beiser, A.; Massaro, J.; Seshadri, S.; D’Agostino, R.B.; DeCarli, C. Stroke risk profile predicts white matter hyperintensity volume: The Framingham Study. Stroke 2004, 35, 1857–1861. [Google Scholar] [CrossRef]
- Hasan, T.F.; Barrett, K.M.; Brott, T.G.; Badi, M.K.; Lesser, E.R.; Hodge, D.O.; Meschia, J.F. Severity of white matter hyperintensities and effects on all-cause mortality in the Mayo Clinic Florida Familial Cerebrovascular Diseases Registry. Mayo Clin. Proc. 2019, 94, 408–416. [Google Scholar] [CrossRef]
- Bokura, H.; Kobayashi, S.; Yamaguchi, S.; Iijima, K.; Nagai, A.; Toyoda, G.; Oguro, H.; Takahashi, K. Silent brain infarction and subcortical white matter lesions increase the risk of stroke and mortality: A prospective cohort study. J. Stroke Cerebrovasc. Dis. 2006, 15, 57–63. [Google Scholar] [CrossRef]
- Johansen, M.C.; Gottesman, R.F.; Kral, B.G.; Vaidya, D.; Yanek, L.R.; Becker, L.C.; Becker, D.M.; Nyquist, P. Association of coronary artery atherosclerosis with brain white matter hyperintensity. Stroke 2021, 52, 2594–2600. [Google Scholar] [CrossRef]
- Wang, S.-Y.; Gong, P.-Y.; E, Y.; Zhang, Y.-D.; Jiang, T. The role of TREML2 in Alzheimer’s disease. J. Alzheimers Dis. 2020, 76, 799–806. [Google Scholar] [CrossRef]
- Thomas, K.A.; King, R.G.; Sestero, C.M.; Justement, L.B. TREM-like transcript 2 is stored in human neutrophil primary granules and is up-regulated in response to inflammatory mediators. J. Leukoc. Biol. 2016, 100, 177–184. [Google Scholar] [CrossRef]
- Zheng, H.; Liu, C.-C.; Atagi, Y.; Chen, X.-F.; Jia, L.; Yang, L.; He, W.; Zhang, X.; Kang, S.S.; Rosenberry, T.L.; et al. Opposing roles of the triggering receptor expressed on myeloid cells 2 and triggering receptor expressed on myeloid cells-like transcript 2 in microglia activation. Neurobiol. Aging 2016, 42, 132–141. [Google Scholar] [CrossRef]
- Wang, S.-Y.; Fu, X.-X.; Duan, R.; Wei, B.; Cao, H.-M.; Yan, E.; Chen, S.-Y.; Zhang, Y.-D.; Jiang, T. The Alzheimer’s disease-associated gene TREML2 modulates inflammation by regulating microglia polarization and NLRP3 inflammasome activation. Neural Regen. Res. 2023, 18, 434. [Google Scholar] [CrossRef]
- Benitez, B.A.; Jin, S.C.; Guerreiro, R.; Graham, R.; Lord, J.; Harold, D.; Sims, R.; Lambert, J.-C.; Gibbs, J.R.; Bras, J.; et al. Missense variant in TREML2 protects against Alzheimer’s disease. Neurobiol. Aging 2014, 35, 1510.e19–1510.e26. [Google Scholar] [CrossRef]
- European Alzheimer’s Disease Initiative (EADI); Genetic and Environmental Risk in Alzheimer’s Disease (GERAD); Alzheimer’s Disease Genetic Consortium (ADGC); Cohorts for Heart and Aging Research in Genomic Epidemiology (CHARGE); Lambert, J.-C.; Ibrahim-Verbaas, C.A.; Harold, D.; Naj, A.C.; Sims, R.; Bellenguez, C.; et al. Meta-analysis of 74,046 individuals identifies 11 new susceptibility loci for Alzheimer’s disease. Nat. Genet. 2013, 45, 1452–1458. [Google Scholar] [CrossRef]
- Jiang, T.; Wan, Y.; Zhou, J.-S.; Tan, M.-S.; Huang, Q.; Zhu, X.-C.; Lu, H.; Wang, H.-F.; Chen, Q.; Tan, L.; et al. A missense variant in TREML2 reduces risk of Alzheimer’s disease in a Han Chinese population. Mol. Neurobiol. 2017, 54, 977–982. [Google Scholar] [CrossRef]
- Song, Y.-N.; Li, J.-Q.; Tan, C.-C.; Wang, H.-F.; Tan, M.-S.; Cao, X.-P.; Yu, J.-T.; Tan, L.; the Alzheimer’s Disease Neuroimaging Initiative. TREML2 mutation mediate Alzheimer’s disease risk by altering neuronal degeneration. Front. Neurosci. 2019, 13, 455. [Google Scholar] [CrossRef]
- Wang, S.; Xue, X.; Duan, R.; Gong, P.; E, Y.; Jiang, T.; Zhang, Y.; For the Alzheimer’s Disease Neuroimaging Initiative. A TREML2 missense variant influences specific hippocampal subfield volumes in cognitively normal elderly subjects. Brain Behav. 2020, 10, e01573. [Google Scholar] [CrossRef]
- Völzke, H.; Schössow, J.; Schmidt, C.O.; Jürgens, C.; Richter, A.; Werner, A.; Werner, N.; Radke, D.; Teumer, A.; Ittermann, T.; et al. Cohort profile update: The Study of Health in Pomerania (SHIP). Int. J. Epidemiol. 2022. [Google Scholar] [CrossRef]
- Genotype-Tissue Expression (GTEx) Portal (gtexportal.org). Available online: Gtexportal.org (accessed on 18 August 2022).
- Carrasquillo, M.M.; Allen, M.; Burgess, J.D.; Wang, X.; Strickland, S.L.; Aryal, S.; Siuda, J.; Kachadoorian, M.L.; Medway, C.; Younkin, C.S.; et al. A candidate regulatory variant at the TREM gene cluster associates with decreased Alzheimer’s disease risk and increased TREML1 and TREM2 brain gene expression. Alzheimers Dement. 2017, 13, 663–673. [Google Scholar] [CrossRef]
- Cruchaga, C.; Kauwe, J.S.K.; Harari, O.; Jin, S.C.; Cai, Y.; Karch, C.M.; Benitez, B.A.; Jeng, A.T.; Skorupa, T.; Carrell, D.; et al. GWAS of cerebrospinal fluid tau levels identifies risk variants for Alzheimer’s disease. Neuron 2013, 78, 256–268. [Google Scholar] [CrossRef]
- Frenzel, S.; Wittfeld, K.; Habes, M.; Klinger-König, J.; Bülow, R.; Völzke, H.; Grabe, H.J. A biomarker for Alzheimer’s disease based on patterns of regional brain atrophy. Front. Psychiatry 2020, 10, 953. [Google Scholar] [CrossRef]
- Mishra, A.; Duplaà, C.; Vojinovic, D.; Suzuki, H.; Sargurupremraj, M.; Zilhão, N.R.; Li, S.; Bartz, T.M.; Jian, X.; Zhao, W.; et al. Gene-mapping study of extremes of cerebral small vessel disease reveals TRIM47 as a strong candidate. Brain 2022, 145, 1992–2007. [Google Scholar] [CrossRef]
- RKing, G.; Herrin, B.R.; Justement, L.B. Trem-like transcript 2 is expressed on cells of the myeloid/granuloid and B lymphoid lineage and is up-regulated in response to inflammation. J. Immunol. 2006, 176, 6012–6021. [Google Scholar] [CrossRef]
- de Freitas, A.; Banerjee, S.; Xie, N.; Cui, H.; Davis, K.I.; Friggeri, A.; Fu, M.; Abraham, E.; Liu, G. Identification of TLT2 as an engulfment receptor for apoptotic cells. J. Immunol. 2012, 188, 6381–6388. [Google Scholar] [CrossRef]
- Halpert, M.M.; Thomas, K.A.; King, R.G.; Justement, L.B. TLT2 potentiates neutrophil antibacterial activity and chemotaxis in response to G protein-coupled receptor-mediated signaling. J. Immunol. 2011, 187, 2346–2355. [Google Scholar] [CrossRef]
- Schwarz, J.M.; Cooper, D.N.; Schuelke, M.; Seelow, D. MutationTaster2: Mutation prediction for the deep-sequencing age. Nat. Methods 2014, 11, 361–362. [Google Scholar] [CrossRef]
- Adzhubei, I.A.; Schmidt, S.; Peshkin, L.; Ramensky, V.E.; Gerasimova, A.; Bork, P.; Kondrashov, A.S.; Sunyaev, S.R. A method and server for predicting damaging missense mutations. Nat. Methods 2010, 7, 248–249. [Google Scholar] [CrossRef]
- Shen, X.; Song, S.; Li, C.; Zhang, J. Synonymous mutations in representative yeast genes are mostly strongly non-neutral. Nature 2022, 606, 725–736. [Google Scholar] [CrossRef]
- Schwahn, C.; Frenzel, S.; Holtfreter, B.; Van der Auwera, S.; Pink, C.; Bülow, R.; Friedrich, N.; Völzke, H.; Biffar, R.; Kocher, T.; et al. Effect of periodontal treatment on preclinical Alzheimer’s disease—Results of a trial emulation approach. Alzheimers Dement. 2022, 18, 127–141. [Google Scholar] [CrossRef]
- Hegenscheid, K.; Kühn, J.; Völzke, H.; Biffar, R.; Hosten, N.; Puls, R. Whole-body magnetic resonance imaging of healthy volunteers: Pilot study results from the population-based SHIP study. RöFo-Fortschr. Auf Dem Geb. Röntgenstrahlen Bildgeb. Verfahr. 2009, 181, 748–759. [Google Scholar] [CrossRef]
- Hosten, N.; Bülow, R.; Völzke, H.; Domin, M.; Schmidt, C.O.; Teumer, A.; Ittermann, T.; Nauck, M.; Felix, S.; Dörr, M.; et al. SHIP-MR and radiology: 12 years of whole-body magnetic resonance imaging in a single center. Healthcare 2021, 10, 33. [Google Scholar] [CrossRef]
- Schmidt, P.; Gaser, C.; Arsic, M.; Buck, D.; Förschler, A.; Berthele, A.; Hoshi, M.; Ilg, R.; Schmid, V.J.; Zimmer, C.; et al. An automated tool for detection of FLAIR-hyperintense white-matter lesions in multiple sclerosis. NeuroImage 2012, 59, 3774–3783. [Google Scholar] [CrossRef]
- Jack, C.R.; Shiung, M.M.; Weigand, S.D.; O’Brien, P.C.; Gunter, J.L.; Boeve, B.F.; Knopman, D.S.; Smith, G.E.; Ivnik, R.J.; Tangalos, E.G.; et al. Brain atrophy rates predict subsequent clinical conversion in normal elderly and amnestic MCI. Neurology 2005, 65, 1227–1231. [Google Scholar] [CrossRef]
- Schurmann, C. Analysis and Integration of Complex Omics Data of the SHIP Study. Ph.D. Thesis, Ernst-Moritz-Arndt-Universität, Greifswald, Germany, 2013. Available online: https://nbn-resolving.org/urn:nbn:de:gbv:9-001589-3 (accessed on 23 November 2021).
- Das, S.; Forer, L.; Schönherr, S.; Sidore, C.; Locke, A.E.; Kwong, A.; Vrieze, S.I.; Chew, E.Y.; Levy, S.; McGue, M.; et al. Next-generation genotype imputation service and methods. Nat. Genet. 2016, 48, 1284–1287. [Google Scholar] [CrossRef]
- Loh, P.-R.; Palamara, P.F.; Price, A.L. Fast and accurate long-range phasing in a UK Biobank cohort. Nat. Genet. 2016, 48, 811–816. [Google Scholar] [CrossRef]
- The Haplotype Reference Consortium. A reference panel of 64,976 haplotypes for genotype imputation. Nat. Genet. 2016, 48, 1279–1283. [Google Scholar] [CrossRef]
- Habes, M.; Toledo, J.B.; Resnick, S.M.; Doshi, J.; Van der Auwera, S.; Erus, G.; Janowitz, D.; Hegenscheid, K.; Homuth, G.; Völzke, H.; et al. Relationship between APOE genotype and structural MRI measures throughout adulthood in the Study of Health in Pomerania population-based cohort. Am. J. Neuroradiol. 2016, 37, 1636–1642. [Google Scholar] [CrossRef]
- Schurmann, C.; Heim, K.; Schillert, A.; Blankenberg, S.; Carstensen, M.; Dörr, M.; Endlich, K.; Felix, S.B.; Gieger, C.; Grallert, H.; et al. Analyzing Illumina gene expression microarray data from different tissues: Methodological aspects of data analysis in the MetaXpress consortium. PLoS ONE 2012, 7, e50938. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 22 July 2022).
- Du, P.; Kibbe, W.A.; Lin, S.M. lumi: A pipeline for processing Illumina microarray. Bioinformatics 2008, 24, 1547–1548. [Google Scholar] [CrossRef]
- Stekhoven, D.J.; Buhlmann, P. MissForest-non-parametric missing value imputation for mixed-type data. Bioinformatics 2012, 28, 112–118. [Google Scholar] [CrossRef]
- Baumeister, S.E.; Alte, D.; Meyer, C.; John, U. Riskanter Alkoholkonsum und alkoholbezogene Störungen in Vorpommern: Die Studie “Leben und Gesundheit in Vorpommern” (SHIP) und der Bundesgesundheitssurvey 1998 im Vergleich. Gesundheitswesen 2005, 67, 39–47. [Google Scholar] [CrossRef]
- Völzke, H.; Schmidt, C.O.; Hoffmann, W. Research: Increasing value, reducing waste. Lancet 2014, 383, 1124. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| SHIP-TREND-0 (N = 4420) | Expression-MRI Sample (N = 869) | Genetic-MRI Sample (N = 1935) | ||||
|---|---|---|---|---|---|---|
| Mean (sd)/N (%) | #NA | Mean (sd)/N (%) | #NA | Mean (sd)/N (%) | #NA | |
| Main variables: | ||||||
| log-transformed WMH volume * | 3.9 (2.8) [0, 4.3, 5.9] | 2275 | 3.5 (2.7) [0, 4.0, 5.4] | 0 | 3.8 (2.8) [0, 4.2, 5.8] | 0 |
| AD score * | −4.3 (1.4) [−5.3, −4.4, −3.5] | 2345 | −4.5 (1.3) [−5.3, −4.5, −3.8] | 5 | −4.3 (1.4) [−5.3, −4.4, −3.5] | 25 |
| TREML2 gene expression * | ----- | ----- | 7.4 (0.3) [7.2, 7.4, 7.6] | 0 | ----- | ----- |
| rs3747742 status (CC, CT, TT) * | 413 (10%), 1805 (44%), 1902 (46%) | 300 | 82 (10%), 387 (45%), 386 (45%) | 14 | 192 (10%), 846 (44%), 897 (46%) | 0 |
| APOE ε4 status (number of ε4 alleles 0, 1, 2) * | 3052 (76%), 882 (22%), 87 (2%) | 399 | 628 (76%), 181 (22%), 19 (2%) | 41 | 1429 (76%), 414 (22%), 43 (2%) | 49 |
| Covariates: | ||||||
| age at blood sampling (years) | 52.0 (15.5) | 0 | 50.1 (13.6) | 0 | 50.8 (13.9) | 0 |
| sex (women) | 2275 (51%) | 0 | 484 (56%) | 0 | 1002 (52%) | 0 |
| white blood cell count (Gpt/L) | 6.2 (2.6) | 15 | 5.7 (1.5) | 1 | 5.9 (2.1) | 2 |
| red blood cell count (Tpt/L) | 4.6 (0.4) | 15 | 4.6 (0.4) | 1 | 4.7 (0.4) | 2 |
| platelet count (Gpt/L) | 228.1 (54.6) | 15 | 225.9 (50.0) | 1 | 226.5 (52.0) | 2 |
| neutrophils (%) | 58.3 (8.6) | 31 | 57.6 (8.2) | 2 | 58.0 (8.4) | 13 |
| monocytes (%) | 8.8 (2.5) | 31 | 9.0 (2.3) | 2 | 8.9 (2.4) | 13 |
| basophils (%) | 0.5 (0.3) | 203 | 0.5 (0.3) | 3 | 0.48 (0.3) | 44 |
| eosinophils (%) | 2.7 (1.8) | 201 | 2.6 (1.8) | 2 | 2.7 (1.9) | 43 |
| RNA integrity number | ----- | ----- | 8.6 (0.5) | 0 | ----- | ----- |
| storage time (days) | ----- | ----- | 216.4 (155.0) | 0 | ----- | ----- |
| intracranial volume (cm3) * | 1586.8 (159.2) | 2270 | 1576.3 (156.7) | 0 | 1586.6 (158.3) | 0 |
| education (years) | 12.2 (2.6) | 68 | 12.8 (2.4) | 7 | 12.7 (2.4) | 14 |
| income (€) | 1361.5 (702.3) | 187 | 1484.9 (742.1) | 33 | 1457.4 (729.7) | 73 |
| alcohol intake (g/d) | 8.4 (13.4) | 51 | 8.5 (13.3) | 7 | 8.1 (12.0) | 16 |
| in partnership | 3418 (78%) | 13 | 695 (80%) | 2 | 1550 (80%) | 4 |
| body mass index (kg/m2) | 28.1 (5.2) | 7 | 27.1 (4.3) | 0 | 27.5 (4.4) | 0 |
| smoking status (never, ex, current) | 1605 (36%), 1610 (37%), 1183 (27%) | 22 | 364 (42%), 318 (37%), 185 (21%) | 2 | 762 (39%), 703 (36%), 466 (24%) | 4 |
| hypertension | 2123 (48%) | 17 | 325 (37%) | 2 | 825 (43%) | 6 |
| serum total/hdl cholesterol ratio | 4.0 (1.3) | 7 | 3.9 (1.1) | 0 | 4.0 (1.2) | 0 |
| triglycerides (mmol/L) | 1.7 (1.2) | 7 | 1.4 (0.9) | 0 | 1.5 (1.1) | 0 |
| WMH~TREML2 Expression | Effect | 95% CI | p-Value | N |
|---|---|---|---|---|
| base model | −0.77 | −1.37; −0.17 | 0.012 | 869 |
| sensitivity analyses: | ||||
| socioeconomic model | −0.79 | −1.39; −0.19 | 0.0098 | 869 |
| cardiovascular model | −0.76 | −1.38; −0.15 | 0.015 | 869 |
| full model | −0.79 | −1.40; −0.17 | 0.012 | 869 |
| AD Score~rs3747742 | Effect | 95% CI | p-Value | N |
|---|---|---|---|---|
| base model | 0.10 | 0.020; 0.19 | 0.015 | 1910 |
| sensitivity analyses: | ||||
| socioeconomic model | 0.10 | 0.018; 0.19 | 0.017 | 1910 |
| cardiovascular model | 0.10 | 0.020; 0.19 | 0.015 | 1910 |
| full model | 0.10 | 0.019; 0.19 | 0.016 | 1910 |
| restriction to TREND-Batch1 | 0.052 | −0.069; 0.17 | 0.40 | 860 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kühn, A.L.; Frenzel, S.; Teumer, A.; Wittfeld, K.; Garvert, L.; Weihs, A.; Homuth, G.; Prokisch, H.; Bülow, R.; Nauck, M.; et al. TREML2 Gene Expression and Its Missense Variant rs3747742 Associate with White Matter Hyperintensity Volume and Alzheimer’s Disease-Related Brain Atrophy in the General Population. Int. J. Mol. Sci. 2022, 23, 13764. https://doi.org/10.3390/ijms232213764
Kühn AL, Frenzel S, Teumer A, Wittfeld K, Garvert L, Weihs A, Homuth G, Prokisch H, Bülow R, Nauck M, et al. TREML2 Gene Expression and Its Missense Variant rs3747742 Associate with White Matter Hyperintensity Volume and Alzheimer’s Disease-Related Brain Atrophy in the General Population. International Journal of Molecular Sciences. 2022; 23(22):13764. https://doi.org/10.3390/ijms232213764
Chicago/Turabian StyleKühn, Annemarie Luise, Stefan Frenzel, Alexander Teumer, Katharina Wittfeld, Linda Garvert, Antoine Weihs, Georg Homuth, Holger Prokisch, Robin Bülow, Matthias Nauck, and et al. 2022. "TREML2 Gene Expression and Its Missense Variant rs3747742 Associate with White Matter Hyperintensity Volume and Alzheimer’s Disease-Related Brain Atrophy in the General Population" International Journal of Molecular Sciences 23, no. 22: 13764. https://doi.org/10.3390/ijms232213764
APA StyleKühn, A. L., Frenzel, S., Teumer, A., Wittfeld, K., Garvert, L., Weihs, A., Homuth, G., Prokisch, H., Bülow, R., Nauck, M., Völker, U., Völzke, H., Grabe, H. J., & Van der Auwera, S. (2022). TREML2 Gene Expression and Its Missense Variant rs3747742 Associate with White Matter Hyperintensity Volume and Alzheimer’s Disease-Related Brain Atrophy in the General Population. International Journal of Molecular Sciences, 23(22), 13764. https://doi.org/10.3390/ijms232213764