



Genetics and Epigenetics of the X and Y Chromosomes in the Sexual Differentiation of the Brain

 , ,
, ,  and
and 
Abstract
1. Sex and Brain: Not All about Gonadal Hormones
2. Sex Chromosome Complement and Brain Sex Differences
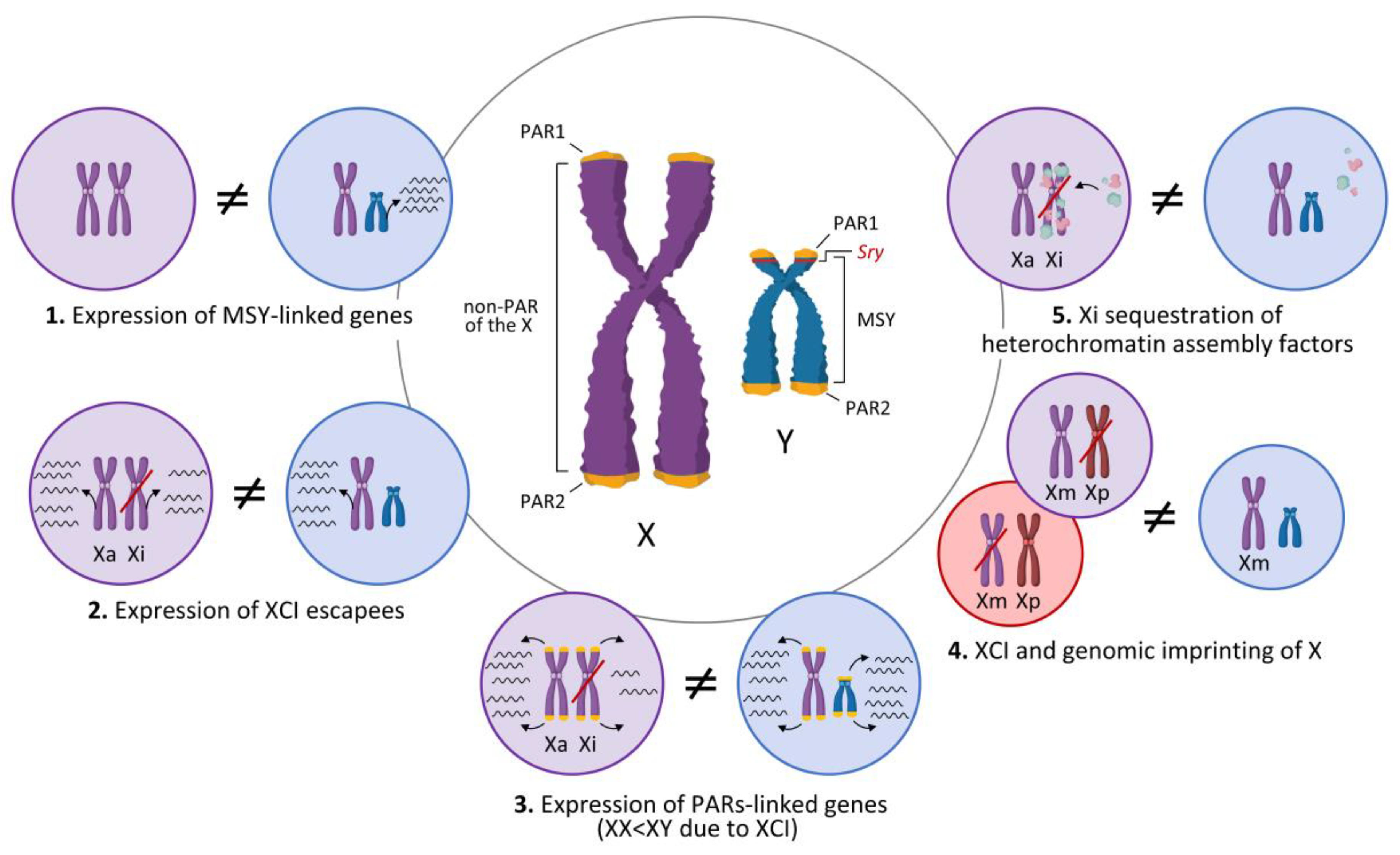
3. Genetics and Epigenetics of the X and Y Chromosomes and Sexual Differentiation
4. X-Linked Genes and Sexual Differentiation of the Brain
5. Y-Linked Genes and Sexual Differentiation of the Brain
6. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Bao, A.M.; Swaab, D.F. Sexual differentiation of the human brain: Relation to gender identity, sexual orientation and neuropsychiatric disorders. Front. Neuroendocrinol. 2011, 32, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Phoenix, C.H.; Goy, R.W.; Gerall, A.A.; Young, W.C. Organizing action of prenatally administered testosterone propionate on the tissues mediating mating behavior in the female guinea pig. Endocrinology 1959, 65, 369–382. [Google Scholar] [CrossRef] [PubMed]
- Beach, F.A.; Noble, R.G.; Orndoff, R.K. Effects of perinatal androgen treatment on responses of male rats to gonadal hormones in adulthood. J. Comp. Physiol. Psychol. 1969, 68, 490–497. [Google Scholar] [CrossRef]
- Stern, J.J. Neonatal castration, androstenedione, and the mating behavior of the male rat. J. Comp. Physiol. Psychol. 1969, 69, 608–612. [Google Scholar] [CrossRef] [PubMed]
- Baum, M.J. A comparison of the effects of methyltrienolone (R 1881) and 5 alpha-dihydrotestosterone on sexual behavior of castrated male rats. Horm. Behav. 1979, 13, 165–174. [Google Scholar] [CrossRef]
- Baum, M.J.; Gallagher, C.A.; Martin, J.T.; Damassa, D.A. Effects of testosterone, dihydrotestosterone, or estradiol administered neonatally on sexual behavior of female ferrets. Endocrinology 1982, 111, 773–780. [Google Scholar] [CrossRef] [PubMed]
- Arnold, A.P.; Gorski, R.A. Gonadal steroid induction of structural sex differences in the central nervous system. Annu. Rev. Neurosci. 1984, 7, 413–442. [Google Scholar] [CrossRef] [PubMed]
- Warren, D.W.; Haltmeyer, G.C.; Eik-Nes, K.B. Testosterone in the fetal rat testis. Biol. Reprod. 1973, 8, 560–565. [Google Scholar] [CrossRef] [PubMed]
- Huhtaniemi, I. Fetal testis—a very special endocrine organ. Eur. J. Endocrinol. 1994, 130, 25–31. [Google Scholar] [CrossRef]
- O’Shaughnessy, P.J.; Baker, P.J.; Johnston, H. The foetal Leydig cell—Differentiation, function and regulation. Int. J. Androl. 2006, 29, 90–95; discussion 105–108. [Google Scholar] [CrossRef]
- MacLusky, N.J.; Naftolin, F. Sexual differentiation of the central nervous system. Science 1981, 211, 1294–1302. [Google Scholar] [CrossRef] [PubMed]
- Wilson, C.A.; Davies, D.C. The control of sexual differentiation of the reproductive system and brain. Reproduction 2007, 133, 331–359. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, M.M. Estradiol and the developing brain. Physiol. Rev. 2008, 88, 91–124. [Google Scholar] [CrossRef] [PubMed]
- Negri-Cesi, P.; Colciago, A.; Pravettoni, A.; Casati, L.; Conti, L.; Celotti, F. Sexual differentiation of the rodent hypothalamus: Hormonal and environmental influences. J. Steroid Biochem. Mol. Biol. 2008, 109, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Arnold, A.P. Sex Differences in the Age of Genetics. In Hormones, Brain, and Behavior, 3rd ed.; Pfaff, D.W., Joëls, M., Eds.; Academic Press, Elsevier: Toronto, ON, Canada, 2017; Volume 5, pp. 33–48. [Google Scholar]
- Arnold, A.P.; Burgoyne, P.S. Are XX and XY brain cells intrinsically different? Trends Endocrinol. Metab. 2004, 15, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Charlesworth, B. The evolution of chromosomal sex determination and dosage compensation. Curr. Biol. 1996, 6, 149–162. [Google Scholar] [CrossRef]
- Goodfellow, P.N.; Lovell-Badge, R. SRY and sex determination in mammals. Annu. Rev. Genet. 1993, 27, 71–92. [Google Scholar] [CrossRef] [PubMed]
- Makela, J.A.; Koskenniemi, J.J.; Virtanen, H.E.; Toppari, J. Testis Development. Endocr. Rev. 2019, 40, 857–905. [Google Scholar] [CrossRef] [PubMed]
- Cambiasso, M.J.; Diaz, H.; Caceres, A.; Carrer, H.F. Neuritogenic effect of estradiol on rat ventromedial hypothalamic neurons co-cultured with homotopic or heterotopic glia. J. Neurosci. Res. 1995, 42, 700–709. [Google Scholar] [CrossRef] [PubMed]
- Carrer, H.F.; Cambiasso, M.J. Sexual differentiation of the brain: Genes, estrogen, and neurotrophic factors. Cell. Mol. Neurobiol. 2002, 22, 479–500. [Google Scholar] [CrossRef] [PubMed]
- Reisert, I.; Pilgrim, C. Sexual differentiation of monoaminergic neurons—Genetic or epigenetic? Trends Neurosci. 1991, 14, 468–473. [Google Scholar] [CrossRef]
- Engele, J.; Pilgrim, C.; Reisert, I. Sexual differentiation of mesencephalic neurons in vitro: Effects of sex and gonadal hormones. Int. J. Dev. Neurosci. 1989, 7, 603–611. [Google Scholar] [CrossRef]
- Beyer, C.; Pilgrim, C.; Reisert, I. Dopamine content and metabolism in mesencephalic and diencephalic cell cultures: Sex differences and effects of sex steroids. J. Neurosci. 1991, 11, 1325–1333. [Google Scholar] [CrossRef] [PubMed]
- Nottebohm, F.; Arnold, A.P. Sexual dimorphism in vocal control areas of the songbird brain. Science 1976, 194, 211–213. [Google Scholar] [CrossRef]
- Gurney, M.E.; Konishi, M. Hormone-induced sexual differentiation of brain and behavior in zebra finches. Science 1980, 208, 1380–1383. [Google Scholar] [CrossRef]
- Simpson, H.B.; Vicario, D.S. Early estrogen treatment of female zebra finches masculinizes the brain pathway for learned vocalizations. J. Neurobiol. 1991, 22, 777–793. [Google Scholar] [CrossRef]
- Holloway, C.C.; Clayton, D.F. Estrogen synthesis in the male brain triggers development of the avian song control pathway in vitro. Nat. Neurosci. 2001, 4, 170–175. [Google Scholar] [CrossRef]
- Arnold, A.P. Sexual differentiation of the zebra finch song system: Positive evidence, negative evidence, null hypotheses, and a paradigm shift. J. Neurobiol. 1997, 33, 572–584. [Google Scholar] [CrossRef]
- Agate, R.J.; Grisham, W.; Wade, J.; Mann, S.; Wingfield, J.; Schanen, C.; Palotie, A.; Arnold, A.P. Neural, not gonadal, origin of brain sex differences in a gynandromorphic finch. Proc. Natl. Acad. Sci. USA 2003, 100, 4873–4878. [Google Scholar] [CrossRef]
- Lovell-Badge, R.; Robertson, E. XY female mice resulting from a heritable mutation in the primary testis-determining gene, Tdy. Development 1990, 109, 635–646. [Google Scholar] [CrossRef]
- Mahadevaiah, S.K.; Odorisio, T.; Elliott, D.J.; Rattigan, A.; Szot, M.; Laval, S.H.; Washburn, L.L.; McCarrey, J.R.; Cattanach, B.M.; Lovell-Badge, R.; et al. Mouse homologues of the human AZF candidate gene RBM are expressed in spermatogonia and spermatids, and map to a Y chromosome deletion interval associated with a high incidence of sperm abnormalities. Hum. Mol. Genet. 1998, 7, 715–727. [Google Scholar] [CrossRef] [PubMed]
- Itoh, Y.; Mackie, R.; Kampf, K.; Domadia, S.; Brown, J.D.; O’Neill, R.; Arnold, A.P. Four core genotypes mouse model: Localization of the Sry transgene and bioassay for testicular hormone levels. BMC Res. Notes 2015, 8, 69. [Google Scholar] [CrossRef] [PubMed]
- Carruth, L.L.; Reisert, I.; Arnold, A.P. Sex chromosome genes directly affect brain sexual differentiation. Nat. Neurosci. 2002, 5, 933–934. [Google Scholar] [CrossRef] [PubMed]
- De Vries, G.J.; Rissman, E.F.; Simerly, R.B.; Yang, L.Y.; Scordalakes, E.M.; Auger, C.J.; Swain, A.; Lovell-Badge, R.; Burgoyne, P.S.; Arnold, A.P. A model system for study of sex chromosome effects on sexually dimorphic neural and behavioral traits. J. Neurosci. 2002, 22, 9005–9014. [Google Scholar] [CrossRef] [PubMed]
- Gatewood, J.D.; Wills, A.; Shetty, S.; Xu, J.; Arnold, A.P.; Burgoyne, P.S.; Rissman, E.F. Sex chromosome complement and gonadal sex influence aggressive and parental behaviors in mice. J. Neurosci. 2006, 26, 2335–2342. [Google Scholar] [CrossRef]
- Simpson, E.R.; Mahendroo, M.S.; Means, G.D.; Kilgore, M.W.; Hinshelwood, M.M.; Graham-Lorence, S.; Amarneh, B.; Ito, Y.; Fisher, C.R.; Michael, M.D.; et al. Aromatase cytochrome P450, the enzyme responsible for estrogen biosynthesis. Endocr. Rev. 1994, 15, 342–355. [Google Scholar] [CrossRef]
- Brooks, D.C.; Coon, V.J.; Ercan, C.M.; Xu, X.; Dong, H.; Levine, J.E.; Bulun, S.E.; Zhao, H. Brain Aromatase and the Regulation of Sexual Activity in Male Mice. Endocrinology 2020, 161, bqaa137. [Google Scholar] [CrossRef]
- Cisternas, C.D.; Garcia-Segura, L.M.; Cambiasso, M.J. Hormonal and genetic factors interact to control aromatase expression in the developing brain. J. Neuroendocrinol. 2018, 30, e12535. [Google Scholar] [CrossRef]
- Cisternas, C.D.; Cabrera Zapata, L.E.; Arevalo, M.A.; Garcia-Segura, L.M.; Cambiasso, M.J. Regulation of aromatase expression in the anterior amygdala of the developing mouse brain depends on ERbeta and sex chromosome complement. Sci. Rep. 2017, 7, 5320. [Google Scholar] [CrossRef]
- Cisternas, C.D.; Tome, K.; Caeiro, X.E.; Dadam, F.M.; Garcia-Segura, L.M.; Cambiasso, M.J. Sex chromosome complement determines sex differences in aromatase expression and regulation in the stria terminalis and anterior amygdala of the developing mouse brain. Mol. Cell. Endocrinol. 2015, 414, 99–110. [Google Scholar] [CrossRef]
- Ruiz-Palmero, I.; Ortiz-Rodriguez, A.; Melcangi, R.C.; Caruso, D.; Garcia-Segura, L.M.; Rune, G.M.; Arevalo, M.A. Oestradiol synthesized by female neurons generates sex differences in neuritogenesis. Sci. Rep. 2016, 6, 31891. [Google Scholar] [CrossRef] [PubMed]
- Davis, E.J.; Broestl, L.; Abdulai-Saiku, S.; Worden, K.; Bonham, L.W.; Minones-Moyano, E.; Moreno, A.J.; Wang, D.; Chang, K.; Williams, G.; et al. A second X chromosome contributes to resilience in a mouse model of Alzheimer’s disease. Sci. Transl. Med. 2020, 12, eaaz5677. [Google Scholar] [CrossRef]
- Gioiosa, L.; Chen, X.; Watkins, R.; Klanfer, N.; Bryant, C.D.; Evans, C.J.; Arnold, A.P. Sex chromosome complement affects nociception in tests of acute and chronic exposure to morphine in mice. Horm. Behav. 2008, 53, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Quinn, J.J.; Hitchcott, P.K.; Umeda, E.A.; Arnold, A.P.; Taylor, J.R. Sex chromosome complement regulates habit formation. Nat. Neurosci. 2007, 10, 1398–1400. [Google Scholar] [CrossRef]
- Smith-Bouvier, D.L.; Divekar, A.A.; Sasidhar, M.; Du, S.; Tiwari-Woodruff, S.K.; King, J.K.; Arnold, A.P.; Singh, R.R.; Voskuhl, R.R. A role for sex chromosome complement in the female bias in autoimmune disease. J. Exp. Med. 2008, 205, 1099–1108. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Watkins, R.; Delot, E.; Reliene, R.; Schiestl, R.H.; Burgoyne, P.S.; Arnold, A.P. Sex difference in neural tube defects in p53-null mice is caused by differences in the complement of X not Y genes. Dev. Neurobiol. 2008, 68, 265–273. [Google Scholar] [CrossRef]
- Caeiro, X.E.; Mir, F.R.; Vivas, L.M.; Carrer, H.F.; Cambiasso, M.J. Sex chromosome complement contributes to sex differences in bradycardic baroreflex response. Hypertension 2011, 58, 505–511. [Google Scholar] [CrossRef]
- Dadam, F.M.; Caeiro, X.E.; Cisternas, C.D.; Macchione, A.F.; Cambiasso, M.J.; Vivas, L. Effect of sex chromosome complement on sodium appetite and Fos-immunoreactivity induced by sodium depletion. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 306, R175–R184. [Google Scholar] [CrossRef]
- Dadam, F.M.; Cisternas, C.D.; Macchione, A.F.; Godino, A.; Antunes-Rodrigues, J.; Cambiasso, M.J.; Vivas, L.M.; Caeiro, X.E. Sex chromosome complement involvement in angiotensin receptor sexual dimorphism. Mol. Cell. Endocrinol. 2017, 447, 98–105. [Google Scholar] [CrossRef]
- Sneddon, E.A.; Rasizer, L.N.; Cavalco, N.G.; Jaymes, A.H.; Ostlie, N.J.; Minshall, B.L.; Masters, B.M.; Hughes, M.R.; Hrncir, H.; Arnold, A.P.; et al. Gonadal hormones and sex chromosome complement differentially contribute to ethanol intake, preference, and relapse-like behaviour in four core genotypes mice. Addict. Biol. 2022, 27, e13222. [Google Scholar] [CrossRef]
- Arnold, A.P. Four Core Genotypes and XY* mouse models: Update on impact on SABV research. Neurosci. Biobehav. Rev. 2020, 119, 1–8. [Google Scholar] [CrossRef]
- Muller, H.J. Genetic Variability, Twin Hybrids and Constant Hybrids, in a Case of Balanced Lethal Factors. Genetics 1918, 3, 422–499. [Google Scholar] [CrossRef] [PubMed]
- Muller, H.J. A gene for the fourth chromosome of Drosophila. J. Exp. Zool. 1914, 17, 325–336. [Google Scholar] [CrossRef]
- Graves, J.A. Sex chromosome specialization and degeneration in mammals. Cell 2006, 124, 901–914. [Google Scholar] [CrossRef] [PubMed]
- Hughes, J.F.; Page, D.C. The Biology and Evolution of Mammalian Y Chromosomes. Annu. Rev. Genet. 2015, 49, 507–527. [Google Scholar] [CrossRef] [PubMed]
- Graves, J.A. Evolution of vertebrate sex chromosomes and dosage compensation. Nat. Rev. Genet. 2016, 17, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Wijchers, P.J.; Festenstein, R.J. Epigenetic regulation of autosomal gene expression by sex chromosomes. Trends Genet. 2011, 27, 132–140. [Google Scholar] [CrossRef]
- Simmler, M.C.; Rouyer, F.; Vergnaud, G.; Nystrom-Lahti, M.; Ngo, K.Y.; de la Chapelle, A.; Weissenbach, J. Pseudoautosomal DNA sequences in the pairing region of the human sex chromosomes. Nature 1985, 317, 692–697. [Google Scholar] [CrossRef]
- Charchar, F.J.; Svartman, M.; El-Mogharbel, N.; Ventura, M.; Kirby, P.; Matarazzo, M.R.; Ciccodicola, A.; Rocchi, M.; D’Esposito, M.; Graves, J.A. Complex events in the evolution of the human pseudoautosomal region 2 (PAR2). Genome Res. 2003, 13, 281–286. [Google Scholar] [CrossRef]
- Monteiro, B.; Arenas, M.; Prata, M.J.; Amorim, A. Evolutionary dynamics of the human pseudoautosomal regions. PLoS Genet. 2021, 17, e1009532. [Google Scholar] [CrossRef]
- Perry, J.; Palmer, S.; Gabriel, A.; Ashworth, A. A short pseudoautosomal region in laboratory mice. Genome Res. 2001, 11, 1826–1832. [Google Scholar] [CrossRef] [PubMed]
- Morgan, A.P.; Bell, T.A.; Crowley, J.J.; Pardo-Manuel de Villena, F. Instability of the Pseudoautosomal Boundary in House Mice. Genetics 2019, 212, 469–487. [Google Scholar] [CrossRef] [PubMed]
- Morey, C.; Avner, P. Genetics and epigenetics of the X chromosome. Ann. N. Y. Acad. Sci. 2010, 1214, E18–E33. [Google Scholar] [CrossRef] [PubMed]
- Raznahan, A.; Parikshak, N.N.; Chandran, V.; Blumenthal, J.D.; Clasen, L.S.; Alexander-Bloch, A.F.; Zinn, A.R.; Wangsa, D.; Wise, J.; Murphy, D.G.M.; et al. Sex-chromosome dosage effects on gene expression in humans. Proc. Natl. Acad. Sci. USA 2018, 115, 7398–7403. [Google Scholar] [CrossRef]
- Bellott, D.W.; Hughes, J.F.; Skaletsky, H.; Brown, L.G.; Pyntikova, T.; Cho, T.J.; Koutseva, N.; Zaghlul, S.; Graves, T.; Rock, S.; et al. Mammalian Y chromosomes retain widely expressed dosage-sensitive regulators. Nature 2014, 508, 494–499. [Google Scholar] [CrossRef] [PubMed]
- Cortez, D.; Marin, R.; Toledo-Flores, D.; Froidevaux, L.; Liechti, A.; Waters, P.D.; Grutzner, F.; Kaessmann, H. Origins and functional evolution of Y chromosomes across mammals. Nature 2014, 508, 488–493. [Google Scholar] [CrossRef] [PubMed]
- Lyon, M.F. Gene action in the X-chromosome of the mouse (Mus musculus L.). Nature 1961, 190, 372–373. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.T.; Bartolomei, M.S. X-inactivation, imprinting, and long noncoding RNAs in health and disease. Cell 2013, 152, 1308–1323. [Google Scholar] [CrossRef]
- Gayen, S.; Maclary, E.; Hinten, M.; Kalantry, S. Sex-specific silencing of X-linked genes by Xist RNA. Proc. Natl. Acad. Sci. USA 2016, 113, E309–E318. [Google Scholar] [CrossRef] [PubMed]
- Penny, G.D.; Kay, G.F.; Sheardown, S.A.; Rastan, S.; Brockdorff, N. Requirement for Xist in X chromosome inactivation. Nature 1996, 379, 131–137. [Google Scholar] [CrossRef]
- Marahrens, Y.; Loring, J.; Jaenisch, R. Role of the Xist gene in X chromosome choosing. Cell 1998, 92, 657–664. [Google Scholar] [CrossRef]
- Disteche, C.M.; Berletch, J.B. X-chromosome inactivation and escape. J. Genet. 2015, 94, 591–599. [Google Scholar] [CrossRef]
- Babak, T.; DeVeale, B.; Tsang, E.K.; Zhou, Y.; Li, X.; Smith, K.S.; Kukurba, K.R.; Zhang, R.; Li, J.B.; van der Kooy, D.; et al. Genetic conflict reflected in tissue-specific maps of genomic imprinting in human and mouse. Nat. Genet. 2015, 47, 544–549. [Google Scholar] [CrossRef] [PubMed]
- Davies, W.; Isles, A.; Smith, R.; Karunadasa, D.; Burrmann, D.; Humby, T.; Ojarikre, O.; Biggin, C.; Skuse, D.; Burgoyne, P.; et al. Xlr3b is a new imprinted candidate for X-linked parent-of-origin effects on cognitive function in mice. Nat. Genet. 2005, 37, 625–629. [Google Scholar] [CrossRef] [PubMed]
- Raefski, A.S.; O’Neill, M.J. Identification of a cluster of X-linked imprinted genes in mice. Nat. Genet. 2005, 37, 620–624. [Google Scholar] [CrossRef] [PubMed]
- Thornhill, A.R.; Burgoyne, P.S. A paternally imprinted X chromosome retards the development of the early mouse embryo. Development 1993, 118, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S.; Isotani, A.; Mise, N.; Yamamoto, M.; Fujihara, Y.; Kaseda, K.; Nakanishi, T.; Ikawa, M.; Hamada, H.; Abe, K.; et al. Comparison of gene expression in male and female mouse blastocysts revealed imprinting of the X-linked gene, Rhox5/Pem, at preimplantation stages. Curr. Biol. 2006, 16, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Berletch, J.B.; Yang, F.; Disteche, C.M. Escape from X inactivation in mice and humans. Genome Biol. 2010, 11, 213. [Google Scholar] [CrossRef] [PubMed]
- Carrel, L.; Willard, H.F. X-inactivation profile reveals extensive variability in X-linked gene expression in females. Nature 2005, 434, 400–404. [Google Scholar] [CrossRef] [PubMed]
- Tukiainen, T.; Villani, A.C.; Yen, A.; Rivas, M.A.; Marshall, J.L.; Satija, R.; Aguirre, M.; Gauthier, L.; Fleharty, M.; Kirby, A.; et al. Landscape of X chromosome inactivation across human tissues. Nature 2017, 550, 244–248. [Google Scholar] [CrossRef]
- Berletch, J.B.; Ma, W.; Yang, F.; Shendure, J.; Noble, W.S.; Disteche, C.M.; Deng, X. Escape from X inactivation varies in mouse tissues. PLoS Genet. 2015, 11, e1005079. [Google Scholar] [CrossRef]
- Peeters, S.B.; Cotton, A.M.; Brown, C.J. Variable escape from X-chromosome inactivation: Identifying factors that tip the scales towards expression. Bioessays 2014, 36, 746–756. [Google Scholar] [CrossRef] [PubMed]
- Balaton, B.P.; Brown, C.J. Escape Artists of the X Chromosome. Trends Genet. 2016, 32, 348–359. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Cobos, M.J.; Balaton, B.P.; Brown, C.J. Genes that escape from X-chromosome inactivation: Potential contributors to Klinefelter syndrome. Am. J. Med. Genet. C Semin. Med. Genet. 2020, 184, 226–238. [Google Scholar] [CrossRef] [PubMed]
- Garieri, M.; Stamoulis, G.; Blanc, X.; Falconnet, E.; Ribaux, P.; Borel, C.; Santoni, F.; Antonarakis, S.E. Extensive cellular heterogeneity of X inactivation revealed by single-cell allele-specific expression in human fibroblasts. Proc. Natl. Acad. Sci. USA 2018, 115, 13015–13020. [Google Scholar] [CrossRef] [PubMed]
- Burgoyne, P.S.; Arnold, A.P. A primer on the use of mouse models for identifying direct sex chromosome effects that cause sex differences in non-gonadal tissues. Biol. Sex Differ. 2016, 7, 68. [Google Scholar] [CrossRef] [PubMed]
- Arnold, A.P. X chromosome agents of sexual differentiation. Nat. Rev. Endocrinol. 2022, 18, 574–583. [Google Scholar] [CrossRef] [PubMed]
- Wijchers, P.J.; Yandim, C.; Panousopoulou, E.; Ahmad, M.; Harker, N.; Saveliev, A.; Burgoyne, P.S.; Festenstein, R. Sexual dimorphism in mammalian autosomal gene regulation is determined not only by Sry but by sex chromosome complement as well. Dev. Cell 2010, 19, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Bermejo-Alvarez, P.; Rizos, D.; Rath, D.; Lonergan, P.; Gutierrez-Adan, A. Sex determines the expression level of one third of the actively expressed genes in bovine blastocysts. Proc. Natl. Acad. Sci. USA 2010, 107, 3394–3399. [Google Scholar] [CrossRef]
- Blencowe, M.; Chen, X.; Zhao, Y.; Itoh, Y.; McQuillen, C.N.; Han, Y.; Shou, B.L.; McClusky, R.; Reue, K.; Arnold, A.P.; et al. Relative contributions of sex hormones, sex chromosomes, and gonads to sex differences in tissue gene regulation. Genome Res. 2022, 32, 807–824. [Google Scholar] [CrossRef] [PubMed]
- Zechner, U.; Wilda, M.; Kehrer-Sawatzki, H.; Vogel, W.; Fundele, R.; Hameister, H. A high density of X-linked genes for general cognitive ability: A run-away process shaping human evolution? Trends Genet. 2001, 17, 697–701. [Google Scholar] [CrossRef]
- Ross, M.T.; Grafham, D.V.; Coffey, A.J.; Scherer, S.; McLay, K.; Muzny, D.; Platzer, M.; Howell, G.R.; Burrows, C.; Bird, C.P.; et al. The DNA sequence of the human X chromosome. Nature 2005, 434, 325–337. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.K.; Disteche, C.M. High expression of the mammalian X chromosome in brain. Brain Res. 2006, 1126, 46–49. [Google Scholar] [CrossRef]
- Deng, X.; Hiatt, J.B.; Nguyen, D.K.; Ercan, S.; Sturgill, D.; Hillier, L.W.; Schlesinger, F.; Davis, C.A.; Reinke, V.J.; Gingeras, T.R.; et al. Evidence for compensatory upregulation of expressed X-linked genes in mammals, Caenorhabditis elegans and Drosophila melanogaster. Nat. Genet. 2011, 43, 1179–1185. [Google Scholar] [CrossRef]
- Nguyen, D.K.; Disteche, C.M. Dosage compensation of the active X chromosome in mammals. Nat. Genet. 2006, 38, 47–53. [Google Scholar] [CrossRef]
- Ropers, H.H.; Hamel, B.C. X-linked mental retardation. Nat. Rev. Genet. 2005, 6, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Chiurazzi, P.; Schwartz, C.E.; Gecz, J.; Neri, G. XLMR genes: Update 2007. Eur. J. Hum. Genet. 2008, 16, 422–434. [Google Scholar] [CrossRef] [PubMed]
- Vacca, M.; Della Ragione, F.; Scalabri, F.; D’Esposito, M. X inactivation and reactivation in X-linked diseases. Semin. Cell Dev. Biol. 2016, 56, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Xu, W.; Lan, F. Histone lysine demethylases in mammalian embryonic development. Exp. Mol. Med. 2017, 49, e325. [Google Scholar] [CrossRef] [PubMed]
- Hoffbuhr, K.C.; Moses, L.M.; Jerdonek, M.A.; Naidu, S.; Hoffman, E.P. Associations between MeCP2 mutations, X-chromosome inactivation, and phenotype. Ment. Retard. Dev. Disabil. Res. Rev. 2002, 8, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Deardorff, M.A.; Porter, N.J.; Christianson, D.W. Structural aspects of HDAC8 mechanism and dysfunction in Cornelia de Lange syndrome spectrum disorders. Protein Sci. 2016, 25, 1965–1976. [Google Scholar] [CrossRef] [PubMed]
- Kadlec, J.; Hallacli, E.; Lipp, M.; Holz, H.; Sanchez-Weatherby, J.; Cusack, S.; Akhtar, A. Structural basis for MOF and MSL3 recruitment into the dosage compensation complex by MSL1. Nat. Struct. Mol. Biol. 2011, 18, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Tominaga, K.; Pereira-Smith, O.M. Emerging role of the MORF/MRG gene family in various biological processes, including aging. Ann. N. Y. Acad. Sci. 2010, 1197, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Jahani-Asl, A.; Cheng, C.; Zhang, C.; Bonni, A. Pathogenesis of Borjeson-Forssman-Lehmann syndrome: Insights from PHF6 function. Neurobiol. Dis. 2016, 96, 227–235. [Google Scholar] [CrossRef]
- Xu, J.; Watkins, R.; Arnold, A.P. Sexually dimorphic expression of the X-linked gene Eif2s3x mRNA but not protein in mouse brain. Gene Expr. Patterns 2006, 6, 146–155. [Google Scholar] [CrossRef]
- Bagni, C.; Oostra, B.A. Fragile X syndrome: From protein function to therapy. Am. J. Med. Genet. A 2013, 161, 2809–2821. [Google Scholar] [CrossRef]
- Snijders Blok, L.; Madsen, E.; Juusola, J.; Gilissen, C.; Baralle, D.; Reijnders, M.R.; Venselaar, H.; Helsmoortel, C.; Cho, M.T.; Hoischen, A.; et al. Mutations in DDX3X Are a Common Cause of Unexplained Intellectual Disability with Gender-Specific Effects on Wnt Signaling. Am. J. Hum. Genet. 2015, 97, 343–352. [Google Scholar] [CrossRef]
- Stegeman, S.; Jolly, L.A.; Premarathne, S.; Gecz, J.; Richards, L.J.; Mackay-Sim, A.; Wood, S.A. Loss of Usp9x disrupts cortical architecture, hippocampal development and TGFbeta-mediated axonogenesis. PLoS ONE 2013, 8, e68287. [Google Scholar] [CrossRef]
- Lu, T.; Chen, R.; Cox, T.C.; Moldrich, R.X.; Kurniawan, N.; Tan, G.; Perry, J.K.; Ashworth, A.; Bartlett, P.F.; Xu, L.; et al. X-linked microtubule-associated protein, Mid1, regulates axon development. Proc. Natl. Acad. Sci. USA 2013, 110, 19131–19136. [Google Scholar] [CrossRef]
- Levine, Z.G.; Walker, S. The Biochemistry of O-GlcNAc Transferase: Which Functions Make It Essential in Mammalian Cells? Annu. Rev. Biochem. 2016, 85, 631–657. [Google Scholar] [CrossRef]
- Gordon, S.L.; Cousin, M.A. X-linked intellectual disability-associated mutations in synaptophysin disrupt synaptobrevin II retrieval. J. Neurosci. 2013, 33, 13695–13700. [Google Scholar] [CrossRef] [PubMed]
- Christophe-Hobertus, C.; Szpirer, C.; Guyon, R.; Christophe, D. Identification of the gene encoding Brain Cell Membrane Protein 1 (BCMP1), a putative four-transmembrane protein distantly related to the Peripheral Myelin Protein 22/Epithelial Membrane Proteins and the Claudins. BMC Genom. 2001, 2, 3. [Google Scholar] [CrossRef] [PubMed]
- Raznahan, A.; Disteche, C.M. X-chromosome regulation and sex differences in brain anatomy. Neurosci. Biobehav. Rev. 2021, 120, 28–47. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Cho, Y.W.; Yu, L.R.; Yu, H.; Veenstra, T.D.; Ge, K. Identification of JmjC domain-containing UTX and JMJD3 as histone H3 lysine 27 demethylases. Proc. Natl. Acad. Sci. USA 2007, 104, 18439–18444. [Google Scholar] [CrossRef] [PubMed]
- Tran, N.; Broun, A.; Ge, K. Lysine Demethylase KDM6A in Differentiation, Development, and Cancer. Mol. Cell. Biol. 2020, 40, e00341-20. [Google Scholar] [CrossRef]
- Shpargel, K.B.; Sengoku, T.; Yokoyama, S.; Magnuson, T. UTX and UTY demonstrate histone demethylase-independent function in mouse embryonic development. PLoS Genet. 2012, 8, e1002964. [Google Scholar] [CrossRef]
- Walport, L.J.; Hopkinson, R.J.; Vollmar, M.; Madden, S.K.; Gileadi, C.; Oppermann, U.; Schofield, C.J.; Johansson, C. Human UTY(KDM6C) is a male-specific N-methyl lysyl demethylase. J. Biol. Chem. 2014, 289, 18302–18313. [Google Scholar] [CrossRef]
- Cho, Y.W.; Hong, T.; Hong, S.; Guo, H.; Yu, H.; Kim, D.; Guszczynski, T.; Dressler, G.R.; Copeland, T.D.; Kalkum, M.; et al. PTIP associates with MLL3- and MLL4-containing histone H3 lysine 4 methyltransferase complex. J. Biol. Chem. 2007, 282, 20395–20406. [Google Scholar] [CrossRef]
- Tie, F.; Banerjee, R.; Conrad, P.A.; Scacheri, P.C.; Harte, P.J. Histone demethylase UTX and chromatin remodeler BRM bind directly to CBP and modulate acetylation of histone H3 lysine 27. Mol. Cell. Biol. 2012, 32, 2323–2334. [Google Scholar] [CrossRef]
- Wang, S.P.; Tang, Z.; Chen, C.W.; Shimada, M.; Koche, R.P.; Wang, L.H.; Nakadai, T.; Chramiec, A.; Krivtsov, A.V.; Armstrong, S.A.; et al. A UTX-MLL4-p300 Transcriptional Regulatory Network Coordinately Shapes Active Enhancer Landscapes for Eliciting Transcription. Mol. Cell 2017, 67, 308–321.e6. [Google Scholar] [CrossRef]
- Wang, C.; Lee, J.E.; Cho, Y.W.; Xiao, Y.; Jin, Q.; Liu, C.; Ge, K. UTX regulates mesoderm differentiation of embryonic stem cells independent of H3K27 demethylase activity. Proc. Natl. Acad. Sci. USA 2012, 109, 15324–15329. [Google Scholar] [CrossRef] [PubMed]
- Greenfield, A.; Carrel, L.; Pennisi, D.; Philippe, C.; Quaderi, N.; Siggers, P.; Steiner, K.; Tam, P.P.; Monaco, A.P.; Willard, H.F.; et al. The UTX gene escapes X inactivation in mice and humans. Hum. Mol. Genet. 1998, 7, 737–742. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Deng, X.; Watkins, R.; Disteche, C.M. Sex-specific differences in expression of histone demethylases Utx and Uty in mouse brain and neurons. J. Neurosci. 2008, 28, 4521–4527. [Google Scholar] [CrossRef] [PubMed]
- Armoskus, C.; Moreira, D.; Bollinger, K.; Jimenez, O.; Taniguchi, S.; Tsai, H.W. Identification of sexually dimorphic genes in the neonatal mouse cortex and hippocampus. Brain Res. 2014, 1562, 23–38. [Google Scholar] [CrossRef]
- Yang, X.; Xu, B.; Mulvey, B.; Evans, M.; Jordan, S.; Wang, Y.D.; Pagala, V.; Peng, J.; Fan, Y.; Patel, A.; et al. Differentiation of human pluripotent stem cells into neurons or cortical organoids requires transcriptional co-regulation by UTX and 53BP1. Nat. Neurosci. 2019, 22, 362–373. [Google Scholar] [CrossRef] [PubMed]
- Shan, Y.; Zhang, Y.; Zhao, Y.; Wang, T.; Zhang, J.; Yao, J.; Ma, N.; Liang, Z.; Huang, W.; Huang, K.; et al. JMJD3 and UTX determine fidelity and lineage specification of human neural progenitor cells. Nat. Commun. 2020, 11, 382. [Google Scholar] [CrossRef]
- Subhramanyam, C.S.; Cao, Q.; Wang, C.; Heng, Z.S.L.; Zhou, Z.; Hu, Q. Role of PIWI-like 4 in modulating neuronal differentiation from human embryonal carcinoma cells. RNA Biol. 2020, 17, 1613–1624. [Google Scholar] [CrossRef]
- Tang, Q.Y.; Zhang, S.F.; Dai, S.K.; Liu, C.; Wang, Y.Y.; Du, H.Z.; Teng, Z.Q.; Liu, C.M. UTX Regulates Human Neural Differentiation and Dendritic Morphology by Resolving Bivalent Promoters. Stem Cell Rep. 2020, 15, 439–453. [Google Scholar] [CrossRef]
- Agger, K.; Cloos, P.A.; Christensen, J.; Pasini, D.; Rose, S.; Rappsilber, J.; Issaeva, I.; Canaani, E.; Salcini, A.E.; Helin, K. UTX and JMJD3 are histone H3K27 demethylases involved in HOX gene regulation and development. Nature 2007, 449, 731–734. [Google Scholar] [CrossRef]
- Lan, F.; Bayliss, P.E.; Rinn, J.L.; Whetstine, J.R.; Wang, J.K.; Chen, S.; Iwase, S.; Alpatov, R.; Issaeva, I.; Canaani, E.; et al. A histone H3 lysine 27 demethylase regulates animal posterior development. Nature 2007, 449, 689–694. [Google Scholar] [CrossRef]
- Shahhoseini, M.; Taghizadeh, Z.; Hatami, M.; Baharvand, H. Retinoic acid dependent histone 3 demethylation of the clustered HOX genes during neural differentiation of human embryonic stem cells. Biochem. Cell Biol. 2013, 91, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Welstead, G.G.; Creyghton, M.P.; Bilodeau, S.; Cheng, A.W.; Markoulaki, S.; Young, R.A.; Jaenisch, R. X-linked H3K27me3 demethylase Utx is required for embryonic development in a sex-specific manner. Proc. Natl. Acad. Sci. USA 2012, 109, 13004–13009. [Google Scholar] [CrossRef]
- Lei, X.; Jiao, J. UTX Affects Neural Stem Cell Proliferation and Differentiation through PTEN Signaling. Stem Cell Rep. 2018, 10, 1193–1207. [Google Scholar] [CrossRef] [PubMed]
- Tang, G.B.; Zeng, Y.Q.; Liu, P.P.; Mi, T.W.; Zhang, S.F.; Dai, S.K.; Tang, Q.Y.; Yang, L.; Xu, Y.J.; Yan, H.L.; et al. The Histone H3K27 Demethylase UTX Regulates Synaptic Plasticity and Cognitive Behaviors in Mice. Front. Mol. Neurosci. 2017, 10, 267. [Google Scholar] [CrossRef] [PubMed]
- Cabrera Zapata, L.E.; Cisternas, C.D.; Sosa, C.; Garcia-Segura, L.M.; Arevalo, M.A.; Cambiasso, M.J. X-linked histone H3K27 demethylase Kdm6a regulates sexually dimorphic differentiation of hypothalamic neurons. Cell. Mol. Life Sci. 2021, 78, 7043–7060. [Google Scholar] [CrossRef]
- Cisternas, C.D.; Cabrera Zapata, L.E.; Mir, F.R.; Scerbo, M.J.; Arevalo, M.A.; Garcia-Segura, L.M.; Cambiasso, M.J. Estradiol-dependent axogenesis and Ngn3 expression are determined by XY sex chromosome complement in hypothalamic neurons. Sci. Rep. 2020, 10, 8223. [Google Scholar] [CrossRef] [PubMed]
- Scerbo, M.J.; Freire-Regatillo, A.; Cisternas, C.D.; Brunotto, M.; Arevalo, M.A.; Garcia-Segura, L.M.; Cambiasso, M.J. Neurogenin 3 mediates sex chromosome effects on the generation of sex differences in hypothalamic neuronal development. Front. Cell. Neurosci. 2014, 8, 188. [Google Scholar] [CrossRef]
- Cabrera Zapata, L.E.; Cambiasso, M.J.; Arevalo, M.A. Epigenetic modifier Kdm6a/Utx controls the specification of hypothalamic neuronal subtypes in a sex-dependent manner. Front. Cell Dev. Biol. 2022, 10, 937875. [Google Scholar] [CrossRef]
- Pelling, M.; Anthwal, N.; McNay, D.; Gradwohl, G.; Leiter, A.B.; Guillemot, F.; Ang, S.L. Differential requirements for neurogenin 3 in the development of POMC and NPY neurons in the hypothalamus. Dev. Biol. 2011, 349, 406–416. [Google Scholar] [CrossRef]
- Anthwal, N.; Pelling, M.; Claxton, S.; Mellitzer, G.; Collin, C.; Kessaris, N.; Richardson, W.D.; Gradwohl, G.; Ang, S.L. Conditional deletion of neurogenin-3 using Nkx2.1iCre results in a mouse model for the central control of feeding, activity and obesity. Dis. Models Mech. 2013, 6, 1133–1145. [Google Scholar] [CrossRef]
- Miyake, N.; Mizuno, S.; Okamoto, N.; Ohashi, H.; Shiina, M.; Ogata, K.; Tsurusaki, Y.; Nakashima, M.; Saitsu, H.; Niikawa, N.; et al. KDM6A point mutations cause Kabuki syndrome. Hum. Mutat. 2013, 34, 108–110. [Google Scholar] [CrossRef] [PubMed]
- Van Laarhoven, P.M.; Neitzel, L.R.; Quintana, A.M.; Geiger, E.A.; Zackai, E.H.; Clouthier, D.E.; Artinger, K.B.; Ming, J.E.; Shaikh, T.H. Kabuki syndrome genes KMT2D and KDM6A: Functional analyses demonstrate critical roles in craniofacial, heart and brain development. Hum. Mol. Genet. 2015, 24, 4443–4453. [Google Scholar] [CrossRef] [PubMed]
- Bogershausen, N.; Gatinois, V.; Riehmer, V.; Kayserili, H.; Becker, J.; Thoenes, M.; Simsek-Kiper, P.O.; Barat-Houari, M.; Elcioglu, N.H.; Wieczorek, D.; et al. Mutation Update for Kabuki Syndrome Genes KMT2D and KDM6A and Further Delineation of X-Linked Kabuki Syndrome Subtype 2. Hum. Mutat. 2016, 37, 847–864. [Google Scholar] [CrossRef]
- Faundes, V.; Goh, S.; Akilapa, R.; Bezuidenhout, H.; Bjornsson, H.T.; Bradley, L.; Brady, A.F.; Brischoux-Boucher, E.; Brunner, H.; Bulk, S.; et al. Clinical delineation, sex differences, and genotype-phenotype correlation in pathogenic KDM6A variants causing X-linked Kabuki syndrome type 2. Genet. Med. 2021, 23, 1202–1210. [Google Scholar] [CrossRef]
- Dubal, D.B. Sex difference in Alzheimer’s disease: An updated, balanced and emerging perspective on differing vulnerabilities. Handb. Clin. Neurol. 2020, 175, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Buckley, R.F.; Mormino, E.C.; Amariglio, R.E.; Properzi, M.J.; Rabin, J.S.; Lim, Y.Y.; Papp, K.V.; Jacobs, H.I.L.; Burnham, S.; Hanseeuw, B.J.; et al. Sex, amyloid, and APOE epsilon4 and risk of cognitive decline in preclinical Alzheimer’s disease: Findings from three well-characterized cohorts. Alzheimers Dement. 2018, 14, 1193–1203. [Google Scholar] [CrossRef] [PubMed]
- Casaletto, K.B.; Elahi, F.M.; Staffaroni, A.M.; Walters, S.; Contreras, W.R.; Wolf, A.; Dubal, D.; Miller, B.; Yaffe, K.; Kramer, J.H. Cognitive aging is not created equally: Differentiating unique cognitive phenotypes in “normal” adults. Neurobiol. Aging 2019, 77, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Jack, C.R., Jr.; Wiste, H.J.; Weigand, S.D.; Knopman, D.S.; Vemuri, P.; Mielke, M.M.; Lowe, V.; Senjem, M.L.; Gunter, J.L.; Machulda, M.M.; et al. Age, Sex, and APOE epsilon4 Effects on Memory, Brain Structure, and beta-Amyloid Across the Adult Life Span. JAMA Neurol. 2015, 72, 511–519. [Google Scholar] [CrossRef]
- Jack, C.R., Jr.; Therneau, T.M.; Wiste, H.J.; Weigand, S.D.; Knopman, D.S.; Lowe, V.J.; Mielke, M.M.; Vemuri, P.; Roberts, R.O.; Machulda, M.M.; et al. Transition rates between amyloid and neurodegeneration biomarker states and to dementia: A population-based, longitudinal cohort study. Lancet Neurol. 2016, 15, 56–64. [Google Scholar] [CrossRef]
- Jack, C.R., Jr.; Wiste, H.J.; Weigand, S.D.; Therneau, T.M.; Knopman, D.S.; Lowe, V.; Vemuri, P.; Mielke, M.M.; Roberts, R.O.; Machulda, M.M.; et al. Age-specific and sex-specific prevalence of cerebral beta-amyloidosis, tauopathy, and neurodegeneration in cognitively unimpaired individuals aged 50–95 years: A cross-sectional study. Lancet Neurol. 2017, 16, 435–444. [Google Scholar] [CrossRef]
- Iwase, S.; Lan, F.; Bayliss, P.; de la Torre-Ubieta, L.; Huarte, M.; Qi, H.H.; Whetstine, J.R.; Bonni, A.; Roberts, T.M.; Shi, Y. The X-linked mental retardation gene SMCX/JARID1C defines a family of histone H3 lysine 4 demethylases. Cell 2007, 128, 1077–1088. [Google Scholar] [CrossRef] [PubMed]
- Christensen, J.; Agger, K.; Cloos, P.A.; Pasini, D.; Rose, S.; Sennels, L.; Rappsilber, J.; Hansen, K.H.; Salcini, A.E.; Helin, K. RBP2 belongs to a family of demethylases, specific for tri-and dimethylated lysine 4 on histone 3. Cell 2007, 128, 1063–1076. [Google Scholar] [CrossRef] [PubMed]
- Qi, S.; Ngwa, C.; Al Mamun, A.; Romana, S.; Wu, T.; Marrelli, S.P.; Arnold, A.P.; McCullough, L.D.; Liu, F. X, but not Y, Chromosomal Complement Contributes to Stroke Sensitivity in Aged Animals. Transl. Stroke Res. 2022. [Google Scholar] [CrossRef]
- Qi, S.; Al Mamun, A.; Ngwa, C.; Romana, S.; Ritzel, R.; Arnold, A.P.; McCullough, L.D.; Liu, F. X chromosome escapee genes are involved in ischemic sexual dimorphism through epigenetic modification of inflammatory signals. J. Neuroinflamm. 2021, 18, 70. [Google Scholar] [CrossRef]
- Lisabeth, L.; Bushnell, C. Stroke risk in women: The role of menopause and hormone therapy. Lancet Neurol. 2012, 11, 82–91. [Google Scholar] [CrossRef]
- Itoh, Y.; Golden, L.C.; Itoh, N.; Matsukawa, M.A.; Ren, E.; Tse, V.; Arnold, A.P.; Voskuhl, R.R. The X-linked histone demethylase Kdm6a in CD4+ T lymphocytes modulates autoimmunity. J. Clin. Investig. 2019, 129, 3852–3863. [Google Scholar] [CrossRef] [PubMed]
- Outchkourov, N.S.; Muino, J.M.; Kaufmann, K.; van Ijcken, W.F.; Groot Koerkamp, M.J.; van Leenen, D.; de Graaf, P.; Holstege, F.C.; Grosveld, F.G.; Timmers, H.T. Balancing of histone H3K4 methylation states by the Kdm5c/SMCX histone demethylase modulates promoter and enhancer function. Cell Rep. 2013, 3, 1071–1079. [Google Scholar] [CrossRef]
- Wei, G.; Deng, X.; Agarwal, S.; Iwase, S.; Disteche, C.; Xu, J. Patient Mutations of the Intellectual Disability Gene KDM5C Downregulate Netrin G2 and Suppress Neurite Growth in Neuro2a Cells. J. Mol. Neurosci. 2016, 60, 33–45. [Google Scholar] [CrossRef]
- Swahari, V.; West, A.E. Histone demethylases in neuronal differentiation, plasticity, and disease. Curr. Opin. Neurobiol. 2019, 59, 9–15. [Google Scholar] [CrossRef]
- Hatch, H.A.M.; Secombe, J. Molecular and cellular events linking variants in the histone demethylase KDM5C to the intellectual disability disorder Claes-Jensen syndrome. FEBS J. 2021. [Google Scholar] [CrossRef]
- Vallianatos, C.N.; Raines, B.; Porter, R.S.; Bonefas, K.M.; Wu, M.C.; Garay, P.M.; Collette, K.M.; Seo, Y.A.; Dou, Y.; Keegan, C.E.; et al. Mutually suppressive roles of KMT2A and KDM5C in behaviour, neuronal structure, and histone H3K4 methylation. Commun. Biol. 2020, 3, 278. [Google Scholar] [CrossRef] [PubMed]
- Jensen, L.R.; Amende, M.; Gurok, U.; Moser, B.; Gimmel, V.; Tzschach, A.; Janecke, A.R.; Tariverdian, G.; Chelly, J.; Fryns, J.P.; et al. Mutations in the JARID1C gene, which is involved in transcriptional regulation and chromatin remodeling, cause X-linked mental retardation. Am. J. Hum. Genet. 2005, 76, 227–236. [Google Scholar] [CrossRef]
- Wu, P.M.; Yu, W.H.; Chiang, C.W.; Wu, C.Y.; Chen, J.S.; Tu, Y.F. Novel Variations in the KDM5C Gene Causing X-Linked Intellectual Disability. Neurol. Genet. 2022, 8, e646. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, T.F.; Goncalves, A.P.; Fintelman Rodrigues, N.; dos Santos, J.M.; Pimentel, M.M.; Santos-Reboucas, C.B. KDM5C mutational screening among males with intellectual disability suggestive of X-Linked inheritance and review of the literature. Eur. J. Med. Genet. 2014, 57, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Abdelhaleem, M. RNA helicases: Regulators of differentiation. Clin. Biochem. 2005, 38, 499–503. [Google Scholar] [CrossRef]
- Garbelli, A.; Beermann, S.; Di Cicco, G.; Dietrich, U.; Maga, G. A motif unique to the human DEAD-box protein DDX3 is important for nucleic acid binding, ATP hydrolysis, RNA/DNA unwinding and HIV-1 replication. PLoS ONE 2011, 6, e19810. [Google Scholar] [CrossRef]
- Cruciat, C.M.; Dolde, C.; de Groot, R.E.; Ohkawara, B.; Reinhard, C.; Korswagen, H.C.; Niehrs, C. RNA helicase DDX3 is a regulatory subunit of casein kinase 1 in Wnt-beta-catenin signaling. Science 2013, 339, 1436–1441. [Google Scholar] [CrossRef]
- Tang, L.; Levy, T.; Guillory, S.; Halpern, D.; Zweifach, J.; Giserman-Kiss, I.; Foss-Feig, J.H.; Frank, Y.; Lozano, R.; Belani, P.; et al. Prospective and detailed behavioral phenotyping in DDX3X syndrome. Mol. Autism 2021, 12, 36. [Google Scholar] [CrossRef] [PubMed]
- Ruzzo, E.K.; Perez-Cano, L.; Jung, J.Y.; Wang, L.K.; Kashef-Haghighi, D.; Hartl, C.; Singh, C.; Xu, J.; Hoekstra, J.N.; Leventhal, O.; et al. Inherited and De Novo Genetic Risk for Autism Impacts Shared Networks. Cell 2019, 178, 850–866.e26. [Google Scholar] [CrossRef]
- Kellaris, G.; Khan, K.; Baig, S.M.; Tsai, I.C.; Zamora, F.M.; Ruggieri, P.; Natowicz, M.R.; Katsanis, N. A hypomorphic inherited pathogenic variant in DDX3X causes male intellectual disability with additional neurodevelopmental and neurodegenerative features. Hum. Genom. 2018, 12, 11. [Google Scholar] [CrossRef]
- Nicola, P.; Blackburn, P.R.; Rasmussen, K.J.; Bertsch, N.L.; Klee, E.W.; Hasadsri, L.; Pichurin, P.N.; Rankin, J.; Raymond, F.L.; Study, D.D.D.; et al. De novo DDX3X missense variants in males appear viable and contribute to syndromic intellectual disability. Am. J. Med. Genet. A 2019, 179, 570–578. [Google Scholar] [CrossRef] [PubMed]
- Hoye, M.L.; Calviello, L.; Poff, A.J.; Ejimogu, N.E.; Newman, C.R.; Montgomery, M.D.; Ou, J.; Floor, S.N.; Silver, D.L. Aberrant cortical development is driven by impaired cell cycle and translational control in a DDX3X syndrome model. eLife 2022, 11, e78203. [Google Scholar] [CrossRef] [PubMed]
- Patmore, D.M.; Jassim, A.; Nathan, E.; Gilbertson, R.J.; Tahan, D.; Hoffmann, N.; Tong, Y.; Smith, K.S.; Kanneganti, T.D.; Suzuki, H.; et al. DDX3X Suppresses the Susceptibility of Hindbrain Lineages to Medulloblastoma. Dev. Cell 2020, 54, 455–470.e5. [Google Scholar] [CrossRef] [PubMed]
- Szappanos, D.; Tschismarov, R.; Perlot, T.; Westermayer, S.; Fischer, K.; Platanitis, E.; Kallinger, F.; Novatchkova, M.; Lassnig, C.; Muller, M.; et al. The RNA helicase DDX3X is an essential mediator of innate antimicrobial immunity. PLoS Pathog. 2018, 14, e1007397. [Google Scholar] [CrossRef]
- Lennox, A.L.; Hoye, M.L.; Jiang, R.; Johnson-Kerner, B.L.; Suit, L.A.; Venkataramanan, S.; Sheehan, C.J.; Alsina, F.C.; Fregeau, B.; Aldinger, K.A.; et al. Pathogenic DDX3X Mutations Impair RNA Metabolism and Neurogenesis during Fetal Cortical Development. Neuron 2020, 106, 404–420.e8. [Google Scholar] [CrossRef]
- Bellott, D.W.; Skaletsky, H.; Pyntikova, T.; Mardis, E.R.; Graves, T.; Kremitzki, C.; Brown, L.G.; Rozen, S.; Warren, W.C.; Wilson, R.K.; et al. Convergent evolution of chicken Z and human X chromosomes by expansion and gene acquisition. Nature 2010, 466, 612–616. [Google Scholar] [CrossRef] [PubMed]
- Skaletsky, H.; Kuroda-Kawaguchi, T.; Minx, P.J.; Cordum, H.S.; Hillier, L.; Brown, L.G.; Repping, S.; Pyntikova, T.; Ali, J.; Bieri, T.; et al. The male-specific region of the human Y chromosome is a mosaic of discrete sequence classes. Nature 2003, 423, 825–837. [Google Scholar] [CrossRef]
- Mueller, J.L.; Skaletsky, H.; Brown, L.G.; Zaghlul, S.; Rock, S.; Graves, T.; Auger, K.; Warren, W.C.; Wilson, R.K.; Page, D.C. Independent specialization of the human and mouse X chromosomes for the male germ line. Nat. Genet. 2013, 45, 1083–1087. [Google Scholar] [CrossRef]
- Ross, J.L.; Bloy, L.; Roberts, T.P.L.; Miller, J.; Xing, C.; Silverman, L.A.; Zinn, A.R. Y chromosome gene copy number and lack of autism phenotype in a male with an isodicentric Y chromosome and absent NLGN4Y expression. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2019, 180, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Maier, M.C.; McInerney, M.A.; Graves, J.A.M.; Charchar, F.J. Noncoding Genes on Sex Chromosomes and Their Function in Sex Determination, Dosage Compensation, Male Traits, and Diseases. Sex Dev. 2021, 15, 432–440. [Google Scholar] [CrossRef]
- Johansson, M.M.; Pottmeier, P.; Suciu, P.; Ahmad, T.; Zaghlool, A.; Halvardson, J.; Darj, E.; Feuk, L.; Peuckert, C.; Jazin, E. Novel Y-Chromosome Long Non-Coding RNAs Expressed in Human Male CNS During Early Development. Front. Genet. 2019, 10, 891. [Google Scholar] [CrossRef] [PubMed]
- De Vries, G.J. Minireview: Sex differences in adult and developing brains: Compensation, compensation, compensation. Endocrinology 2004, 145, 1063–1068. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, M.M.; Arnold, A.P. Reframing sexual differentiation of the brain. Nat. Neurosci. 2011, 14, 677–683. [Google Scholar] [CrossRef] [PubMed]
- Arnold, A.P. Rethinking sex determination of non-gonadal tissues. Curr. Top. Dev. Biol. 2019, 134, 289–315. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Chr | Gene (GO Database) | ||
|---|---|---|---|
| Symbol | Name | Function | |
| Y | Sry | sex determining region of Chr Y | DNA-binding transcription factor activity |
| Kdm6c-Uty | histone demethylase UTY | Histone demethylase activity | |
| Kdm5d | lysine (K)-specific demethylase 5D | Histone demethylase activity | |
| Ddx3y | DEAD box helicase 3, Y-linked | ATP-dependent RNA helicase dbp3 | |
| Usp9y | ubiquitin specific peptidase 9, Y chromosome | Cysteine-type deubiquitinase activity | |
| Eif2s3y | eukaryotic translation initiation factor 2, subunit 3, structural gene Y-linked | Eukaryotic translation initiation factor 2 subunit 3 family member | |
| Nlgn4y | neuroligin-4, Y-linked | Cell adhesion molecule | |
| X | Kdm6a-Utx | lysine (K)-specific demethylase 6A | Histone demethylase activity |
| Kdm5c | lysine (K)-specific demethylase 5C | Lysine-specific demethylase | |
| Ddx3x | DEAD box helicase 3, X-linked | ATP-dependent RNA helicase dbp3 | |
| Usp9x | ubiquitin specific peptidase 9, X chromosome | Ubiquitin carboxyl-terminal hydrolase | |
| Eif2s3x | eukaryotic translation initiation factor 2, subunit 3, structural gene X-linked | Eukaryotic translation initiation factor 2 subunit 3 family member | |
| Mecp2 | methyl CpG binding protein 2 | DNA binding | |
| Hdac8 | histone deacetylase 8 | Histone deacetylase activity | |
| Morf4l2 | mortality factor 4 like 2 | Histone acetylation | |
| Msl3 | MSL3 like 2 | Histone acetylation | |
| Phf6 | PHD finger protein 6 | DNA metabolism protein | |
| Fmr1 | fragile X messenger ribonucleoprotein 1 | Regulation of alternative mRNA splicing | |
| Mid1 | midline 1 | E3 ubiquitin-protein ligase trim36-related | |
| Ogt | O-linked N-acetylglucosamine (GlcNAc) transferase (UDP-N-acetylglucosamine:polypeptide-N-acetylglucosaminyl transferase) | DNA binding | |
| Syp | synaptophysin | Lipid binding | |
| Tmem47 | transmembrane protein 47 | Plasma membrane | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cabrera Zapata, L.E.; Garcia-Segura, L.M.; Cambiasso, M.J.; Arevalo, M.A. Genetics and Epigenetics of the X and Y Chromosomes in the Sexual Differentiation of the Brain. Int. J. Mol. Sci. 2022, 23, 12288. https://doi.org/10.3390/ijms232012288
Cabrera Zapata LE, Garcia-Segura LM, Cambiasso MJ, Arevalo MA. Genetics and Epigenetics of the X and Y Chromosomes in the Sexual Differentiation of the Brain. International Journal of Molecular Sciences. 2022; 23(20):12288. https://doi.org/10.3390/ijms232012288
Chicago/Turabian StyleCabrera Zapata, Lucas E., Luis Miguel Garcia-Segura, María Julia Cambiasso, and Maria Angeles Arevalo. 2022. "Genetics and Epigenetics of the X and Y Chromosomes in the Sexual Differentiation of the Brain" International Journal of Molecular Sciences 23, no. 20: 12288. https://doi.org/10.3390/ijms232012288
APA StyleCabrera Zapata, L. E., Garcia-Segura, L. M., Cambiasso, M. J., & Arevalo, M. A. (2022). Genetics and Epigenetics of the X and Y Chromosomes in the Sexual Differentiation of the Brain. International Journal of Molecular Sciences, 23(20), 12288. https://doi.org/10.3390/ijms232012288