

Peroxisome Proliferator FpPEX11 Is Involved in the Development and Pathogenicity in Fusarium pseudograminearum


Abstract
1. Introduction
2. Results
2.1. Identification and Knockout of FpPEX11 in F. pseudograminearum
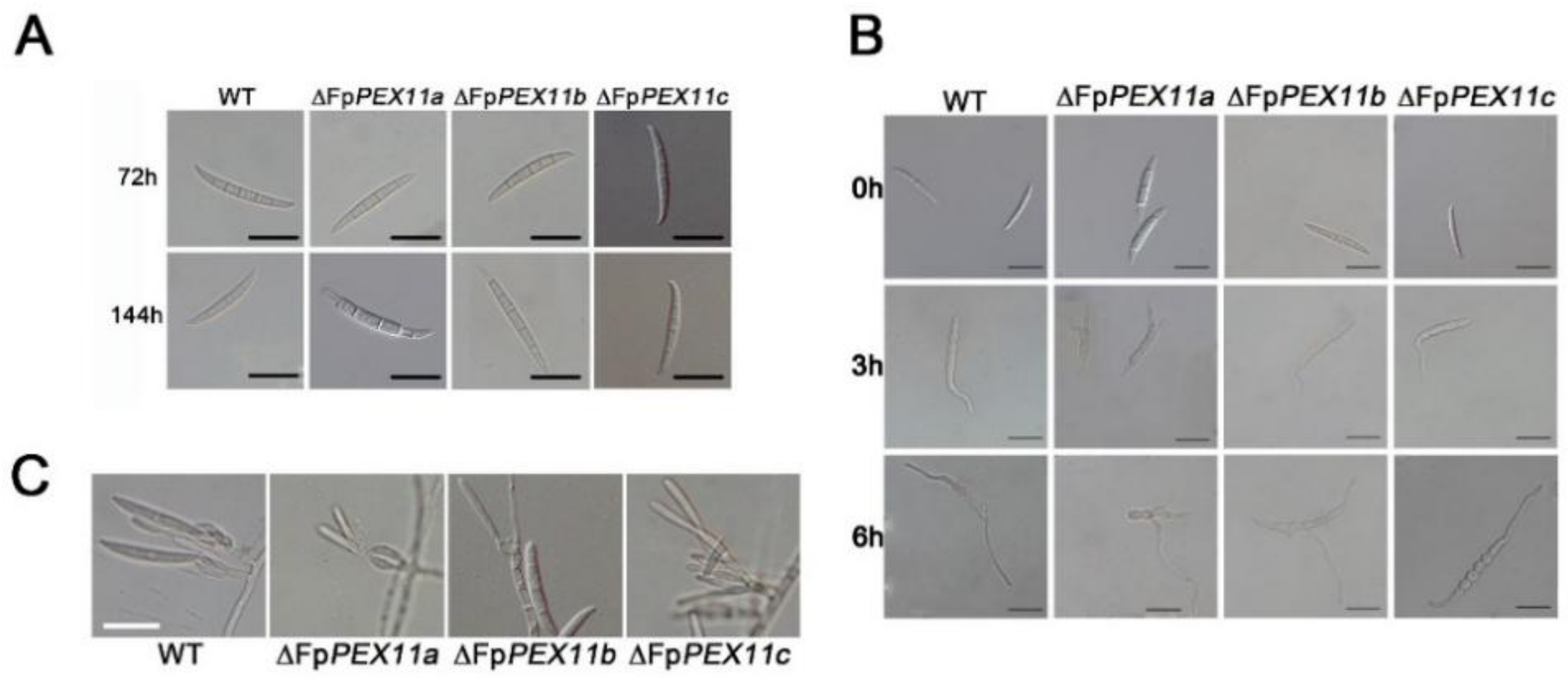
2.2. FpPEX11 Is Involved in Vegetative Growth and Asexual Reproduction
2.3. FpPEX11 Regulates ROS Metabolism
2.4. FpPEX11 Is Involved in Lipid Metabolism
2.5. FpPEX11 Is Involved in Responding to Cell Membrane and Cell Wall Stresses
2.6. The ∆FpPEX11 Exhibits Abnormal Peroxisome Number
2.7. FpPEX11 Regulates Expression of Peroxisome-Proliferator-Associated Gene
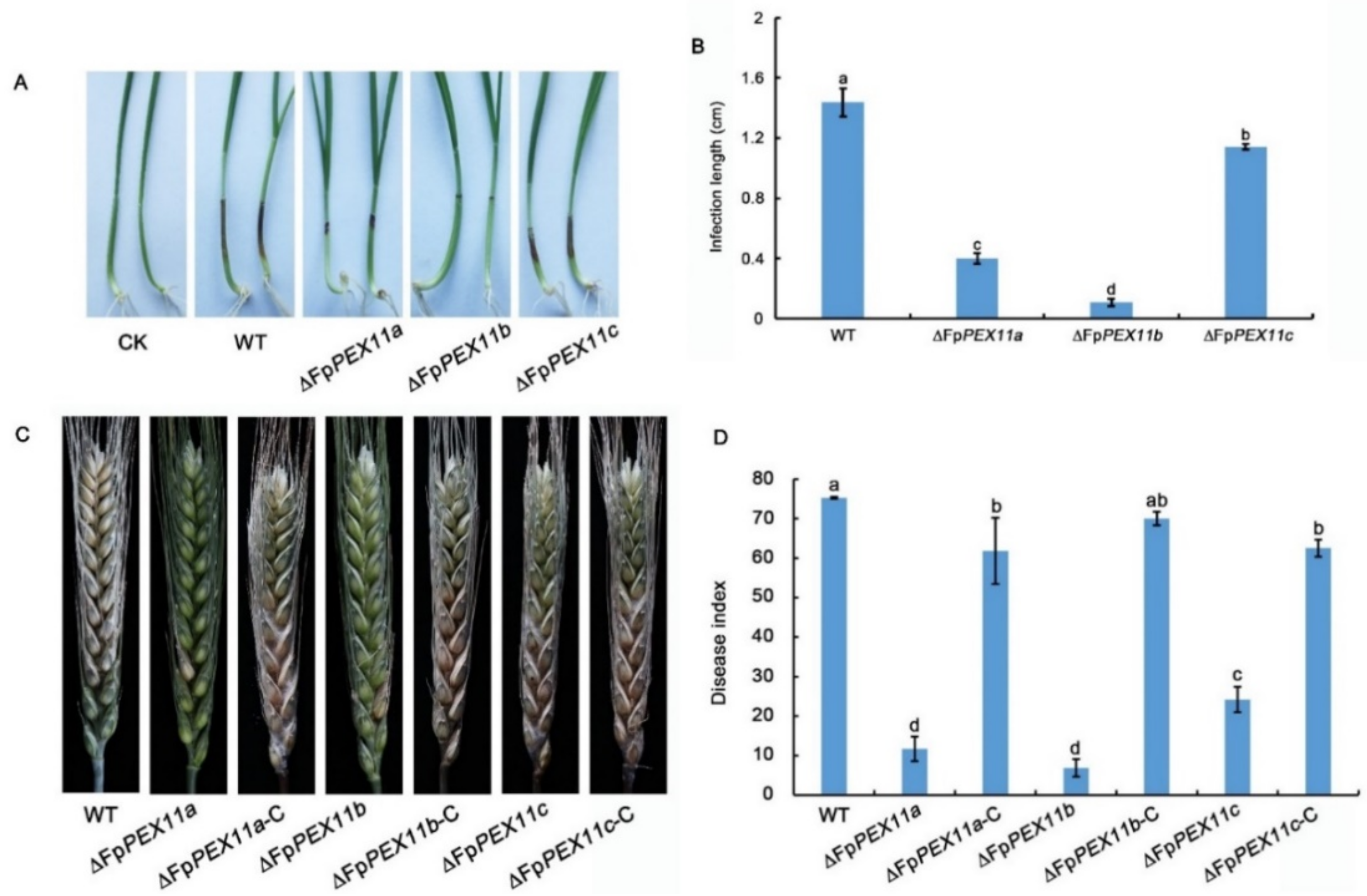
2.8. FpPEX11 Is Essential for the Pathogenicity of F. pseudograminearum
3. Discussion
4. Materials and Methods
4.1. Fungal Strain and Culture Medium
4.2. Sequence Analysis
4.3. Strain Construction
4.4. Vegetative Growth Detection
4.5. ROS Detection
4.6. Fluorescence Microscopy
4.7. Relative Quantitative Real-Time PCR
4.8. Pathogenicity Test
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| DON | Deoxynivalenol |
| FCR | Fusarium crown rot |
| ER | Endoplasmic reticulum |
| Reactive oxygen species | |
| ZS | Zellweger syndrome |
| WT | Wild type |
| PDA | Potato dextrose agar |
| CM | Complete medium |
| YEPD | Yeast extract peptone dextrose |
| SDS | Sodium dodecyl sulfate |
References
- Hogg, A.C.; Johnston, R.H.; Johnston, J.A.; Klouser, L.; Kephart, K.D.; Dyer, A.T. Monitoring fusarium crown rot populations in spring wheat residues using quantitative real-time polymerase chain reaction. Phytopathology 2010, 100, 149–157. [Google Scholar] [CrossRef]
- Smiley, R.W.; Gourlie, J.A.; Easley, S.A.; Patterson, L.-M.; Whittaker, R.G. Crop Damage Estimates for Crown Rot of Wheat and Barley in the Pacific Northwest. Plant Dis. 2005, 89, 595–604. [Google Scholar] [CrossRef]
- Burgess, L.W.; Backhouse, D.; Summerell, B.A.; Swan, L.J. Crown Rot of Wheat. In Fusarium: Paul E. Nelson Memorial Symposium; The American Phytopathological Society: St Paul, MN, USA, 2001. [Google Scholar]
- Knight, N.L.; Sutherland, M.W. Histopathological Assessment of Fusarium pseudograminearum Colonization of Cereal Culms During Crown Rot Infections. Plant Dis. 2016, 100, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Obanor, F.; Neate, S.; Simpfendorfer, S.; Sabburg, R.; Wilson, P.; Chakraborty, S. Fusarium graminearumandFusarium pseudograminearumcaused the 2010 head blight epidemics in Australia. Plant Pathol. 2013, 62, 79–91. [Google Scholar] [CrossRef]
- Chen, C.; Payne, W.A.; Smiley, R.W.; Stoltz, M.A. Yield and water-use efficiency of eight wheat cultivars planted on seven dates in Northeastern Oregon. Agron. J. 2003, 95, 836. [Google Scholar] [CrossRef]
- Papendick, R.I.; Cook, R.J. Plant water stress and development of Fusarium foot rot in wheat subjected to different cultural practices. Phytopathology 1974, 64, 358–363. [Google Scholar] [CrossRef]
- Paulitz, T.C. Low Input No-till Cereal Production in the Pacific Northwest of the U.S.: The Challenges of Root Diseases. Eur. J. Plant Pathol. 2006, 115, 271–281. [Google Scholar] [CrossRef]
- Al-Fahdawi, M.S.; Able, J.A.; Evans, M.; Able, A.J. Response of durum wheat to different levels of zinc and Fusarium pseudograminearum. Crop. Pasture Sci. 2014, 65, 61–73. [Google Scholar] [CrossRef]
- Veenhuis, M.; Salomons, F.A.; Klei, I.J.V.D. Peroxisome biogenesis and degradation in yeast a structurefunction analysis. Microsc. Res. Tech. 2000, 15, 584–600. [Google Scholar] [CrossRef]
- Fransen, M. Peroxisome Dynamics: Molecular Players, Mechanisms, and (Dys)functions. ISRN Cell. Biol. 2012, 2012, 1–24. [Google Scholar] [CrossRef]
- Fagarasanu, A.; Mast, F.D.; Knoblach, B.; Rachubinski, R.A. Molecular mechanisms of organelle inheritance: Lessons from peroxisomes in yeast. Nat. Rev. Mol. Cell. Biol. 2010, 11, 644–654. [Google Scholar] [CrossRef] [PubMed]
- Nagotu, S.; Veenhuis, M.; van der Klei, I.J. Divide et impera: The dictum of peroxisomes. Traffic 2010, 11, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Huber, A.; Koch, J.; Kragler, F.; Brocard, C.; Hartig, A. A subtle interplay between three Pex11 proteins shapes de novo formation and fission of peroxisomes. Traffic 2012, 13, 157–167. [Google Scholar] [CrossRef]
- Kiel, J.A.; van den Berg, M.; Bovenberg, R.A.; van der Klei, I.J.; Veenhuis, M. Penicillium chrysogenum Pex5p mediates differential sorting of PTS1 proteins to microbodies of the methylotrophic yeast Hansenula polymorpha. Fungal. Genet. Biol. 2004, 41, 708–720. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zheng, S.; Ju, Z.; Zhang, C.; Tang, G.; Wang, J.; Wen, Z.; Chen, W.; Ma, Z. Contribution of peroxisomal docking machinery to mycotoxin biosynthesis, pathogenicity and pexophagy in the plant pathogenic fungus Fusarium graminearum. Environ. Microbiol. 2018, 20, 3224–3245. [Google Scholar] [CrossRef]
- Fransen, M.; Nordgren, M.; Wang, B.; Apanasets, O. Role of peroxisomes in ROS/RNS-metabolism: Implications for human disease. Biochim. Biophys. Acta 2012, 1822, 1363–1373. [Google Scholar] [CrossRef]
- Van Veldhoven, P.P. Biochemistry and genetics of inherited disorders of peroxisomal fatty acid metabolism. J. Lipid. Res. 2010, 51, 2863–2895. [Google Scholar] [CrossRef]
- Waterham, H.R.; Ferdinandusse, S.; Wanders, R.J. Human disorders of peroxisome metabolism and biogenesis. Biochim. Biophys. Acta 2016, 1863, 922–933. [Google Scholar] [CrossRef]
- Cipolla, C.M.; Lodhi, I.J. Peroxisomal Dysfunction in Age-Related Diseases. Trends Endocrinol. Metab. 2017, 28, 297–308. [Google Scholar] [CrossRef]
- Hu, J.; Aguirre, M.; Peto, C.; Alonso, J.; Ecker, J.; Chory, J. A Role for Peroxisomes in Photomorphogenesis and Development of Arabidopsis. Science 2002, 297, 405–409. [Google Scholar] [CrossRef]
- Hu, J.; Baker, A.; Bartel, B.; Linka, N.; Mullen, R.T.; Reumann, S.; Zolman, B.K. Plant peroxisomes: Biogenesis and function. Plant Cell. 2012, 24, 2279–2303. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, L.A. Peroxisomes as a cellular source of reactive nitrogen species signal molecules. Arch. Biochem. Biophys. 2011, 506, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sparkes, I.A.; Brandizzi, F.; Slocombe, S.P.; El-Shami, M.; Hawes, C.; Baker, A. An Arabidopsis pex10 null mutant is embryo lethal, implicating peroxisomes in an essential role during plant embryogenesis. Plant Physiol. 2003, 133, 1809–1819. [Google Scholar] [CrossRef]
- Shabab, M. Role of plant peroxisomes in protection against herbivores. Subcell Biochem. 2013, 69, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Kresge, N.; Simoni, R.D.; Hill, R.L. The discovery of glyoxysomes: The work of Harry Beevers, 1961. J. Biol. Chem. 2010, 285, e6–e7. [Google Scholar] [CrossRef] [PubMed]
- Marshall, P.A.; Krimkevich, Y.I.; Lark, R.H.; Dyer, J.M.; Veenhuis, M.; Goodman, J.M. Pmp27 promotes peroxisomal proliferation. J. Cell Biol. 1995, 129, 345–355. [Google Scholar] [CrossRef]
- Orth, T.; Reumann, S.; Zhang, X.; Fan, J.; Wenzel, D.; Quan, S.; Hu, J. The PEROXIN11 protein family controls peroxisome proliferation in Arabidopsis. Plant Cell 2007, 19, 333–350. [Google Scholar] [CrossRef] [PubMed]
- Koch, J.; Pranjic, K.; Huber, A.; Ellinger, A.; Hartig, A.; Kragler, F.; Brocard, C. PEX11 family members are membrane elongation factors that coordinate peroxisome proliferation and maintenance. J. Cell Sci. 2010, 123, 3389–3400. [Google Scholar] [CrossRef]
- Weng, H.; Ji, X.; Naito, Y.; Endo, K.; Ma, X.; Takahashi, R.; Shen, C.; Hirokawa, G.; Fukushima, Y.; Iwai, N. Pex11alpha deficiency impairs peroxisome elongation and division and contributes to nonalcoholic fatty liver in mice. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E187–E196. [Google Scholar] [CrossRef]
- Wang, J.; Li, L.; Zhang, Z.; Qiu, H.; Li, D.; Fang, Y.; Jiang, H.; Chai, R.Y.; Mao, X.; Wang, Y.; et al. One of Three Pex11 Family Members Is Required for Peroxisomal Proliferation and Full Virulence of the Rice Blast Fungus Magnaporthe oryzae. PLoS ONE 2015, 10, e0134249. [Google Scholar] [CrossRef]
- Tam, Y.Y.; Torres-Guzman, J.C.; Vizeacoumar, F.J.; Smith, J.J.; Marelli, M.; Aitchison, J.D.; Rachubinski, R.A. Pex11-related proteins in peroxisome dynamics: A role for the novel peroxin Pex27p in controlling peroxisome size and number in Saccharomyces cerevisiae. Mol. Biol. Cell 2003, 14, 4089–4102. [Google Scholar] [CrossRef] [PubMed]
- Rottensteiner, H.; Stein, K.; Sonnenhol, E.; Erdmann, R. Conserved function of pex11p and the novel pex25p and pex27p in peroxisome biogenesis. Mol. Biol. Cell 2003, 14, 4316–4328. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Latruffe, N.; Vamecq, J.; Cherkaoui Malki, M. Genetic-dependency of peroxisomal cell functions-emerging aspects. J. Cell Mol. Med. 2003, 7, 238–248. [Google Scholar] [CrossRef] [PubMed]
- Ram, A.F.; Klis, F.M. Identification of fungal cell wall mutants using susceptibility assays based on Calcofluor white and Congo red. Nat. Protoc. 2006, 1, 2253–2256. [Google Scholar] [CrossRef]
- Lazarow, P.B.; Fujiki, Y. Biogenesis of peroxisomes. Annu. Rev. Cell Biol. 1985, 1, 489–530. [Google Scholar] [CrossRef]
- Hettema, E.H.; Motley, A.M. How peroxisomes multiply. J. Cell Sci. 2009, 122, 2331–2336. [Google Scholar] [CrossRef]
- Snyder, W.B.; Koller, A.; Choy, A.J.; Subramani, S. The peroxin Pex19p interacts with multiple, integral membrane proteins at the peroxisomal membrane. J. Cell Biol. 2000, 149, 1171–1178. [Google Scholar] [CrossRef]
- William, B.S.; Faber, K.N.; Thibaut, J.W.; Koller, A.; Luers, G.H.; Rangell, L.; Keller, G.A.; Subramani, S. Pex19p interacts with Pex3p and Pex10p and is essential for peroxisome biogenesis in Pichia pastoris. Mol. Biol. Cell 1999, 10, 1745–1761. [Google Scholar] [CrossRef][Green Version]
- Cichocki, B.A.; Krumpe, K.; Vitali, D.G.; Rapaport, D. Pex19 is involved in importing dually targeted tail-anchored proteins to both mitochondria and peroxisomes. Traffic 2018, 19, 770–785. [Google Scholar] [CrossRef]
- Koch, J.; Brocard, C. PEX11 proteins attract Mff and human Fis1 to coordinate peroxisomal fission. J. Cell Sci. 2012, 125, 3813–3826. [Google Scholar] [CrossRef]
- Williams, C.; Opalinski, L.; Landgraf, C.; Costello, J.; Schrader, M.; Krikken, A.M.; Knoops, K.; Kram, A.M.; Volkmer, R.; van der Klei, I.J. The membrane remodeling protein Pex11p activates the GTPase Dnm1p during peroxisomal fission. Proc. Natl. Acad. Sci. USA 2015, 112, 6377–6382. [Google Scholar] [CrossRef] [PubMed]
- Rottensteiner, H.; Kal, A.J.; Filipits, M.; Binder, M.; Hamilton, B.; Tabak, H.F.; Ruis, H. Pip2p_ a transcriptional regulator of peroxisome proliferation in the yeast Saccharomyces cerevisiae. EMBO J. 1996, 15, 2924–2934. [Google Scholar] [CrossRef] [PubMed]
- Tower, R.J.; Fagarasanu, A.; Aitchison, J.D.; Rachubinski, R.A. The peroxin Pex34p functions with the Pex11 family of peroxisomal divisional proteins to regulate the peroxisome population in yeast. Mol. Biol. Cell 2011, 22, 1727–1738. [Google Scholar] [CrossRef] [PubMed]
- Kimura, A.; Takano, Y.; Furusawa, I.; Okuno, T.; Notes, A. Peroxisomal Metabolic Function Is Required for Appressorium-Mediated Plant Infection by Colletotrichum lagenarium. Plant Cell 2001, 13, 1945–1957. [Google Scholar] [CrossRef]
- Bowman, S.M.; Free, S.J. The structure and synthesis of the fungal cell wall. Bioessays 2006, 28, 799–808. [Google Scholar] [CrossRef]
- Li, L.; Wang, J.; Zhang, Z.; Wang, Y.; Liu, M.; Jiang, H.; Chai, R.; Mao, X.; Qiu, H.; Liu, F.; et al. MoPex19, which is essential for maintenance of peroxisomal structure and woronin bodies, is required for metabolism and development in the rice blast fungus. PLoS ONE 2014, 9, e85252. [Google Scholar] [CrossRef]
- Wanders, R.J.; Waterham, H.R. Biochemistry of mammalian peroxisomes revisited. Annu. Rev. Biochem. 2006, 75, 295–332. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, J.; Zhang, Z.; Wang, Y.; Li, L.; Chai, R.; Mao, X.; Jiang, H.; Qiu, H.; Du, X.; et al. PTS1 Peroxisomal Import Pathway Plays Shared and Distinct Roles to PTS2 Pathway in Development and Pathogenicity of Magnaporthe oryzae. PLoS ONE 2013, 8, e55554. [Google Scholar] [CrossRef]
- Bonekamp, N.A.; Volkl, A.; Fahimi, H.D.; Schrader, M. Reactive oxygen species and peroxisomes: Struggling for balance. Biofactors 2009, 35, 346–355. [Google Scholar] [CrossRef]
- Ma, H.; Wang, M.; Gai, Y.; Fu, H.; Zhang, B.; Ruan, R.; Chung, K.-R.; Li, H. Thioredoxin and Glutaredoxin Systems Required for Oxidative Stress Resistance, Fungicide Sensitivity, and Virulence of Alternaria alternata. Appl. Environ. Microbiol. 2018, 84, e00086-18. [Google Scholar] [CrossRef]
- Mudge, A.M.; Dill-Macky, R.; Dong, Y.; Gardiner, D.M.; White, R.G.; Manners, J.M. A role for the mycotoxin deoxynivalenol in stem colonisation during crown rot disease of wheat caused by Fusarium graminearum and Fusarium pseudograminearum. Physiol. Mol. Plant Pathol. 2006, 69, 73–85. [Google Scholar] [CrossRef]
- Wang, L.-m.; Zhang, Y.-f.; Du, Z.-l.; Kang, R.-j.; Chen, L.-l.; Xing, X.-p.; Yuan, H.; Ding, S.-l.; Li, H.-l. FpPDE1 function of Fsarium pseudograminearum on pathogenesis in wheat. J. Integr. Agric. 2017, 16, 2504–2512. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, C.; Wang, L.; Sun, S.; Liu, A.; Liang, Y.; Yu, J.; Dong, H. FgPEX1 and FgPEX10 are required for the maintenance of Woronin bodies and full virulence of Fusarium graminearum. Curr. Genet 2019, 65, 1383–1396. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Geng, X.; Ma, Y.; Zhao, J.; Li, T.; Ding, S.; Shi, Y.; Li, H. Identification of basic helix-loop-helix transcription factors reveals candidate genes involved in pathogenicity of Fusarium pseudograminearum. Can. J. Plant Pathol. 2019, 41, 200–208. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, L.; Liang, Y.; Yu, J. FgPEX4 is involved in development, pathogenicity, and cell wall integrity in Fusarium graminearum. Curr. Genet. 2019, 65, 747–758. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, L.; Liang, S.; Zhang, P.; Kang, R.; Zhang, M.; Wang, M.; Chen, L.; Yuan, H.; Ding, S.; et al. FpDep1, a component of Rpd3L histone deacetylase complex, is important for vegetative development, ROS accumulation, and pathogenesis in Fusarium pseudograminearum. Fungal. Genet. Biol. 2020, 135, 103299. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Rubella, S.; Goswami, H.C.K. Pathogenicity and In Planta Mycotoxin Accumulation Among Members of the Fusarium graminearum Species Complex on Wheat and Rice. Phytopathology 2005, 95, 1397–1404. [Google Scholar] [CrossRef]
- Tang, L.; Yu, X.; Zhang, L.; Zhang, L.; Chen, L.; Zou, S.; Liang, Y.; Yu, J.; Dong, H. Mitochondrial FgEch1 is responsible for conidiation and full virulence in Fusarium graminearum. Curr. Genet. 2020, 66, 361–371. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Conidiation (106 Conidia/mL) | Germination (%) | |
|---|---|---|---|
| 3 h | 6 h | ||
| WT | 2.59 ± 0.15 a | 51.6 ± 0.75 a | 100.00 ± 0.00 a |
| ∆FpPEX11a | 1.21 ± 0.10 b | 27.2 ± 3.32 b | 97.00 ± 3.00 a |
| ∆FpPEX11b | 0.75 ± 0.09 b | 49.2 ± 1.16 a | 95.00 ± 1.00 a |
| ∆FpPEX11c | 2.37 ± 0.35 a | 52.8 ± 1.59 a | 99.00 ± 1.00 a |
| DON Production | Relative Expression Levels of TRI | ||||
|---|---|---|---|---|---|
| ELISA (μg/mg) | HPLC (μg/g) * | TRI5 | TRI6 | TRI10 | |
| WT | 133.78 ± 18.36 a | 14.59 ± 3.47 b | 1.00 ± 0.09 | 1.00 ± 0.22 | 1.00 ± 0.22 |
| ∆FpPEX11a | 0 | 0.16 ± 0.01 c | 0.11 ± 0.04 | 0.16 ± 0.06 | 0.19 ± 0.02 |
| ∆FpPEX11b | 0 | 0.00763 ± 0 c | 0.13 ± 0.02 | 0.10 ± 0.02 | 0.15 ± 0.08 |
| ∆FpPEX11c | 121.43 ± 21.65 a | 32.05 ± 6.00 a | 7.198 ± 0.09 | 1.82 ± 0.24 | 0.90 ± 0.21 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Xu, H.; Liu, C.; Tao, Y.; Wang, X.; Liang, Y.; Zhang, L.; Yu, J. Peroxisome Proliferator FpPEX11 Is Involved in the Development and Pathogenicity in Fusarium pseudograminearum. Int. J. Mol. Sci. 2022, 23, 12184. https://doi.org/10.3390/ijms232012184
Wang M, Xu H, Liu C, Tao Y, Wang X, Liang Y, Zhang L, Yu J. Peroxisome Proliferator FpPEX11 Is Involved in the Development and Pathogenicity in Fusarium pseudograminearum. International Journal of Molecular Sciences. 2022; 23(20):12184. https://doi.org/10.3390/ijms232012184
Chicago/Turabian StyleWang, Mingyu, Hao Xu, Chunjie Liu, Yilin Tao, Xiaofeng Wang, Yuancun Liang, Li Zhang, and Jinfeng Yu. 2022. "Peroxisome Proliferator FpPEX11 Is Involved in the Development and Pathogenicity in Fusarium pseudograminearum" International Journal of Molecular Sciences 23, no. 20: 12184. https://doi.org/10.3390/ijms232012184
APA StyleWang, M., Xu, H., Liu, C., Tao, Y., Wang, X., Liang, Y., Zhang, L., & Yu, J. (2022). Peroxisome Proliferator FpPEX11 Is Involved in the Development and Pathogenicity in Fusarium pseudograminearum. International Journal of Molecular Sciences, 23(20), 12184. https://doi.org/10.3390/ijms232012184