
Biphasic Functions of Sodium Fluoride (NaF) in Soft and in Hard Periodontal Tissues

 , ,
, ,  , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
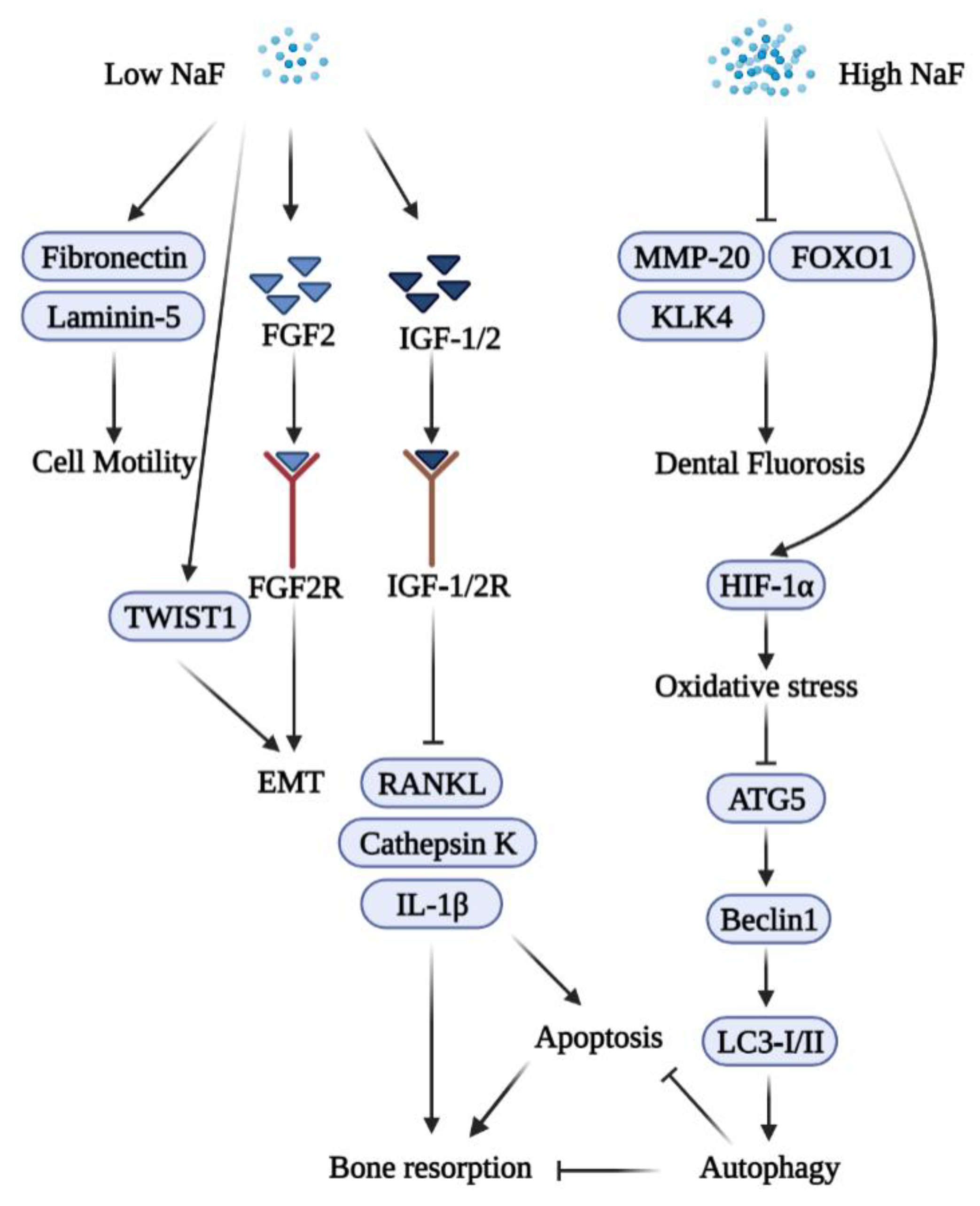
2. Fluoride and Epithelial Cells
2.1. High Concentration Fluoride and Epithelial Cells
2.2. Low Concentration Fluoride and Epithelial Cells
2.3. Fluoride and Epithelial–Mesenchymal Interactions
3. Fluoride and Bone Marrow Mesenchymal Stem Cells (BMMSCs)
4. Fluoride and Bone Metabolism
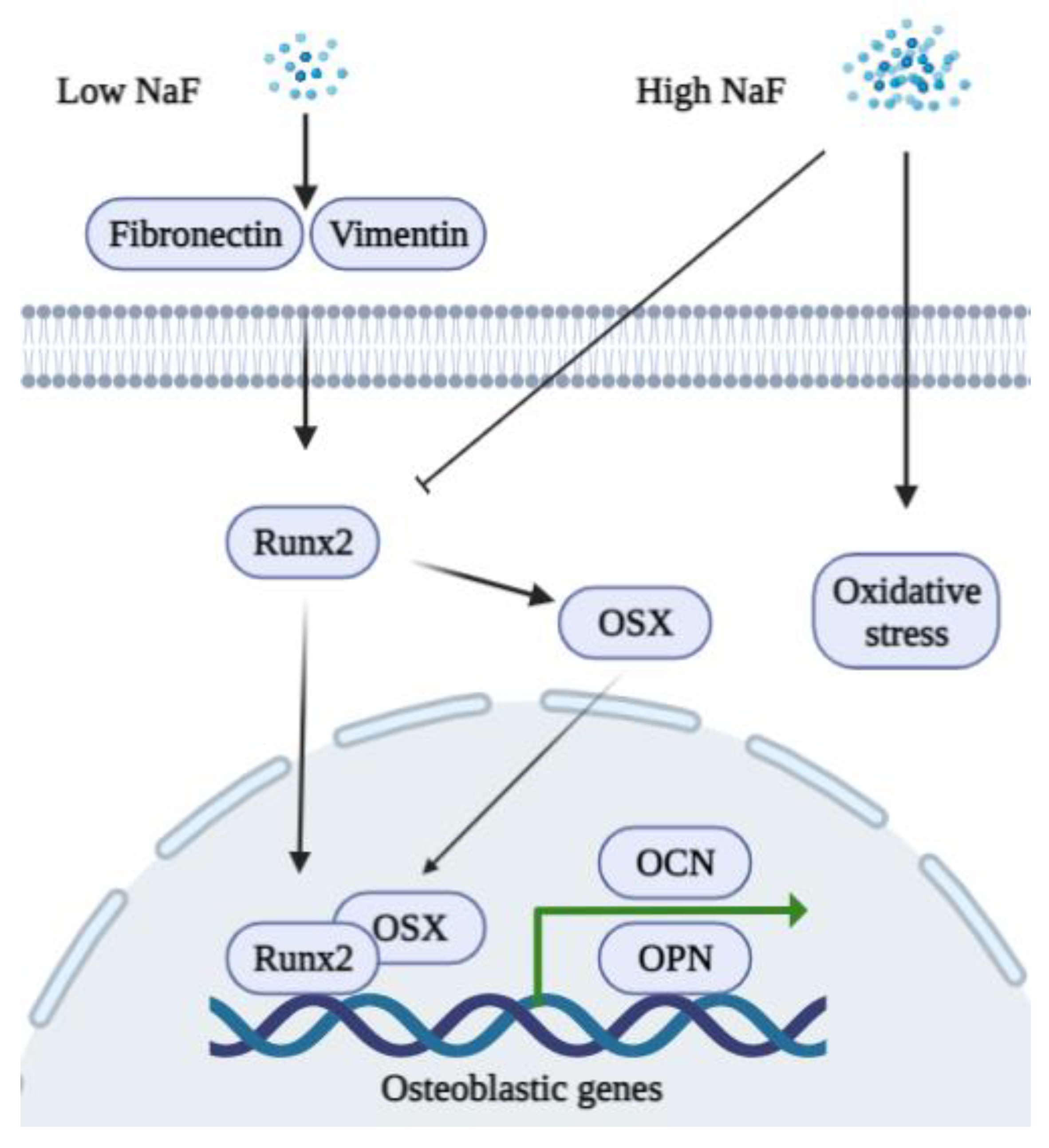
4.1. Fluoride and Osteoblasts
4.2. Fluoride and Osteoclasts
5. Fluoride and Periodontal Diseases
6. Fluoride and microRNA
7. Clinical Applications
8. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Whelton, H.; Spencer, A.; Do, L.; Rugg-Gunn, A. Fluoride Revolution and Dental Caries: Evolution of Policies for Global Use. J. Dent. Res. 2019, 98, 837–846. [Google Scholar] [CrossRef] [PubMed]
- Cate, J.T.; Buzalaf, M. Fluoride Mode of Action: Once There Was an Observant Dentist. J. Dent. Res. 2019, 98, 725–730. [Google Scholar] [CrossRef] [PubMed]
- DenBesten, P.K. Biological mechanisms of dental fluorosis relevant to the use of fluoride supplements. Community Dent. Oral Epidemiol. 1999, 27, 41–47. [Google Scholar] [CrossRef]
- Whitford, G.M. The metabolism and toxicity of fluoride. Monogr. Oral Sci. 1996, 16, 1–153. [Google Scholar]
- Gupta, A.R.; Dey, S.; Saini, M.; Swarup, D. Toxic effect of sodium fluoride on hydroxyproline level and expression of collagen-1 gene in rat bone and its amelioration by Tamrindus indica L. fruit pulp extract. Interdiscip. Toxicol. 2016, 9, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, D.A.; Yujra, V.Q.; Da Silva, V.H.P.; Claudio, S.R.; Estadella, D.; Viana, M.D.B.; Oshima, C.T.F. Putative mechanisms of genotoxicity induced by fluoride: A comprehensive review. Environ. Sci. Pollut. Res. 2017, 24, 15254–15259. [Google Scholar] [CrossRef]
- Shenoy, P.S.; Sen, U.; Kapoor, S.; Ranade, A.V.; Chowdhury, C.R.; Bose, B. Sodium fluoride induced skeletal muscle changes: Degradation of proteins and signaling mechanism. Environ. Pollut. 2018, 244, 534–548. [Google Scholar] [CrossRef] [PubMed]
- Nakade, O.; Koyama, H.; Arai, J.; Ariji, H.; Takada, J.; Kaku, T. Stimulation by low concentrations of fluoride of the proliferation and alkaline phosphatase activity of human dental pulp cells in vitro. Arch. Oral Biol. 1999, 44, 89–92. [Google Scholar] [CrossRef]
- Boivin, G.; Chavassieux, P.; Chapuy, M.; Baud, C.; Meunier, P. Skeletal fluorosis: Histomorphometric analysis of bone changes and bone fluoride content in 29 patients. Bone 1989, 10, 89–99. [Google Scholar] [CrossRef]
- Zipkin, I.; Bernick, S.; Menczel, J.A. Morphological study of the effect of fluoride on the periodontium of the hydrocorti-sone-treated rat. Periodontics 1965, 3, 111–114. [Google Scholar] [CrossRef]
- Levy, B.M.; Dreizen, S.; Bernick, S.; Hampton, J.K. Studies on the Biology of the Periodontium of Marmosets: IX. Effect of Parathyroid Hormone on the Alveolar Bone of Marmosets Pretreated with Fluoridated and Nonfluoridated Drinking Water. J. Dent. Res. 1970, 49, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Ammann, P.; Rizzoli, R.; Caverzasio, J.; Bonjour, J.-P. Fluoride Potentiates the Osteogenic Effects of IGF-I in Aged Ovariectomized Rats. Bone 1998, 22, 39–43. [Google Scholar] [CrossRef]
- Wang, S.-S.; Tang, Y.-L.; Pang, X.; Zheng, M.; Liang, X.-H. The maintenance of an oral epithelial barrier. Life Sci. 2019, 227, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Kawano, S.; Saito, M.; Handa, K.; Morotomi, T.; Toyono, T.; Seta, Y.; Nakamura, N.; Uchida, T.; Toyoshima, K.; Ohishi, M.; et al. Characterization of Dental Epithelial Progenitor Cells Derived from Cervical-loop Epithelium in a Rat Lower Incisor. J. Dent. Res. 2004, 83, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Crivelini, M.M.; Oliveira, D.T.; De Mesquita, R.A.; De Sousa, S.C.O.M.; Loyola, A.M. Kallikrein 4 and matrix metalloproteinase-20 immunoexpression in malignant, benign and infiltrative odontogenic tumors. J. Oral Maxillofac. Pathol. 2016, 20, 246–251. [Google Scholar] [CrossRef]
- Mu, Y.; Tian, R.; Xiao, L.; Sun, D.; Zhang, Z.; Xu, S.; Yang, G. Molecular Evolution of Tooth-Related Genes Provides New Insights into Dietary Adaptations of Mammals. J. Mol. Evol. 2021, 89, 458–471. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Gao, Y.; Wang, C.; Zhao, L.; Sun, D. Excessive fluoride induces endoplasmic reticulum stress and interferes enamel proteinases secretion. Environ. Toxicol. 2011, 28, 332–341. [Google Scholar] [CrossRef] [PubMed]
- DenBesten, P.K.; Zhu, L.; Li, W.; Tanimoto, K.; Liu, H.; Witkowska, H.E. Fluoride incorporation into apatite crystals delays amelogenin hydrolysis. Eur. J. Oral Sci. 2011, 119, 3–7. [Google Scholar] [CrossRef][Green Version]
- DenBesten, P.K.; Yan, Y.; Featherstone, J.D.B.; Hilton, J.F.; Smith, C.E.; Li, W. Effects of fluoride on rat dental enamel matrix proteinases. Arch. Oral Biol. 2002, 47, 763–770. [Google Scholar] [CrossRef]
- Sharma, R.; Tsuchiya, M.; Skobe, Z.; Tannous, B.A.; Bartlett, J.D. The Acid Test of Fluoride: How pH Modulates Toxicity. PLoS ONE 2010, 5, e10895. [Google Scholar] [CrossRef]
- Gao, J.; Ruan, J.; Gao, L. Excessive fluoride reducesFoxo1expression in dental epithelial cells of the rat incisor. Eur. J. Oral Sci. 2014, 122, 317–323. [Google Scholar] [CrossRef]
- Zuckerbraun, H.L.; Babich, H.; May, R.; Sinensky, M.C. Triclosan, cytotoxicity, mode of action, and induction of apoptosis in human gingival cells in vitro. Eur. J. Oral Sci. 1998, 106, 628–636. [Google Scholar] [CrossRef]
- Yan, Q.; Zhang, Y.; Li, W.; DenBesten, P. Micromolar Fluoride Alters Ameloblast Lineage Cells in vitro. J. Dent. Res. 2007, 86, 336–340. [Google Scholar] [CrossRef]
- Jorgensen, S.N.; Sanders, J.R. Mathematical models of wound healing and closure: A comprehensive review. Med. Biol. Eng. Comput. 2015, 54, 1297–1316. [Google Scholar] [CrossRef] [PubMed]
- Arakawa, Y.; Bhawal, U.K.; Ikoma, T.; Kimoto, K.; Kuroha, K.; Kubota, T.; Hamada, N.; Kubota, E.; Arakawa, H. Low concentration fluoride stimulates cell motility of epithelial cells in vitro. Biomed. Res. 2009, 30, 271–277. [Google Scholar] [CrossRef]
- Wixler, V. The role of FHL2 in wound healing and inflammation. FASEB J. 2019, 33, 7799–7809. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.-D.; Kim, K.-H.; Lee, Y.-M.; Ku, Y.; Seol, Y.-J. Periodontal Wound Healing and Tissue Regeneration: A Narrative Review. Pharmaceuticals 2021, 14, 456. [Google Scholar] [CrossRef] [PubMed]
- Mossahebi-Mohammadi, M.; Quan, M.; Zhang, J.-S.; Li, X. FGF Signaling Pathway: A Key Regulator of Stem Cell Pluripotency. Front. Cell Dev. Biol. 2020, 8, 79. [Google Scholar] [CrossRef]
- El-Baz, L.M.; Shoukry, N.M.; Hafez, H.S.; Guzy, R.D.; Salem, M.L. Fibroblast Growth Factor 2 Augments Transforming Growth Factor Beta 1 Induced Epithelial-mesenchymal Transition in Lung Cell Culture Model. Iran. J. Allergy Asthma Immunol. 2020, 19, 348–361. [Google Scholar] [CrossRef]
- Kinoshita-Ise, M.; Tsukashima, A.; Kinoshita, T.; Yamazaki, Y.; Ohyama, M. Altered FGF expression profile in human scalp-derived fibroblasts upon WNT activation: Implication of their role to provide folliculogenetic microenvironment. Inflamm. Regen. 2020, 40, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Chen, S.; Han, J.-X.; Qian, B.; Wang, X.-R.; Zhong, W.-L.; Qin, Y.; Zhang, H.; Gao, W.-F.; Lei, Y.-Y.; et al. Twist1 Regulates Vimentin through Cul2 Circular RNA to Promote EMT in Hepatocellular Carcinoma. Cancer Res. 2018, 78, 4150–4162. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Bhawal, U.K.; Hamada, N.; Kuboyama, N.; Abiko, Y.; Arakawa, H. Low Level Fluoride Stimulates Epithelial-Mesenchymal Interaction in Oral Mucosa. J. Hard Tissue Biol. 2013, 22, 59–66. [Google Scholar] [CrossRef][Green Version]
- Chen, F.; Han, Y.; Kang, Y. Bone marrow niches in the regulation of bone metastasis. Br. J. Cancer 2021, 214, 1912–1920. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Li, X.; Liu, Y.; Gao, X.; Zhu, T.; Lu, L. Acceleration of Bone Regeneration in Critical-Size Defect Using BMP-9-Loaded nHA/ColI/MWCNTs Scaffolds Seeded with Bone Marrow Mesenchymal Stem Cells. BioMed Res. Int. 2019, 2019, 7343957. [Google Scholar] [CrossRef]
- Bhawal, U.K.; Li, X.; Suzuki, M.; Taguchi, C.; Oka, S.; Arikawa, K.; Tewari, N.; Liu, Y. Treatment with low-level sodium fluoride on wound healing and the osteogenic differentiation of bone marrow mesenchymal stem cells. Dent. Traumatol. 2019, 36, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Garcia, A.L.; Picinini, J.; Silveira, M.D.; Camassola, M.; Visentim, A.P.; Salvador, M.; da Silva, J. Fluorosilicic acid induces DNA damage and oxidative stress in bone marrow mesenchymal stem cells. Mutat. Res. Toxicol. Environ. Mutagen. 2020, 861–862, 503297. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Xie, F.-N.; Dong, P.; Li, Q.-C.; Yu, G.-Y.; Xiao, R. High-Dose Fluoride Impairs the Properties of Human Embryonic Stem Cells via JNK Signaling. PLoS ONE 2016, 11, e0148819. [Google Scholar] [CrossRef]
- Xu, S.; Xie, X.; Li, C.; Liu, Z.; Zuo, D. Micromolar sodium fluoride promotes osteo/odontogenic differentiation in dental pulp stem cells by inhibiting PI3K/AKT pathway. Arch. Oral Biol. 2021, 131, 105265. [Google Scholar] [CrossRef]
- Pan, Y.; Li, Z.; Wang, Y.; Yan, M.; Wu, J.; Beharee, R.G.; Yu, J. Sodium fluoride regulates the osteo/odontogenic differentiation of stem cells from apical papilla by modulating autophagy. J. Cell. Physiol. 2019, 234, 16114–16124. [Google Scholar] [CrossRef]
- Ngoc, T.D.N.; Son, Y.-O.; Lim, S.-S.; Shi, X.; Kim, J.-G.; Heo, J.S.; Choe, Y.; Jeon, Y.-M.; Lee, J.-C. Sodium fluoride induces apoptosis in mouse embryonic stem cells through ROS-dependent and caspase- and JNK-mediated pathways. Toxicol. Appl. Pharmacol. 2012, 259, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, J.A.; Partridge, N.C. Physiological Bone Remodeling: Systemic Regulation and Growth Factor Involvement. Physiology 2016, 31, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, Z.; Duan, N.; Zhu, G.; Schwarz, E.M.; Xie, C. Osteoblast–osteoclast interactions. Connect. Tissue Res. 2017, 59, 99–107. [Google Scholar] [CrossRef]
- Sims, N.A.; Martin, T.J. Osteoclasts Provide Coupling Signals to Osteoblast Lineage Cells Through Multiple Mechanisms. Annu. Rev. Physiol. 2020, 82, 507–529. [Google Scholar] [CrossRef] [PubMed]
- Antonarakis, G.S.; Moseley, R.; Waddington, R.J. Differential influence of fluoride concentration on the synthesis of bone matrix glycoproteins within mineralizing bone cellsin vitro. Acta Odontol. Scand. 2014, 72, 1066–1069. [Google Scholar] [CrossRef] [PubMed]
- Simon, M.J.K.; Beil, F.T.; Riedel, C.; Lau, G.; Tomsia, A.; Zimmermann, E.A.; Koehne, T.; Ueblacker, P.; Rüther, W.; Pogoda, P.; et al. Deterioration of teeth and alveolar bone loss due to chronic environmental high-level fluoride and low calcium exposure. Clin. Oral Investig. 2016, 20, 2361–2370. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zhou, H.; Liu, H.; Ji, H.; Fei, W.; Luo, E. Fluorine-contained hydroxyapatite suppresses bone resorption through inhibiting osteoclasts differentiation and function in vitro and in vivo. Cell Prolif. 2019, 52, e12613. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M. Fluoride and bone metabolism. Clin. Calcium 2007, 17, 217–223. [Google Scholar]
- Kurdi, M.S. Chronic fluorosis: The disease and its anaesthetic implications. Indian J. Anaesth. 2016, 60, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Qiao, W.; Liu, Q.; Li, Z.; Zhang, H.; Chen, Z. Changes in physicochemical and biological properties of porcine bone derived hydroxyapatite induced by the incorporation of fluoride. Sci. Technol. Adv. Mater. 2017, 18, 110–121. [Google Scholar] [CrossRef]
- Epsley, S.; Tadros, S.; Farid, A.; Kargilis, D.; Mehta, S.; Rajapakse, C.S. The Effect of Inflammation on Bone. Front. Physiol. 2021, 11. [Google Scholar] [CrossRef]
- Zoch, M.L.; Clemens, T.L.; Riddle, R.C. New insights into the biology of osteocalcin. Bone 2015, 82, 42–49. [Google Scholar] [CrossRef]
- Icer, M.A.; Gezmen-Karadag, M. The multiple functions and mechanisms of osteopontin. Clin. Biochem. 2018, 59, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.-Y.; Wu, M.; Zhao, D.; Edwards, D.; McVicar, A.; Luo, Y.; Zhu, G.; Wang, Y.; Zhou, H.-D.; Chen, W.; et al. Runx1 is a central regulator of osteogenesis for bone homeostasis by orchestrating BMP and WNT signaling pathways. PLoS Genet. 2021, 17, e1009233. [Google Scholar] [CrossRef]
- Takahata, Y.; Hagino, H.; Kimura, A.; Urushizaki, M.; Kobayashi, S.; Wakamori, K.; Fujiwara, C.; Nakamura, E.; Yu, K.; Kiyonari, H.; et al. Smoc1 and Smoc2 regulate bone formation as downstream molecules of Runx2. Commun. Biol. 2021, 4, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Iaquinta, M.R.; Lanzillotti, C.; Mazziotta, C.; Bononi, I.; Frontini, F.; Mazzoni, E.; Oton-Gonzalez, L.; Rotondo, J.C.; Torreggiani, E.; Tognon, M.; et al. The role of microRNAs in the osteogenic and chondrogenic differentiation of mesenchymal stem cells and bone pathologies. Theranostics 2021, 11, 6573–6591. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.-Q.; Zhao, Z.-T.; Zhang, X.-Y.; Wang, Y.; Wang, H.; Liu, D.-W.; Li, G.-S.; Jing, L. Fluoride Affects Calcium Homeostasis and Osteogenic Transcription Factor Expressions Through L-type Calcium Channels in Osteoblast Cell Line. Biol. Trace Element Res. 2014, 162, 219–226. [Google Scholar] [CrossRef]
- Lee, M.; Arikawa, K.; Nagahama, F. Micromolar Levels of Sodium Fluoride Promote Osteoblast Differentiation Through Runx2 Signaling. Biol. Trace Element Res. 2017, 178, 283–291. [Google Scholar] [CrossRef]
- Volobaev, V.P.; Serdyukova, E.S.; Kalyuzhnaya, E.E.; Schetnikova, E.A.; Korotkova, A.D.; Naik, A.A.; Bach, S.N.; Prosekov, A.Y.; Larionov, A.V. Investigation of the genotoxic effects of fluoride on a bone tissue model. Toxicol. Res. 2020, 36, 337–342. [Google Scholar] [CrossRef]
- Burgener, D.; Bonjour, J.-P.; Caverzasio, J. Fluoride increases tyrosine kinase activity in osteoblast-like cells: Regulatory role for the stimulation of cell proliferation and Pi transport across the plasma membrane. J. Bone Miner. Res. 2009, 10, 164–171. [Google Scholar] [CrossRef]
- Farley, J.R.; Wergedal, J.E.; Baylink, D.J. Fluoride Directly Stimulates Proliferation and Alkaline Phosphatase Activity of Bone-Forming Cells. Science 1983, 222, 330–332. [Google Scholar] [CrossRef]
- Pak, C.Y.; Zerwekh, J.E.; Antich, P. Anabolic effects of fluoride on bone. Trends Endocrinol. Metab. 1995, 6, 229–234. [Google Scholar] [CrossRef]
- Rochette, L.; Meloux, A.; Rigal, E.; Zeller, M.; Cottin, Y.; Vergely, C. The Role of Osteoprotegerin and Its Ligands in Vascular Function. Int. J. Mol. Sci. 2019, 20, 705. [Google Scholar] [CrossRef]
- Yang, S.; Wang, Z.; Farquharson, C.; Alkasir, R.; Zahra, M.; Ren, G.; Han, B. Sodium fluoride induces apoptosis and alters bcl-2 family protein expression in MC3T3-E1 osteoblastic cells. Biochem. Biophys. Res. Commun. 2011, 410, 910–915. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.-P.; Wang, H.-W.; Liu, J.; Zhang, Z.-H.; Zhu, S.-Q.; Zhou, B.-H. Mitochondrial respiratory chain complex abnormal expressions and fusion disorder are involved in fluoride-induced mitochondrial dysfunction in ovarian granulosa cells. Chemosphere 2018, 215, 619–625. [Google Scholar] [CrossRef]
- Wu, S.; Yan, W.; Qiu, B.; Liao, Y.; Gu, J.; Wei, S.; Zhang, A.; Pan, X. Aberrant methylation-induced dysfunction of p16 is associated with osteoblast activation caused by fluoride. Environ. Toxicol. 2018, 34, 37–47. [Google Scholar] [CrossRef]
- Simonet, W.; Lacey, D.; Dunstan, C.; Kelley, M.; Chang, M.-S.; Lüthy, R.; Nguyen, H.; Wooden, S.; Bennett, L.; Boone, T.; et al. Osteoprotegerin: A Novel Secreted Protein Involved in the Regulation of Bone Density. Cell 1997, 89, 309–319. [Google Scholar] [CrossRef]
- Usui, M.; Onizuka, S.; Sato, T.; Kokabu, S.; Ariyoshi, W.; Nakashima, K. Mechanism of alveolar bone destruction in periodontitis—Periodontal bacteria and inflammation. Jpn. Dent. Sci. Rev. 2021, 57, 201–208. [Google Scholar] [CrossRef] [PubMed]
- McDonald, M.M.; Kim, A.S.; Mulholland, B.S.; Rauner, M. New Insights Into Osteoclast Biology. JBMR Plus 2021, 5, e10539. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Patil, S.; Xu, F.; Lin, X.; Qian, A. Role of Biomolecules in Osteoclasts and Their Therapeutic Potential for Osteoporosis. Biomolecules 2021, 11, 747. [Google Scholar] [CrossRef]
- Kitamura, N.; Kaminuma, O. Isoform-Selective NFAT Inhibitor: Potential Usefulness and Development. Int. J. Mol. Sci. 2021, 22, 2725. [Google Scholar] [CrossRef]
- Bhawal, U.K.; Lee, H.-J.; Arikawa, K.; Shimosaka, M.; Suzuki, M.; Toyama, T.; Sato, T.; Kawamata, R.; Taguchi, C.; Hamada, N.; et al. Micromolar sodium fluoride mediates anti-osteoclastogenesis in Porphyromonas gingivalis-induced alveolar bone loss. Int. J. Oral Sci. 2015, 7, 242–249. [Google Scholar] [CrossRef]
- Okuda, A.; Kanehisa, J.; Heersche, J.N.M. The effects of sodium fluoride on the resorptive activity of isolated osteoclasts. J. Bone Miner. Res. 2010, 5, S115–S120. [Google Scholar] [CrossRef] [PubMed]
- Łukomska, A.; Baranowska-Bosiacka, I.; Dec, K.; Pilutin, A.; Tarnowski, M.; Jakubczyk, K.; Żwierełło, W.; Skórka-Majewicz, M.; Chlubek, D.; Gutowska, I. Changes in Gene and Protein Expression of Metalloproteinase-2 and -9 and their Inhibitors TIMP2 and TIMP3 in Different Parts of Fluoride-Exposed Rat Brain. Int. J. Mol. Sci. 2020, 22, 391. [Google Scholar] [CrossRef] [PubMed]
- Cvikl, B.; Lussi, A.; Carvalho, T.S.; Moritz, A.; Gruber, R. Stannous chloride and stannous fluoride are inhibitors of matrix metalloproteinases. J. Dent. 2018, 78, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Miyauchi, Y.; Ninomiya, K.; Miyamoto, H.; Sakamoto, A.; Iwasaki, R.; Hoshi, H.; Miyamoto, K.; Hao, W.; Yoshida, S.; Morioka, H.; et al. The Blimp1–Bcl6 axis is critical to regulate osteoclast differentiation and bone homeostasis. J. Exp. Med. 2010, 207, 751–762. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Ma, Y.; Zhong, N.; Pei, J. The Inverted U-Curve Association of Fluoride and Osteoclast Formation in Mice. Biol. Trace Elem. Res. 2019, 191, 419–425. [Google Scholar] [CrossRef]
- Jiang, N.; Guo, F.; Xu, W.; Zhang, Z.; Jin, H.; Shi, L.; Zhang, X.; Gao, J.; Xu, H. Effect of fluoride on osteocyte-driven osteoclastic differentiation. Toxicology 2020, 436, 152429. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Guo, F.; Sun, B.; Zhang, X.; Xu, H. Different Effects of Fluoride Exposure on the Three Major Bone Cell Types. Biol. Trace Elem. Res. 2019, 193, 226–233. [Google Scholar] [CrossRef]
- Haffajee, A.D.; Socransky, S.S. Introduction to microbial aspects of periodontal biofilm communities, development and treatment. Periodontology 2000 2006, 42, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Graves, D.; Cochran, D. The Contribution of Interleukin-1 and Tumor Necrosis Factor to Periodontal Tissue Destruction. J. Periodontol. 2003, 74, 391–401. [Google Scholar] [CrossRef]
- Armitage, G.C. Periodontal diagnoses and classification of periodontal diseases. Periodontology 2000 2004, 34, 9–21. [Google Scholar] [CrossRef]
- Preshaw, P.M.; Taylor, J. How has research into cytokine interactions and their role in driving immune responses impacted our understanding of periodontitis? J. Clin. Periodontol. 2011, 38, 60–84. [Google Scholar] [CrossRef] [PubMed]
- Kornman, K.S. Mapping the Pathogenesis of Periodontitis: A New Look. J. Periodontol. 2008, 79, 1560–1568. [Google Scholar] [CrossRef]
- Lee, H.-J.; Choi, C.-H. Anti-inflammatory effects of bamboo salt and sodium fluoride in human gingival fibroblasts—An in vitro study. Kaohsiung J. Med. Sci. 2015, 31, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.B.; Florez, J.C. Genetics of diabetes mellitus and diabetes complications. Nat. Rev. Nephrol. 2020, 16, 377–390. [Google Scholar] [CrossRef]
- Preshaw, P.M.; Bissett, S.M. Periodontitis and diabetes. Br. Dent. J. 2019, 227, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Pirih, F.Q.; Monajemzadeh, S.; Singh, N.; Sinacola, R.S.; Shin, J.M.; Chen, T.; Fenno, J.C.; Kamarajan, P.; Rickard, A.H.; Travan, S.; et al. Association between metabolic syndrome and periodontitis: The role of lipids, inflammatory cytokines, altered host response, and the microbiome. Periodontology 2000 2021, 87, 50–75. [Google Scholar] [CrossRef]
- Tuleta, I.; Frangogiannis, N.G. Diabetic fibrosis. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2020, 1867, 166044. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, M.R.; Atkinson, M.A.; Brusko, T.M. Pleiotropic roles of the insulin-like growth factor axis in type 1 diabetes. Curr. Opin. Endocrinol. Diabetes Obes. 2019, 26, 188–194. [Google Scholar] [CrossRef]
- Nackiewicz, D.; Dan, M.; Speck, M.; Chow, S.Z.; Chen, Y.-C.; Pospisilik, J.A.; Verchere, C.B.; Ehses, J.A. Islet Macrophages Shift to a Reparative State following Pancreatic Beta-Cell Death and Are a Major Source of Islet Insulin-like Growth Factor-1. iScience 2019, 23, 100775. [Google Scholar] [CrossRef]
- Camaya, I.; Mok, T.Y.; Lund, M.; To, J.; Braidy, N.; Robinson, M.W.; Santos, J.; O’Brien, B.; Donnelly, S. The parasite-derived peptide FhHDM-1 activates the PI3K/Akt pathway to prevent cytokine-induced apoptosis of β-cells. J. Mol. Med. 2021, 99, 1605–1621. [Google Scholar] [CrossRef]
- Rachdaoui, N. Insulin: The Friend and the Foe in the Development of Type 2 Diabetes Mellitus. Int. J. Mol. Sci. 2020, 21, 1770. [Google Scholar] [CrossRef]
- Yakar, S.; Werner, H.; Rosen, C.J. 40 YEARS OF IGF1: Insulin-like growth factors: Actions on the skeleton. J. Mol. Endocrinol. 2018, 61, T115–T137. [Google Scholar] [CrossRef]
- Lee, H.-J.; Arikawa, K. Influence of Low Level Sodium Fluoride on Expression of IGF-1 and IGF-2 Protein in Experimental Type 2 Diabetes with Periodontitis Model. J. Hard Tissue Biol. 2015, 24, 319–324. [Google Scholar] [CrossRef]
- Lv, X.-H.; Zhao, D.-H.; Cai, S.-Z.; Luo, S.-Y.; You, T.; Xu, B.-L.; Chen, K. Autophagy plays a protective role in cell death of osteoblasts exposure to lead chloride. Toxicol. Lett. 2015, 239, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Yoshii, S.; Kuma, A.; Mizushima, N. Transgenic rescue of Atg5-null mice from neonatal lethality with neuron-specific expression of ATG5: Systemic analysis of adult Atg5-deficient mice. Autophagy 2017, 13, 763–764. [Google Scholar] [CrossRef] [PubMed]
- Otomo, C.; Metlagel, Z.; Takaesu, G.; Otomo, T. Structure of the human ATG12~ATG5 conjugate required for LC3 lipidation in autophagy. Nat. Struct. Mol. Biol. 2013, 20, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Zhu, Q.; Dee, R.; Opheim, Z.; Mack, C.P.; Cyr, D.M.; Taylor, J.M. Focal Adhesion Kinase-mediated Phosphorylation of Beclin1 Protein Suppresses Cardiomyocyte Autophagy and Initiates Hypertrophic Growth. J. Biol. Chem. 2017, 292, 2065–2079. [Google Scholar] [CrossRef]
- Zhang, S.; Niu, Q.; Gao, H.; Ma, R.; Lei, R.; Zhang, C.; Xia, T.; Li, P.; Xu, C.; Wang, C.; et al. Excessive apoptosis and defective autophagy contribute to developmental testicular toxicity induced by fluoride. Environ. Pollut. 2016, 212, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sato, F.; Tanimoto, K.; Rajeshwaran, N.; Thangavelu, L.; Makishima, M.; Bhawal, U.K. The Potential Roles of Dec1 and Dec2 in Periodontal Inflammation. Int. J. Mol. Sci. 2021, 22, 10349. [Google Scholar] [CrossRef] [PubMed]
- Oka, S.; Li, X.; Zhang, F.; Taguchi, C.; Tewari, N.; Kim, I.S.; Zhong, L.; Arikawa, K.; Liu, Y.; Bhawal, U.K. Oral toxicity to high level sodium fluoride causes impairment of autophagy. Res. Sq. 2021, 1–22. [Google Scholar] [CrossRef]
- Anuradha, C.D.; Kanno, S.; Hirano, S. Oxidative damage to mitochondria is a preliminary step to caspase-3 activation in fluoride-induced apoptosis in HL-60 cells. Free. Radic. Biol. Med. 2001, 31, 367–373. [Google Scholar] [CrossRef]
- Lee, J.-H.; Jung, J.-Y.; Jeong, Y.-J.; Park, J.-H.; Yang, K.-H.; Choi, N.-K.; Kim, S.-H.; Kim, W.-J. Involvement of both mitochondrial- and death receptor-dependent apoptotic pathways regulated by Bcl-2 family in sodium fluoride-induced apoptosis of the human gingival fibroblasts. Toxicology 2008, 243, 340–347. [Google Scholar] [CrossRef]
- Suzuki, M.; Bandoski, C.; Bartlett, J.D. Fluoride induces oxidative damage and SIRT1/autophagy through ROS-mediated JNK signaling. Free Radic. Biol. Med. 2015, 89, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Jin, T.; Lu, Y. AntimiR-30b Inhibits TNF-α Mediated Apoptosis and Attenuated Cartilage Degradation through Enhancing Autophagy. Cell. Physiol. Biochem. 2016, 40, 883–894. [Google Scholar] [CrossRef] [PubMed]
- Daiwile, A.; Sivanesan, S.; Izzotti, A.; Bafana, A.; Naoghare, P.K.; Arrigo, P.; Purohit, H.J.; Parmar, D.; Kannan, K. Noncoding RNAs: Possible Players in the Development of Fluorosis. BioMed Res. Int. 2015, 2015, 1–10. [Google Scholar] [CrossRef]
- Ouyang, T.; Qin, Y.; Luo, K.; Han, X.; Yu, C.; Zhang, A.; Pan, X. miR-486-3p regulates CyclinD1 and promotes fluoride-induced osteoblast proliferation and activation. Environ. Toxicol. 2021, 36, 1817–1828. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Qin, Y.; Luo, K.; Wang, X.; Yu, C.; Zhang, A.; Pan, X. Downregulation of miR-4755-5p promotes fluoride-induced osteoblast activation via tageting Cyclin D1. J. Trace Elem. Med. Biol. 2020, 62, 126626. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Yang, Y.; Wang, H.; Darko, G.; Sun, D.; Gao, Y. Identification of miR-200c-3p as a major regulator of SaoS2 cells activation induced by fluoride. Chemosphere 2018, 199, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, X.; Zhao, Z.; Xu, H. Preliminary Analysis of MicroRNAs Expression Profiling in MC3T3-E1 Cells Exposed to Fluoride. Biol. Trace Element Res. 2016, 176, 367–373. [Google Scholar] [CrossRef]
- Li, C.; Qin, Y.; Ouyang, T.; Yao, M.; Zhang, A.; Luo, P.; Pan, X. miR-122-5p Mediates Fluoride-Induced Osteoblast Activation by Targeting CDK4. Biol. Trace Element Res. 2020, 199, 1215–1227. [Google Scholar] [CrossRef] [PubMed]
- Deng, Q.; Yang, J.; Zhouyang, J.; Sheng, W.; Gao, S.; Zhang, Y.; Haopeng, L.; Bingxin, B.; Mengting, W. Preliminary screening of fluorine-stained osteoblastic apoptosis-related microRNA. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2021, 305, 359–372. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Li, Z.; Xu, Z.; Chen, C.; Wang, J.; Zhu, J.; Dong, Z. miR-378d is Involved in the Regulation of Apoptosis and Autophagy of and E2 Secretion from Cultured Ovarian Granular Cells Treated by Sodium Fluoride. Biol. Trace Elem. Res. 2021, 199, 4119–4128. [Google Scholar] [CrossRef]
- Sun, Z.; Zhang, W.; Li, S.; Xue, X.; Niu, R.; Shi, L.; Li, B.; Wang, X.; Wang, J. Altered miRNAs expression profiling in sperm of mice induced by fluoride. Chemosphere 2016, 155, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cheng, M.; Zhao, Y.; Wang, J.; Wang, J. Effects of fluoride on PIWI-interacting RNA expression profiling in testis of mice. Chemosphere 2020, 269, 128727. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Y.; Guo, Z.; Li, R.; Xue, X.; Sun, Z.; Niu, R. Effects of perinatal fluoride exposure on the expressions of miR-124 and miR-132 in hippocampus of mouse pups. Chemosphere 2018, 197, 117–122. [Google Scholar] [CrossRef]
- Thylstrup, A.; Fejerskov, O. Textbook of Clinical Cariology, 2nd ed.; Munksgaard: Copenhagen, Denmark, 1994. [Google Scholar]
- Silva, M.C.C.; Lima, C.C.B.; Lima, M.D.D.M.D.; Moura, L.D.F.A.D.D.; Tabchoury, C.P.M.; de Moura, M.S. Effect of fluoridated water on dental caries and fluorosis in schoolchildren who use fluoridated dentifrice. Braz. Dent. J. 2021, 32, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Manchanda, S.; Sardana, D.; Liu, P.; Lee, G.H.; Li, K.Y.; Lo, E.C.; Yiu, C.K. Topical fluoride to prevent early childhood caries: Systematic review with network meta-analysis. J. Dent. 2021, 116, 103885. [Google Scholar] [CrossRef]
- Chaurasiya, A.; Gojanur, S. Evaluation of the clinical efficacy of 38% silver diamine fluoride in arresting dental caries in primary teeth and its parental acceptance. J. Indian Soc. Pedod. Prev. Dent. 2021, 39, 85–89. [Google Scholar]
- Nanda, T.; Jain, S.; Kaur, H.; Kapoor, D.; Nanda, S.; Jain, R. Root conditioning in periodontology—Revisited. J. Nat. Sci. Biol. Med. 2014, 5, 356–358. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.C.; Moura, C.C.G.; Ferreira, S.P.B.; De Magalhães, D.; Dechichi, P.; Soares, P.B.F. Biological Effects of a Root Conditioning Treatment on Periodontally Affected Teeth—An In Vitro Analysis. Braz. Dent. J. 2016, 27, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Shreehari, A.; Darekar, H.; Borthakur, R. A comparative analysis of root surface biomodification with ethylene diamine tetra acetic acid and tetracycline hydrochloride: An in vitro scanning electron microscopic study. Med. J. Armed Forces India 2016, 72, 145–151. [Google Scholar] [CrossRef]
- Andreasen, J.O.; Borum, M.K.; Jacobsen, H.L.; Andreasen, F.M. Replantation of 400 avulsed permanent incisors. 4. Factors related to periodontal ligament healing. Dent. Traumatol. 1995, 11, 76–89. [Google Scholar] [CrossRef] [PubMed]
- Hasanuddin, S.; Reddy, J.S. Sequelae of delayed replantation of maxillary permanent incisors after avulsion: A case series with 24-month follow-up and clinical review. J. Indian Soc. Pedod. Prev. Dent. 2018, 36, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Sardana, D.; Goyal, A.; Gauba, K. Delayed replantation of avulsed tooth with 15-h extra-oral time: 3-year follow-up. Singap. Dent. J. 2014, 35, 71–76. [Google Scholar] [CrossRef]
- Mesquita, G.C.; Soares, P.B.F.; Moura, C.C.G.; Roscoe, M.G.; Paiva, S.M.; Soares, C. A 12-Year Retrospective Study of Avulsion Cases in a Public Brazilian Dental Trauma Service. Braz. Dent. J. 2017, 28, 749–756. [Google Scholar] [CrossRef] [PubMed]
- Lauridsen, E.; Andreasen, J.O.; Bouaziz, O.; Andersson, L. Risk of ankylosis of 400 avulsed and replanted human teeth in relation to length of dry storage: A re-evaluation of a long-term clinical study. Dent. Traumatol. 2019, 36, 108–116. [Google Scholar] [CrossRef]
- Kameyama, Y.; Nakane, S.; Maeda, H.; Saito, T.; Konishi, S.; Ito, N. Effect of fluoride on root resorption caused by mechanical injuries of the periodontal soft tissues in rats. Dent. Traumatol. 1994, 10, 210–214. [Google Scholar] [CrossRef] [PubMed]
- Foo, M.; Jones, A.; Darendeliler, M.A. Physical properties of root cementum: Part 9. Effect of systemic fluoride intake on root resorption in rats. Am. J. Orthod. Dentofac. Orthop. 2007, 131, 34–43. [Google Scholar] [CrossRef]
- Rubin, M.R.; Bilezikian, J.P. New anabolic therapies in osteoporosis. Curr. Opin. Rheumatol. 2002, 14, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, R.N.; Basha, S.; Al-Thomali, Y.; Alshamrani, A.S.; Alzahrani, F.S.; Enan, E.T. Self-assembling peptide P11-4 in remineralization of enamel caries—A systematic review of in-vitro studies. Acta Odontol. Scand. 2020, 79, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Pithon, M.M.; Baião, F.S.; Sant’Anna, L.I.D.; Tanaka, O.M.; Maia, L. Effectiveness of casein phosphopeptide-amorphous calcium phosphate-containing products in the prevention and treatment of white spot lesions in orthodontic patients: A systematic review. J. Investig. Clin. Dent. 2019, 10, e12391. [Google Scholar] [CrossRef] [PubMed]
- Butera, A.; Pascadopoli, M.; Gallo, S.; Lelli, M.; Tarterini, F.; Giglia, F.; Scribante, A. SEM/EDS Evaluation of the Mineral Deposition on a Polymeric Composite Resin of a Toothpaste Containing Biomimetic Zn-Carbonate Hydroxyapatite (microRepair®) in Oral Environment: A Randomized Clinical Trial. Polymers 2021, 13, 2740. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Tewari, N.; Sato, F.; Tanimoto, K.; Thangavelu, L.; Makishima, M.; Bhawal, U.K. Biphasic Functions of Sodium Fluoride (NaF) in Soft and in Hard Periodontal Tissues. Int. J. Mol. Sci. 2022, 23, 962. https://doi.org/10.3390/ijms23020962
Wang X, Tewari N, Sato F, Tanimoto K, Thangavelu L, Makishima M, Bhawal UK. Biphasic Functions of Sodium Fluoride (NaF) in Soft and in Hard Periodontal Tissues. International Journal of Molecular Sciences. 2022; 23(2):962. https://doi.org/10.3390/ijms23020962
Chicago/Turabian StyleWang, Xingzhi, Nitesh Tewari, Fuyuki Sato, Keiji Tanimoto, Lakshmi Thangavelu, Makoto Makishima, and Ujjal K. Bhawal. 2022. "Biphasic Functions of Sodium Fluoride (NaF) in Soft and in Hard Periodontal Tissues" International Journal of Molecular Sciences 23, no. 2: 962. https://doi.org/10.3390/ijms23020962
APA StyleWang, X., Tewari, N., Sato, F., Tanimoto, K., Thangavelu, L., Makishima, M., & Bhawal, U. K. (2022). Biphasic Functions of Sodium Fluoride (NaF) in Soft and in Hard Periodontal Tissues. International Journal of Molecular Sciences, 23(2), 962. https://doi.org/10.3390/ijms23020962