
Fluorescence Spectroscopy of Low-Level Endogenous β-Adrenergic Receptor Expression at the Plasma Membrane of Differentiating Human iPSC-Derived Cardiomyocytes

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Generation and Characterization of WT and CRISPRed hiPSC-CM
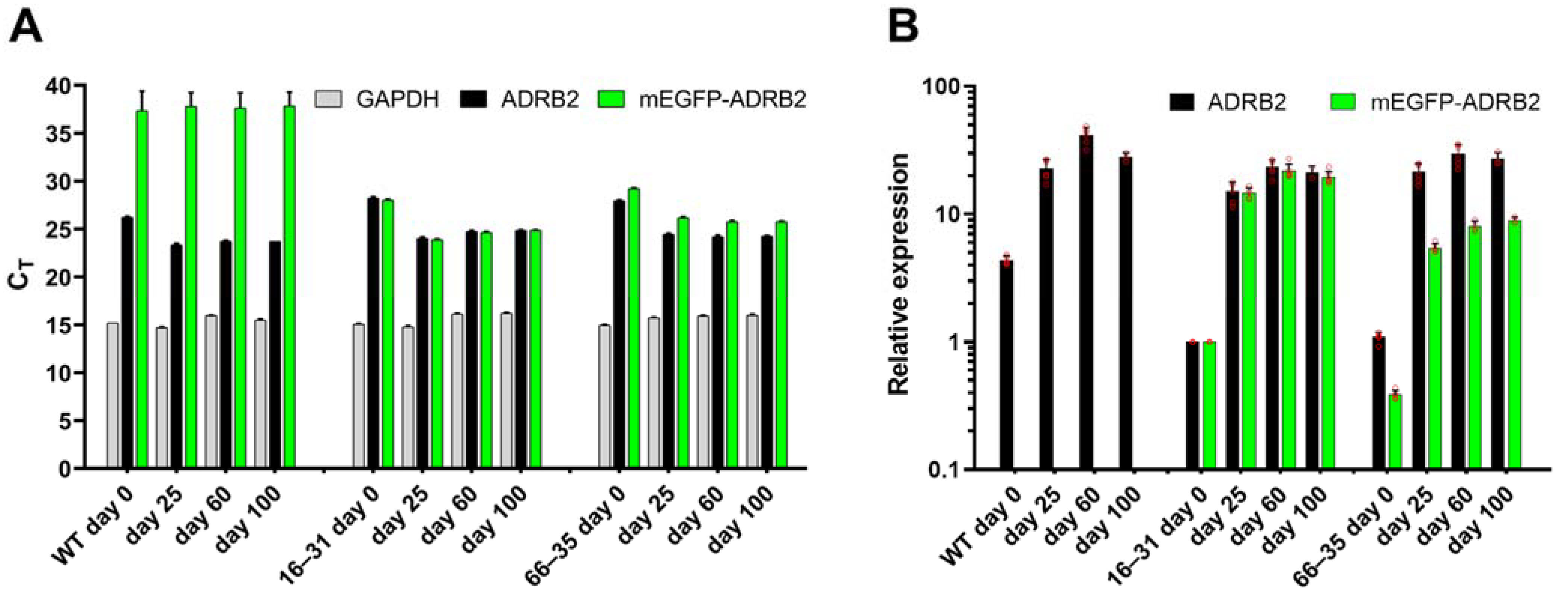
2.2. Evolution of ADRB2 Transcript Levels during Differentiation and Maturation
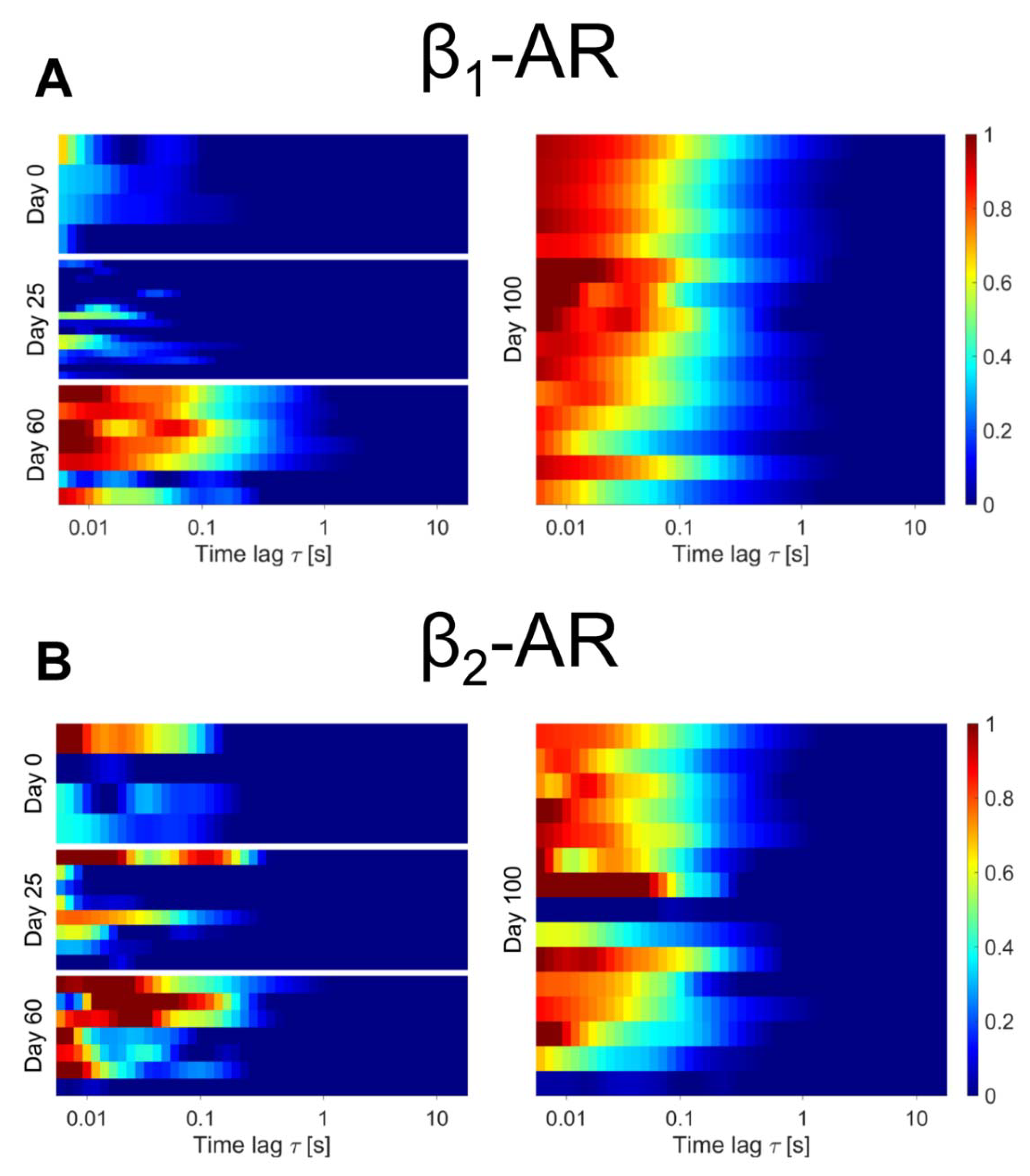
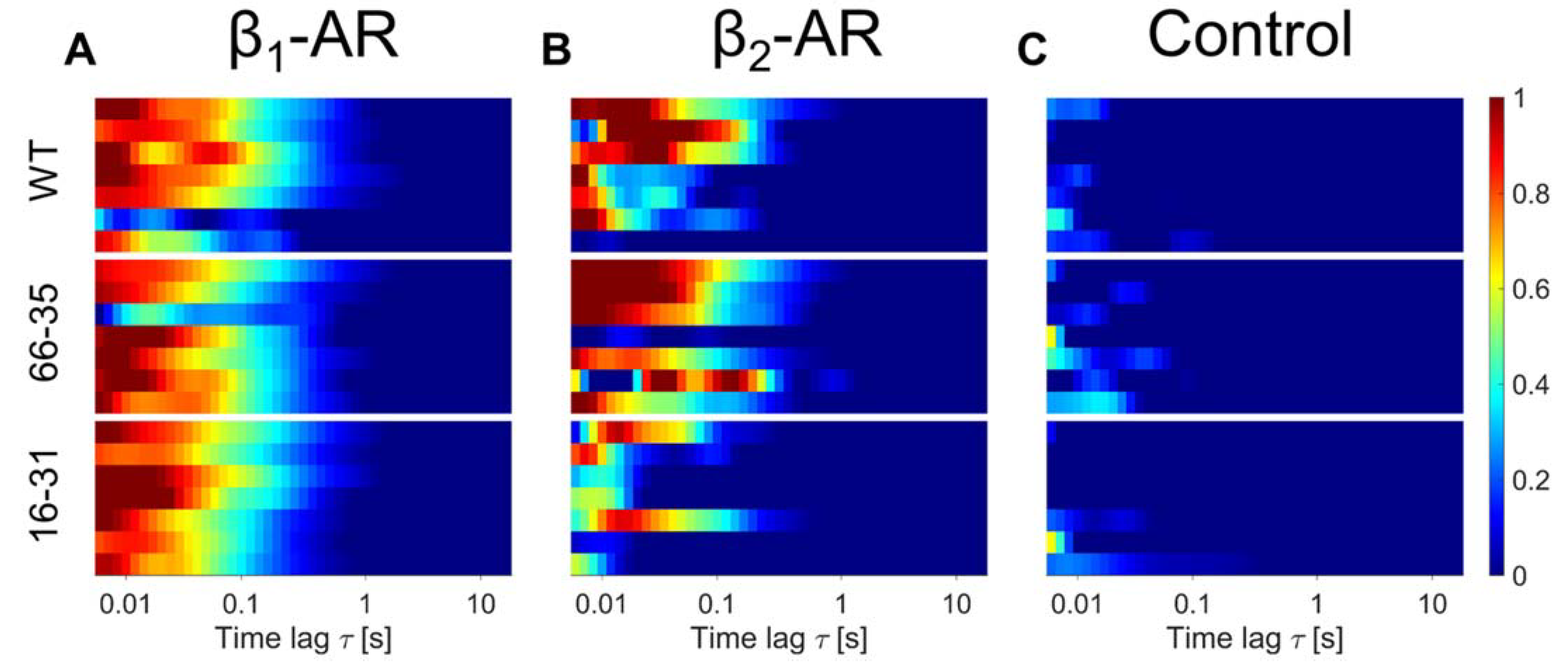
2.3. β-Adrenergic Receptor Expression at the Plasma Membrane of Differentiating hiPSC-CM
3. Discussion
4. Materials and Methods
4.1. Oligonucleotides
4.2. Differentiation and Culture of hiPSC-CM
4.3. Culture of H9c2 Cells
4.4. CRISPR Strategy
4.5. Flow Cytometry
4.6. Contractility Assay
4.7. qRT-PCR
4.8. Cell Preparation for Confocal Imaging
4.9. Immunofluorescence
4.10. Acquisition of Linescans
4.11. Analysis of Confocal Linescans
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guo, Y.; Pu, W.T. Cardiomyocyte Maturation: New Phase in Development. Circ. Res. 2020, 126, 1086–1106. [Google Scholar] [CrossRef] [PubMed]
- Babiarz, J.E.; Ravon, M.; Sridhar, S.; Ravindran, P.; Swanson, B.; Bitter, H.; Weiser, T.; Chiao, E.; Certa, U.; Kolaja, K.L. Determination of the Human Cardiomyocyte mRNA and miRNA Differentiation Network by Fine-Scale Profiling. Stem Cells Dev. 2012, 21, 1956–1965. [Google Scholar] [CrossRef] [PubMed]
- Churko, J.M.; Garg, P.; Treutlein, B.; Venkatasubramanian, M.; Wu, H.; Lee, J.; Wessells, Q.N.; Chen, S.-Y.; Chen, W.-Y.; Chetal, K.; et al. Defining human cardiac transcription factor hierarchies using integrated single-cell heterogeneity analysis. Nat. Commun. 2018, 9, 4906. [Google Scholar] [CrossRef] [PubMed]
- Grancharova, T.; Gerbin, K.A.; Rosenberg, A.B.; Roco, C.M.; Arakaki, J.E.; DeLizo, C.M.; Dinh, S.Q.; Donovan-Maiye, R.M.; Hirano, M.; Nelson, A.M.; et al. A comprehensive analysis of gene expression changes in a high replicate and open-source dataset of differentiating hiPSC-derived cardiomyocytes. Sci. Rep. 2021, 11, 15845. [Google Scholar] [CrossRef]
- Liu, Q.; Jiang, C.; Xu, J.; Zhao, M.-T.; Van Bortle, K.; Cheng, X.; Wang, G.; Chang, H.Y.; Wu, J.C.; Snyder, M.P. Genome-Wide Temporal Profiling of Transcriptome and Open Chromatin of Early Cardiomyocyte Differentiation Derived From hiPSCs and hESCs. Circ. Res. 2017, 121, 376–391. [Google Scholar] [CrossRef] [PubMed]
- Brodde, O.E. Beta 1- and beta 2-adrenoceptors in the human heart: Properties, function, and alterations in chronic heart failure. Pharmacol Rev. 1991, 43, 203–242. [Google Scholar] [PubMed]
- Engelhardt, S.; Böhm, M.; Erdmann, E.; Lohse, M.J. Analysis of beta-adrenergic receptor mRNA levels in human ventricular biopsy specimens by quantitative polymerase chain reactions: Progressive reduction of beta1-adrenergic receptor mRNA in heart failure. J. Am. Coll. Cardiol. 1996, 27, 146–154. [Google Scholar] [CrossRef]
- Wu, H.; Lee, J.; Vincent, L.G.; Wang, Q.; Gu, M.; Lan, F.; Churko, J.M.; Sallam, K.I.; Matsa, E.; Sharma, A.; et al. Epigenetic Regulation of Phosphodiesterases 2A and 3A Underlies Compromised β-Adrenergic Signaling in an iPSC Model of Dilated Cardiomyopathy. Cell Stem Cell 2015, 17, 89–100. [Google Scholar] [CrossRef]
- Jung, G.; Fajardo, G.; Ribeiro, A.J.S.; Kooiker, K.B.; Coronado, M.; Zhao, M.; Hu, D.; Reddy, S.; Kodo, K.; Sriram, K.; et al. Time-dependent evolution of functional vs. remodeling signaling in induced pluripotent stem cell-derived cardiomyocytes and induced maturation with biomechanical stimulation. FASEB J. 2016, 30, 1464–1479. [Google Scholar] [CrossRef]
- Hasan, A.; Mohammadi, N.; Nawaz, A.; Kodagoda, T.; Diakonov, I.; Harding, S.E.; Gorelik, J. Age-Dependent Maturation of iPSC-CMs Leads to the Enhanced Compartmentation of β2AR-cAMP Signalling. Cells 2020, 9, 2275. [Google Scholar] [CrossRef]
- Kondrashov, A.; Hoang, M.D.; Smith, J.G.; Bhagwan, J.R.; Duncan, G.; Mosqueira, D.; Munoz, M.B.; Vo, N.T.; Denning, C. Simplified Footprint-Free Cas9/CRISPR Editing of Cardiac-Associated Genes in Human Pluripotent Stem Cells. Stem Cells Dev. 2018, 27, 391–404. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Jia, Z.; Cui, J.; Yang, H.; Yang, H.; Zhang, Y.; Zhou, C. Beta-adrenergic signals regulate cardiac differentiation of mouse embryonic stem cells via mitogen-activated protein kinase pathways. Dev. Growth Differ. 2011, 53, 772–779. [Google Scholar] [CrossRef]
- Bathe-Peters, M.; Gmach, P.; Boltz, H.-H.; Einsiedel, J.; Gotthardt, M.; Hübner, H.; Gmeiner, P.; Lohse, M.J.; Annibale, P. Visualization of β-adrenergic receptor dynamics and differential localization in cardiomyocytes. Proc. Natl. Acad. Sci. USA 2021, 118. [Google Scholar] [CrossRef]
- Nikolaev, V.O.; Moshkov, A.; Lyon, A.R.; Miragoli, M.; Novak, P.; Paur, H.; Lohse, M.J.; Korchev, Y.E.; Harding, S.E.; Gorelik, J. Beta2-adrenergic receptor redistribution in heart failure changes cAMP compartmentation. Science 2010, 327, 1653–1657. [Google Scholar] [CrossRef] [PubMed]
- Bathe-Peters, M.; Gmach, P.; Annibale, P.; Lohse, M.J. Linescan microscopy data to extract diffusion coefficient of a fluorescent species using a commercial confocal microscope. Data Brief. 2020, 29, 105063. [Google Scholar] [CrossRef] [PubMed]
- Guan, X.; Kobilka, T.; Kobilka, B. Enhancement of membrane insertion and function in a type IIIb membrane protein following introduction of a cleavable signal peptide. J. Biol. Chem. 1992, 267, 21995–21998. [Google Scholar] [CrossRef]
- Işbilir, A.; Serfling, R.; Möller, J.; Thomas, R.; De Faveri, C.; Zabel, U.; Scarselli, M.; Beck-Sickinger, A.G.; Bock, A.; Coin, I.; et al. Determination of G-protein–coupled receptor oligomerization by molecular brightness analyses in single cells. Nat. Protoc. 2021, 16, 1419–1451. [Google Scholar] [CrossRef]
- Qian, T.; Heaster, T.M.; Houghtaling, A.R.; Sun, K.; Samimi, K.; Skala, M.C. Label-free imaging for quality control of cardiomyocyte differentiation. Nat. Commun. 2021, 12, 4580. [Google Scholar] [CrossRef]
- Chorvat, D.; Kirchnerova, J.; Cagalinec, M.; Smolka, J.; Mateasik, A.; Chorvatova, A. Spectral Unmixing of Flavin Autofluorescence Components in Cardiac Myocytes. Biophys. J. 2005, 89, L55–L57. [Google Scholar] [CrossRef]
- Goulding, J.; Kondrashov, A.; Mistry, S.J.; Melarangi, T.; Vo, N.T.; Hoang, D.M.; White, C.W.; Denning, C.; Briddon, S.J.; Hill, S.J. The use of fluorescence correlation spectroscopy to monitor cell surface β2-adrenoceptors at low expression levels in human embryonic stem cell-derived cardiomyocytes and fibroblasts. FASEB J. 2021, 35, e21398. [Google Scholar] [CrossRef]
- Soave, M.; Stoddart, L.A.; White, C.W.; Kilpatrick, L.E.; Goulding, J.; Briddon, S.J.; Hill, S.J. Detection of genome-edited and endogenously expressed G protein-coupled receptors. FEBS J. 2021, 288, 2585–2601. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Beyer, A.; Aebersold, R. On the Dependency of Cellular Protein Levels on mRNA Abundance. Cell 2016, 165, 535–550. [Google Scholar] [CrossRef] [PubMed]
- Tahmasebi, S.; Amiri, M.; Sonenberg, N. Translational Control in Stem Cells. Front. Genet. 2018, 9, 709. [Google Scholar] [CrossRef] [PubMed]
- Patwardhan, A.; Cheng, N.; Trejo, J. Post-Translational Modifications of G Protein–Coupled Receptors Control Cellular Signaling Dynamics in Space and Time. Pharmacol. Rev. 2021, 73, 120–151. [Google Scholar] [CrossRef]
- Cuello, F.; E Knaust, A.; Saleem, U.; Loos, M.; Raabe, J.; Mosqueira, D.; Laufer, S.; Schweizer, M.; van der Kraak, P.; Flenner, F.; et al. Impairment of the ER/mitochondria compartment in human cardiomyocytes with PLN p.Arg14del mutation. EMBO Mol. Med. 2021, 13, e13074. [Google Scholar] [CrossRef]
- Burridge, P.W.; Matsa, E.; Shukla, P.; Lin, Z.C.; Churko, J.M.; Ebert, A.D.; Lan, F.; Diecke, S.; Huber, B.; Mordwinkin, N.M.; et al. Chemically defined generation of human cardiomyocytes. Nat. Methods 2014, 11, 855–860. [Google Scholar] [CrossRef]
- Lian, X.; Hsiao, C.; Wilson, G.; Zhu, K.; Hazeltine, L.B.; Azarin, S.M.; Raval, K.K.; Zhang, J.; Kamp, T.J.; Palecek, S.P. Robust cardiomyocyte differentiation from human pluripotent stem cells via temporal modulation of canonical Wnt signaling. Proc. Natl. Acad. Sci. USA 2012, 109, E1848–E1857. [Google Scholar] [CrossRef]
- Tohyama, S.; Hattori, F.; Sano, M.; Hishiki, T.; Nagahata, Y.; Matsuura, T.; Hashimoto, H.; Suzuki, T.; Yamashita, H.; Satoh, Y.; et al. Distinct Metabolic Flow Enables Large-Scale Purification of Mouse and Human Pluripotent Stem Cell-Derived Cardiomyocytes. Cell Stem Cell 2013, 12, 127–137. [Google Scholar] [CrossRef]
- Miller, D.C.; Genehr, C.; Telugu, N.S.; Kurths, S.; Diecke, S. Simple Workflow and Comparison of Media for hPSC-Cardiomyocyte Cryopreservation and Recovery. Curr. Protoc. Stem Cell Biol. 2020, 55, e125. [Google Scholar] [CrossRef]
- Dokshin, G.A.; Ghanta, K.S.; Piscopo, K.M.; Mello, C.C. Robust Genome Editing with Short Single-Stranded and Long, Partially Single-Stranded DNA Donors in Caenorhabditis elegans. Genetics 2018, 210, 781–787. [Google Scholar] [CrossRef] [Green Version]
- Vallone, V.F.; Telugu, N.S.; Fischer, I.; Miller, D.; Schommer, S.; Diecke, S.; Stachelscheid, H. Methods for Automated Single Cell Isolation and Sub-Cloning of Human Pluripotent Stem Cells. Curr. Protoc. Stem Cell Biol. 2020, 55, e123. [Google Scholar] [CrossRef] [PubMed]
- Grune, T.; Ott, C.; Häseli, S.; Höhn, A.; Jung, T. The “MYOCYTER”—Convert cellular and cardiac contractions into numbers with ImageJ. Sci. Rep. 2019, 9, 15112. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence from 5′ to 3′ | Description |
|---|---|---|
| crRNA- 103forw | CCTGCCAGACTGCGCGCCAT | crRNA used with generic tracrRNA to build the gRNA for Cas9 targeting to ADRB2 locus. Binds over start codon for knock-in of mEGFP |
| Donor plasmid for knock-in of mEGFP | AAGCTTAACGGGCAGAACGCACTGCGA AGCGGCTTCTTCAGAGCACGGGCTGGAACTGGCAGGCA CCGCGAGCCCCTAGCACCCGACAAGCTGAGTGTGCAGGACGA GTCCCCACCACACCCACACCACAGCCGCTGAA TGAGGCTTCCAGGCGTCCGCTCGCGGCCCGCAGA GCCCCGCCGTGGGTCCGCCCGCTGAGGCGCCCCCAGCCAGTGCGCTCA CCTGCCAGACTGCGCGCCATGGTGAGCAAGGGCGAGGAGC TGTTCACCGGGGTGGTGCCCATCCTGGTCGAGCTGGACGGCGA CGTAAACGGCCACAAGTTCAGCGTGTCCGGCGAGGGCGAG GGCGATGCCACCTACGGCAAGCTGACCCTGAAGTTCATCTGCA CCACCGGCAAGCTGCCCGTGCCCTGGCCCACCCTCGTGAC CACCCTGACCTACGGCGTGCAGTGCTTCAGCCGCTACCCCGAC CACATGAAGCAGCACGACTTCTTCAAGTCCGCCATGCCCG AAGGCTACGTCCAGGAGCGCACCATCTTCTTCAAGGACGACGG CAACTACAAGACCCGCGCCGAGGTGAAGTTCGAGGGCGA CACCCTGGTGAACCGCATCGAGCTGAAGGGCATCGACTTCAAGG AGGACGGCAACATCCTGGGGCACAAGCTGGAGTACAACTA CAACAGCCACAACGTCTATATCATGGCCGACAAGCAGAAGAAC GGCATCAAGGTGAACTTCAAGATCCGCCACAACATCGAGG ACGGCAGCGTGCAGCTCGCCGACCACTACCAGCAGAACACCCC CATCGGCGACGGCCCCGTGCTGCTGCCCGACAACCACTAC CTGAGCACCCAGTCCAAGCTGAGCAAAGACCCCAACGAGAAGC GCGATCACATGGTCCTGCTGGAGTTCGTGACCGCCGCCGG GATCACTCTCGGCATGGACGAGCTGTACAAGtctagaGGGCAA CCCGGGAACGGCAGCGCCTTCTTGCTGGCACCCAATAGAA GCCATGCGCCGGACCACGACGTCACGCAGCAAAGGGACGAGGT GTGGGTGGTGGGCATGGGCATCGTCATGTCTCTCATCGTC CTGGCCATCGTGTTTGGCAATGTGCTGGTCATCACAGCCATTG CCAAGTTCGAGCGTCTGCAGACGGTCACCAACTACTTCAT CACTTCACTGGCCTGTGCTGAATTC | Donor sequence (1.2 kbp) encoding for mEGFP plus 230 bp homology arms flanked by HindIII (5′) and EcoRI (3′) restriction enzyme sites, which were used for cloning into a pcDNA3.1(+) vector backbone. Served as template to generate the partially single-stranded hybrid HDR donor cocktail for knock-in of mEGFP at the ADRB2 locus using the following 4 primers: mEGFP_N-term_f mEGFP_C-term_linker_r b2AR-hum_120bp_5′arm_f1 b2AR-hum_120bp_3′arm_r1 |
| crRNA- 69forw | CTGCGCGCCATGGTGAGCAA | crRNA used with generic tracrRNA to build the gRNA for Cas9 targeting to the mEGFP-ADRB2 locus. Binds over start codon for knock-in of HA-SS |
| ssODN for knock-in of HA-SS | TGGGTCCGCCCGCTGAGGCGCCCCCA GCCAGTGCGCTCACCTGCCAGACTGCGCGCCATGA AGACGATCATCGCCCTGAGCTACATCTTCTGCCTGGTA TTCGCCGTGAGCAAGGGCGAGGAGCTGTTCACCGG GGTGGTGCCCATCCTGGTCGAGCTGGACGGC | ssODN consisting of 60 bp homology arms used as HDR donor template for knock-in of HA signal sequence at the N-terminus of mEGFP-ADRB2 locus. |
| mEGFP_ N-term_f | ATGGTGAGCAAGGGCGAGGA | Forward primer for generation of partially single-stranded hybrid HDR donor, binds N-terminus of mEGFP |
| mEGFP_ C-term_ linker_r | tctagaCTTGTACAGCTCGTCC | Reverse primer for generation of partially single-stranded hybrid HDR donor, binds C-terminus of mEGFP including a linker sequence |
| b2AR-hum_120bp_ 5′arm_f1 | CCCACACCACAGCCGCTGAA | Forward primer for generation of partially single-stranded hybrid HDR donor, binds to 5′UTR of ADRB2 |
| b2AR-hum_120bp_ 3′arm_r1 | AGACATGACGATGCCCATGC | Reverse primer for generation of partially single-stranded hybrid HDR donor, binds to N-terminus of ADRB2 |
| b2AR-hum_ mEGFP- ADRB2_f1 | ATTGGCCGAAAGTTCCCGTA | Forward sequencing primer: verification of the endogenous insertion of HDR donor template at the ADRB2 locus, binds to 5′UTR of ADRB2 upstream of 5′-homology arm of HDR donor |
| b2AR-hum_ mEGFP- ADRB2_r1 | GTCCAAAACTCGCACCAGAA | Reverse sequencing primer: verification of the endogenous insertion of HDR donor template at the ADRB2 locus, binds to N-terminus of ADRB2 downstream of the 3′homology arm of HDR donor |
| 2nd qPCR_ GFP_Fwd | TGAGCAAAGACCCCAACGAG | Forward qPCR primer for amplification of mEGFP-ADRB2. Binds to C-terminus of mEGFP |
| 2nd qPCR_ ADRB2_Rev | CGCATGGCTTCTATTGGGTG | Reverse qPCR primer for amplification of mEGFP-ADRB2. Binds to N-terminus of ADRB2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gmach, P.; Bathe-Peters, M.; Telugu, N.; Miller, D.C.; Annibale, P. Fluorescence Spectroscopy of Low-Level Endogenous β-Adrenergic Receptor Expression at the Plasma Membrane of Differentiating Human iPSC-Derived Cardiomyocytes. Int. J. Mol. Sci. 2022, 23, 10405. https://doi.org/10.3390/ijms231810405
Gmach P, Bathe-Peters M, Telugu N, Miller DC, Annibale P. Fluorescence Spectroscopy of Low-Level Endogenous β-Adrenergic Receptor Expression at the Plasma Membrane of Differentiating Human iPSC-Derived Cardiomyocytes. International Journal of Molecular Sciences. 2022; 23(18):10405. https://doi.org/10.3390/ijms231810405
Chicago/Turabian StyleGmach, Philipp, Marc Bathe-Peters, Narasimha Telugu, Duncan C. Miller, and Paolo Annibale. 2022. "Fluorescence Spectroscopy of Low-Level Endogenous β-Adrenergic Receptor Expression at the Plasma Membrane of Differentiating Human iPSC-Derived Cardiomyocytes" International Journal of Molecular Sciences 23, no. 18: 10405. https://doi.org/10.3390/ijms231810405
APA StyleGmach, P., Bathe-Peters, M., Telugu, N., Miller, D. C., & Annibale, P. (2022). Fluorescence Spectroscopy of Low-Level Endogenous β-Adrenergic Receptor Expression at the Plasma Membrane of Differentiating Human iPSC-Derived Cardiomyocytes. International Journal of Molecular Sciences, 23(18), 10405. https://doi.org/10.3390/ijms231810405