
Icariin Alleviates Escherichia coli Lipopolysaccharide-Mediated Endometritis in Mice by Inhibiting Inflammation and Oxidative Stress

 ,
,  , ,
, , 

Abstract
:
1. Introduction
2. Results
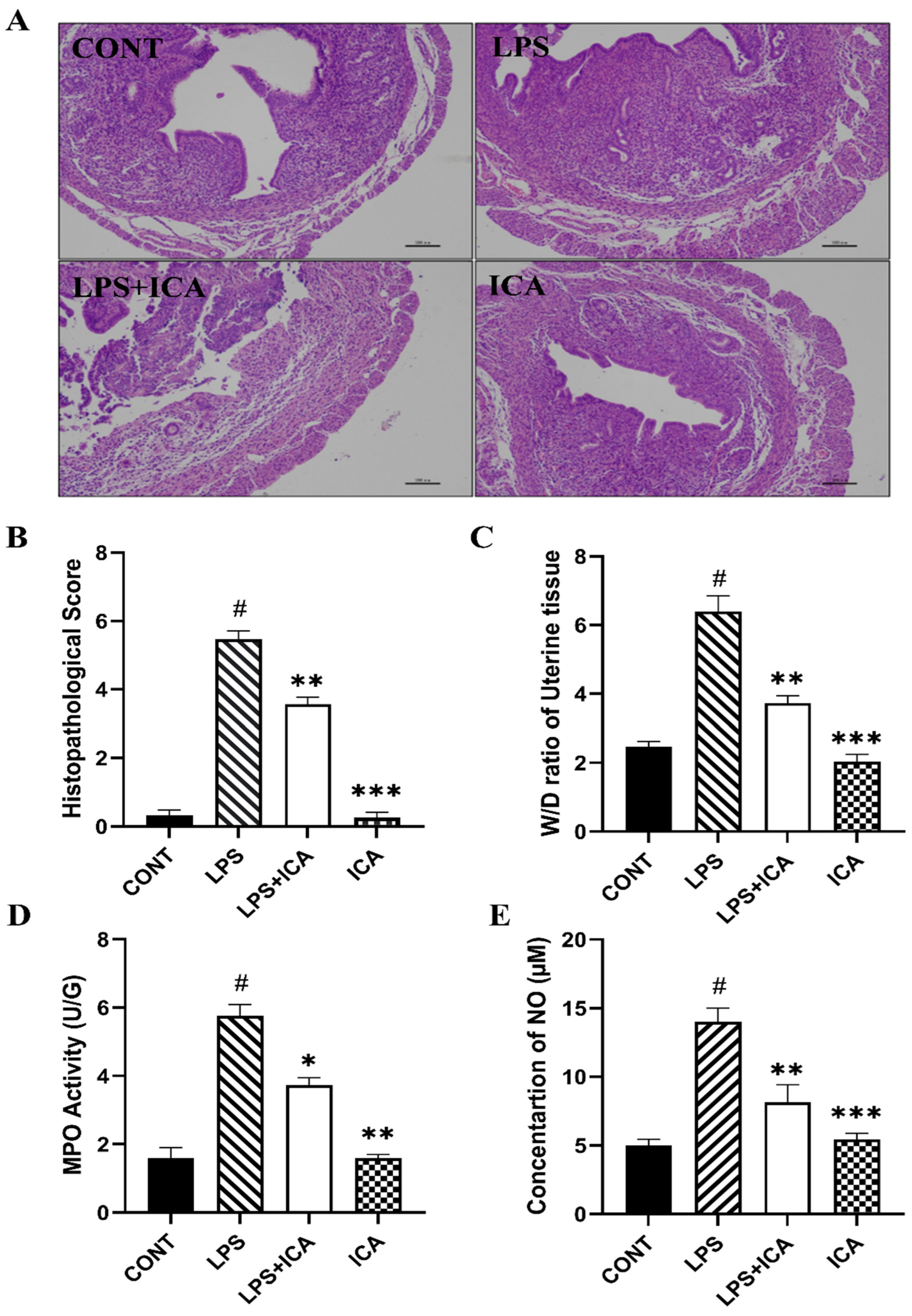
2.1. Effect of ICA against LPS-Inflicted Murine Endometritis
2.2. Effect of ICA on W/D Ratio, MPO Activity, and NO Concentration Assay
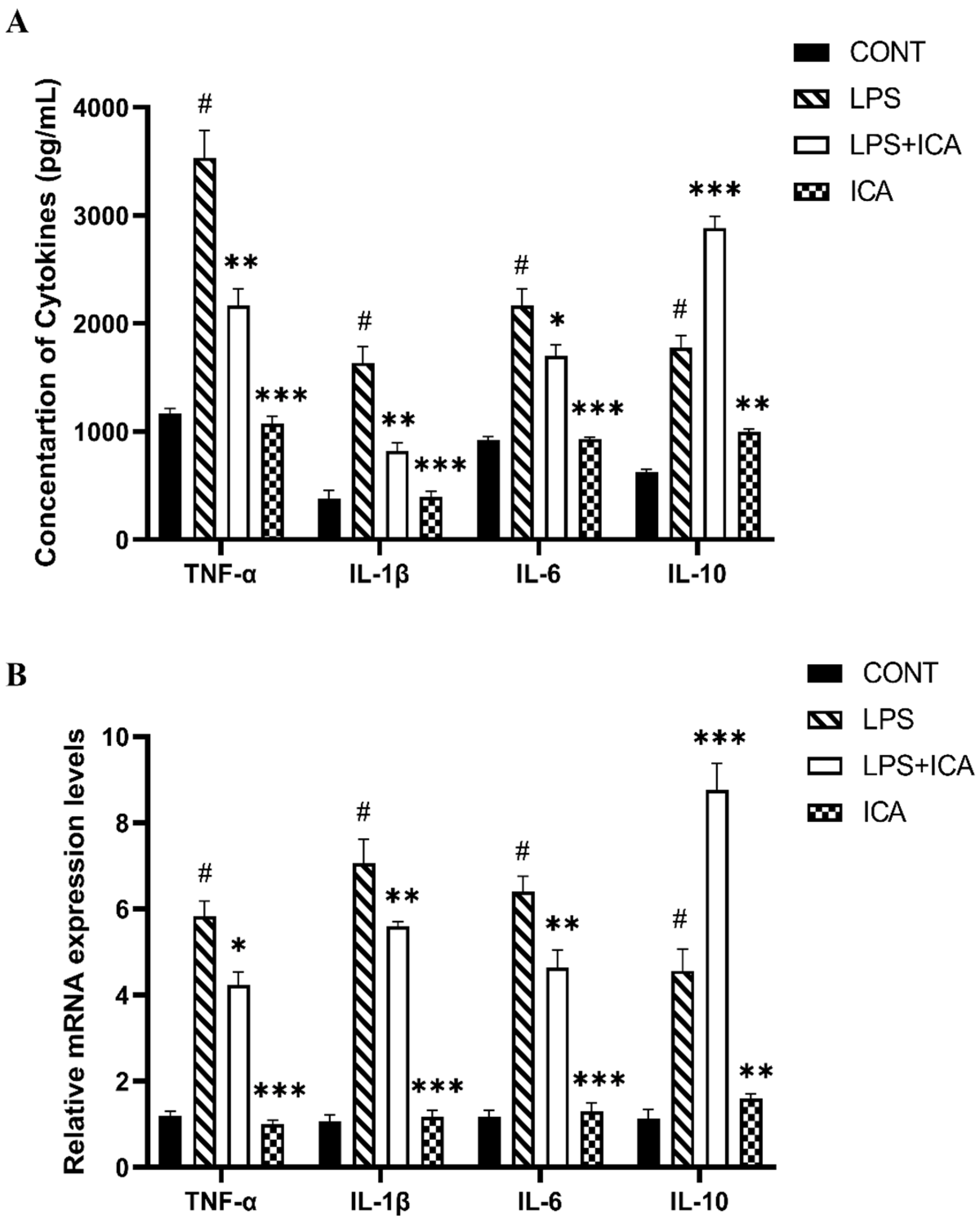
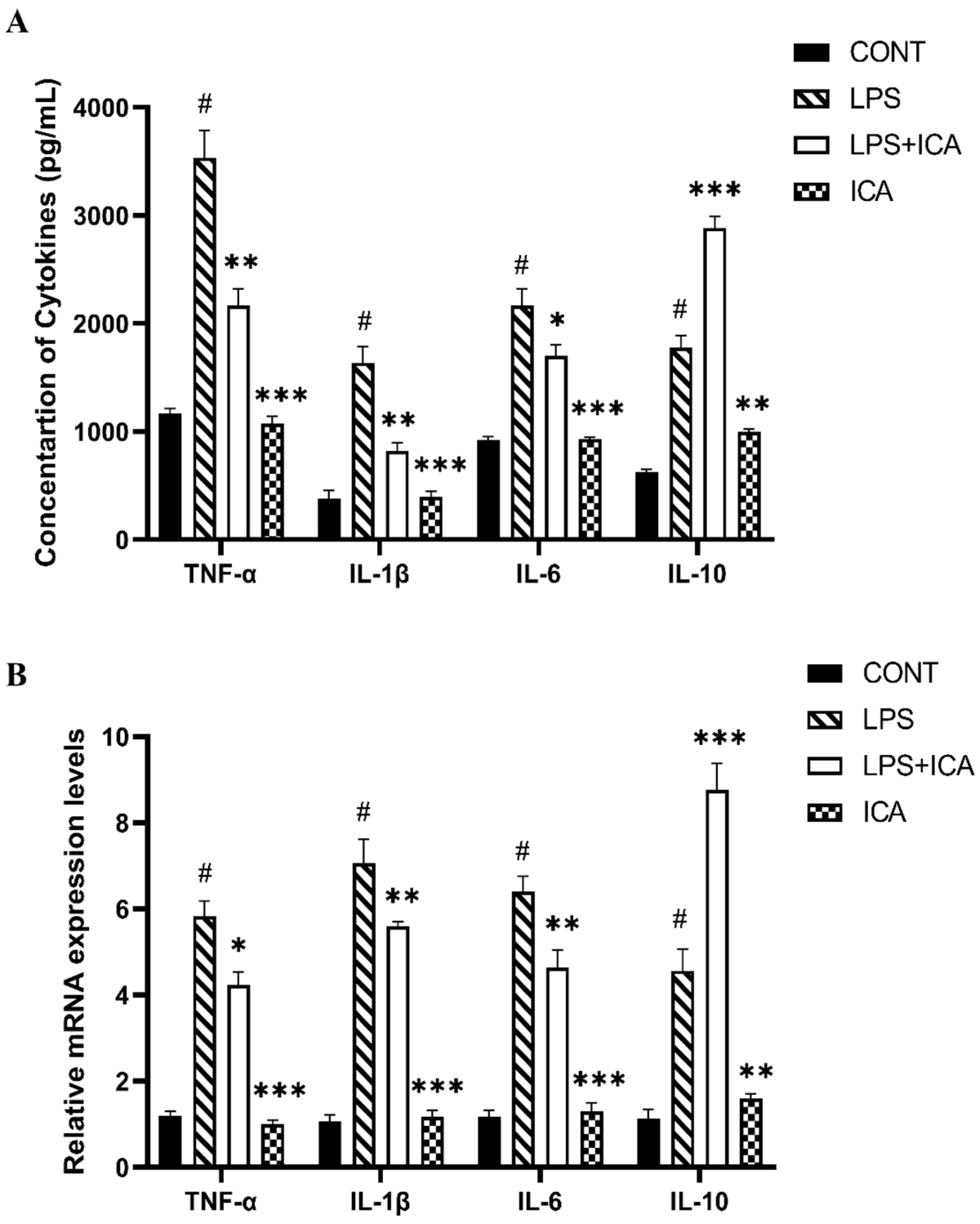
2.3. Outcome of ICA on Cytokines
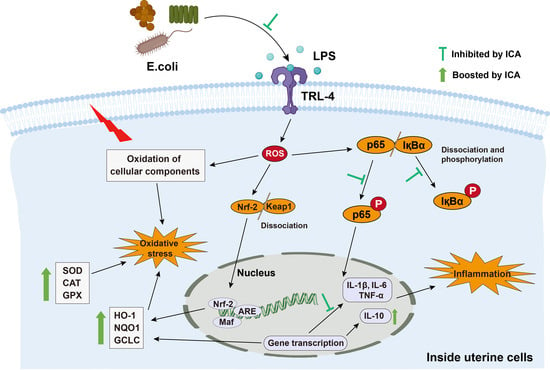
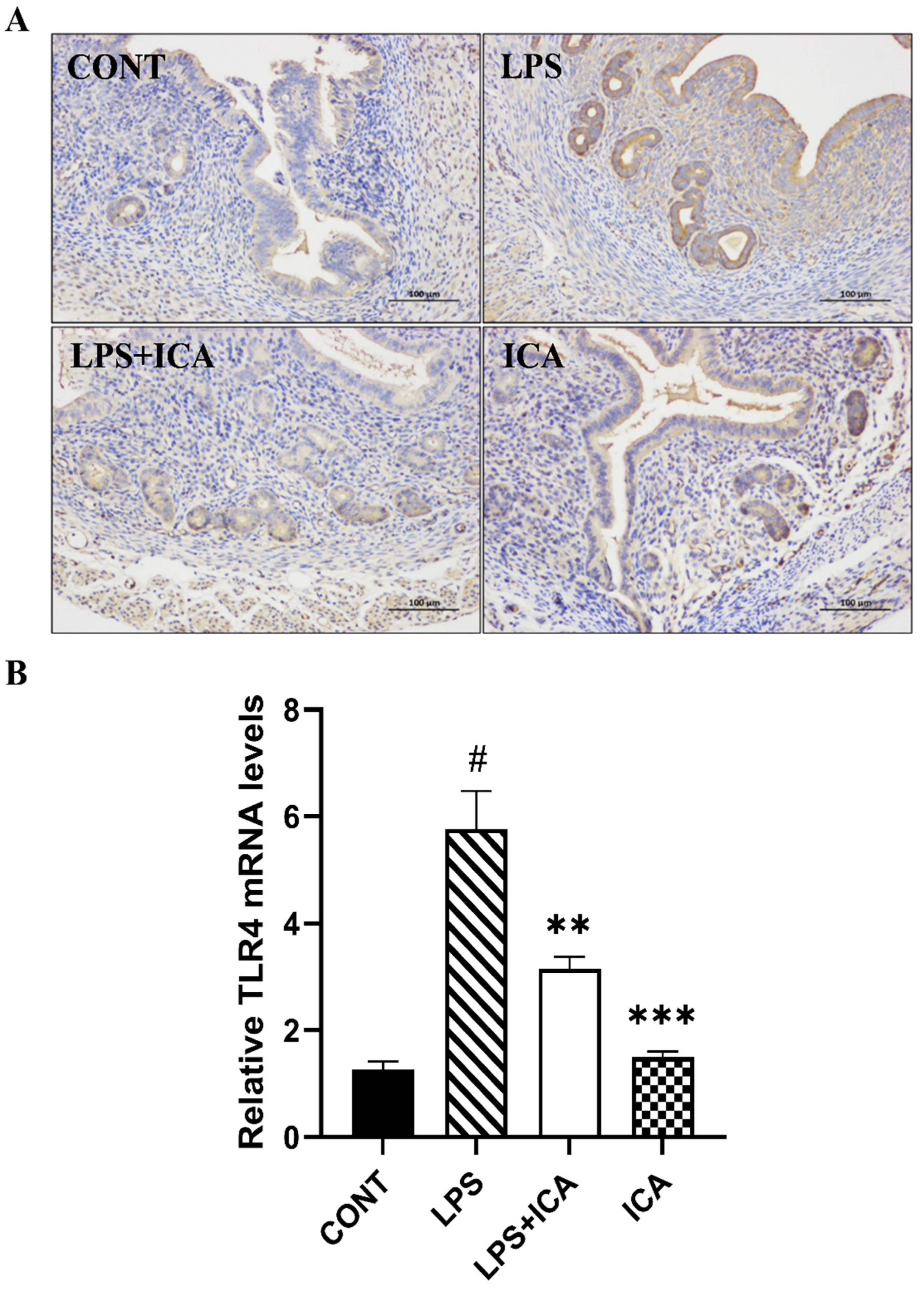
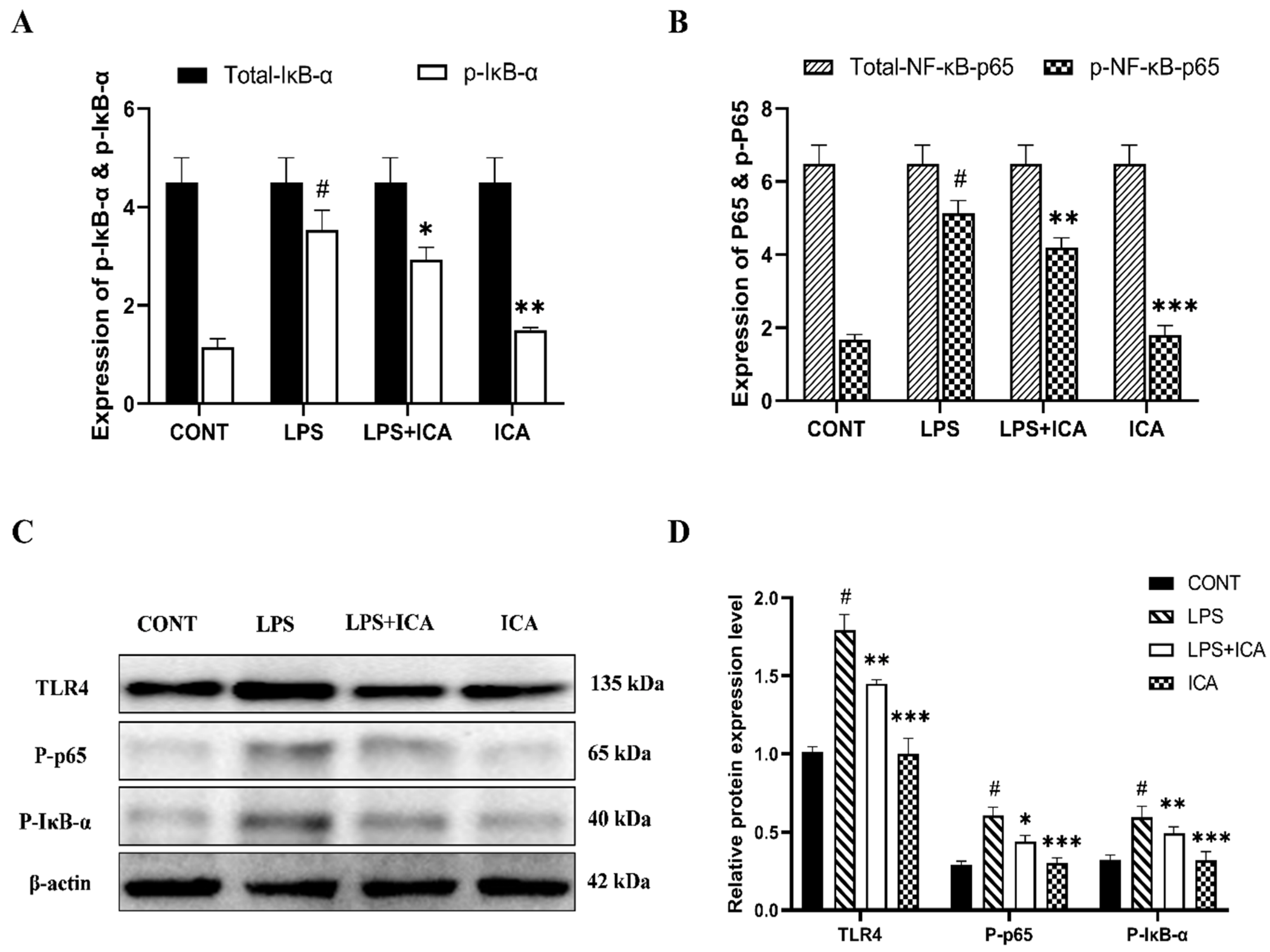
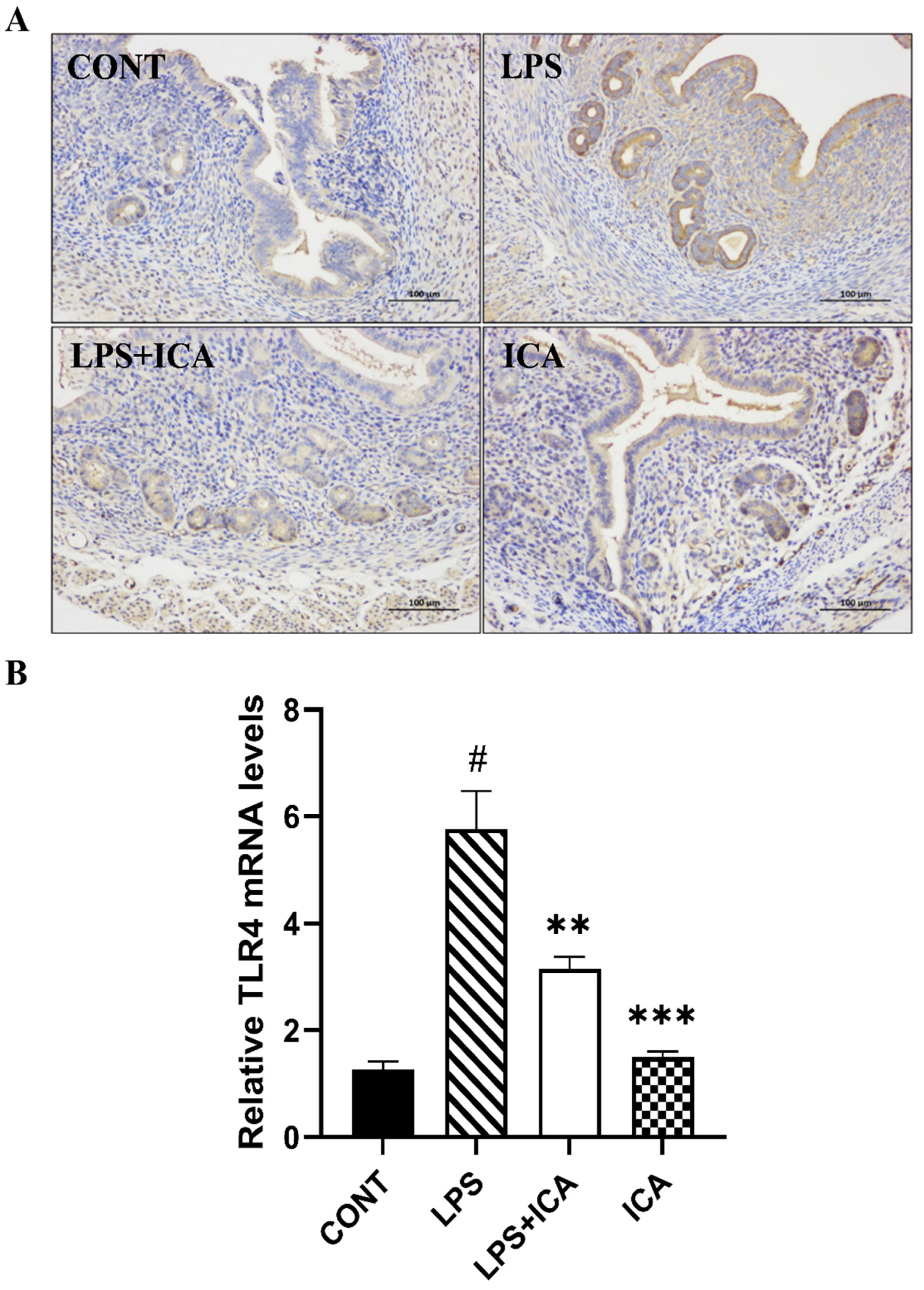
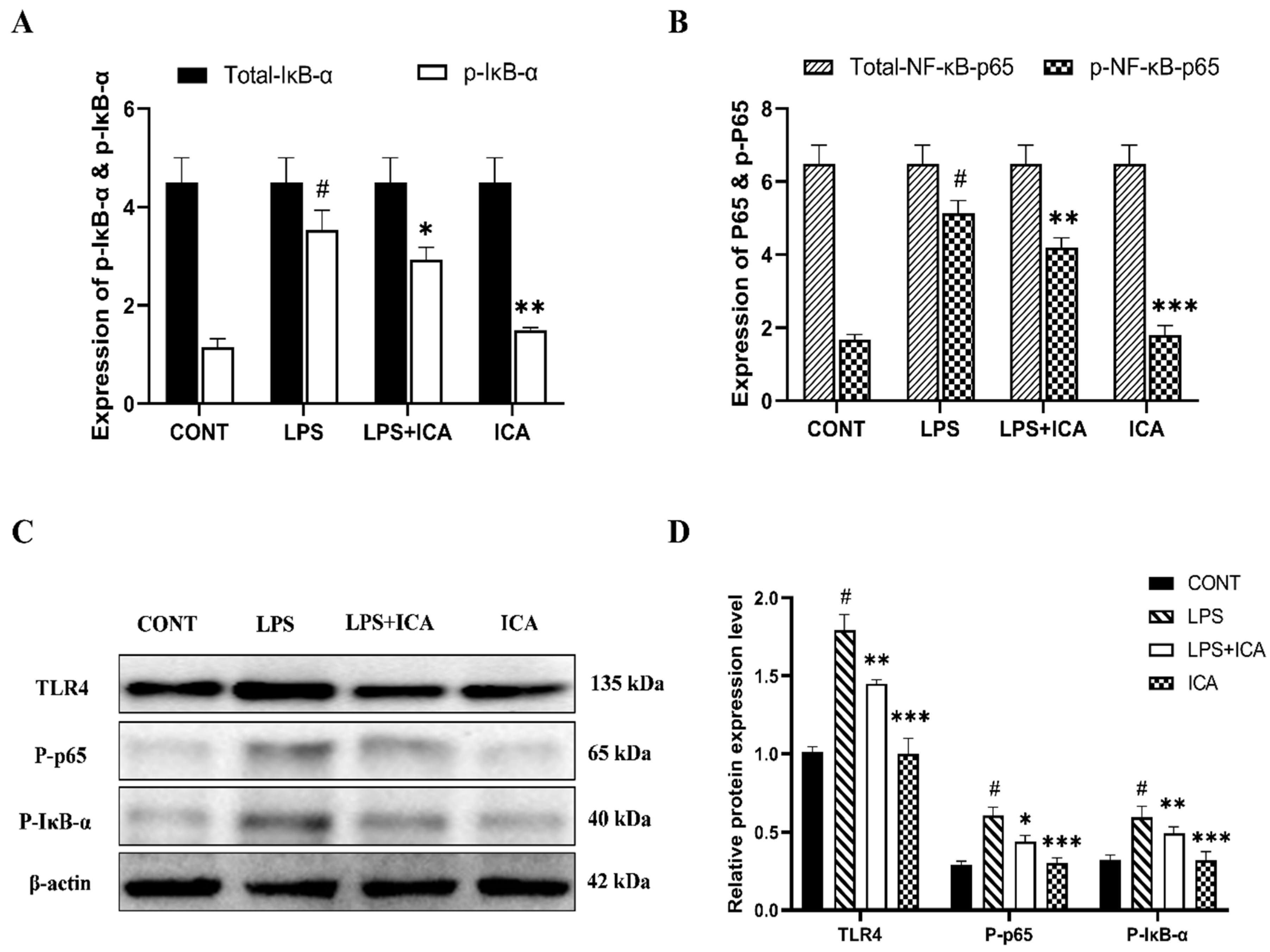
2.4. Effect of ICA on LPS-Triggered TLR4-Mediated NF-κB Pathway
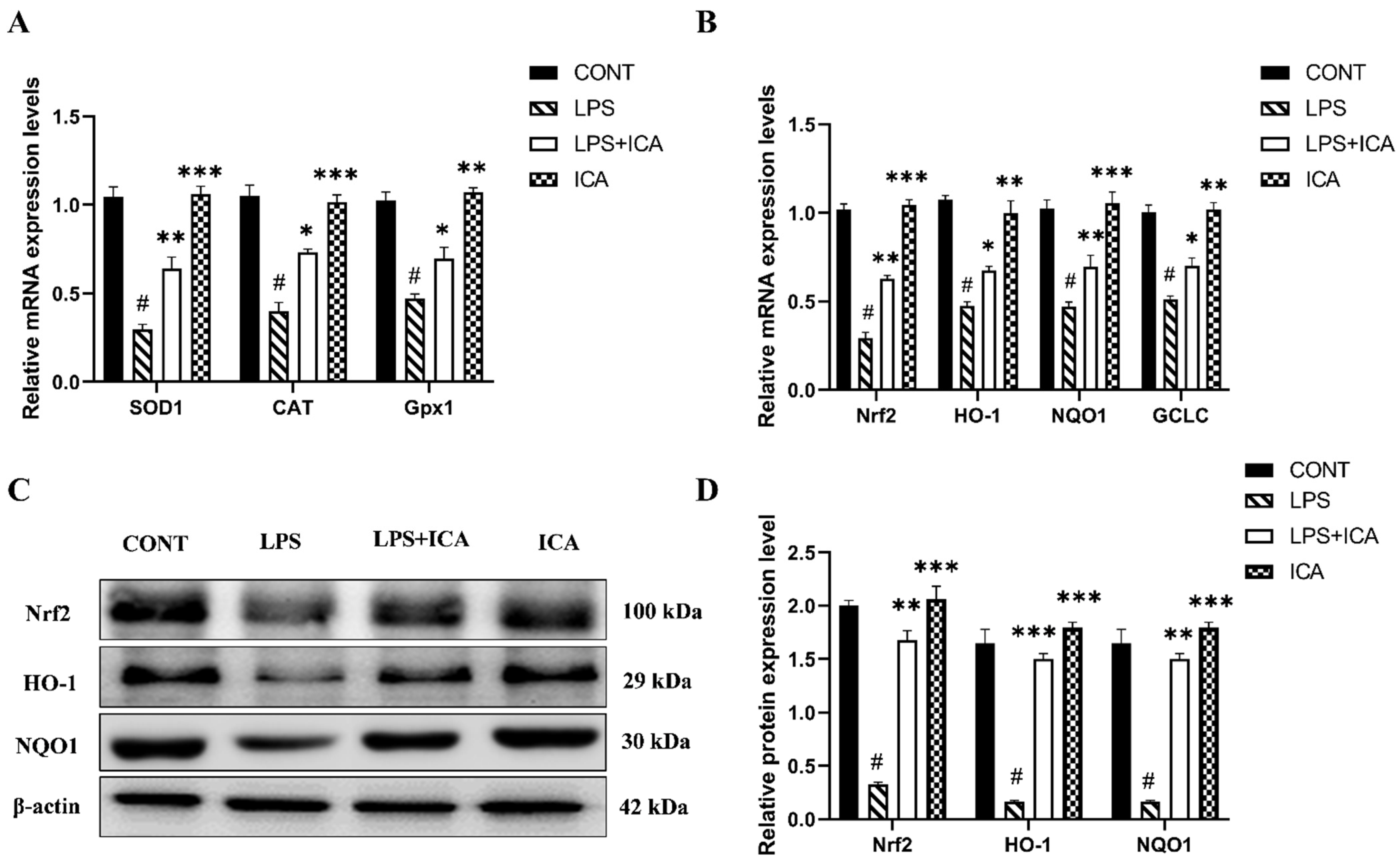
2.5. Effect of ICA on LPS-Triggered Oxidative Stress Markers in Endometritis
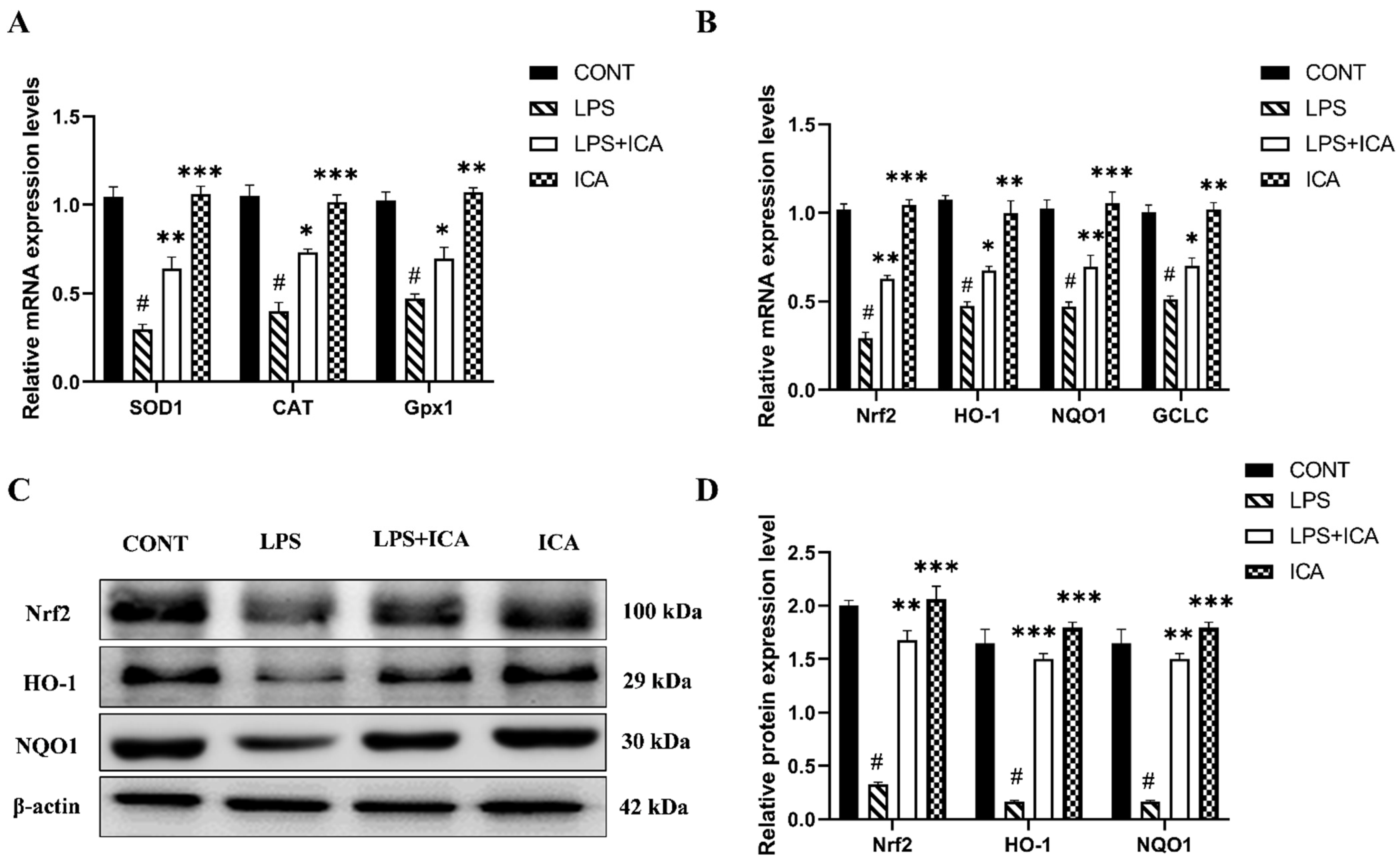
2.6. Effect of ICA against LPS-Triggered Activation of Nrf2 Pathway
3. Discussion
4. Materials and Methods
4.1. Ethical Statement
4.2. Reagents
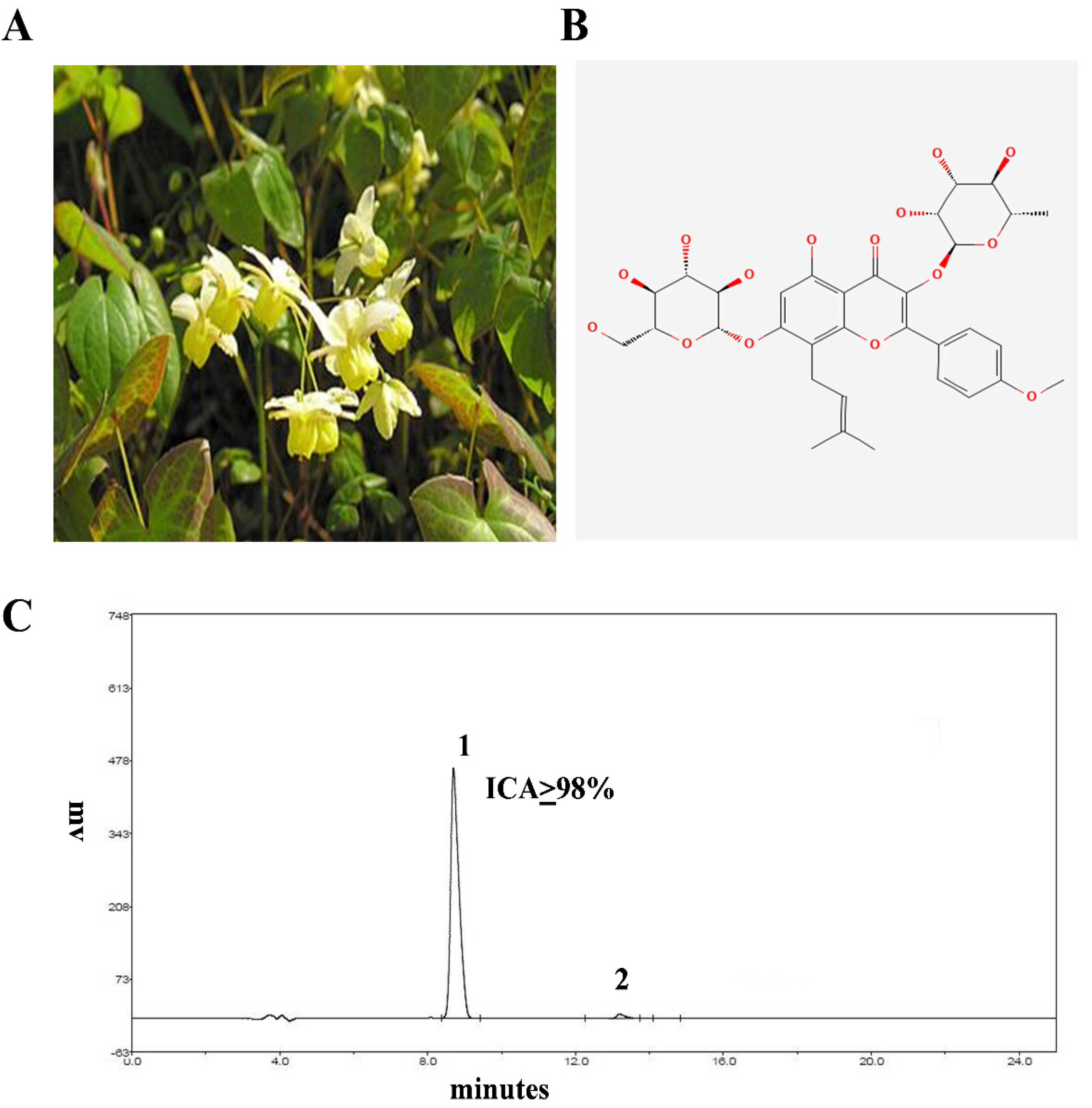
4.3. HPLC Analysis of ICA
4.4. Animals
4.5. Treatment Design
4.6. Histopathologic Assay of Uterine Tissue
4.7. Wet to Dry Weight (W/D) Ratio of Uterine Tissue
4.8. MPO Activity Assay and Determination of NO Concentration
4.9. ELISA Analysis
4.10. Determination of Oxidative Stress Markers
4.11. qRT-PCR Analysis
4.12. Immunohistochemistry
4.13. Western Blot Analysis
4.14. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jiang, K.; Guo, S.; Yang, J.; Liu, J.; Shaukat, A.; Zhao, G.; Wu, H.; Deng, G. Matrine alleviates Staphylococcus aureus lipoteichoic acid-induced endometritis via suppression of TLR2-mediated NF-κB activation. Int. Immunopharmacol. 2019, 70, 201–207. [Google Scholar] [CrossRef]
- Wu, H.; Zhao, G.; Jiang, K.; Li, C.; Qiu, C.; Deng, G. Engeletin alleviates lipopolysaccharide-induced endometritis in mice by inhibiting TLR4-mediated NF-κB activation. J. Agric. Food Chem. 2016, 64, 6171–6178. [Google Scholar] [CrossRef]
- Liu, J.; Guo, S.; Jiang, K.; Zhang, T.; Zhiming, W.; Yaping, Y.; Jing, Y.; Shaukat, A.; Deng, G. miR-488 mediates negative regulation of the AKT/NF-κB pathway by targeting Rac1 in LPS-induced inflammation. J. Cell. Physiol. 2020, 235, 4766–4777. [Google Scholar] [CrossRef]
- Wu, H.; Jiang, K.; Yin, N.; Ma, X.; Zhao, G.; Qiu, C.; Deng, G. Thymol mitigates lipopolysaccharide-induced endometritis by regulating the TLR4- and ROS-mediated NF-kappaB signaling pathways. Oncotarget 2017, 8, 20042–20055. [Google Scholar] [CrossRef]
- Lv, X.; Fu, K.; Li, W.; Wang, Y.; Wang, J.; Li, H.; Tian, W.; Cao, R. TIIA attenuates LPS-induced mouse endometritis by suppressing the NF-κB signaling pathway. Can. J. Physiol. Pharmacol. 2015, 93, 967–971. [Google Scholar] [CrossRef]
- Ibraimi, Z.; Shehi, A.; Murtezani, A.; Krasniqi, S.; Agani, Z. Kosovo’s Public Health Damage from Abusive Use of Antibiotics in Dairy Cattle. Mater. Socio-Med. 2015, 27, 149. [Google Scholar] [CrossRef]
- Wang, X.; Yuan, T.; Yin, N.; Ma, X.; Zhang, Z.; Zhu, Z.; Shaukat, A.; Deng, G. Luteoloside Protects the Uterus from Staphylococcus aureus-Induced Inflammation, Apoptosis, and Injury. Inflammation 2018, 41, 1702–1716. [Google Scholar] [CrossRef]
- Piras, C.; Guo, Y.; Soggiu, A.; Chanrot, M.; Greco, V.; Urbani, A.; Charpigny, G.; Bonizzi, L.; Roncada, P.; Humblot, P. Changes in protein expression profiles in bovine endometrial epithelial cells exposed to E. coli LPS challenge. Mol. BioSystems 2017, 13, 392–405. [Google Scholar] [CrossRef]
- Zhang, H.; Wu, Z.-M.; Yang, Y.-P.; Shaukat, A.; Yang, J.; Guo, Y.-F.; Zhang, T.; Zhu, X.-Y.; Qiu, J.-X.; Deng, G.-Z. Catalpol ameliorates LPS-induced endometritis by inhibiting inflammation and TLR4/NF-κB signaling. J. Zhejiang Univ. Sci. B 2019, 20, 816–827. [Google Scholar] [CrossRef]
- Zhao, G.; Jiang, K.; Yang, Y.; Zhang, T.; Wu, H.; Shaukat, A.; Qiu, C.; Deng, G. The potential therapeutic role of miR-223 in bovine endometritis by targeting the NLRP3 inflammasome. Front. Immunol. 2018, 9, 1916. [Google Scholar] [CrossRef] [Green Version]
- Dreyfuss, D.; Ricard, J.-D. Acute lung injury and bacterial infection. Clin. Chest Med. 2005, 26, 105–112. [Google Scholar] [CrossRef]
- Jiang, K.; Yang, J.; Xue, G.; Dai, A.; Wu, H. Fisetin ameliorates the inflammation and oxidative stress in lipopolysaccharide-induced endometritis. J. Inflamm. Res. 2021, 14, 2963. [Google Scholar] [CrossRef] [PubMed]
- Di, A.; Gao, X.-P.; Qian, F.; Kawamura, T.; Han, J.; Hecquet, C.; Richard, D.Y.; Vogel, S.M.; Malik, A.B. The redox-sensitive cation channel TRPM2 modulates phagocyte ROS production and inflammation. Nat. Immunol. 2012, 13, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Jiang, K.; Ye, W.; Bai, Q.; Cai, J.; Wu, H.; Li, X. Therapeutic Role of miR-30a in Lipoteichoic Acid-Induced Endometritis via Targeting the MyD88/Nox2/ROS Signaling. Oxidative Med. Cell. Longev. 2021, 2021, 5042048. [Google Scholar] [CrossRef] [PubMed]
- Shaukat, A.; Shaukat, I.; Rajput, S.A.; Shukat, R.; Hanif, S.; Jiang, K.; Zhang, T.; Akhtar, M.; Shaukat, I.; Ma, X. Ginsenoside Rb1 protects from Staphylococcus aureus-induced oxidative damage and apoptosis through endoplasmic reticulum-stress and death receptor-mediated pathways. Ecotoxicol. Environ. Saf. 2021, 219, 112353. [Google Scholar] [CrossRef] [PubMed]
- Shaukat, A.; Yang, C.; Yang, Y.; Guo, Y.-F.; Jiang, K.; Guo, S.; Liu, J.; Zhang, T.; Zhao, G.; Ma, X.; et al. Ginsenoside Rb 1: A novel therapeutic agent in Staphylococcus aureus-induced Acute Lung Injury with special reference to Oxidative stress and Apoptosis. Microb. Pathog. 2020, 143, 104109. [Google Scholar] [CrossRef] [PubMed]
- Jiang, K.; Guo, S.; Yang, C.; Yang, J.; Chen, Y.; Shaukat, A.; Zhao, G.; Wu, H.; Deng, G. Barbaloin protects against lipopolysaccharide (LPS)-induced acute lung injury by inhibiting the ROS-mediated PI3K/AKT/NF-κB pathway. Int. Immunopharmacol. 2018, 64, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Jiang, K.; Yang, J.; Song, C.; He, F.; Yang, L.; Li, X. Enforced expression of miR-92b blunts E. coli lipopolysaccharide-mediated inflammatory injury by activating the PI3K/AKT/β-catenin pathway via targeting PTEN. Int. J. Biol. Sci. 2021, 17, 1289. [Google Scholar] [CrossRef]
- Akhtar, M.; Shaukat, A.; Zahoor, A.; Chen, Y.; Wang, Y.; Yang, M.; Umar, T.; Guo, M.; Deng, G. Hederacoside-C inhibition of Staphylococcus aureus-induced mastitis via TLR2 & TLR4 and their downstream signaling NF-κB and MAPKs pathways in vivo and in vitro. Inflammation 2020, 43, 579–594. [Google Scholar]
- Akhtar, M.; Shaukat, A.; Zahoor, A.; Chen, Y.; Wang, Y.; Yang, M.; Umar, T.; Guo, M.; Deng, G. Anti-inflammatory effects of Hederacoside-C on Staphylococcus aureus induced inflammation via TLRs and their downstream signal pathway in vivo and in vitro. Microb. Pathog. 2019, 137, 103767. [Google Scholar] [CrossRef]
- Zeng, K.-W.; Fu, H.; Liu, G.-X.; Wang, X.-M. Icariin attenuates lipopolysaccharide-induced microglial activation and resultant death of neurons by inhibiting TAK1/IKK/NF-κB and JNK/p38 MAPK pathways. Int. Immunopharmacol. 2010, 10, 668–678. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.-R.; Vuorela, P.; Vuorela, H.; Hiltunen, R. Isolation and immunomodulating effect of flavonol glycosides from Epimedium hunanense. Planta Med. 1997, 63, 316–319. [Google Scholar] [CrossRef]
- Wang, Y.-K.; Huang, Z.-Q. Protective effects of icariin on human umbilical vein endothelial cell injury induced by H2O2 in vitro. Pharmacol. Res. 2005, 52, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Kong, L.; Xia, X.; Zhang, W.; Xia, Z.; Jiang, F. Antidepressant-like effect of icariin and its possible mechanism in mice. Pharmacol. Biochem. Behav. 2005, 82, 686–694. [Google Scholar] [CrossRef]
- Ding, L.; Liang, X.-G.; Hu, Y.; Zhu, D.-Y.; Lou, Y.-J. Involvement of p38MAPK and reactive oxygen species in icariin-induced cardiomyocyte differentiation of murine embryonic stem cells in vitro. Stem Cells Dev. 2008, 17, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Chung, B.-H.; Kim, J.-D.; Kim, C.-K.; Kim, J.H.; Won, M.-H.; Lee, H.-S.; Dong, M.-S.; Ha, K.-S.; Kwon, Y.-G.; Kim, Y.-M. Icariin stimulates angiogenesis by activating the MEK/ERK-and PI3K/Akt/eNOS-dependent signal pathways in human endothelial cells. Biochem. Biophys. Res. Commun. 2008, 376, 404–408. [Google Scholar] [CrossRef]
- Xu, C.-Q.; Liu, B.-J.; Wu, J.-F.; Xu, Y.-C.; Duan, X.-H.; Cao, Y.-X.; Dong, J.-C. Icariin attenuates LPS-induced acute inflammatory responses: Involvement of PI3K/Akt and NF-κB signaling pathway. Eur. J. Pharmacol. 2010, 642, 146–153. [Google Scholar] [CrossRef]
- Li, F.; Du, B.-W.; Lu, D.-F.; Wu, W.-X.; Wongkrajang, K.; Wang, L.; Pu, W.-C.; Liu, C.-L.; Liu, H.-W.; Wang, M.-K. Flavonoid glycosides isolated from Epimedium brevicornum and their estrogen biosynthesis-promoting effects. Sci. Rep. 2017, 7, 7760. [Google Scholar] [CrossRef]
- El-Shitany, N.A.; Eid, B.G. Icariin modulates carrageenan-induced acute inflammation through HO-1/Nrf2 and NF-kB signaling pathways. Biomed. Pharmacother. 2019, 120, 109567. [Google Scholar] [CrossRef]
- Yin, N.; Yang, Y.; Wang, X.; Yang, C.; Ma, X.; Shaukat, A.; Zhao, G.; Deng, G. MiR-19a mediates the negative regulation of the NF-κB pathway in lipopolysaccharide-induced endometritis by targeting TBK1. Inflamm. Res. 2019, 68, 231–240. [Google Scholar] [CrossRef]
- Shaukat, A.; Shaukat, I.; Rajput, S.A.; Shukat, R.; Hanif, S.; Shaukat, I.; Zhang, X.; Chen, C.; Sun, X.; Ye, T.; et al. Ginsenoside Rb1 Mitigates Escherichia coli Lipopolysaccharide-Induced Endometritis through TLR4-Mediated NF-κB Pathway. Molecules 2021, 26, 7089. [Google Scholar] [CrossRef] [PubMed]
- Ferrero-Miliani, L.; Nielsen, O.; Andersen, P.; Girardin, S. Chronic inflammation: Importance of NOD2 and NALP3 in interleukin-1β generation. Clin. Exp. Immunol. 2007, 147, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; He, S.; Yuan, J.; Miao, S.; Gao, H.; Zhang, J.; Li, Y.; Peng, W.; Wu, P. Lipoxin A4 attenuates LPS-induced mouse acute lung injury via Nrf2-mediated E-cadherin expression in airway epithelial cells. Free Radic. Biol. Med. 2016, 93, 52–66. [Google Scholar] [CrossRef] [PubMed]
- Umar, T.; Yin, B.; Umer, S.; Ma, X.; Jiang, K.; Umar, Z.; Akhtar, M.; Shaukat, A.; Deng, G. MicroRNA: Could It Play a Role in Bovine Endometritis? Inflammation 2021, 44, 1683–1695. [Google Scholar] [CrossRef]
- Li, W.; Fu, K.; Lv, X.; Wang, Y.; Wang, J.; Li, H.; Tian, W.; Cao, R. Lactoferrin suppresses lipopolysaccharide-induced endometritis in mice via down-regulation of the NF-κB pathway. Int. Immunopharmacol. 2015, 28, 695–699. [Google Scholar] [CrossRef]
- Jiang, P.-Y.; Zhu, X.-J.; Zhang, Y.-N.; Zhou, F.-F.; Yang, X.-F. Protective effects of apigenin on LPS-induced endometritis via activating Nrf2 signaling pathway. Microb. Pathog. 2018, 123, 139–143. [Google Scholar] [CrossRef]
- Miller, A.M.; McPhaden, A.R.; Preston, A.; Wadsworth, R.M.; Wainwright, C.L. TNFα increases the inflammatory response to vascular balloon injury without accelerating neointimal formation. Atherosclerosis 2005, 179, 51–59. [Google Scholar] [CrossRef]
- Xiong, W.; Ma, H.; Zhang, Z.; Jin, M.; Wang, J.; Xu, Y.; Wang, Z. The protective effect of icariin and phosphorylated icariin against LPS-induced intestinal goblet cell dysfunction. Innate Immun. 2020, 26, 97–106. [Google Scholar] [CrossRef]
- Tak, P.P.; Firestein, G.S. NF-κB: A key role in inflammatory diseases. J. Clin. Investig. 2001, 107, 7–11. [Google Scholar] [CrossRef]
- Shaukat, A.; Guo, Y.F.; Jiang, K.; Zhao, G.; Wu, H.; Zhang, T.; Yang, Y.; Guo, S.; Yang, C.; Zahoor, A.; et al. Ginsenoside Rb1 ameliorates Staphylococcus aureus-induced Acute Lung Injury through attenuating NF-kappaB and MAPK activation. Microb. Pathog. 2019, 132, 302–312. [Google Scholar] [CrossRef]
- Yingkun, N.; Zhenyu, W.; Jing, L.; Xiuyun, L.; Huimin, Y. Stevioside protects LPS-induced acute lung injury in mice. Inflammation 2013, 36, 242–250. [Google Scholar] [CrossRef]
- Chen, Y.; Jin, S.; Teng, X.; Hu, Z.; Zhang, Z.; Qiu, X.; Tian, D.; Wu, Y. Hydrogen sulfide attenuates LPS-induced acute kidney injury by inhibiting inflammation and oxidative stress. Oxidative Med. Cell. Longev. 2018, 2018, 6717212. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.; Li, L.; Lu, S.; Li, K.; Su, Z.; Wang, Y.; Fan, X.; Li, X.; Zhao, G. The protective effect of dexmedetomidine on LPS-induced acute lung injury through the HMGB1-mediated TLR4/NF-κB and PI3K/Akt/mTOR pathways. Mol. Immunol. 2018, 94, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.-Y.; Lin, D.P.-C.; Wu, C.-Y.; Teng, M.-C.; Sun, C.-Y.; Tsai, Y.-T.; Su, K.-C.; Wang, S.-R.; Chang, H.-H. Dietary zerumbone prevents mouse cornea from UVB-induced photokeratitis through inhibition of NF-κB, iNOS, and TNF-α expression and reduction of MDA accumulation. Mol. Vis. 2011, 17, 854. [Google Scholar]
- Leung, W.-S.; Yang, M.-L.; Lee, S.-S.; Kuo, C.-W.; Ho, Y.-C.; Huang-Liu, R.; Lin, H.-W.; Kuan, Y.-H. Protective effect of zerumbone reduces lipopolysaccharide-induced acute lung injury via antioxidative enzymes and Nrf2/HO-1 pathway. Int. Immunopharmacol. 2017, 46, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Kong, M.; Mao, J.; Luo, B.; Qin, Z. Role of transcriptional factor Nrf2 in the acute lung injury of mice. Int. J. Clin. Exp. Pathol. 2015, 8, 10929. [Google Scholar]
- Moffit, J.S.; Aleksunes, L.M.; Kardas, M.J.; Slitt, A.L.; Klaassen, C.D.; Manautou, J.E. Role of NAD (P) H: Quinone oxidoreductase 1 in clofibrate-mediated hepatoprotection from acetaminophen. Toxicology 2007, 230, 197–206. [Google Scholar] [CrossRef]
- Otterbein, L.E.; Choi, A.M. Heme oxygenase: Colors of defense against cellular stress. Am. J. Physiol. Lung Cell. Mol. Physiol. 2000, 279, L1029–L1037. [Google Scholar] [CrossRef]
- Luo, H.; Zhang, R. Icariin enhances cell survival in lipopolysaccharide-induced synoviocytes by suppressing ferroptosis via the Xc-/GPX4 axis. Exp. Ther. Med. 2021, 21, 72. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhu, G.; He, J.; Wang, G.; Li, D.; Zhang, F. Icariin targets Nrf2 signaling to inhibit microglia-mediated neuroinflammation. Int. Immunopharmacol. 2019, 73, 304–311. [Google Scholar] [CrossRef]
- Hu, L.; Li, L.; Zhang, H.; Li, Q.; Jiang, S.; Qiu, J.; Sun, J.; Dong, J. Inhibition of airway remodeling and inflammatory response by Icariin in asthma. BMC Complementary Altern. Med. 2019, 19, 316. [Google Scholar] [CrossRef] [PubMed]
- Rajput, S.A.; Shaukat, A.; Wu, K.; Rajput, I.R.; Baloch, D.M.; Akhtar, R.W.; Raza, M.A.; Najda, A.; Rafał, P.; Albrakati, A. Luteolin Alleviates AflatoxinB1-Induced Apoptosis and Oxidative Stress in the Liver of Mice through Activation of Nrf2 Signaling Pathway. Antioxidants 2021, 10, 1268. [Google Scholar] [CrossRef] [PubMed]
- Rajput, S.A.; Shaukat, A.; Rajput, I.R.; Kamboh, A.A.; Iqbal, Z.; Saeed, M.; Akhtar, R.W.; Shah, S.A.H.; Raza, M.A.; El Askary, A. Ginsenoside Rb1 prevents deoxynivalenol-induced immune injury via alleviating oxidative stress and apoptosis in mice. Ecotoxicol. Environ. Saf. 2021, 220, 112333. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | CONT | LPS | LPS + ICA | ICA |
|---|---|---|---|---|
| ROS (florescence/mg protein) | 69.23 ± 4.11 | 276.63 ± 24.64 # | 180.19 ± 11.41 ** | 78.64 ± 6.49 **** |
| MDA (nmol/mg protein) | 2.11 ± 0.31 | 5.47 ± 0.58 # | 4.16 ± 0.33 * | 1.99 ± 0.18 *** |
| SOD (U/mg protein) | 41.85 ± 5.44 | 13.73 ± 2.14 # | 27.41 ± 6.97 ** | 43.44 ± 4.67 *** |
| CAT (U/mg protein) | 69.73 ± 6.45 | 21.07 ± 5.61 # | 47.79 ± 3.31 ** | 71.34 ± 7.41 *** |
| Gpx1 (U/mg protein) | 199.22 ± 14.42 | 69.57 ± 5.19 # | 127.89 ± 11.18 ** | 205.01 ± 10.29 *** |
| Treatments | Hours | ||||
|---|---|---|---|---|---|
| 0 | 24 | 30 | 36 | 42 | |
| CONT | PBS | _ | _ | _ | Euthanized |
| LPS | LPS | _ | _ | _ | Euthanized |
| LPS + ICA | LPS | ICA | ICA | ICA | Euthanized |
| ICA | PBS | ICA | ICA | ICA | Euthanized |
| Target Gene | Primer | Primer Sequence (5’  3’) 3’) | Accession No. | Product Size |
|---|---|---|---|---|
| TLR4 | Forward | ATTCAGAGCCGTTGGTGTATC | NM_021297.2 | 109 |
| Reverse | GGGACTTCTCAACCTTCTCAAG | |||
| TNF-α | Forward | GGGCTTTACCTCATCTACTCA | NM_013693.3 | 198 |
| Reverse | GCTCTTGATGGCAGACAGG | |||
| IL-1β | Forward | CCTGGGCTGTCCTGATGAGAG | NM_008361.4 | 131 |
| Reverse | TCCACGGGAAAGACACAGGTA | |||
| IL-6 | Forward | GGCGGATCGGATGTTGTGAT | NM_031168.1 | 199 |
| Reverse | GGACCCCAGACAATCGGTTG | |||
| IL-10 | Forward | ACAGCCGGGAAGACAATAACT | NM_010548.2 | 66 |
| Reverse | GCAGCTCTAGGAGCATGTGG | |||
| SOD1 | Forward | GGTCTCCAACATGCCTCTCT | NM_011434.2 | 203 |
| Reverse | AACCATCCACTTCGAGCAGA | |||
| CAT | Forward | CACTGACGAGATGGCACACT | NM_009804.2 | 175 |
| Reverse | TGTGGAGAATCGAACGGCAA | |||
| Gpx1 | Forward | GTACTTGGGGTCGGTCATGA | NM_001329527.1 | 222 |
| Reverse | GGTTTCCCGTGCAATCAGTT | |||
| Nrf2 | Forward | TCCTATGCGTGAATCCCAAT | NM_010902.3 | 103 |
| Reverse | GCGGCTTGAATGTTTGTCTT | |||
| HO-1 | Forward | GGGCTGTGAACTCTGTCCAATGT | NM_010442.2 | 162 |
| Reverse | TTGGTGAGGGAACTGTGTCAGG | |||
| NQO1 | Forward | TTCTGTGGCTTCCAGGTCTTAG | NM_008706.5 | 156 |
| Reverse | GTCAAACAGGCTGCTTGGAGCAA | |||
| GCLC | Forward | ACAAGGACGTGCTCAAGTGG | NM_010295.2 | 199 |
| Reverse | CCAGGCGTTCCTTCGATCAT | |||
| GAPDH | Forward | CAATGTGTCCGTCGTGGATCT | NM_001289726.1 | 124 |
| Reverse | GTCCTCAGTGTAGCCCAAGATG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shaukat, A.; Shaukat, I.; Rajput, S.A.; Shukat, R.; Hanif, S.; Huang, S.; Aleem, M.T.; Li, K.; Li, Q.; Chen, C.; et al. Icariin Alleviates Escherichia coli Lipopolysaccharide-Mediated Endometritis in Mice by Inhibiting Inflammation and Oxidative Stress. Int. J. Mol. Sci. 2022, 23, 10219. https://doi.org/10.3390/ijms231810219
Shaukat A, Shaukat I, Rajput SA, Shukat R, Hanif S, Huang S, Aleem MT, Li K, Li Q, Chen C, et al. Icariin Alleviates Escherichia coli Lipopolysaccharide-Mediated Endometritis in Mice by Inhibiting Inflammation and Oxidative Stress. International Journal of Molecular Sciences. 2022; 23(18):10219. https://doi.org/10.3390/ijms231810219
Chicago/Turabian StyleShaukat, Aftab, Irfan Shaukat, Shahid Ali Rajput, Rizwan Shukat, Sana Hanif, Shucheng Huang, Muhammad Tahir Aleem, Kun Li, Qiao Li, Chao Chen, and et al. 2022. "Icariin Alleviates Escherichia coli Lipopolysaccharide-Mediated Endometritis in Mice by Inhibiting Inflammation and Oxidative Stress" International Journal of Molecular Sciences 23, no. 18: 10219. https://doi.org/10.3390/ijms231810219
APA StyleShaukat, A., Shaukat, I., Rajput, S. A., Shukat, R., Hanif, S., Huang, S., Aleem, M. T., Li, K., Li, Q., Chen, C., Zhang, X., Lv, H., Yao, Z., & Yang, L. (2022). Icariin Alleviates Escherichia coli Lipopolysaccharide-Mediated Endometritis in Mice by Inhibiting Inflammation and Oxidative Stress. International Journal of Molecular Sciences, 23(18), 10219. https://doi.org/10.3390/ijms231810219