
Detection of Unprecedented CYP74 Enzyme in Mammal: Hydroperoxide Lyase CYP74C44 of the Bat Sturnira hondurensis

Abstract
:1. Introduction
2. Results
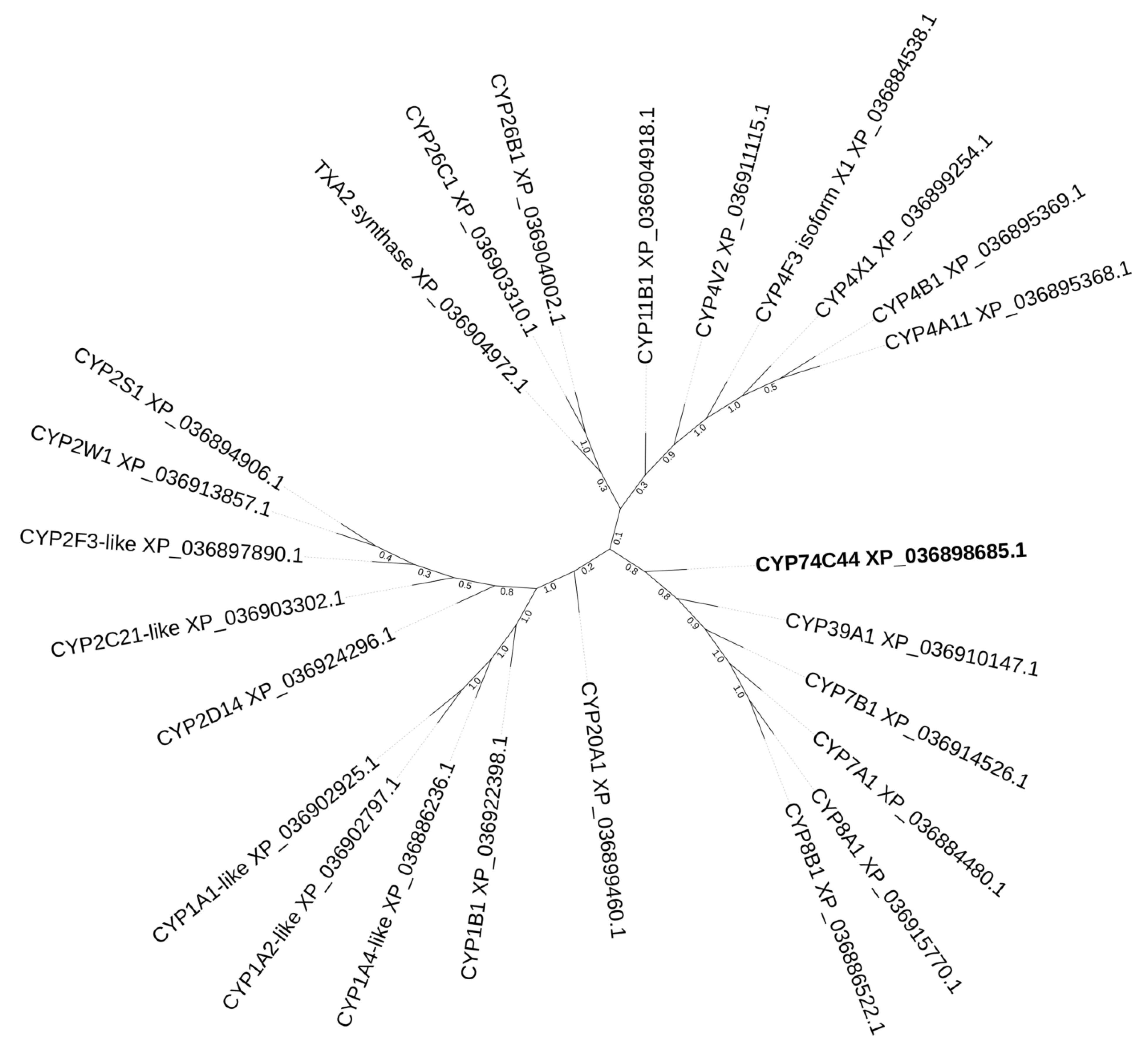
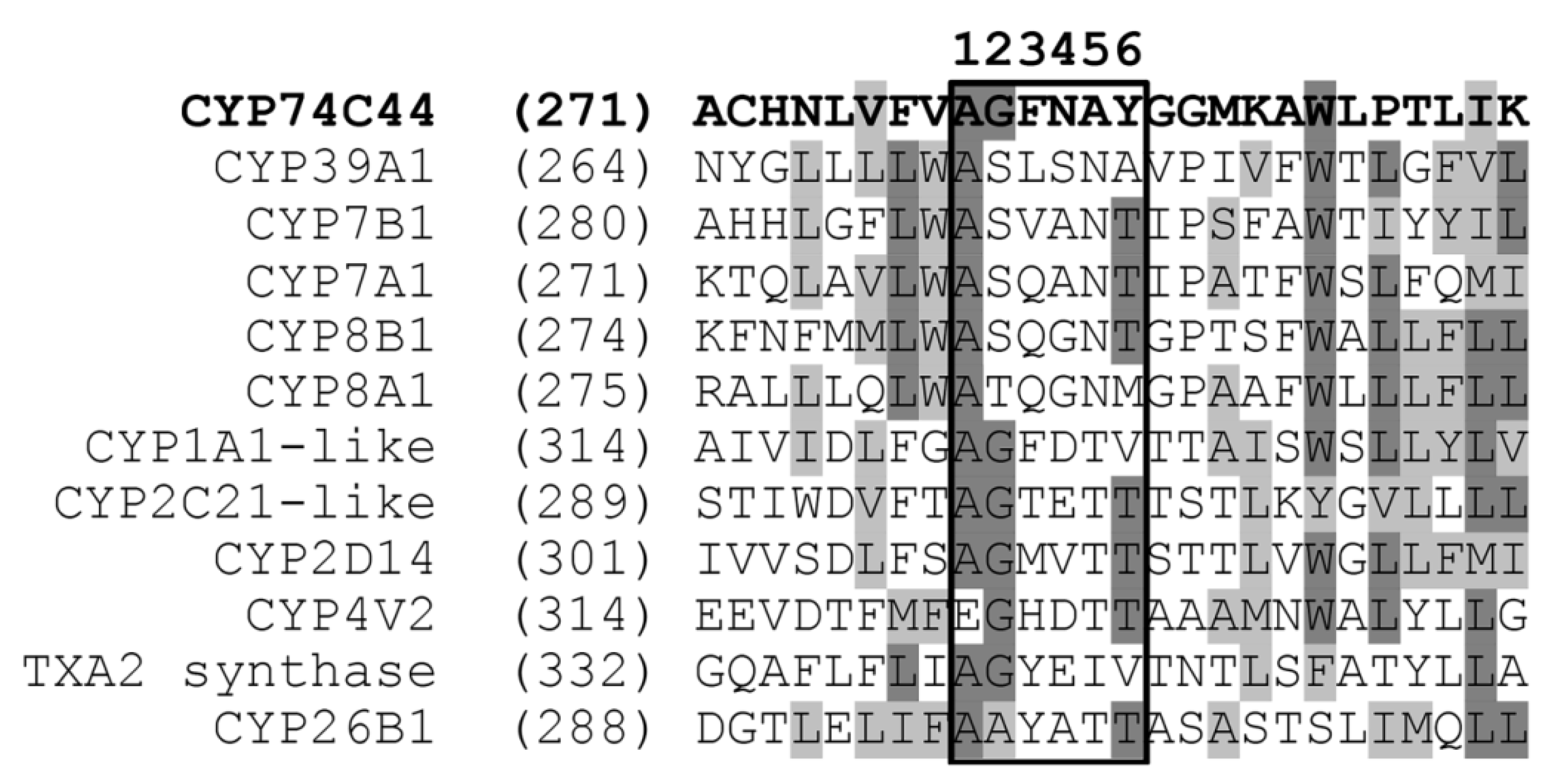
2.1. Bioinformatics Analyses
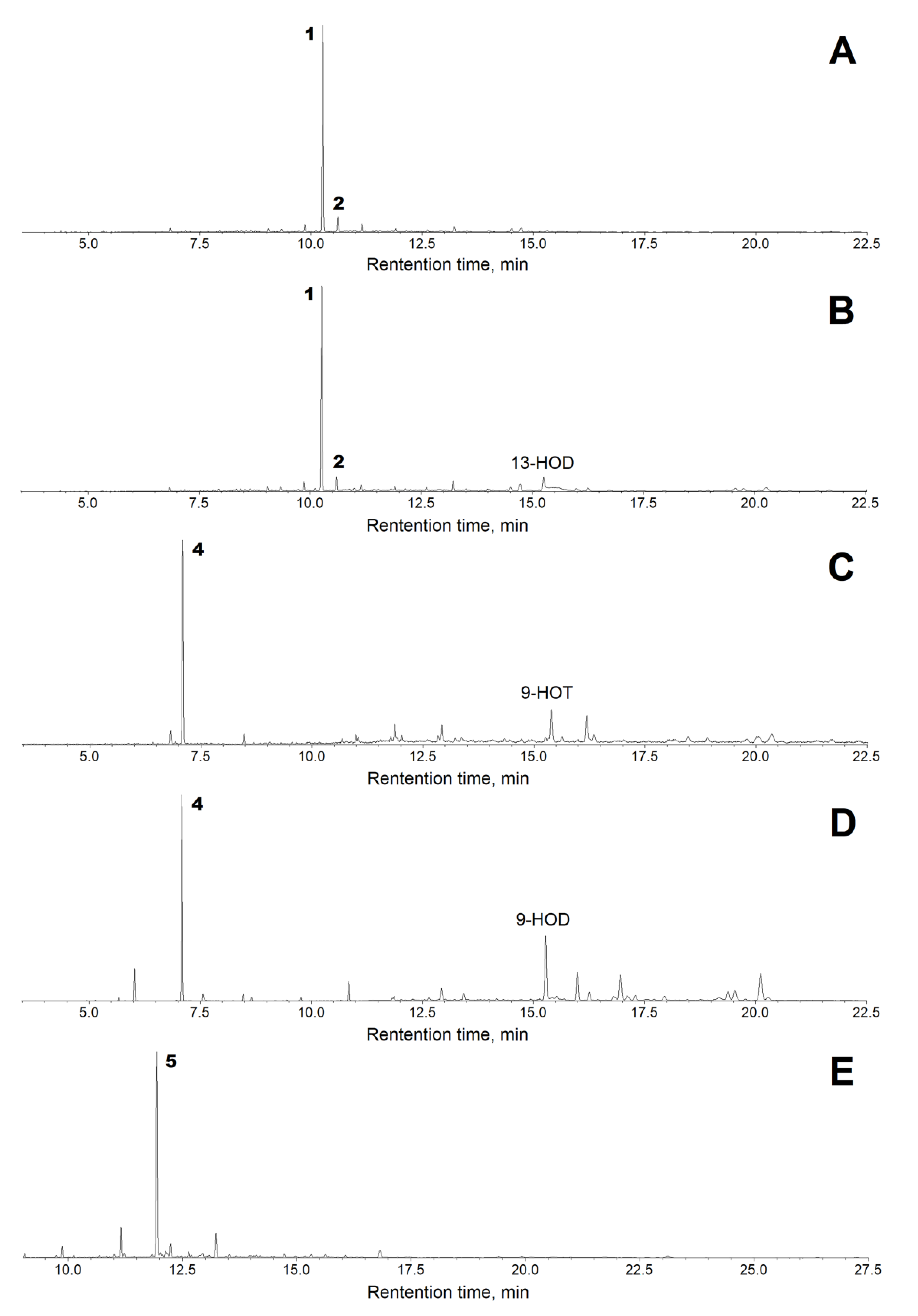
2.2. Kinetics and Substrate Specificity of the Recombinant CYP74C44
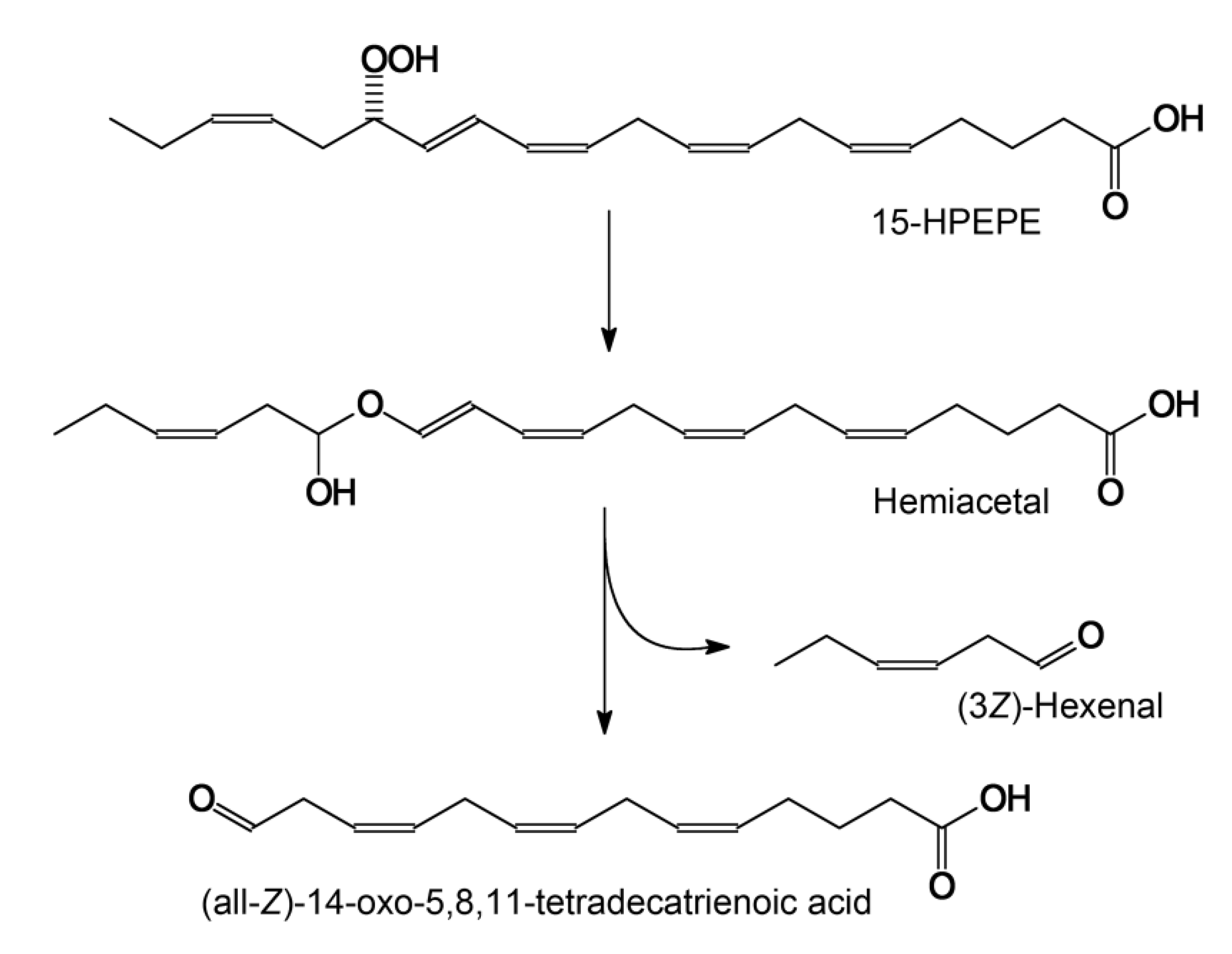
2.3. Substrate and Product Specificities of the Recombinant CYP74C44
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Bioinformatic Methods
4.3. Expression and Purification of Recombinant Enzyme
4.4. Kinetic Studies
4.5. Incubations of Recombinant Enzyme with Substrates
4.6. Methods of Spectral Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Denisov, I.G.; Makris, T.M.; Sligar, S.G.; Schlichting, I. Structure and chemistry of cytochrome P450. Chem. Rev. 2005, 105, 2253–2278. [Google Scholar] [CrossRef] [PubMed]
- Werck-Reichhart, D.; Feyereisen, R. Cytochromes P450: A success story. Genome Biol. 2000, 1, REVIEWS3003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, D.R.; Schuler, M.A.; Paquette, S.M.; Werck-Reichhart, D.; Bak, S. Comparative genomics of rice and Arabidopsis. Analysis of 727 cytochrome P450 genes and pseudogenes from a monocot and a dicot. Plant Physiol. 2004, 135, 756–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansuy, D.; Renaud, J.P. Heme-thiolate proteins different from cytochromes P450 catalyzing monooxygenations. In Cytochrome P450: Structure, Mechanism, and Biochemistry, 2nd ed.; Ortiz de Montellano, P.R., Ed.; Plenum Press: New York, NY, USA, 1995; pp. 537–574. [Google Scholar]
- Guengerich, F.P.; Munro, A.W. Unusual cytochrome P450 enzymes and reactions. J. Biol. Chem. 2013, 288, 17065–17073. [Google Scholar] [CrossRef] [Green Version]
- Poulos, T.L. Heme enzyme structure and function. Chem. Rev. 2014, 114, 3919–3962. [Google Scholar] [CrossRef] [Green Version]
- Hughes, R.K.; De Domenico, S.; Santino, A. Plant cytochrome CYP74 family: Biochemical features, endocellular localisation, activation mechanism in plant defence and improvements for industrial applications. ChemBioChem 2009, 10, 1122–1133. [Google Scholar] [CrossRef]
- Brash, A.R. Mechanistic aspects of CYP74 allene oxide synthases and related cytochrome P450 enzymes. Phytochemistry 2009, 70, 1522–1531. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.S.; Nioche, P.; Hamberg, M.; Raman, C.S. Structural insights into the evolutionary paths of oxylipin biosynthetic enzymes. Nature 2008, 455, 363–368. [Google Scholar] [CrossRef]
- Wang, K.; Tian, S.; Galindo-González, J.; Dávalos, L.M.; Zhang, Y.; Zhao, H. Molecular adaptation and convergent evolution of frugivory in Old World and neotropical fruit bats. Mol. Ecol. 2020, 29, 4366–4381. [Google Scholar] [CrossRef]
- Toporkova, Y.Y.; Askarova, E.K.; Gorina, S.S.; Ogorodnikova, A.V.; Mukhtarova, L.S.; Grechkin, A.N. Epoxyalcohol synthase activity of the CYP74B enzymes of higher plants. Biochim. Biophys. Acta 2020, 1865, 158743. [Google Scholar] [CrossRef]
- Grechkin, A.N. Hydroperoxide lyase and divinyl ether synthase. Prostaglandins Other Lipid Mediat. 2002, 68–69, 457–470. [Google Scholar] [CrossRef]
- Matsui, K. Green leaf volatiles: Hydroperoxide lyase pathway of oxylipin metabolism. Curr. Opin. Plant Biol. 2006, 9, 274–280. [Google Scholar] [CrossRef]
- Tijet, N.; Schneider, C.; Muller, B.L.; Brash, A.R. Biogenesis of volatile aldehydes from fatty acid hydroperoxides: Molecular cloning of a hydroperoxide lyase (CYP74C) with specificity for both the 9-and 13-hydroperoxides of linoleic and linolenic acids. Arch. Biochem. Biophys. 2001, 386, 281–289. [Google Scholar] [CrossRef]
- Kuroda, H.; Oshima, T.; Kaneda, H.; Takashio, M. Identification and functional analyses of two cDNAs that encode fatty acid 9-/13-hydroperoxide lyase (CYP74C) in rice. Biosci. Biotechnol. Biochem. 2005, 69, 1545–1554. [Google Scholar] [CrossRef] [Green Version]
- Mita, G.; Quarta, A.; Fasano, P.; De Paolis, A.; Di Sansebastiano, G.P.; Perrotta, C.; Iannacone, R.; Belfield, E.; Hughes, R.; Tsesmetzis, N.; et al. Molecular cloning and characterization of an almond 9-hydroperoxide lyase, a new CYP74 targeted to lipid bodies. J. Exp. Bot. 2005, 56, 2321–2333. [Google Scholar] [CrossRef] [Green Version]
- Grechkin, A.N.; Brühlmann, F.; Mukhtarova, L.S.; Gogolev, Y.V.; Hamberg, M. Hydroperoxide lyases (CYP74C and CYP74B) catalyze the homolytic isomerization of fatty acid hydroperoxides into hemiacetals. Biochim. Biophys. Acta 2006, 1761, 1419–1428. [Google Scholar] [CrossRef]
- Toporkova, Y.Y.; Gorina, S.S.; Bessolitsyna, E.K.; Smirnova, E.O.; Fatykhova, V.S.; Brühlmann, F.; Ilyina, T.M.; Mukhtarova, L.S.; Grechkin, A.N. Double function hydroperoxide lyases/epoxyalcohol synthases (CYP74C) of higher plants: Identification and conversion into allene oxide synthases by site-directed mutagenesis. Biochim. Biophys. Acta 2018, 1863, 369–378. [Google Scholar] [CrossRef]
- Gorshkov, V.Y.; Toporkova, Y.Y.; Tsers, I.D.; Smirnova, E.O.; Ogorodnikova, A.V.; Gogoleva, N.E.; Parfirova, O.I.; Petrova, O.E.; Gogolev, Y.V. Differential modulation of the lipoxygenase cascade during typical and latent Pectobacterium atrosepticum infections. Ann. Bot. 2022, 129, 271–286. [Google Scholar] [CrossRef]
- Zhu, B.Q.; Xu, X.Q.; Wu, Y.W.; Duan, C.Q.; Pan, Q.H. Isolation and characterization of two hydroperoxide lyase genes from grape berries. Mol. Biol. Rep. 2012, 39, 7443–7455. [Google Scholar] [CrossRef]
- Itoh, A.; Schilmiller, A.L.; McCaig, B.C.; Howe, G.A. Identification of a jasmonate-regulated allene oxide synthase that metabolizes 9-hydroperoxides of linoleic and linolenic acids. J. Biol. Chem. 2002, 277, 46051–46058. [Google Scholar] [CrossRef] [Green Version]
- Stumpe, M.; Göbel, C.; Demchenko, K.; Hoffmann, M.; Klösgen, R.B.; Pawlowski, K.; Feussner, I. Identification of an allene oxide synthase (CYP74C) that leads to formation of α-ketols from 9-hydroperoxides of linoleic and linolenic acid in below-ground organs of potato. Plant J. 2006, 47, 883–896. [Google Scholar] [CrossRef]
- Grechkin, A.N.; Mukhtarova, L.S.; Latypova, L.R.; Gogolev, Y.; Toporkova, Y.Y.; Hamberg, M. Tomato CYP74C3 is a multifunctional enzyme not only synthesizing allene oxide but also catalyzing its hydrolysis and cyclization. ChemBioChem 2008, 9, 2498–2505. [Google Scholar] [CrossRef]
- Brash, A.R.; Boeglin, W.E.; Stec, D.F.; Voehler, M.; Schneider, C.; Cha, J.K. Isolation and characterization of two geometric allene oxide isomers synthesized from 9S-hydroperoxylinoleic acid by cytochrome P450 CYP74C3: Stereochemical assignment of natural fatty acid allene oxides. J. Biol. Chem. 2013, 288, 20797–20806. [Google Scholar] [CrossRef] [Green Version]
- Toporkova, Y.Y.; Smirnova, E.O.; Lantsova, N.V.; Mukhtarova, L.S.; Grechkin, A.N. Detection of the First Epoxyalcohol Synthase/Allene Oxide Synthase (CYP74 Clan) in the Lancelet (Branchiostoma belcheri, Chordata). Int. J. Mol. Sci. 2021, 22, 4737. [Google Scholar] [CrossRef]
- Toporkova, Y.Y.; Gorina, S.S.; Mukhitova, F.K.; Hamberg, M.; Ilyina, T.M.; Mukhtarova, L.S.; Grechkin, A.N. Identification of CYP443D1 (CYP74 clan) of Nematostella vectensis as a first cnidarian epoxyalcohol synthase and insights into its catalytic mechanism. Biochim. Biophys. Acta 2017, 1862, 1099–1109. [Google Scholar] [CrossRef]
- Gorina, S.S.; Toporkova, Y.Y.; Mukhtarova, L.S.; Grechkin, A.N. The CYP443C1 (CYP74 clan) cytochrome of sea anemone Nematostella vectensis—The first metazoan enzyme possessing hydroperoxide lyase/epoxyalcohol synthase activity. Dokl. Biochem. Biophys. 2019, 486, 192–196. [Google Scholar] [CrossRef]
- Grechkin, A.N.; Hamberg, M. The “heterolytic hydroperoxide lyase” is an isomerase producing a short-lived fatty acid hemiacetal. Biochim. Biophys. Acta 2004, 1636, 47–58. [Google Scholar] [CrossRef]
- Mukhtarova, L.S.; Brühlmann, F.; Hamberg, M.; Khairutdinov, B.I.; Grechkin, A.N. Plant hydroperoxide-cleaving enzymes (CYP74 family) function as hemiacetal synthases: Structural proof of hemiacetals by NMR spectroscopy. Biochim. Biophys. Acta 2018, 1863, 1316–1322. [Google Scholar] [CrossRef] [PubMed]
- Gölzer, P.; Janzowski, C.; Pool-Zobel, B.L.; Eisenbrand, G. (E)-2-Hexenal-Induced DNA Damage and Formation of Cyclic 1, N2-(1, 3-Propano)-2’-deoxyguanosine Adducts in Mammalian Cells. Chem. Res. Toxicol. 1996, 9, 1207–1213. [Google Scholar] [CrossRef] [PubMed]
- Padhi, S.; Dias, I.; Korn, V.L.; Bennett, J.W. Pseudogymnoascus destructans: Causative agent of white-nose syndrome in bats is inhibited by safe volatile organic compounds. J. Fungi 2018, 4, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korn, V.L.; Pennerman, K.K.; Padhi, S.; Bennett, J.W. Trans-2-hexenal downregulates several pathogenicity genes of Pseudogymnoascus destructans, the causative agent of white-nose syndrome in bats. J. Ind. Microbiol. Biotechnol. 2021, 48, kuab060. [Google Scholar] [CrossRef]
- Corey, E.J.; Iguchi, S.; Albright, J.O.; De, B. Studies on the conformational mobility of arachidonic acid. Facile macrolactonization of 20-hydroxyarachidonic acid. Tetrahedron Lett. 1983, 24, 37–40. [Google Scholar] [CrossRef]
- Manna, S.; Falck, J.R.; Chacos, N.; Capdevila, J. Synthesis of arachidonic acid metabolites produced by purified kidney cortex microsomal cytochrome P-450. Tetrahedron Lett. 1983, 24, 33–36. [Google Scholar] [CrossRef]
- Wilson, R.A.; Gardner, H.W.; Keller, N.P. Cultivar-dependent expression of a maize lipoxygenase responsive to seed infesting fungi. Mol. Plant-Microbe Interact. 2001, 14, 980–987. [Google Scholar] [CrossRef] [Green Version]
- Chechetkin, I.R.; Osipova, E.V.; Tarasova, N.B.; Mukhitova, F.K.; Hamberg, M.; Gogolev, Y.V.; Grechkin, A.N. Specificity of oxidation of linoleic acid homologs by plant lipoxygenases. Biochemistry 2009, 74, 855–861. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Zuckerkandl, E.; Pauling, L. Evolutionary divergence and convergence in proteins. In Evolving Genes and Proteins; Bryson, V., Vogel, H.J., Eds.; Academic Press: New York, NY, USA, 1965; pp. 97–166. [Google Scholar]
- Gorina, S.S.; Mukhitova, F.K.; Ilyina, T.M.; Toporkova, Y.Y.; Grechkin, A.N. Detection of unprecedented allene oxide synthase member of CYP74B subfamily: CYP74B33 of carrot (Daucus carota). Biochim. Biophys. Acta 2019, 1864, 1580–1590. [Google Scholar] [CrossRef]
- Gogolev, Y.V.; Gorina, S.S.; Gogoleva, N.E.; Toporkova, Y.Y.; Chechetkin, I.R.; Grechkin, A.N. Green leaf divinyl ether synthase: Gene detection, molecular cloning and identification of a unique CYP74B subfamily member. Biochim. Biophys. Acta 2012, 1821, 287–294. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | kcat (s−1) | Km (μM) | kcat/Km (μM−1∙s−1) | Catalytic Efficiency, % 15(S)-HPEPE |
|---|---|---|---|---|
| 15-HPEPE | 275 ± 16 | 8.17 ± 1.6 | 33.7 | 100 |
| 9-HPOT | 254.2 ± 26 | 20.9 ± 5.6 | 12.2 | 36.2 |
| 9-HPOD | 551.3 ± 19 | 46.6 ± 7.2 | 11.8 | 35 |
| 13-HPOT | 189 ± 18 | 28.4 ± 6.7 | 6.7 | 19.9 |
| 13-HPOD | 291.8 ± 28 | 57.9 ± 13.3 | 5.1 | 15.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gorina, S.S.; Iljina, T.M.; Mukhtarova, L.S.; Toporkova, Y.Y.; Grechkin, A.N. Detection of Unprecedented CYP74 Enzyme in Mammal: Hydroperoxide Lyase CYP74C44 of the Bat Sturnira hondurensis. Int. J. Mol. Sci. 2022, 23, 8009. https://doi.org/10.3390/ijms23148009
Gorina SS, Iljina TM, Mukhtarova LS, Toporkova YY, Grechkin AN. Detection of Unprecedented CYP74 Enzyme in Mammal: Hydroperoxide Lyase CYP74C44 of the Bat Sturnira hondurensis. International Journal of Molecular Sciences. 2022; 23(14):8009. https://doi.org/10.3390/ijms23148009
Chicago/Turabian StyleGorina, Svetlana S., Tatiana M. Iljina, Lucia S. Mukhtarova, Yana Y. Toporkova, and Alexander N. Grechkin. 2022. "Detection of Unprecedented CYP74 Enzyme in Mammal: Hydroperoxide Lyase CYP74C44 of the Bat Sturnira hondurensis" International Journal of Molecular Sciences 23, no. 14: 8009. https://doi.org/10.3390/ijms23148009
APA StyleGorina, S. S., Iljina, T. M., Mukhtarova, L. S., Toporkova, Y. Y., & Grechkin, A. N. (2022). Detection of Unprecedented CYP74 Enzyme in Mammal: Hydroperoxide Lyase CYP74C44 of the Bat Sturnira hondurensis. International Journal of Molecular Sciences, 23(14), 8009. https://doi.org/10.3390/ijms23148009