
Enhanced Muscle Fibers of Epinephelus coioides by Myostatin Autologous Nucleic Acid Vaccine

 ,
,
Abstract
:1. Introduction
2. Results
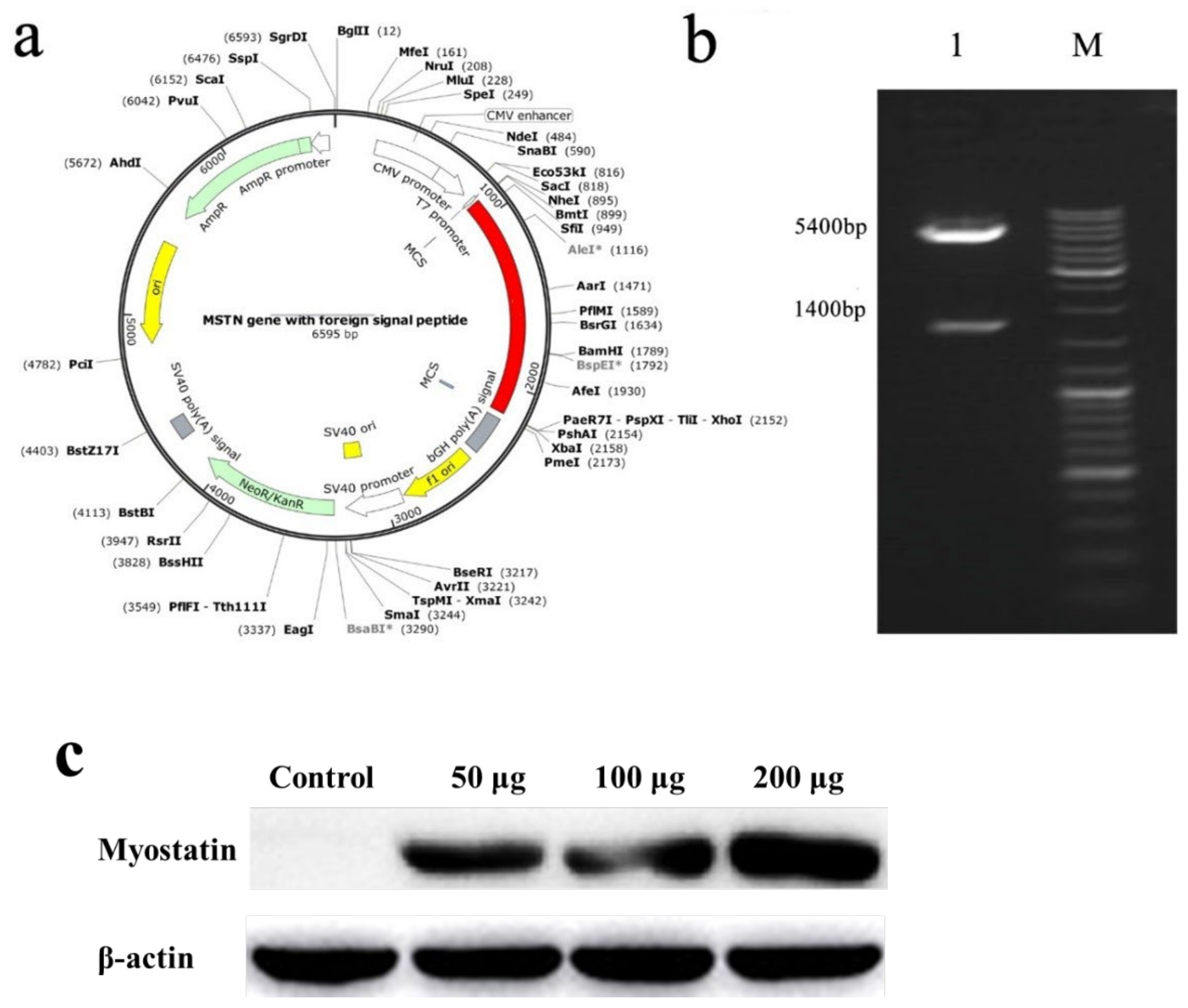
2.1. Construction of Nucleic Acid Vaccine
2.2. Effects of Myostatin Autologous Nucleic Acid Vaccine on Body Weight and Length
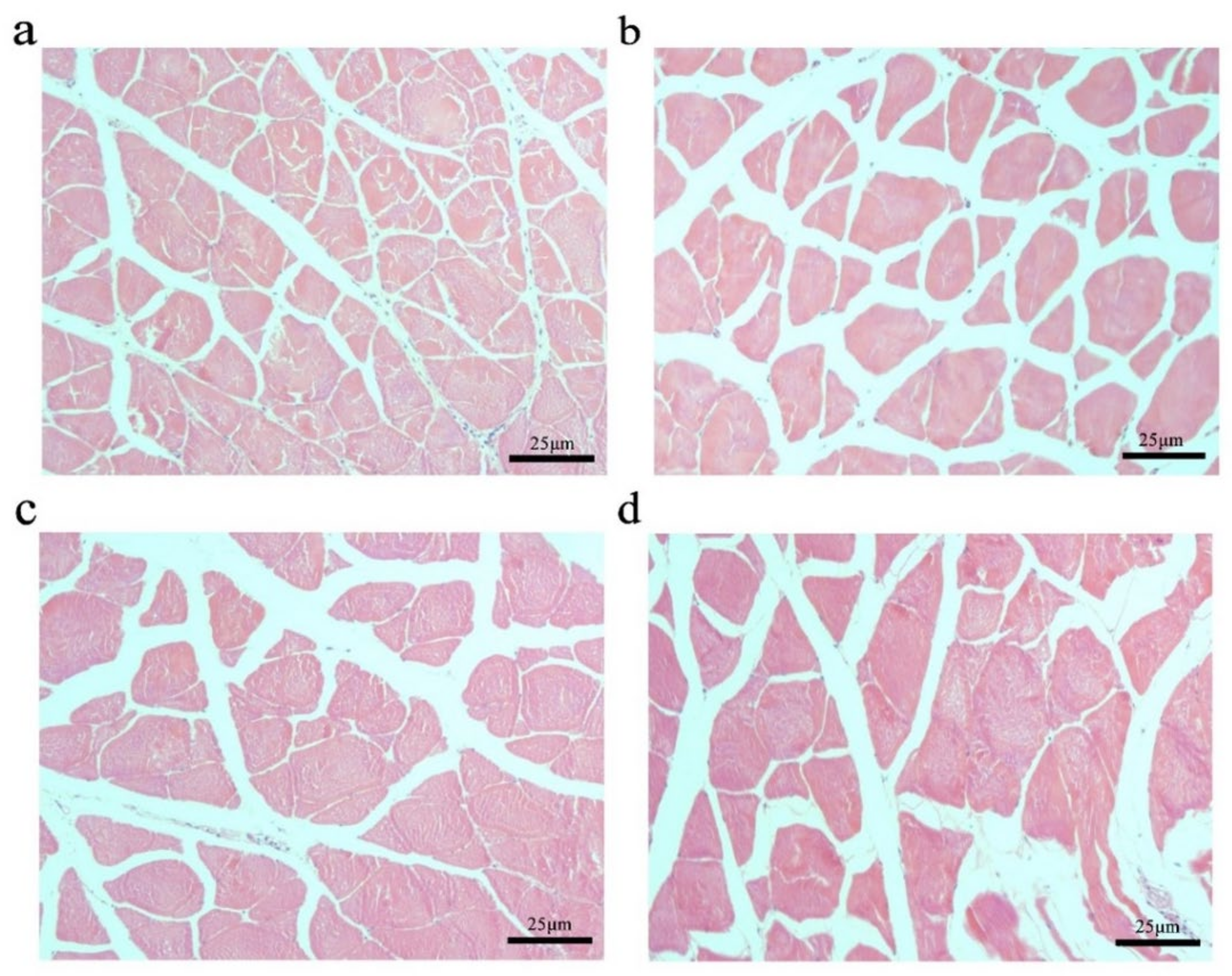
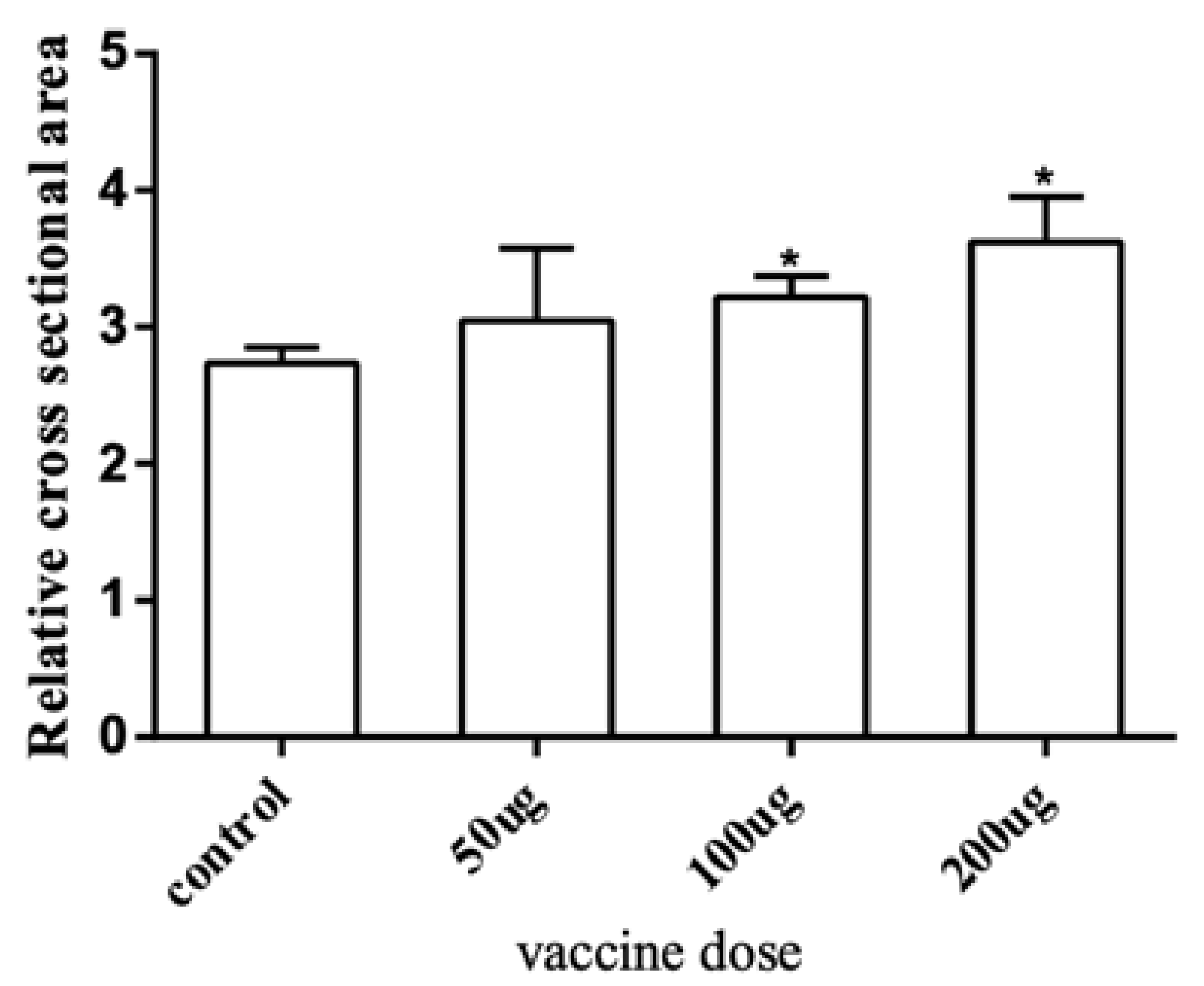
2.3. Effects of Myostatin Autologous Nucleic Acid Vaccine on the Muscle Tissue
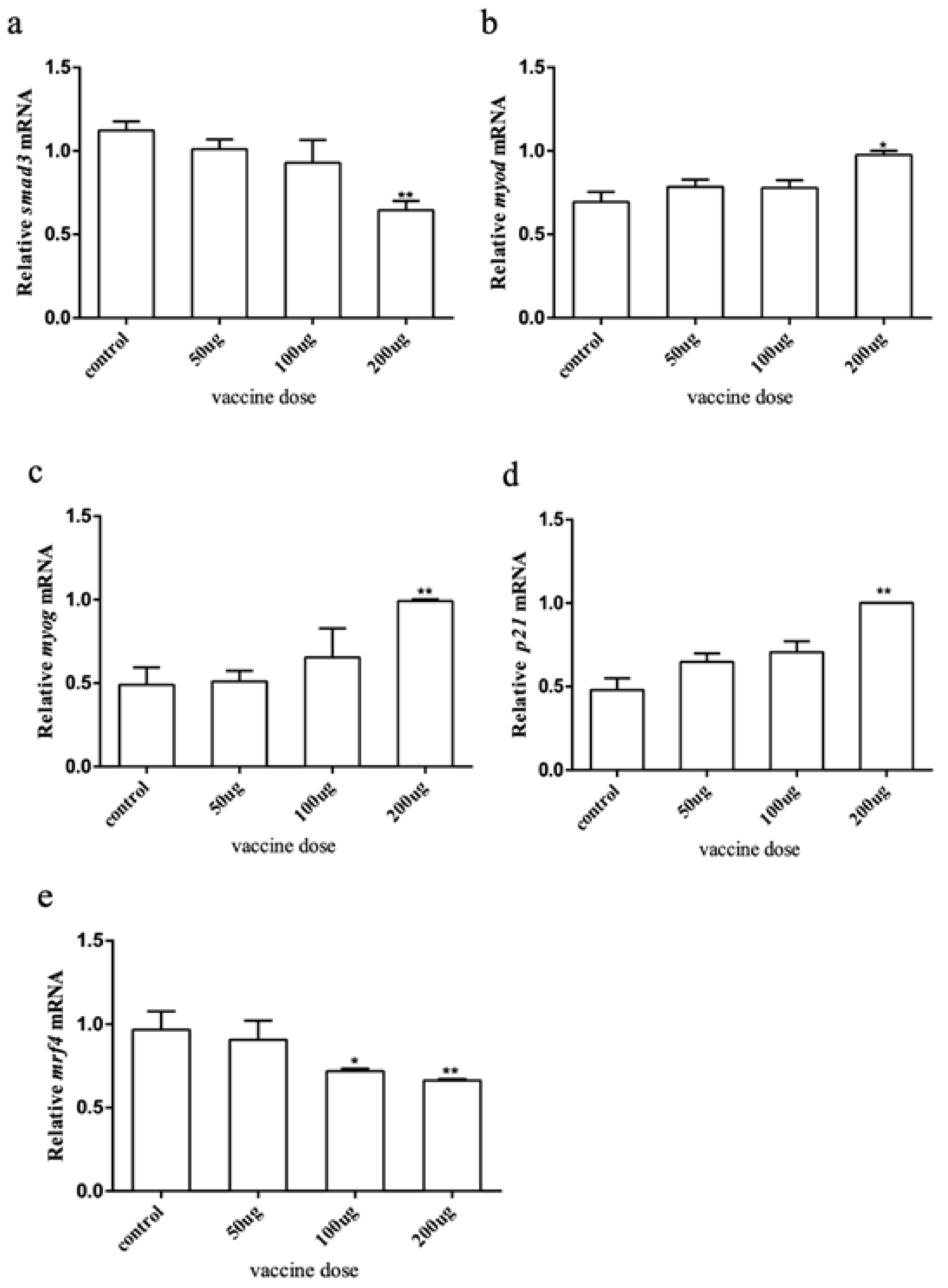
2.4. Effects of Myostatin Autologous Nucleic Acid Vaccine on the Downstream and Related Genes of the Myostatin Signaling Pathway
3. Discussion
4. Materials and Methods
4.1. Construction of Nucleic Acid Vaccine
4.2. Vaccine Production
4.3. Animals
4.4. Immunization Protocol and Sample Collection
4.5. Muscle Histology
4.6. RNA Extraction, cDNA Synthesis and qRT-PCR
4.7. Western Blot Analysis
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Saunders, M.A.; Good, J.M.; Lawrence, E.C.; Ferrell, R.E.; Li, W.H.; Nachman, M.W. Human adaptive evolution at Myostatin (GDF-8), a regulator of muscle growth. Am. J. Hum. Genet. 2006, 79, 1089–1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manickam, R.; Pena, R.N.; Whitelaw, C.B. Mammary gland differentiation inversely correlates with GDF-8 expression. Mol. Reprod. Dev. 2008, 75, 1783–1788. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.L.; Huang, Y.Y.; Ho, W.K.; Poon, H.K.; Cheung, P.L.; Wai, S.O.; Chow, P.H. Growth-differentiation factor-8 (GDF-8) in the uterus: Its identification and functional significance in the golden hamster. Reprod. Biol. Endocrinol. RBE 2009, 7, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peiris, H.N.; Mitchell, M.D. The expression and potential functions of placental myostatin. Placenta 2012, 33, 902–907. [Google Scholar] [CrossRef]
- Buehring, B.; Binkley, N. Myostatin—The holy grail for muscle, bone, and fat? Curr. Osteoporos. Rep. 2013, 11, 407–414. [Google Scholar] [CrossRef]
- Yasaka, N.; Suzuki, K.; Kishioka, Y.; Wakamatsu, J.; Nishimura, T. Laminin binds to myostatin and attenuates its signaling. Anim. Sci. J. Nihon Chikusan Gakkaiho 2013, 84, 663–668. [Google Scholar] [CrossRef]
- McPherron, A.C.; Lawler, A.M.; Lee, S.J. Regulation of skeletal muscle mass in mice by a new TGF-beta superfamily member. Nature 1997, 387, 83–90. [Google Scholar] [CrossRef]
- McPherron, A.C.; Lee, S.J. Double muscling in cattle due to mutations in the myostatin gene. Proc. Natl. Acad. Sci. USA 1997, 23, 12457–12461. [Google Scholar] [CrossRef] [Green Version]
- Hua, L.S.; Zhu, X.Y.; Rong, Z.; Yun, L.A.; Yan, D.C.; Wen, J.S.; Gang, L.M.; Qin, W.Y.; Chun, C.G. Polymorphism of Porcine Myostatin Gene. Acta Genet. Sin. 2002, 29, 326–331. [Google Scholar]
- Tay, G.K.; Iaschi, S.P.A.; Bellinge, R.H.S.; Chong, F.N.; Hui, J. The development of sequence-based-typing of myostatin (GDF-8) to identify the double muscling phenotype in the goat. Small Rumin. Res. 2004, 52, 1–12. [Google Scholar] [CrossRef]
- Gu, H.; Cao, Y.; Qiu, B.; Zhou, Z.; Deng, R.; Chen, Z.; Li, R.; Li, X.; Wei, Q.; Xia, X.; et al. Establishment and phenotypic analysis of an Mstn knockout rat. Biochem. Biophys. Res. Commun. 2016, 477, 115–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostbye, T.K.; Galloway, T.F.; Nielsen, C.; Gabestad, I.; Bardal, T.; Andersen, O. The two myostatin genes of Atlantic salmon (Salmo salar) are expressed in a variety of tissues. Eur. J. Biochem. 2001, 268, 5249–5257. [Google Scholar] [CrossRef] [PubMed]
- Rescan, P.Y.; Jutel, I.; Ralliere, C. Two myostatin genes are differentially expressed in myotomal muscles of the trout (Oncorhynchus mykiss). J. Exp. Biol. 2001, 204 Pt 20, 3523–3529. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.Q.; Chen, S.L.; Sha, Z.X.; Liu, Y. Molecular cloning and expression analysis of the myostatin gene in sea perch (Lateolabrax japonicus). Mar. Biotechnol. 2007, 9, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Y.; Sun, Y.; Zhang, X.; Wan, H.; Yao, C.; Liang, K.; Li, L.; Liu, B.; Zhong, J.; Zhang, Z.; et al. Characterization of two myostatin genes in pufferfish Takifugu bimaculatus: Sequence, genomic structure, and expression. PeerJ 2020, 8, e9655. [Google Scholar] [CrossRef]
- Wu, L.; Li, Y.; Xu, Y.; Wang, L.; Ma, X.; Dong, C.; Zhao, X.; Tian, X.; Li, X.; Kong, X. The roles of two myostatins and immune effects after inhibition in Qi river crucian carp (Carassius auratus). Fish Shellfish Immunol. 2020, 98, 710–719. [Google Scholar] [CrossRef]
- Gabillard, J.C.; Biga, P.R.; Rescan, P.Y.; Seiliez, I. Revisiting the paradigm of myostatin in vertebrates: Insights from fishes. Gen. Comp. Endocrinol. 2013, 194, 45–54. [Google Scholar] [CrossRef]
- Gao, Y.; Dai, Z.; Shi, C.; Zhai, G.; Jin, X.; He, J.; Lou, Q.; Yin, Z. Depletion of Myostatin b Promotes Somatic Growth and Lipid Metabolism in Zebrafish. Front. Endocrinol. 2016, 7, 88. [Google Scholar] [CrossRef] [Green Version]
- Stinckens, A.; Georges, M.; Buys, N. Mutations in the myostatin gene leading to hypermuscularity in mammals: Indications for a similar mechanism in fish? Anim. Genet. 2011, 42, 229–234. [Google Scholar] [CrossRef]
- Helterline, D.L.; Garikipati, D.; Stenkamp, D.L.; Rodgers, B.D. Embryonic and tissue-specific regulation of myostatin-1 and -2 gene expression in zebrafish. Gen. Comp. Endocrinol. 2007, 151, 90–97. [Google Scholar] [CrossRef] [Green Version]
- Grobet, L.; Martin, L.J.; Poncelet, D.; Pirottin, D.; Brouwers, B.; Riquet, J.; Schoeberlein, A.; Dunner, S.; Menissier, F.; Massabanda, J.; et al. A deletion in the bovine myostatin gene causes the double-muscled phenotype in cattle. Nat. Genet. 1997, 17, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Kambadur, R.; Sharma, M.; Smith, T.P.; Bass, J.J. Mutations in myostatin (GDF8) in double-muscled Belgian Blue and Piedmontese cattle. Genome Res. 1997, 7, 910–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosher, D.S.; Quignon, P.; Bustamante, C.D.; Sutter, N.B.; Mellersh, C.S.; Parker, H.G.; Ostrander, E.A. A mutation in the myostatin gene increases muscle mass and enhances racing performance in heterozygote dogs. PLoS Genet. 2007, 3, e79. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Ouyang, H.; Xie, Z.; Yao, C.; Guo, N.; Li, M.; Jiao, H.; Pang, D. Efficient Generation of Myostatin Mutations in Pigs Using the CRISPR/Cas9 System. Sci. Rep. 2015, 5, 16623. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.; Fujimura, T.; Matsunari, H.; Sakuma, T.; Nakano, K.; Watanabe, M.; Asano, Y.; Kitagawa, E.; Yamamoto, T.; Nagashima, H. Efficient modification of the myostatin gene in porcine somatic cells and generation of knockout piglets. Mol. Reprod. Dev. 2016, 83, 61–70. [Google Scholar] [CrossRef]
- He, Z.; Zhang, T.; Jiang, L.; Zhou, M.; Wu, D.; Mei, J.; Cheng, Y. Use of CRISPR/Cas9 technology efficiently targetted goat myostatin through zygotes microinjection resulting in double-muscled phenotype in goats. Biosci. Rep. 2018, 38, BSR20180742. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.Y.; Hu, S.Y.; Gong, H.Y.; Chen, M.H.; Lu, J.K.; Wu, J.L. Suppression of myostatin with vector-based RNA interference causes a double-muscle effect in transgenic zebrafish. Biochem. Biophys. Res. Commun. 2009, 387, 766–771. [Google Scholar] [CrossRef]
- Whittemore, L.-A.; Song, K.; Li, X.; Aghajanian, J.; Davies, M.; Girgenrath, S.; Hill, J.J.; Jalenak, M.; Kelley, P.; Knight, A.; et al. Inhibition of myostatin in adult mice increases skeletal muscle mass and strength. Biochem. Biophys. Res. Commun. 2003, 300, 965–971. [Google Scholar] [CrossRef]
- Tang, L.; Yan, Z.; Wan, Y.; Han, W.; Zhang, Y. Myostatin DNA vaccine increases skeletal muscle mass and endurance in mice. Muscle Nerve 2007, 36, 342–348. [Google Scholar] [CrossRef]
- Nelson, C.E.; Gersbach, C.A. Engineering Delivery Vehicles for Genome Editing. Annu. Rev. Chem. Biomol. Eng. 2016, 7, 637–662. [Google Scholar] [CrossRef] [Green Version]
- Chiang, Y.A.; Kinoshita, M.; Maekawa, S.; Kulkarni, A.; Lo, C.F.; Yoshiura, Y.; Wang, H.C.; Aoki, T. TALENs-mediated gene disruption of myostatin produces a larger phenotype of medaka with an apparently compromised immune system. Fish Shellfish Immunol. 2016, 48, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Mao, D.G.; Yang, L.G.; Cao, S.X.; Zhang, Z.L.; He, X.H. Construction of expression vector of follicular inhibin and HBsAg fusion gene and expression in Hela cell. Chin. J. Immunol. 2003, 19, 775–778. [Google Scholar]
- Cao, S.X.; Yang, L.G.; Zhang, W.W.; Mao, D.G.; He, X.H. Construction and Expression of Somatostatin DNA Vaccine pEGS/2SS in Hela. Chin. J. Vet. Sci. 2005, 025, 499–502. [Google Scholar]
- Comai, G.; Tajbakhsh, S. Molecular and cellular regulation of skeletal myogenesis. Curr. Top. Dev. Biol. 2014, 110, 1–73. [Google Scholar] [PubMed]
- Buckingham, M.; Rigby, P.W. Gene regulatory networks and transcriptional mechanisms that control myogenesis. Dev. Cell 2014, 28, 225–238. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Hernandez, J.M.; Garcia-Gonzalez, E.G.; Brun, C.E.; Rudnicki, M.A. The myogenic regulatory factors, determinants of muscle development, cell identity and regeneration. Semin. Cell Dev. Biol. 2017, 72, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Acosta, J.; Carpio, Y.; Borroto, I.; González, O.; Estrada, M.P. Myostatin gene silenced by RNAi show a zebrafish giant phenotype. J. Biotechnol. 2005, 119, 324–331. [Google Scholar] [CrossRef]
- Wang, X.; Niu, Y.; Zhou, J.; Zhu, H.; Ma, B.; Yu, H.; Yan, H.; Hua, J.; Huang, X.; Qu, L.; et al. CRISPR/Cas9-mediated MSTN disruption and heritable mutagenesis in goats causes increased body mass. Anim. Genet. 2018, 49, 43–51. [Google Scholar] [CrossRef]
- Ji, S.; Losinski, R.L.; Cornelius, S.G.; Frank, G.R.; Willis, G.M.; Gerrard, D.E.; Depreux, F.F.; Spurlock, M.E. Myostatin expression in porcine tissues: Tissue specificity and developmental and postnatal regulation. Am. J. Physiol. 1998, 275 Pt 2, R1265–R1273. [Google Scholar] [CrossRef]
- Lyons, J.A.; Haring, J.S.; Biga, P.R. Myostatin expression, lymphocyte population, and potential cytokine production correlate with predisposition to high-fat diet induced obesity in mice. PLoS ONE 2010, 5, e12928. [Google Scholar] [CrossRef]
- Liu, H.; Mao, H.; Dong, X.; Cao, H.; Liu, K.; Yin, Z. Expression of MSTN gene and its correlation with pectoralis muscle fiber traits in the domestic pigeons (Columba livia). Poult. Sci. 2019, 98, 5265–5271. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; McPherron, A.C. Regulation of myostatin activity and muscle growth. Proc. Natl. Acad. Sci. USA 2001, 98, 9306–9311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Black, B.L.; Derynck, R. TGF-beta inhibits muscle differentiation through functional repression of myogenic transcription factors by Smad3. Genes Dev. 2001, 15, 2950–2966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rebbapragada, A.; Benchabane, H.; Wrana, J.L.; Celeste, A.J.; Attisano, L. Myostatin signals through a transforming growth factor beta-like signaling pathway to block adipogenesis. Mol. Cell. Biol. 2003, 23, 7230–7242. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Topouzis, S.; Liang, L.F.; Stotish, R.L. Myostatin signaling through Smad2, Smad3 and Smad4 is regulated by the inhibitory Smad7 by a negative feedback mechanism. Cytokine 2004, 26, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; McFarlane, C.; Vajjala, A.; Lokireddy, S.; Ng, Z.H.; Tan, C.K.; Tan, N.S.; Wahli, W.; Sharma, M.; Kambadur, R. Smad3 signaling is required for satellite cell function and myogenic differentiation of myoblasts. Cell Res. 2011, 21, 1591–1604. [Google Scholar] [CrossRef] [Green Version]
- Du, W. Effect of Immunization with Myostatin DNA Vaccine and Propeptide on Animal Muscle Growth. Ph.D. Thesis, Shihezi University, Shihezi, China, 2018. [Google Scholar]
- Wang, H.N.; Sun, H.X.; Zhang, Y.J.; Liu, Y.Q.; Gu, Z.H.; Shi, X.F. Effects of Interfering MSTN on Proliferation and Differentiation of Sheep Myoblasts and Expression of Related Genes. Chin. J. Anim. Vet. Sci. 2018, 49, 46–54. [Google Scholar]
- Xia, J. Study on Tissues Distribution and Preparation Technology of Avian Influenza DNA Vaccine. Master’s Thesis, Xinjiang Agricultural University, Urumqi, China, 2007. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′–3′) |
|---|---|
| KOZAK | GCCACC |
| signal peptide of Japanese encephalitis virus | ATGGGAAAGAGAAGCGCCGGCAGCATCATGTGGCTGGCCAGCCTGGCCGTCGTGATCGCCTGTGCCGGCGCC |
| Surface core antigen of hepatitis B virus | ATGGAGAACATCGCATCTGGACTCCTTGGACCCCTGCTCGTGCTGCAGGCCGGGTTTTTCCTGCTGACAAGAATCCTCACAATACCACAGAGTCTCGACTCCTGGTGGACTTCTCTCAATTTTCTCGGGGGAACACCCGTGTGCCTTGGCCAAAACTCCCAGTCCCAAACCTCCAGTCATAGCCCAACCTGCTGTCCTCCAATTTGTCCTGGTTATCGCTGGATGTGTCTGCGGAGGTTTATCATCTTTCTCTGCATCCTGCTGCTCTGCCTCATCTTCCTGCTGGTTCTTCTGGACTATCAGGGTATGCTGCCCGTTTGTCCTCTCATTCCAGGAAGCAGCACGACCAGCACCGGCCCATGCAAGACCTGCACCAGTCCTGCTCAGGGCACCTCTATGTTTCCCAGCTGCTGTTGTACTAAACCTACGGATGGCAACTGCACCTGCATTCCCATCCCAAGCTCTTGGGCTTTCGCCAAGTACCTCTGGGAATGGGCCAGCGTCCGATTCTCTTGGCTCAGTCTGCTCGTGCCTTTTGTGCAGTCCTTCGTGGGTCTTTCCCCCACTGTCTGGCTTAGTGTGATATGGATGATGTGGTACTGGGGCCCTAGTCTGTACAACATCCTGAGTCCCTTCATACCGCTGCTGCCGATATTCTTTTGCCTTTGGGTCTAC |
| Th epitope | CAGTATATAAAAGCAAATTCTAAATTTATAGGTATAACTGAA |
| somatostatin | GCCGGCTGTAAGAACTTCTTTTGGAAAACATTCACCAGCTGC |
| linker | GGAGGCGGTGGATCCGGAGGCGGAGGAAGCGGAGGTGGCGGCTCT |
| myostatin mature peptide gene | TCAGGCCTGGACTGTGACGAGAACTCTCCAGAGTCCCGGTGCTGCCGCTATCCGCTCACAGTGGACTTTGAAGACTTTGGCTGGGACTGGATTATTGCCCCAAAGCGCTACAAGGCCAACTATTGCTCCGGGGAGTGTGAGTACATGCACTTGCAGAAGTACCCACACACCCACCTGGTGAACAAAGCCAATCCCAGAGGGACCGCAGGCCCCTGCTGCACCCCCACCAAGATGTCACCCATCAACATGCTCTACTTTAACCGAAAAGAGCAGATCATCTATGGCAAGATCCCCTCCATGGTGGTGGACCGTTGTGGATGCTCTTGA |
| Primer | Sequence (5′-3′) |
|---|---|
| myod-RT-F | CGGCGGCTTGGTAA |
| myod-RT-R | CATCGGAGCAGTTGGA |
| p21-RT-F | GGTGGTAGAAAGAAAGAT |
| p21-RT-R | TTGCCTGTAGAGTCGTA |
| β-actin-RT-F | TCTTCCAGCCATCCTTCCTTGG |
| β-actin-RT-R | CTGCATACGGTCAGCAATGCC |
| myog-RT-F | GATGGGCTTATGTGGG |
| myog-RT-R | GGTAACCGTCTTCCTTTT |
| mrf4-RT-F | TACAACGGCAACGACA |
| mrf4-RT-R | GCCCACATGAGGCACT |
| smad3-RT-F | GCATAACCATACCCAGAT |
| smad3-RT-R | CGCAGACTTCGTCCTT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, B.; Yang, J.; Yang, Y.; Xia, J.; He, Y.; Wang, Q.; Zhao, H.; Yang, H. Enhanced Muscle Fibers of Epinephelus coioides by Myostatin Autologous Nucleic Acid Vaccine. Int. J. Mol. Sci. 2022, 23, 6997. https://doi.org/10.3390/ijms23136997
Fu B, Yang J, Yang Y, Xia J, He Y, Wang Q, Zhao H, Yang H. Enhanced Muscle Fibers of Epinephelus coioides by Myostatin Autologous Nucleic Acid Vaccine. International Journal of Molecular Sciences. 2022; 23(13):6997. https://doi.org/10.3390/ijms23136997
Chicago/Turabian StyleFu, Bing, Jinzeng Yang, Yan Yang, Jun Xia, Yinglin He, Qing Wang, Huihong Zhao, and Huirong Yang. 2022. "Enhanced Muscle Fibers of Epinephelus coioides by Myostatin Autologous Nucleic Acid Vaccine" International Journal of Molecular Sciences 23, no. 13: 6997. https://doi.org/10.3390/ijms23136997
APA StyleFu, B., Yang, J., Yang, Y., Xia, J., He, Y., Wang, Q., Zhao, H., & Yang, H. (2022). Enhanced Muscle Fibers of Epinephelus coioides by Myostatin Autologous Nucleic Acid Vaccine. International Journal of Molecular Sciences, 23(13), 6997. https://doi.org/10.3390/ijms23136997