
Hypercholesterolemia Negatively Regulates P2X7-Induced Cellular Function in CD4+ and CD8+ T-Cell Subsets from B6 Mice Fed a High-Fat Diet


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
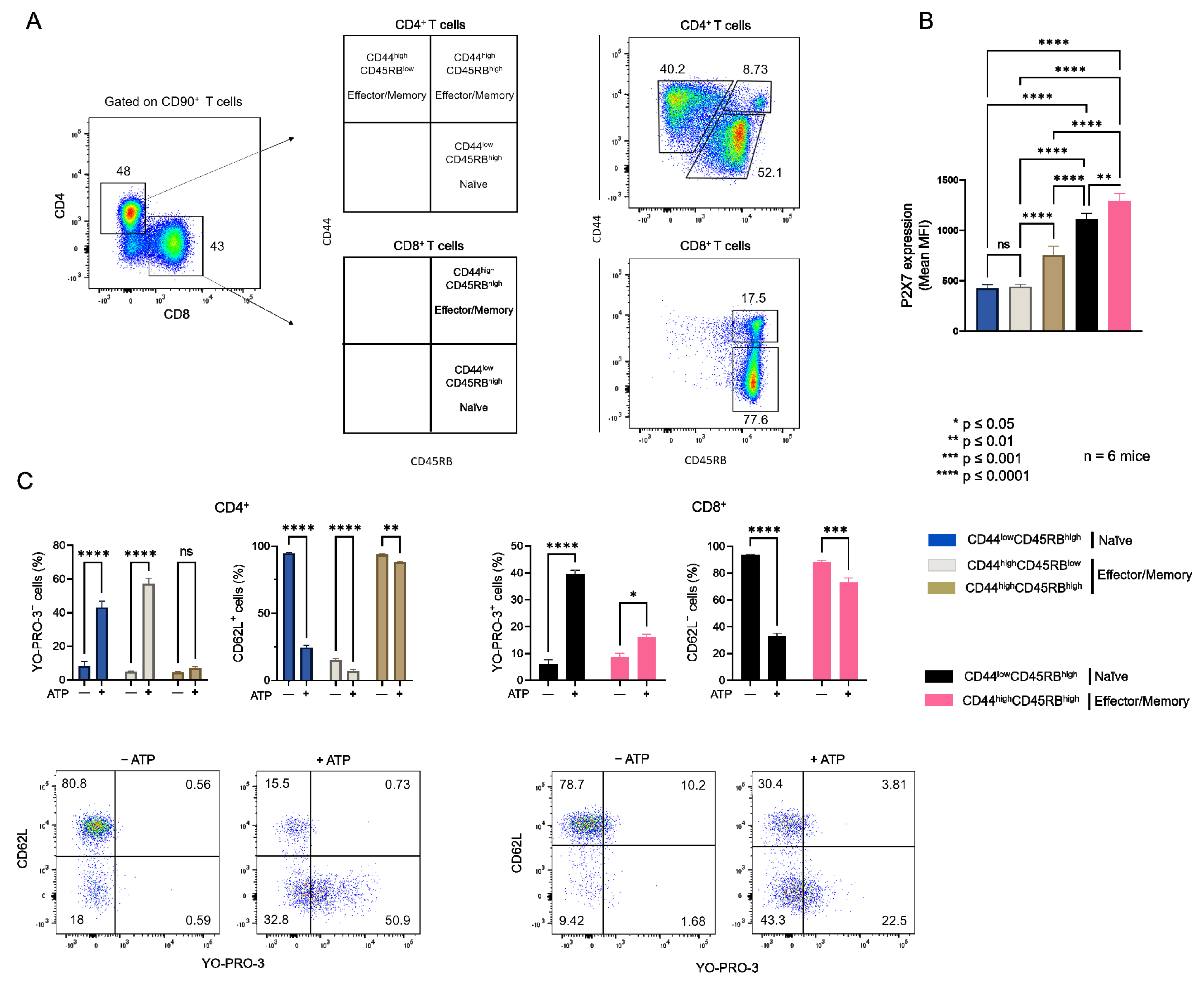
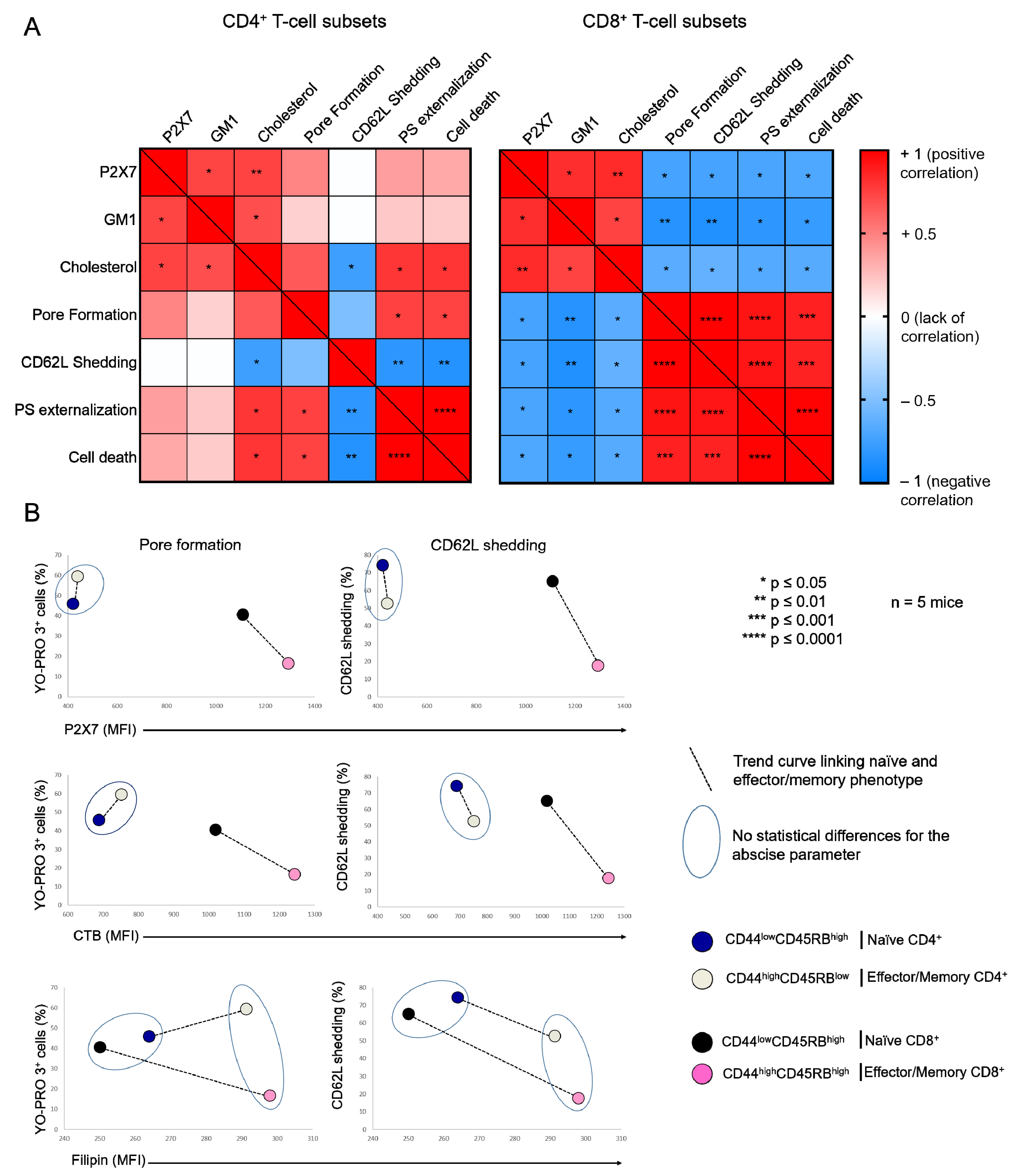
2.1. Plasma Membrane Levels of Cholesterol and GM1 Gangliosides in T Cells at Different Stages of Differentiation and the Consequences on Their Sensitivity to eATP
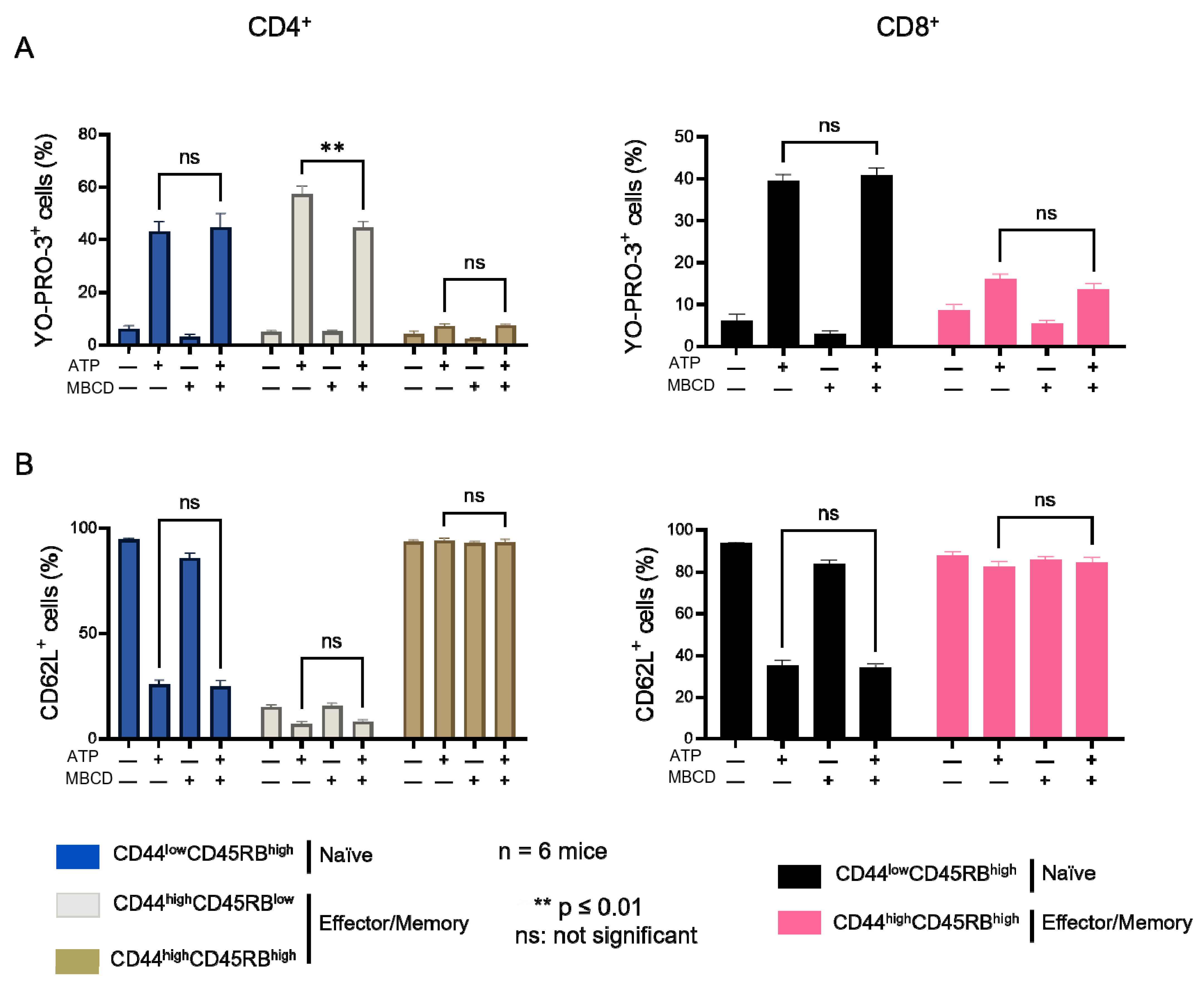
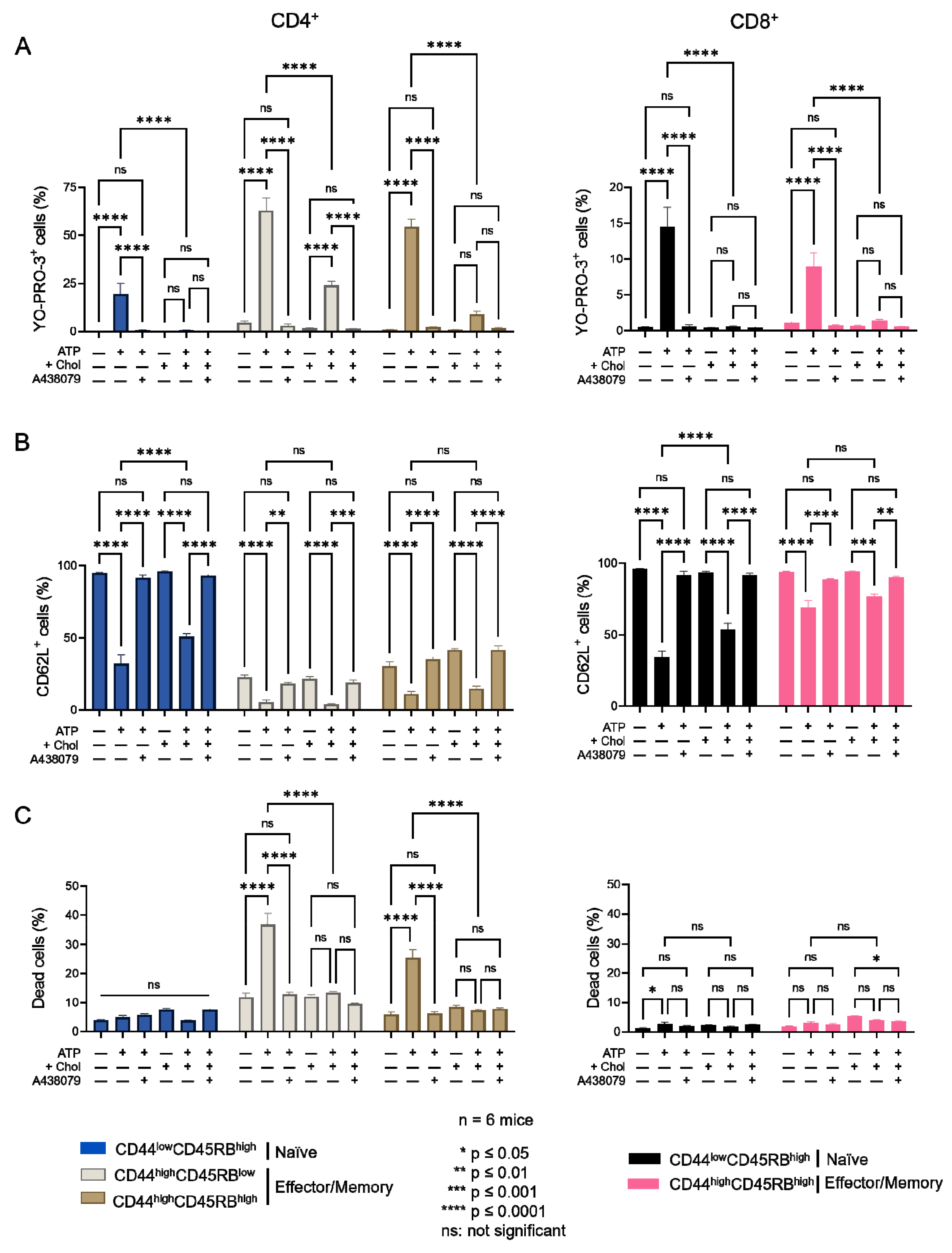
2.2. Amplitude of P2X7-Induced Cellular Responses in T Cells Following Plasma Membrane Cholesterol Modulation
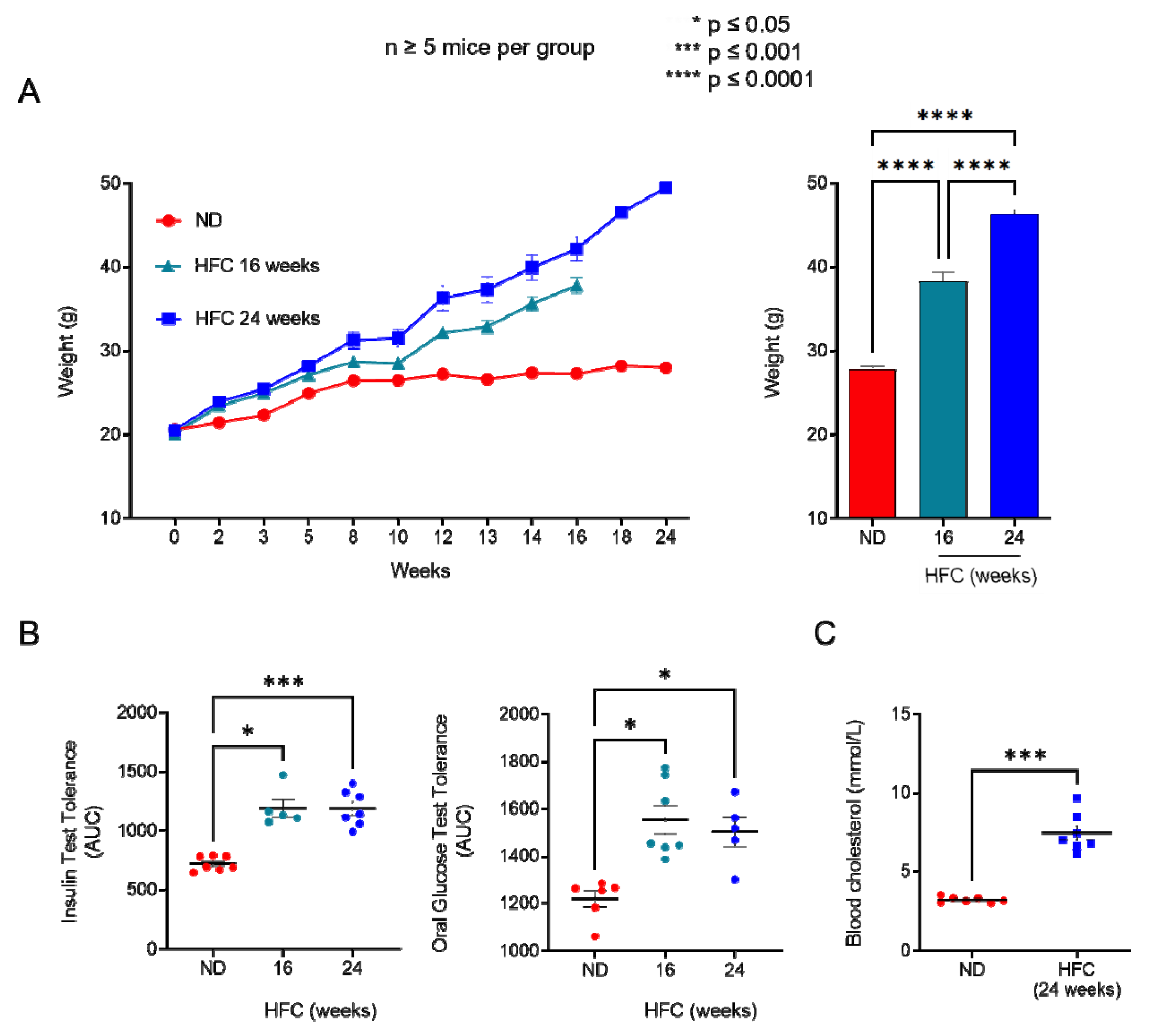
2.3. Diet-Induced Hypercholesterolemia Increase Effector/Memory CD4+ and CD8+ T Cell Numbers in B6 Mice
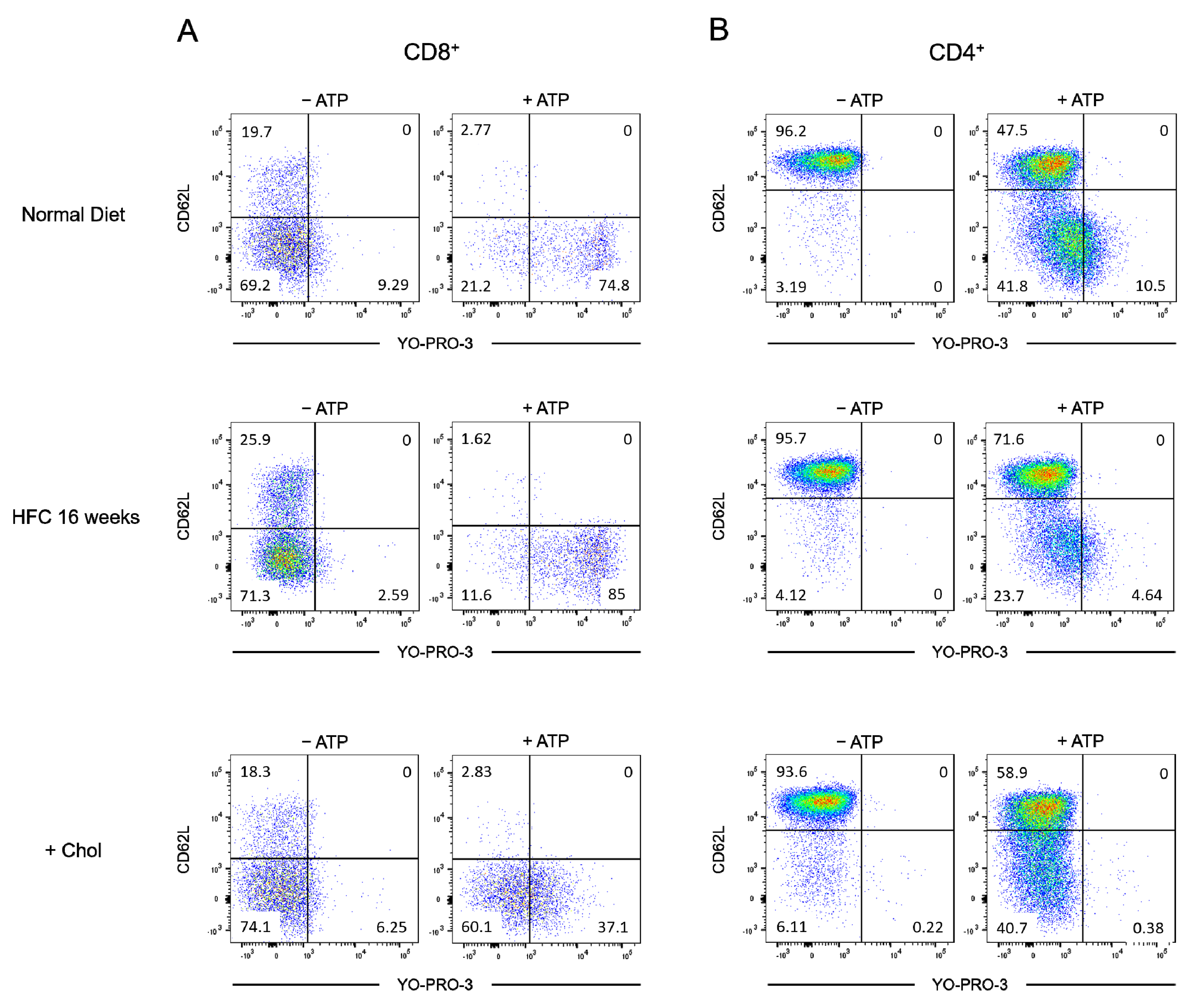
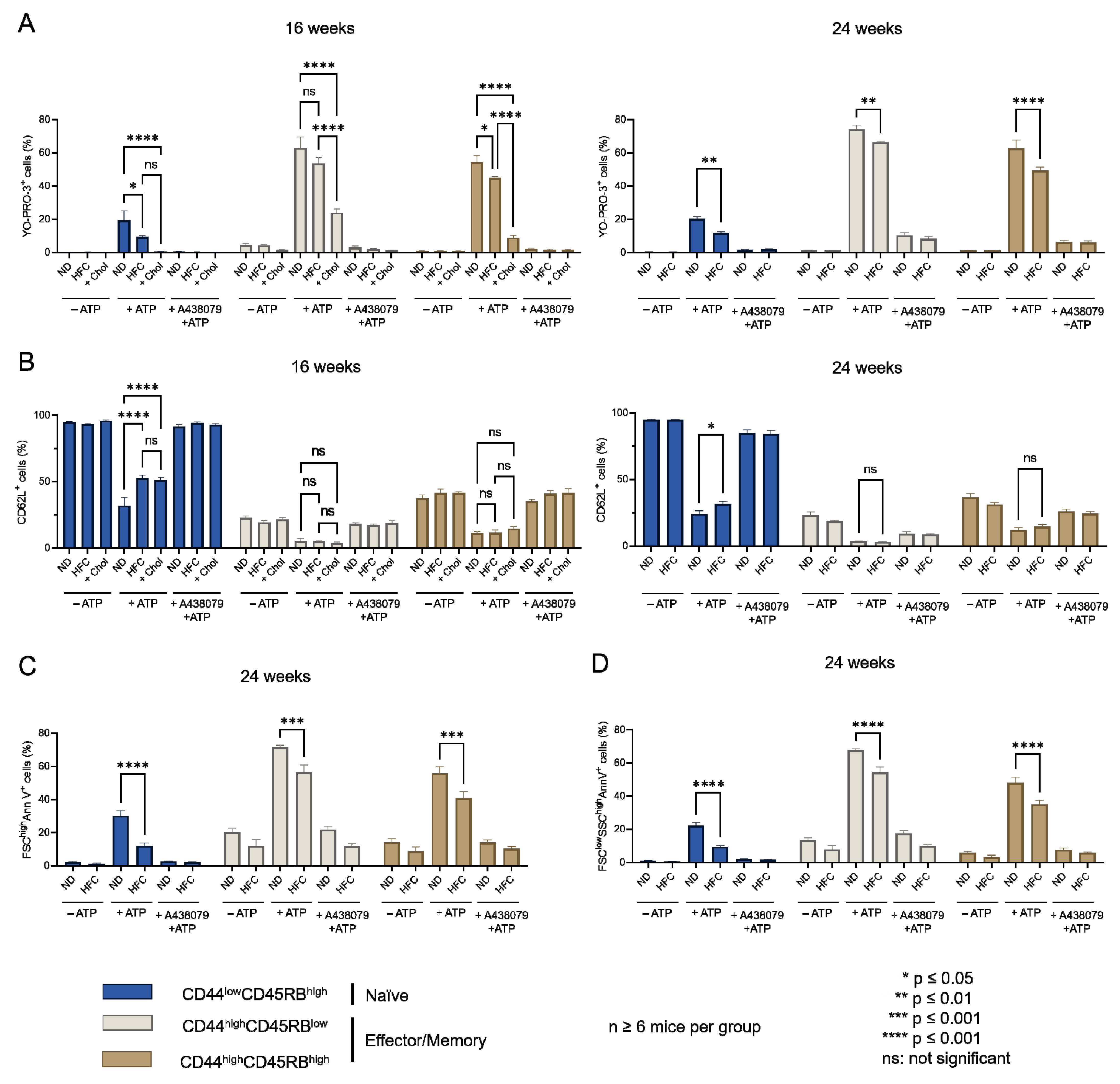
2.4. Diet-Induced Hypercholesterolemia Affected P2X7-Induced Cellular Responses in Naive and Effector/Memory CD4+ and CD8+ T Cells
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Mice
4.3. Diet
4.4. Metabolic Assay
4.5. Serological Parameters
4.6. Plasma Membrane Cholesterol Modulation
4.7. T Lymphocyte Phenotyping
4.8. ATP-Mediated Cellular Function Assays
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
References
- Sluyter, R. The P2X7 Receptor. Adv. Exp. Med. Biol. 2017, 1051, 17–53. [Google Scholar] [PubMed]
- Di Virgilio, F.; Dal Ben, D.; Sarti, A.C.; Giuliani, A.L.; Falzoni, S. The P2X7 Receptor in Infection and Inflammation. Immunity 2017, 47, 15–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiley, J.S.; Chen, R.; Wiley, M.J.; Jamieson, G.P. The ATP4—Receptor-operated ion channel of human lymphocytes: Inhibition of ion fluxes by amiloride analogs and by extracellular sodium ions. Arch. Biochem. Biophys. 1992, 292, 411–418. [Google Scholar] [CrossRef]
- Mackenzie, A.B.; Young, M.T.; Adinolfi, E.; Surprenant, A. Pseudoapoptosis Induced by Brief Activation of ATP-gated P2X7 Receptors. J. Biol. Chem. 2005, 280, 33968–33976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinberg, T.H.; Newman, A.S.; Swanson, J.A.; Silverstein, S.C. ATP4—Permeabilizes the plasma membrane of mouse macrophages to fluorescent dyes. J. Biol. Chem. 1987, 262, 8884–8888. [Google Scholar] [CrossRef]
- Surprenant, A.; Rassendren, F.; Kawashima, E.; North, R.A.; Buell, G. The cytolytic P2Z receptor for extracellular ATP identified as a P2X receptor (P2X7). Science 1996, 272, 735–738. [Google Scholar] [CrossRef]
- Ferrari, D.; Los, M.; Bauer, M.K.; Vandenabeele, P.; Wesselborg, S.; Schulze-Osthoff, K. P2Z purinoreceptor ligation induces activation of caspases with distinct roles in apoptotic and necrotic alterations of cell death. FEBS Lett. 1999, 447, 71–75. [Google Scholar] [CrossRef] [Green Version]
- Humphreys, B.D.; Rice, J.; Kertesy, S.B.; Dubyak, G.R. Stress-activated Protein Kinase/JNK Activation and Apoptotic Induction by the Macrophage P2X7 Nucleotide Receptor. J. Biol. Chem. 2000, 275, 26792–26798. [Google Scholar] [CrossRef]
- Auger, R.; Motta, I.; Benihoud, K.; Ojcius, D.M.; Kanellopoulos, J.M. A role for mitogen-activated protein kinase(Erk1/2) activation and non-selective pore formation in P2X7 receptor-mediated thymocyte death. J. Biol. Chem. 2005, 280, 28142–28151. [Google Scholar] [CrossRef] [Green Version]
- Adinolfi, E.; Callegari, M.G.; Cirillo, M.; Pinton, P.; Giorgi, C.; Cavagna, D.; Di Virgilio, F. Expression of the P2X7 receptor increases the Ca2+ content of the endoplasmic reticulum, activates NFATc1, and protects from apoptosis. J. Biol. Chem. 2009, 284, 10120–10128. [Google Scholar] [CrossRef] [Green Version]
- Adinolfi, E.; Cirillo, M.; Woltersdorf, R.; Falzoni, S.; Chiozzi, P.; Pellegatti, P.; Callegari, M.G.; Sandonà, D.; Markwardt, F.; Schmalzing, G.; et al. Trophic activity of a naturally occurring truncated isoform of the P2X7 receptor. FASEB J. 2010, 24, 3393–3404. [Google Scholar] [CrossRef] [PubMed]
- Baricordi, O.R.; Ferrari, D.; Melchiorri, L.; Chiozzi, P.; Hanau, S.; Chiari, E.; Rubini, M.; Di Virgilio, F. An ATP-activated channel is involved in mitogenic stimulation of human T lymphocytes. Blood 1996, 87, 682–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giuliani, A.L.; Sarti, A.C.; Falzoni, S.; Di Virgilio, F. The P2X7 Receptor-Interleukin-1 Liaison. Front. Pharmacol. 2017, 8, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Gall, S.M.; Bobé, P.; Reiss, K.; Horiuchi, K.; Niu, X.D.; Lundell, D.; Blobel, C.P. ADAMs 10 and 17 represent differentially regulated components of a general shedding machinery for membrane proteins such as transforming growth factor alpha, L-selectin, and tumor necrosis factor alpha. Mol. Biol. Cell 2009, 20, 1785–1794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopp, R.; Krautloher, A.; Ramírez-Fernández, A.; Nicke, A. P2X7 Interactions and Signaling—Making Head or Tail of It. Front. Mol. Neurosci. 2019, 12, 183. [Google Scholar] [CrossRef]
- Safya, H.; Mellouk, A.; Legrand, J.; Le Gall, S.M.; Benbijja, M.; Kanellopoulos-Langevin, C.; Kanellopoulos, J.M.; Bobé, P. Variations in Cellular Responses of Mouse T Cells to Adenosine-5′-Triphosphate Stimulation Do Not Depend on P2X7 Receptor Expression Levels but on Their Activation and Differentiation Stage. Front. Immunol. 2018, 9, 360. [Google Scholar] [CrossRef] [Green Version]
- Mellouk, A.; Bobé, P. CD8(+), but not CD4(+) effector/memory T cells, express the CD44 (high) CD45RB (high) phenotype with aging, which displays reduced expression levels of P2X7 receptor and ATP-induced cellular responses. FASEB J. 2019, 33, 3225–3236. [Google Scholar] [CrossRef]
- Le Gall, S.M.; Legrand, J.; Benbijja, M.; Safya, H.; Benihoud, K.; Kanellopoulos, J.M.; Bobé, P. Loss of P2X7 Receptor Plasma Membrane Expression and Function in Pathogenic B220+ Double-Negative T Lymphocytes of Autoimmune MRL/lpr Mice. PLoS ONE 2012, 7, e52161. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Marcos, M.; Peérez-Andreés, E.; Tandel, S.; Fontanils, U.; Kumps, A.; Kabreé, E.; Goémez-Muñoz, A.; Marino, A.; Dehaye, J.-P.; Pochet, S. Coupling of two pools of P2X7 receptors to distinct intracellular signaling pathways in rat submandibular gland. J. Lipid Res. 2006, 47, 705–714. [Google Scholar] [CrossRef] [Green Version]
- Gonnord, P.; Delarasse, C.; Auger, R.; Benihoud, K.; Prigent, M.; Cuif, M.H.; Lamaze, C.; Kanellopoulos, J.M. Palmitoylation of the P2X7 receptor, an ATP-gated channel, controls its expression and association with lipid rafts. FASEB J. 2008, 23, 795–805. [Google Scholar] [CrossRef]
- Murrell-Lagnado, R.D. Regulation of P2X Purinergic Receptor Signaling by Cholesterol. Curr. Top. Membr. 2017, 80, 211–232. [Google Scholar] [PubMed]
- McCarthy, A.E.; Yoshioka, C.; Mansoor, S.E. Full-Length P2X7 Structures Reveal How Palmitoylation Prevents Channel Desensitization. Cell 2019, 179, 659–670. [Google Scholar] [CrossRef] [PubMed]
- Karasawa, A.; Michalski, K.; Mikhelzon, P.; Kawate, T. The P2X7 receptor forms a dye-permeable pore independent of its intracellular domain but dependent on membrane lipid composition. eLife 2017, 6, e31186. [Google Scholar] [CrossRef] [PubMed]
- Robinson, L.E.; Shridar, M.; Smith, P.; Murrell-Lagnado, R.D. Plasma Membrane Cholesterol as a Regulator of Human and Rodent P2X7 Receptor Activation and Sensitization. J. Biol. Chem. 2014, 289, 31983–31994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bietz, A.; Zhu, H.; Xue, M.; Xu, C. Cholesterol Metabolism in T Cells. Front. Immunol. 2017, 8, 1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zumerle, S.; Molon, B.; Viola, A. Membrane Rafts in T Cell Activation: A Spotlight on CD28 Costimulation. Front. Immunol. 2017, 8, 1467. [Google Scholar] [CrossRef]
- Mahammad, S.; Parmryd, I. Cholesterol depletion using methyl-beta-cyclodextrin. Methods Mol. Biol. 2015, 1232, 91–102. [Google Scholar]
- Hanada, K.; Nishijima, M.; Akamatsu, Y.; Pagano, R.E. Both sphingolipids and cholesterol participate in the detergent insolubility of alkaline phosphatase, a glycosylphosphatidylinositol-anchored protein, in mammalian membranes. J. Biol. Chem. 1995, 270, 6254–6260. [Google Scholar] [CrossRef] [Green Version]
- Hutteau-Hamel, T. P2X7 Receptor Activation by Extracellular ATP in Mouse T-Cell Lines Depends on its Plasma Membrane Localization Driven by Membrane Cholesterol Content. Ph.D. Thesis, Paris-Saclay University, Clamart, France, 2022. [Google Scholar]
- Sezgin, E.; Schwille, P. Fluorescence techniques to study lipid dynamics. Cold Spring Harb. Perspect. Biol. 2011, 3, a009803. [Google Scholar] [CrossRef] [Green Version]
- Brumeanu, T.-D.; Preda-Pais, A.; Stoica, C.; Bona, C.; Casares, S. Differential partitioning and trafficking of GM gangliosides and cholesterol-rich lipid rafts in thymic and splenic CD4 T cells. Mol. Immunol. 2007, 44, 530–540. [Google Scholar] [CrossRef]
- Thomas, S.; Kumar, R.; Casares, S.; Brumeanu, T.-D. Sensitive detection of GM1 lipid rafts and TCR partitioning in the T cell membrane. J. Immunol. Methods 2003, 275, 161–168. [Google Scholar] [CrossRef]
- Bonacina, F.; Coe, D.; Wang, G.; Longhi, M.P.; Baragetti, A.; Moregola, A.; Garlaschelli, K.; Uboldi, P.; Pellegatta, F.; Grigore, L.; et al. Myeloid apolipoprotein E controls dendritic cell antigen presentation and T cell activation. Nat. Commun. 2018, 9, 3083. [Google Scholar] [CrossRef] [PubMed]
- Gargett, C.E.; Cornish, J.E.; Wiley, J.S. ATP, a partial agonist for the P2Z receptor of human lymphocytes. J. Cereb. Blood Flow Metab. 1997, 122, 911–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zidovetzki, R.; Levitan, I. Use of cyclodextrins to manipulate plasma membrane cholesterol content: Evidence, misconceptions and control strategies. Biochim. Biophys. Acta 2007, 1768, 1311–1324. [Google Scholar] [CrossRef] [Green Version]
- Avtanski, D.; Pavlov, V.A.; Tracey, K.J.; Poretsky, L. Characterization of inflammation and insulin resistance in high-fat diet-induced male C57BL/6J mouse model of obesity. Anim. Model. Exp. Med. 2019, 2, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Reiss, K.; Leitzke, S.; Seidel, J.; Sperrhacke, M.; Bhakdi, S. Scramblases as Regulators of Proteolytic ADAM Function. Membranes 2022, 12, 185. [Google Scholar] [CrossRef]
- Ferrere, G.; Leroux, A.; Wrzosek, L.; Puchois, V.; Gaudin, F.; Ciocan, D.; Renoud, M.-L.; Naveau, S.; Perlemuter, G.; Cassard, A.-M. Activation of Kupffer Cells Is Associated with a Specific Dysbiosis Induced by Fructose or High Fat Diet in Mice. PLoS ONE 2016, 11, e0146177. [Google Scholar] [CrossRef] [Green Version]
- Benbijja, M.; Mellouk, A.; Bobé, P. Sensitivity of leukemic T-cell lines to arsenic trioxide cytotoxicity is dependent on the induction of phosphatase B220/CD45R expression at the cell surface. Mol. Cancer 2014, 13, 251. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hutteau-Hamel, T.; Mellouk, A.; Trainel, N.; Cassard, A.-M.; Bobé, P. Hypercholesterolemia Negatively Regulates P2X7-Induced Cellular Function in CD4+ and CD8+ T-Cell Subsets from B6 Mice Fed a High-Fat Diet. Int. J. Mol. Sci. 2022, 23, 6730. https://doi.org/10.3390/ijms23126730
Hutteau-Hamel T, Mellouk A, Trainel N, Cassard A-M, Bobé P. Hypercholesterolemia Negatively Regulates P2X7-Induced Cellular Function in CD4+ and CD8+ T-Cell Subsets from B6 Mice Fed a High-Fat Diet. International Journal of Molecular Sciences. 2022; 23(12):6730. https://doi.org/10.3390/ijms23126730
Chicago/Turabian StyleHutteau-Hamel, Tom, Amine Mellouk, Nicolas Trainel, Anne-Marie Cassard, and Pierre Bobé. 2022. "Hypercholesterolemia Negatively Regulates P2X7-Induced Cellular Function in CD4+ and CD8+ T-Cell Subsets from B6 Mice Fed a High-Fat Diet" International Journal of Molecular Sciences 23, no. 12: 6730. https://doi.org/10.3390/ijms23126730
APA StyleHutteau-Hamel, T., Mellouk, A., Trainel, N., Cassard, A.-M., & Bobé, P. (2022). Hypercholesterolemia Negatively Regulates P2X7-Induced Cellular Function in CD4+ and CD8+ T-Cell Subsets from B6 Mice Fed a High-Fat Diet. International Journal of Molecular Sciences, 23(12), 6730. https://doi.org/10.3390/ijms23126730