
NMDA and P2X7 Receptors Require Pannexin 1 Activation to Initiate and Maintain Nociceptive Signaling in the Spinal Cord of Neuropathic Rats

 , , , , and
, , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
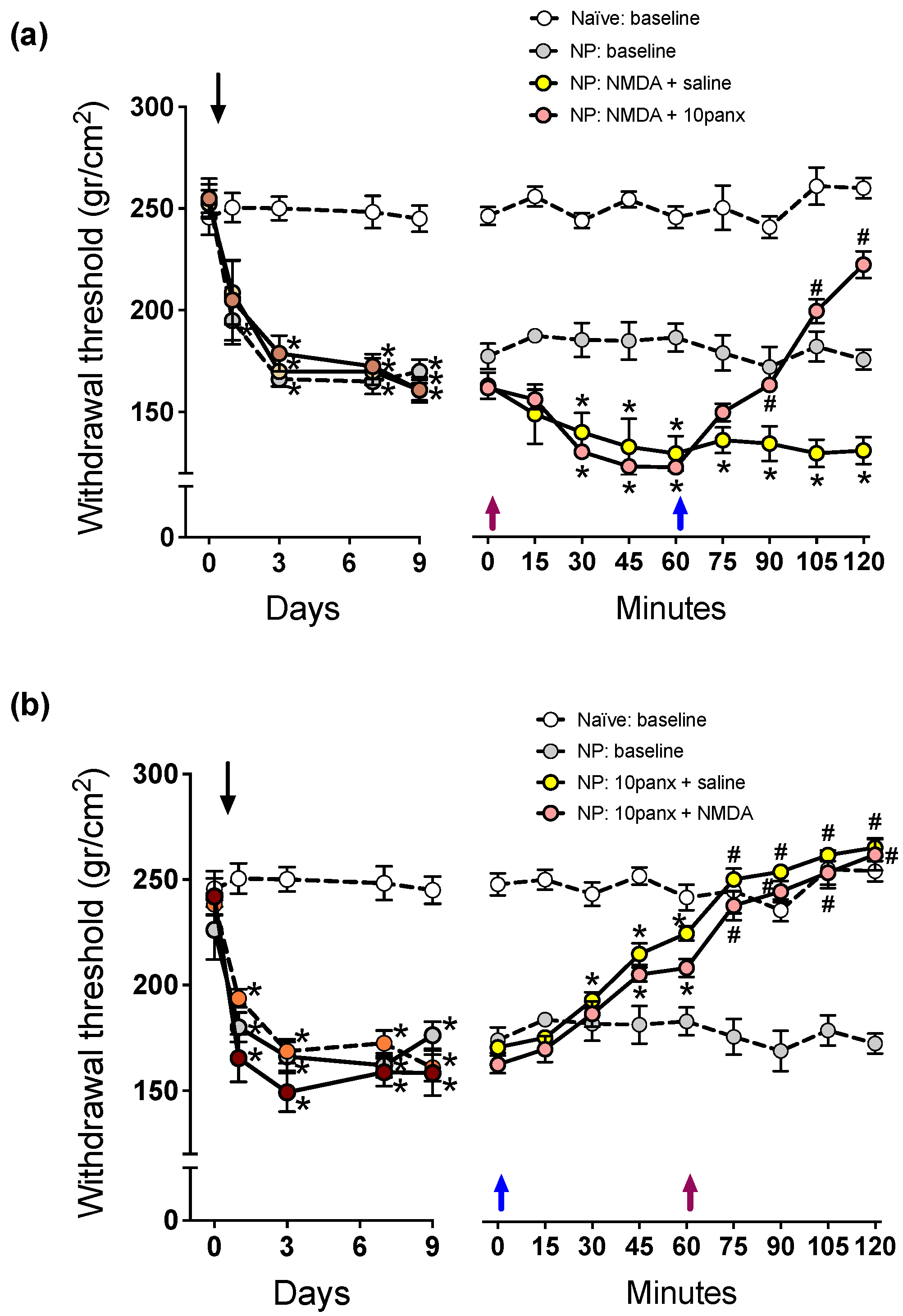
2.1. NMDAR-Mediated Mechanical Hyperalgesia Requires Panx1 Channel Activation to Initiate and Maintain Nociceptive Signaling in Neuropathic Rats
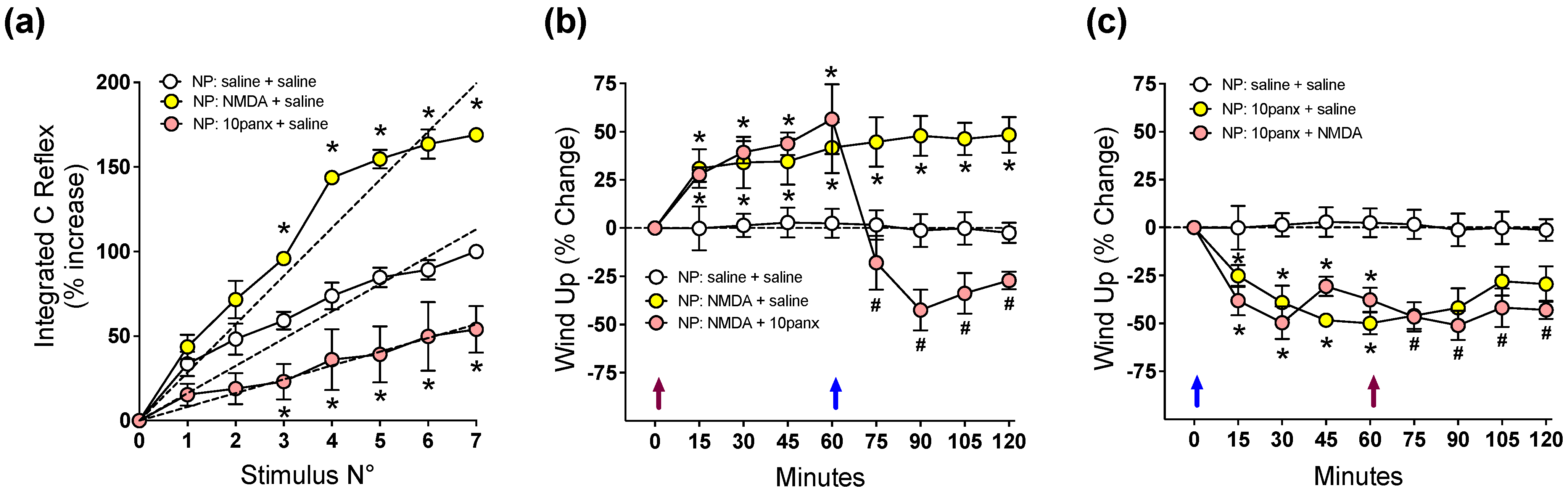
2.2. NMDAR-Evoked C-Reflex Wind-Up Requires Panx1 Channel Activation to Maintain Nociceptive Signaling in the Spinal Cord of Neuropathic Rats
2.3. P2X7R-Mediated Mechanical Hyperalgesia Requires Panx1 Channel Activation to Initiate and Maintain Nociceptive Signaling in Neuropathic Rats
2.4. P2X7R-Evoked C-Reflex Wind-Up Requires Panx1 Channel Activation to Maintain Nociceptive Signaling in the Spinal Cord of Neuropathic Rats
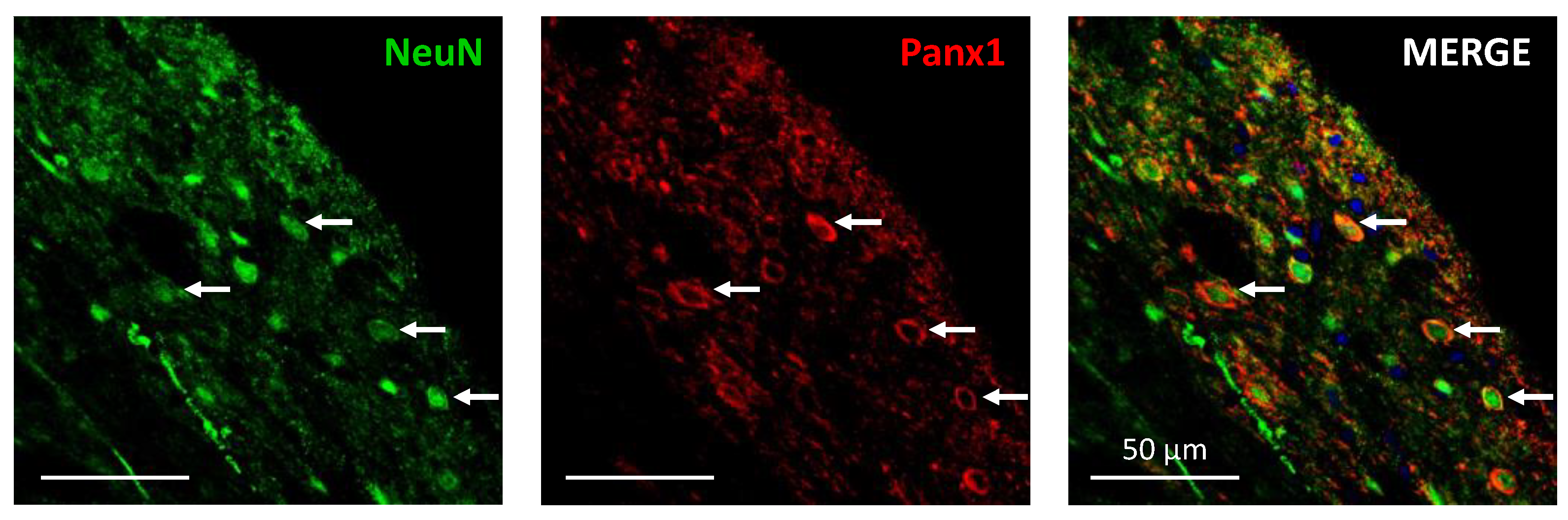
2.5. Pannexin Channels Are Localized in Intrinsic Dorsal Horn Neurons
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Induction of Neuropathy
4.3. Drugs and Treatments
4.4. Behavioral Assessment of Nociceptive Behavior (Algesimetry)
4.5. Electrophysiological Recording of Nociceptive C-Reflex (Electromyography)
4.6. Expression of Panx1 Protein in Dorsal Horn Neurons (Immunofluorescence)
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Latremoliere, A.; Woolf, C.J. Central Sensitization: A Generator of Pain Hypersensitivity by Central Neural Plasticity. J. Pain 2009, 10, 895–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuner, R. Central mechanisms of pathological pain. Nat. Med. 2010, 16, 1258–1266. [Google Scholar] [CrossRef] [PubMed]
- Bannister, K.; Sachau, J.; Baron, R.; Dickenson, A.H. Neuropathic Pain: Mechanism-Based Therapeutics. Annu. Rev. Pharmacol. Toxicol. 2020, 60, 257–274. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Tsuda, M. Purinergic systems, neuropathic pain and the role of microglia. Exp. Neurol. 2011, 234, 293–301. [Google Scholar] [CrossRef]
- Ji, R.-R.; Berta, T.; Nedergaard, M. Glia and pain: Is chronic pain a gliopathy? Pain 2013, 154, S10–S28. [Google Scholar] [CrossRef]
- Donnelly, C.R.; Andriessen, A.S.; Chen, G.; Wang, K.; Jiang, C.; Maixner, W.; Ji, R.-R. Central Nervous System Targets: Glial Cell Mechanisms in Chronic Pain. Neurotherapeutics 2020, 17, 846–860. [Google Scholar] [CrossRef]
- Bravo, D.; Ibarra, P.; Retamal, J.; Pelissier, T.; Laurido, C.; Hernandez, A.; Constandil, L. Pannexin 1: A novel participant in neuropathic pain signaling in the rat spinal cord. Pain 2014, 155, 2108–2115. [Google Scholar] [CrossRef]
- Mannelli, L.D.C.; Marcoli, M.; Micheli, L.; Zanardelli, M.; Maura, G.; Ghelardini, C.; Cervetto, C. Oxaliplatin evokes P2X7-dependent glutamate release in the cerebral cortex: A pain mechanism mediated by Pannexin 1. Neuropharmacology 2015, 97, 133–141. [Google Scholar] [CrossRef]
- Mousseau, M.; Burma, N.E.; Lee, K.Y.; Leduc-Pessah, H.; Kwok, C.H.T.; Reid, A.R.; O’Brien, M.; Sagalajev, B.; Stratton, J.A.; Patrick, N.; et al. Microglial pannexin-1 channel activation is a spinal determinant of joint pain. Sci. Adv. 2018, 4, eaas9846. [Google Scholar] [CrossRef] [Green Version]
- Ray, A.; Zoidl, G.; Weickert, S.; Wahle, P.; Dermietzel, R. Site-specific and developmental expression of pannexin1 in the mouse nervous system. Eur. J. Neurosci. 2005, 21, 3277–3290. [Google Scholar] [CrossRef]
- Swayne, L.A.; Boyce, A.K.J. Regulation of Pannexin 1 Surface Expression by Extracellular ATP: Potential Implications for Nervous System Function in Health and Disease. Front. Cell. Neurosci. 2017, 11, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, R.J.; Jackson, M.F.; Olah, M.E.; Rungta, R.L.; Hines, D.J.; Beazely, M.A.; MacDonald, J.F.; MacVicar, B.A. Activation of Pannexin-1 Hemichannels Augments Aberrant Bursting in the Hippocampus. Science 2008, 322, 1555–1559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoidl, G.; Petrasch-Parwez, E.; Ray, A.; Meier, C.; Bunse, S.; Habbes, H.-W.; Dahl, G.; Dermietzel, R. Localization of the pannexin1 protein at postsynaptic sites in the cerebral cortex and hippocampus. Neuroscience 2007, 146, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Sheng, H.; Chen, L.; Hao, B.; Shi, X.; Chen, Y. Effect of pannexin-1 on the release of glutamate and cytokines in astrocytes. J. Clin. Neurosci. 2016, 23, 135–141. [Google Scholar] [CrossRef]
- Koyanagi, S.; Kusunose, N.; Taniguchi, M.; Akamine, T.; Kanado, Y.; Ozono, Y.; Masuda, T.; Kohro, Y.; Matsunaga, N.; Tsuda, M.; et al. Glucocorticoid regulation of ATP release from spinal astrocytes underlies diurnal exacerbation of neuropathic mechanical allodynia. Nat. Commun. 2016, 7, 13102. [Google Scholar] [CrossRef]
- Narahari, A.K.; Kreutzberger, A.J.; Gaete, P.S.; Chiu, Y.-H.; Leonhardt, S.A.; Medina, C.B.; Jin, X.; Oleniacz, P.W.; Kiessling, V.; Barrett, P.Q.; et al. ATP and large signaling metabolites flux through caspase-activated Pannexin 1 channels. eLife 2021, 10, e64784. [Google Scholar] [CrossRef]
- Bravo, D.; Maturana, C.; Pelissier, T.; Hernández, A.; Constandil, L. Interactions of pannexin 1 with NMDA and P2X7 receptors in central nervous system pathologies: Possible role on chronic pain. Pharmacol. Res. 2015, 101, 86–93. [Google Scholar] [CrossRef]
- Seo, J.; Dalal, M.; Contreras, J. Pannexin-1 Channels as Mediators of Neuroinflammation. Int. J. Mol. Sci. 2021, 22, 5189. [Google Scholar] [CrossRef]
- E Isakson, B.; Thompson, R.J. Pannexin-1 as a potentiator of ligand-gated receptor signaling. Channels 2014, 8, 118–123. [Google Scholar] [CrossRef] [Green Version]
- Kaindl, A.M.; Degos, V.; Peineau, S.; Gouadon, E.; Chhor, V.; Loron, G.; Le Charpentier, T.; Josserand, J.; Ali, C.; Vivien, D.; et al. Activation of microglial N-methyl-D-aspartate receptors triggers inflammation and neuronal cell death in the developing and mature brain. Ann. Neurol. 2012, 72, 536–549. [Google Scholar] [CrossRef]
- Weng, X.; Tan, Y.; Chu, X.; Wu, X.-F.; Liu, R.; Tian, Y.; Li, L.; Guo, F.; Ouyang, Q.; Li, L. N-methyl-D-aspartic acid receptor 1 (NMDAR1) aggravates secondary inflammatory damage induced by hemin-NLRP3 pathway after intracerebral hemorrhage. Chin. J. Traumatol. 2015, 18, 254–258. [Google Scholar] [CrossRef] [PubMed]
- Starobova, H.; Nadar, E.I.; Vetter, I. The NLRP3 Inflammasome: Role and Therapeutic Potential in Pain Treatment. Front. Physiol. 2020, 11, 1016. [Google Scholar] [CrossRef] [PubMed]
- Mortezaee, K.; Khanlarkhani, N.; Beyer, C.; Zendedel, A. Inflammasome: Its role in traumatic brain and spinal cord injury. J. Cell. Physiol. 2017, 233, 5160–5169. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Mei, X. Role of NLRP3 Inflammasomes in Neuroinflammation Diseases. Eur. Neurol. 2020, 83, 576–580. [Google Scholar] [CrossRef]
- Zhang, H.; Li, F.; Li, W.-W.; Stary, C.; Clark, J.; Xu, S.; Xiong, X. The inflammasome as a target for pain therapy. Br. J. Anaesth. 2016, 117, 693–707. [Google Scholar] [CrossRef] [Green Version]
- D’Amico, R.; Fusco, R.; Siracusa, R.; Impellizzeri, D.; Peritore, A.; Gugliandolo, E.; Interdonato, L.; Sforza, A.; Crupi, R.; Cuzzocrea, S.; et al. Inhibition of P2X7 Purinergic Receptor Ameliorates Fibromyalgia Syndrome by Suppressing NLRP3 Pathway. Int. J. Mol. Sci. 2021, 22, 6471. [Google Scholar] [CrossRef]
- Lapato, A.; Tiwari-Woodruff, S.K. Connexins and pannexins: At the junction of neuro-glial homeostasis & disease. J. Neurosci. Res. 2017, 96, 31–44. [Google Scholar] [CrossRef]
- Bao, L.; Locovei, S.; Dahl, G. Pannexin membrane channels are mechanosensitive conduits for ATP. FEBS Lett. 2004, 572, 65–68. [Google Scholar] [CrossRef] [Green Version]
- Eide, P.K. Wind-up and the NMDA receptor complex from a clinical perspective. Eur. J. Pain 2000, 4, 5–15. [Google Scholar] [CrossRef]
- Zhou, Q.; Price, D.D.; Callam, C.S.; Woodruff, M.A.; Verne, G.N. Effects of the N-Methyl-D-Aspartate Receptor on Temporal Summation of Second Pain (Wind-up) in Irritable Bowel Syndrome. J. Pain 2011, 12, 297–303. [Google Scholar] [CrossRef] [Green Version]
- Donnelly-Roberts, D.; McGaraughty, S.; Shieh, C.-C.; Honore, P.; Jarvis, M.F. Painful Purinergic Receptors. J. Pharmacol. Exp. Ther. 2007, 324, 409–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, Y.-X.; Zhang, Y.; Zhang, Y.-Q.; Zhao, Z.-Q. Involvement of microglial P2X7 receptors and downstream signaling pathways in long-term potentiation of spinal nociceptive responses. Brain, Behav. Immun. 2010, 24, 1176–1189. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.K.; Adelstein, R.S.; Kawamoto, S. Identification of Neuronal Nuclei (NeuN) as Fox-3, a New Member of the Fox-1 Gene Family of Splicing Factors. J. Biol. Chem. 2009, 284, 31052–31061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dickenson, A.; Chapman, V.; Green, G. The pharmacology of excitatory and inhibitory amino acid-mediated events in the transmission and modulation of pain in the spinal cord. Gen. Pharmacol. Vasc. Syst. 1997, 28, 633–638. [Google Scholar] [CrossRef]
- Kreutzwiser, D.; Tawfic, Q.A. Expanding Role of NMDA Receptor Antagonists in the Management of Pain. CNS Drugs 2019, 33, 347–374. [Google Scholar] [CrossRef]
- Weilinger, N.L.; Tang, P.L.; Thompson, R.J. Anoxia-Induced NMDA Receptor Activation Opens Pannexin Channels via Src Family Kinases. J. Neurosci. 2012, 32, 12579–12588. [Google Scholar] [CrossRef]
- Weilinger, N.L.; Lohman, A.W.; Rakai, B.D.; Ma, E.M.M.; Bialecki, J.; Maslieieva, V.; Rilea, T.; Bandet, M.V.; Ikuta, N.T.; Scott, L.; et al. Metabotropic NMDA receptor signaling couples Src family kinases to pannexin-1 during excitotoxicity. Nat. Neurosci. 2016, 19, 432–442. [Google Scholar] [CrossRef]
- Dore, K.; Stein, I.S.; Brock, J.A.; Castillo, P.E.; Zito, K.; Sjöström, P.J. Unconventional NMDA Receptor Signaling. J. Neurosci. 2017, 37, 10800–10807. [Google Scholar] [CrossRef] [Green Version]
- Balderas, P.M.D.O. Flux-Independent NMDAR Signaling: Molecular Mediators, Cellular Functions, and Complexities. Int. J. Mol. Sci. 2018, 19, 3800. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Cao, J.; Yang, X.; Suo, Z.-W.; Shi, L.; Liu, Y.-N.; Yang, H.-B.; Hu, X.-D. NR2B phosphorylation at tyrosine 1472 in spinal dorsal horn contributed to N-methyl-D-aspartate-induced pain hypersensitivity in mice. J. Neurosci. Res. 2011, 89, 1869–1876. [Google Scholar] [CrossRef]
- Liu, X.J.; Gingrich, J.R.; Vargas-Caballero, M.; Na Dong, Y.; Sengar, A.; Beggs, S.; Wang, S.-H.; Ding, H.K.; Frankland, P.W.; Salter, M.W. Treatment of inflammatory and neuropathic pain by uncoupling Src from the NMDA receptor complex. Nat. Med. 2008, 14, 1325–1332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, M.; Chen, S.-R.; Pan, H.-L. Presynaptic NMDA receptors control nociceptive transmission at the spinal cord level in neuropathic pain. Cell. Mol. Life Sci. 2019, 76, 1889–1899. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Laumet, G.; Chen, S.-R.; Hittelman, W.N.; Pan, H.-L. Pannexin-1 Up-regulation in the Dorsal Root Ganglion Contributes to Neuropathic Pain Development. J. Biol. Chem. 2015, 290, 14647–14655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bardoni, R.; Goldstein, P.A.; Lee, C.J.; Gu, J.G.; MacDermott, A.B. ATP P2X Receptors Mediate Fast Synaptic Transmission in the Dorsal Horn of the Rat Spinal Cord. J. Neurosci. 1997, 17, 5297–5304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharp, C.J.; Reeve, A.J.; Collins, S.D.; Martindale, J.C.; Summerfield, S.G.; Sargent, B.S.; Bate, S.T.; Chessell, I.P. Investigation into the role of P2X3 /P2X2/3 receptors in neuropathic pain following chronic constriction injury in the rat: An electrophysiological study. Br. J. Pharmacol. 2006, 148, 845–852. [Google Scholar] [CrossRef] [Green Version]
- Gao, P.; Ding, X.; Khan, T.M.; Rong, W.; Franke, H.; Illes, P. P2X7 receptor-sensitivity of astrocytes and neurons in the substantia gelatinosa of organotypic spinal cord slices of the mouse depends on the length of the culture period. Neuroscience 2017, 349, 195–207. [Google Scholar] [CrossRef]
- Munoz, F.M.; Gao, R.; Tian, Y.; Henstenburg, B.A.; Barrett, J.E.; Hu, H. Neuronal P2X7 receptor-induced reactive oxygen species production contributes to nociceptive behavior in mice. Sci. Rep. 2017, 7, 3539. [Google Scholar] [CrossRef]
- Iglesias, R.; Locovei, S.; Roque, A.; Alberto, A.P.; Dahl, G.; Spray, D.C.; Scemes, E. P2X7 receptor-Pannexin1 complex: Pharmacology and signaling. Am. J. Physiol. Physiol. 2008, 295, C752–C760. [Google Scholar] [CrossRef] [Green Version]
- Abeele, F.V.; Bidaux, G.; Gordienko, D.; Beck, B.; Panchin, Y.; Baranova, A.; Ivanov, D.; Skryma, R.; Prevarskaya, N. Functional implications of calcium permeability of the channel formed by pannexin 1. J. Cell Biol. 2006, 174, 535–546. [Google Scholar] [CrossRef] [Green Version]
- Dahl, G.; Keane, R.W. Pannexin: From discovery to bedside in 11 ± 4 years? Brain Res. 2012, 1487, 150–159. [Google Scholar] [CrossRef] [Green Version]
- Weaver, J.L.; Arandjelovic, S.; Brown, G.; Mendu, S.K.; Schappe, M.S.; Buckley, M.W.; Chiu, Y.-H.; Shu, S.; Kim, J.K.; Chung, J.; et al. Hematopoietic pannexin 1 function is critical for neuropathic pain. Sci. Rep. 2017, 7, srep42550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuda, T.; Ozono, Y.; Mikuriya, S.; Kohro, Y.; Tozaki-Saitoh, H.; Iwatsuki, K.; Uneyama, H.; Ichikawa, K.I.H.U.R.; Salter, M.W.; Tsuda, T.M.Y.K.H.T.-S.M.; et al. Dorsal horn neurons release extracellular ATP in a VNUT-dependent manner that underlies neuropathic pain. Nat. Commun. 2016, 7, 12529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hide, I.; Tanaka, M.; Inoue, A.; Nakajima, K.; Kohsaka, S.; Inoue, K.; Nakata, Y. Extracellular ATP Triggers Tumor Necrosis Factor-α Release from Rat Microglia. J. Neurochem. 2000, 75, 965–972. [Google Scholar] [CrossRef]
- Suzuki, T.; Hide, I.; Ido, K.; Kohsaka, S.; Inoue, K.; Nakata, Y. Production and Release of Neuroprotective Tumor Necrosis Factor by P2X7 Receptor-Activated Microglia. J. Neurosci. 2004, 24, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyce, A.K.; Epp, A.L.; Nagarajan, A.; Swayne, L.A. Transcriptional and post-translational regulation of pannexins. Biochim. Biophys. Acta Biomembr. 2018, 1860, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Zappalà, A.; Cicero, D.; Serapide, M.F.; Paz, C.; Catania, M.V.; Falchi, M.; Parenti, R.; Pantò, M.; La Delia, F.; Cicirata, F. Expression of pannexin1 in the CNS of adult mouse: Cellular localization and effect of 4-aminopyridine-induced seizures. Neuroscience 2006, 141, 167–178. [Google Scholar] [CrossRef]
- Yeung, A.K.; Patil, C.S.; Jackson, M.F. Pannexin-1 in the CNS: Emerging concepts in health and disease. J. Neurochem. 2020, 154, 468–485. [Google Scholar] [CrossRef] [PubMed]
- Cone, A.C.; Ambrosi, C.; Scemes, E.; Martone, M.E.; Sosinsky, G.E. A Comparative Antibody Analysis of Pannexin1 Expression in Four Rat Brain Regions Reveals Varying Subcellular Localizations. Front. Pharmacol. 2013, 4, 6. [Google Scholar] [CrossRef] [Green Version]
- National Research Council. Guide for the Care and Use of Laboratory Animals, 8th ed.; The National Academies Press: Washington, DC, USA, 2011; pp. 1–120. ISBN 978-0-309-38629-6. [Google Scholar] [CrossRef]
- Zimmermann, M. Ethical guidelines for investigations of experimental pain in conscious animals. Pain 1983, 16, 109–110. [Google Scholar] [CrossRef]
- Decosterd, I.; Woolf, C.J. Spared nerve injury: An animal model of persistent peripheral neuropathic pain. Pain 2000, 87, 149–158. [Google Scholar] [CrossRef]
- Peyronnard, J.-M.; Charron, L. Motor and sensory neurons of the rat sural nerve: A horseradish peroxidase study. Muscle Nerve 1982, 5, 654–660. [Google Scholar] [CrossRef] [PubMed]
- Salter, M.W.; Pitcher, G.M. Dysregulated Src upregulation of NMDA receptor activity: A common link in chronic pain and schizophrenia. FEBS J. 2012, 279, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Tsuda, M. Nociceptive signaling mediated by P2X3, P2X4 and P2X7 receptors. Biochem. Pharmacol. 2020, 187, 114309. [Google Scholar] [CrossRef] [PubMed]
- Siegan, J.B.; Sagen, J. Attenuation of NMDA-induced spinal hypersensitivity by adrenal medullary transplants. Brain Res. 1995, 680, 88–98. [Google Scholar] [CrossRef]
- Song, J.; Ying, Y.; Wang, W.; Liu, X.-G.; Xu, X.; Wei, X.; Ruan, X. The role of P2X7R/ERK signaling in dorsal root ganglia satellite glial cells in the development of chronic postsurgical pain induced by skin/muscle incision and retraction (SMIR). Brain Behav. Immun. 2018, 69, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Randall, L.O.; Selitto, J.J. A method for measurement of analgesic activity on inflamed tissue. Arch. Int. Pharmacodyn. Ther. 1957, 111, 409–419. [Google Scholar]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bravo, D.; Zepeda-Morales, K.; Maturana, C.J.; Retamal, J.S.; Hernández, A.; Pelissier, T.; Barra, R.; Sáez-Briones, P.; Burgos, H.; Constandil, L. NMDA and P2X7 Receptors Require Pannexin 1 Activation to Initiate and Maintain Nociceptive Signaling in the Spinal Cord of Neuropathic Rats. Int. J. Mol. Sci. 2022, 23, 6705. https://doi.org/10.3390/ijms23126705
Bravo D, Zepeda-Morales K, Maturana CJ, Retamal JS, Hernández A, Pelissier T, Barra R, Sáez-Briones P, Burgos H, Constandil L. NMDA and P2X7 Receptors Require Pannexin 1 Activation to Initiate and Maintain Nociceptive Signaling in the Spinal Cord of Neuropathic Rats. International Journal of Molecular Sciences. 2022; 23(12):6705. https://doi.org/10.3390/ijms23126705
Chicago/Turabian StyleBravo, David, Katherine Zepeda-Morales, Carola J. Maturana, Jeffri S. Retamal, Alejandro Hernández, Teresa Pelissier, Rafael Barra, Patricio Sáez-Briones, Héctor Burgos, and Luis Constandil. 2022. "NMDA and P2X7 Receptors Require Pannexin 1 Activation to Initiate and Maintain Nociceptive Signaling in the Spinal Cord of Neuropathic Rats" International Journal of Molecular Sciences 23, no. 12: 6705. https://doi.org/10.3390/ijms23126705
APA StyleBravo, D., Zepeda-Morales, K., Maturana, C. J., Retamal, J. S., Hernández, A., Pelissier, T., Barra, R., Sáez-Briones, P., Burgos, H., & Constandil, L. (2022). NMDA and P2X7 Receptors Require Pannexin 1 Activation to Initiate and Maintain Nociceptive Signaling in the Spinal Cord of Neuropathic Rats. International Journal of Molecular Sciences, 23(12), 6705. https://doi.org/10.3390/ijms23126705