
Cichoric Acid May Play a Role in Protecting Hair Cells from Ototoxic Drugs


 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
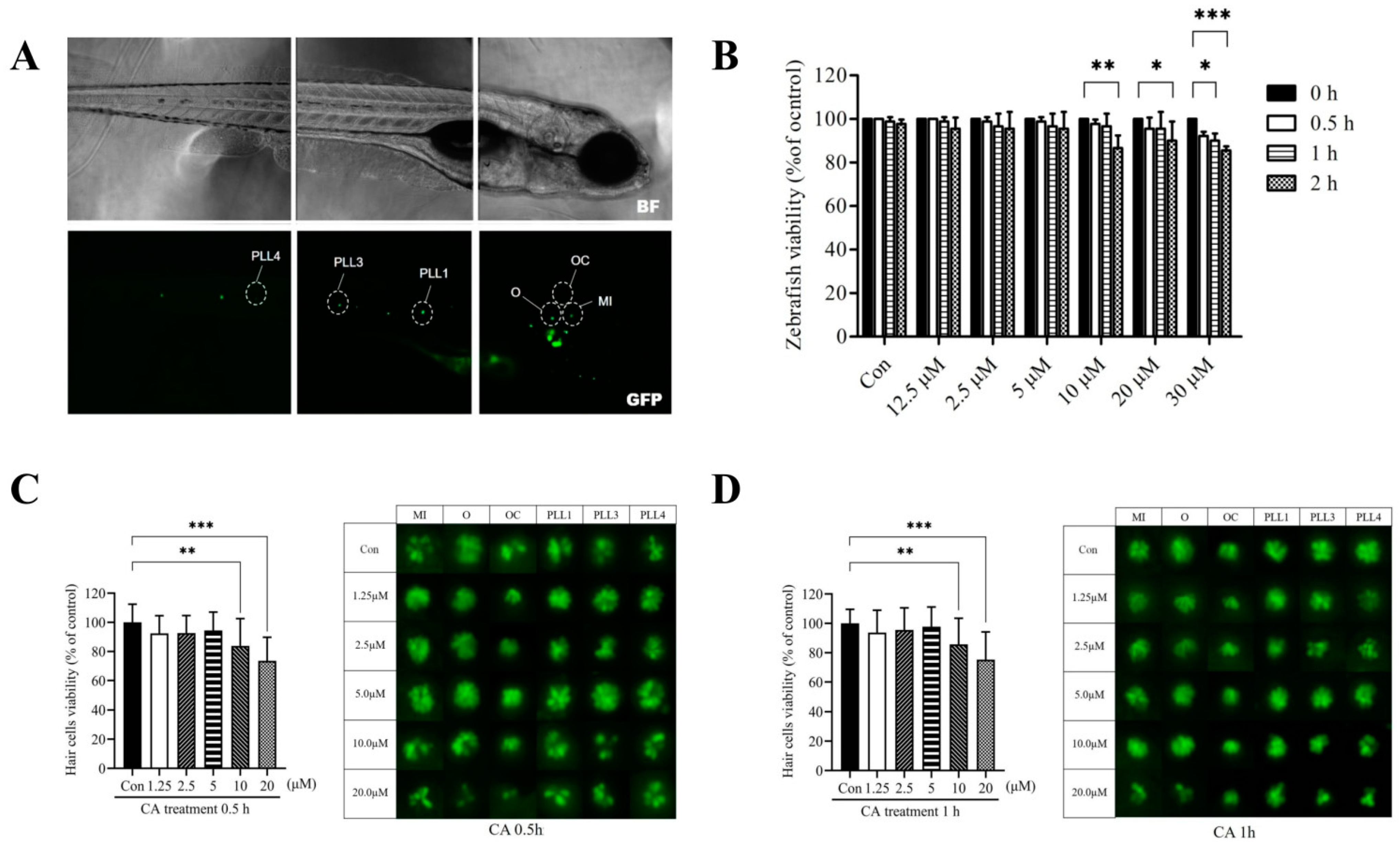
2.1. The Dose-and Time-Response Testing of CA on Transgenic Zebrafish Larvae
2.2. Cytotoxicity of CA on Hair Cells in Neuromasts
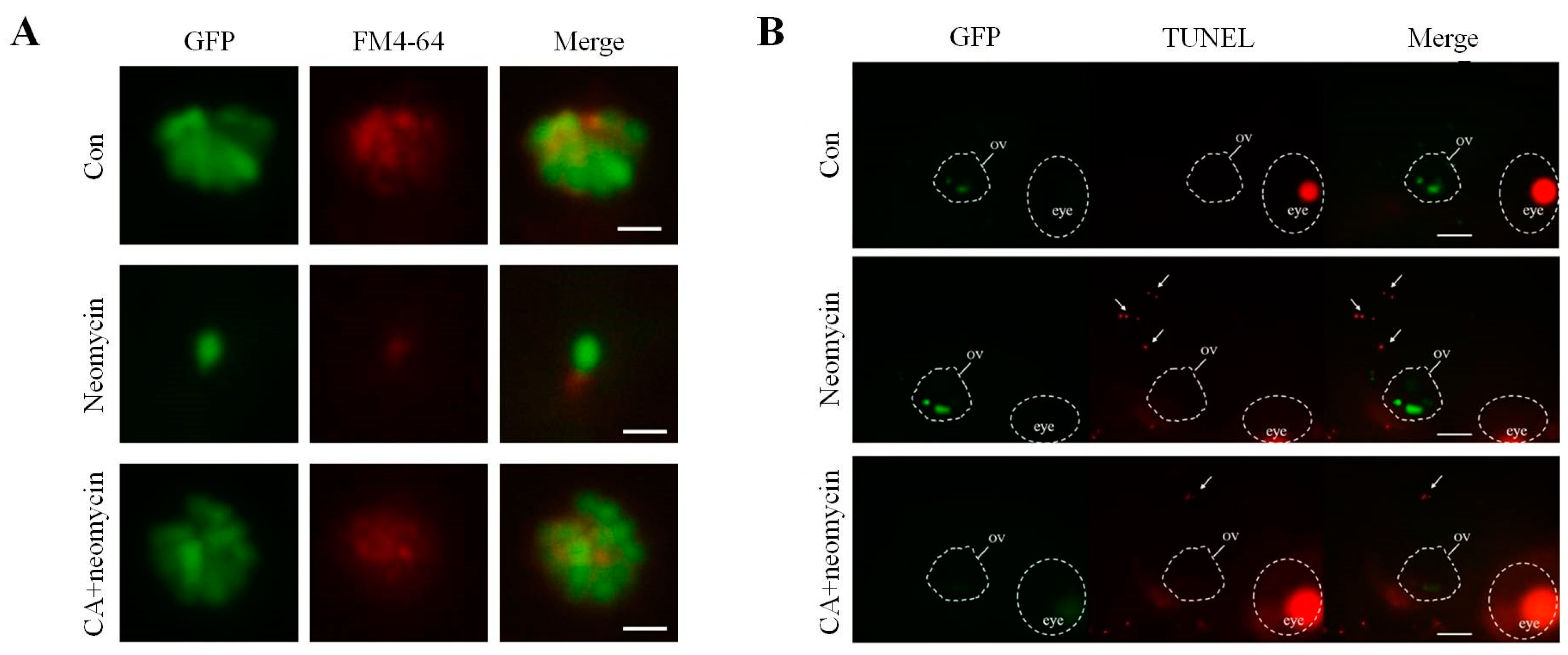
2.3. Protective Effect of CA Concentrations on Neomycin-Induced Hair Cells Loss in Transgenic Zebrafish Larvae
2.4. The Effects of CA on Apoptosis after Neomycin Treatment
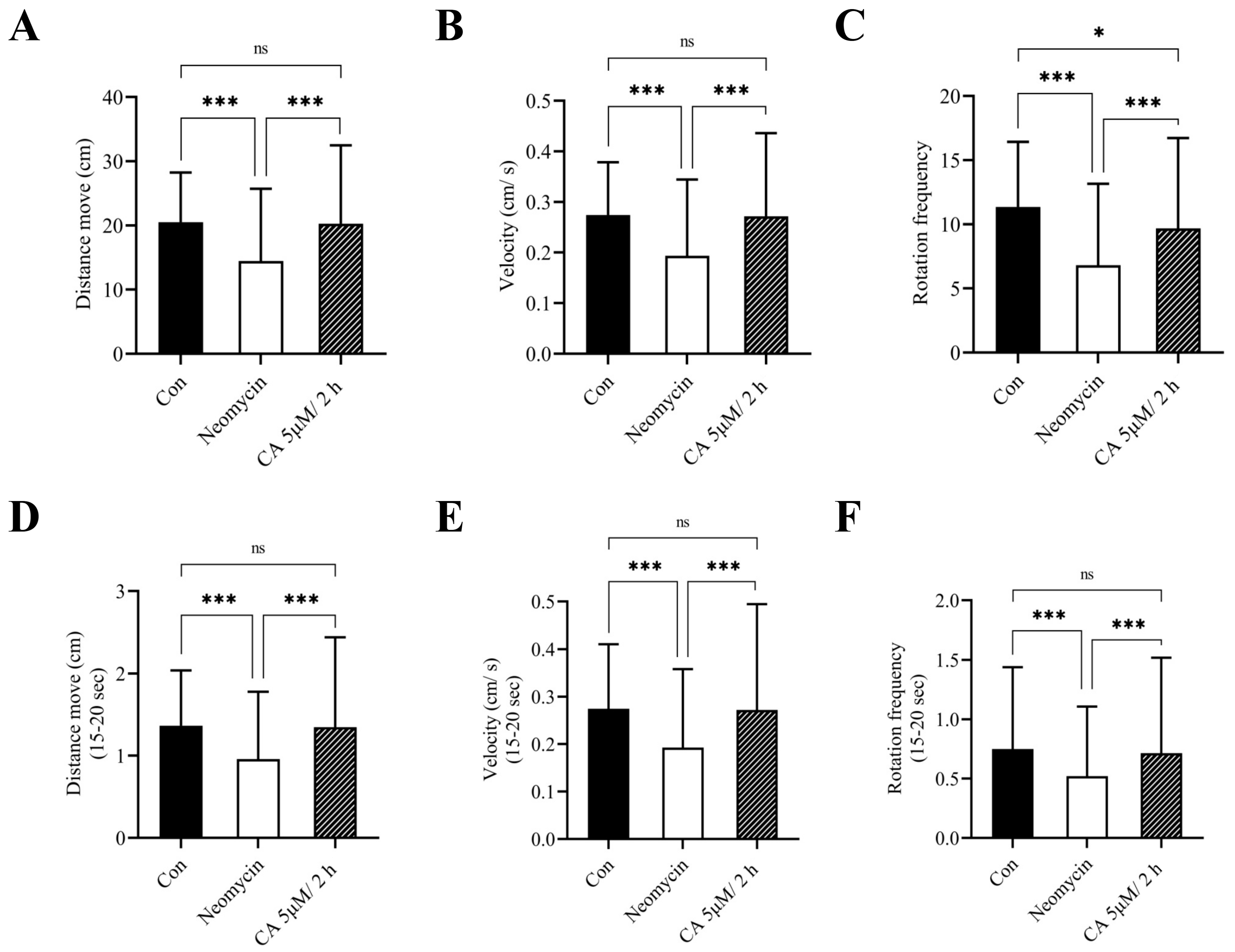
2.5. Effect of CA on the Behavioral Activities of Transgenic Zebrafish Larvae
3. Discussion
4. Materials and Methods
4.1. Zebrafish Preparation and Maintenance
4.2. Acute Toxicity Test
4.3. Washout Test
4.4. FM4-64 Staining
4.5. TUNEL Assay
4.6. Behavioral Analysis
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ganesan, P.; Schmiedge, J.; Manchaiah, V.; Swapna, S.; Dhandayutham, S.; Kothandaraman, P.P. Ototoxicity: A Challenge in Diagnosis and Treatment. J. Audiol. Otol. 2018, 22, 59–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aleman, M.R.; True, A.; Scalco, R.; Crowe, C.M.; Costa, L.R.R.; Chigerwe, M. Gentamicin-induced sensorineural auditory loss in healthy adult horses. J. Vet. Intern. Med. 2021, 35, 2486–2494. [Google Scholar] [CrossRef]
- Ryals, M.; Morell, R.J.; Martin, D.; Boger, E.T.; Wu, P.; Raible, D.W.; Cunningham, L.L. The Inner Ear Heat Shock Transcriptional Signature Identifies Compounds That Protect against Aminoglycoside Ototoxicity. Front. Cell Neurosci. 2018, 12, 445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmgren, M.; Sheets, L. Using the Zebrafish Lateral Line to Understand the Roles of Mitochondria in Sensorineural Hearing Loss. Front. Cell Dev. Biol. 2020, 8, 628712. [Google Scholar] [CrossRef]
- Breitzler, L.; Lau, I.H.; Fonseca, P.J.; Vasconcelos, R.O. Noise-induced hearing loss in zebrafish: Investigating structural and functional inner ear damage and recovery. Hear. Res. 2020, 391, 107952. [Google Scholar] [CrossRef]
- Rowe, C.J.; Crowley-Perry, M.; McCarthy, E.; Davidson, T.L.; Connaughton, V.P. The Three-Chamber Choice Behavioral Task using Zebrafish as a Model System. J. Vis. Exp. 2021, 170, e61934. [Google Scholar] [CrossRef]
- Zhong, Z.; Fu, X.; Li, H.; Chen, J.; Wang, M.; Gao, S.; Zhang, L.; Cheng, C.; Zhang, Y.; Li, P.; et al. Citicoline Protects Auditory Hair Cells against Neomycin-Induced Damage. Front. Cell Dev. Biol. 2020, 8, 712. [Google Scholar] [CrossRef]
- Gao, S.; Cheng, C.; Wang, M.; Jiang, P.; Zhang, L.; Wang, Y.; Wu, H.; Zeng, X.; Wang, H.; Gao, X.; et al. Blebbistatin Inhibits Neomycin-Induced Apoptosis in Hair Cell-Like HEI-OC-1 Cells and in Cochlear Hair Cells. Front. Cell Neurosci. 2019, 13, 590. [Google Scholar] [CrossRef]
- Ruhl, D.; Du, T.T.; Wagner, E.L.; Choi, J.H.; Li, S.; Reed, R.; Kim, K.; Freeman, M.; Hashisaki, G.; Lukens, J.R.; et al. Necroptosis and Apoptosis Contribute to Cisplatin and Aminoglycoside Ototoxicity. J. Neurosci. 2019, 39, 2951–2964. [Google Scholar] [CrossRef] [Green Version]
- Priuska, E.M.; Schacht, J. Mechanism and prevention of aminoglycoside ototoxicity: Outer hair cells as targets and tools. Ear Nose Throat J. 1997, 76, 164–166,168,170. [Google Scholar] [CrossRef]
- Hayward, R.S.; Harding, J.; Molloy, R.; Land, L.; Longcroft-Neal, K.; Moore, D.; Ross, J.D.C. Adverse effects of a single dose of gentamicin in adults: A systematic review. Br. J. Clin. Pharmacol. 2018, 84, 223–238. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Shanmugam, M.K.; Xiang, P.; Yam, T.Y.A.; Kumar, V.; Chew, W.S.; Chang, J.K.; Ali, M.Z.B.; Reolo, M.J.Y.; Peh, Y.X.; et al. Sphingosine 1-Phosphate Receptor 2 Induces Otoprotective Responses to Cisplatin Treatment. Cancers 2020, 12, 211. [Google Scholar] [CrossRef] [Green Version]
- Jia, Z.; He, Q.; Shan, C.; Li, F. Tauroursodeoxycholic acid attenuates gentamicin-induced cochlear hair cell death in vitro. Toxicol. Lett. 2018, 294, 20–26. [Google Scholar] [CrossRef]
- He, Y.; Li, W.; Zheng, Z.; Zhao, L.; Li, W.; Wang, Y.; Li, H. Inhibition of Protein arginine methyltransferase 6 reduces reactive oxygen species production and attenuates aminoglycoside- and cisplatin-induced hair cell death. Theranostics 2020, 10, 133–150. [Google Scholar] [CrossRef]
- Zhu, D.; Zhang, X.; Niu, Y.; Diao, Z.; Ren, B.; Li, X.; Liu, Z.; Liu, X. Cichoric acid improved hyperglycaemia and restored muscle injury via activating antioxidant response in MLD-STZ-induced diabetic mice. Food Chem. Toxicol. 2017, 107 Pt A, 138–149. [Google Scholar] [CrossRef]
- Guo, R.; Zhao, B.; Wang, Y.; Wu, D.; Wang, Y.; Yu, Y.; Yan, Y.; Zhang, W.; Liu, Z.; Liu, X. Cichoric Acid Prevents Free-Fatty-Acid-Induced Lipid Metabolism Disorders via Regulating Bmal1 in HepG2 Cells. J. Agric. Food Chem. 2018, 66, 9667–9678. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, J.; Wu, Z.; Wang, X. Chicoric acid attenuates hyperglycemia-induced endothelial dysfunction through AMPK-dependent inhibition of oxidative/nitrative stresses. J. Recept. Signal Transduct. Res. 2021, 41, 378–392. [Google Scholar] [CrossRef]
- Landmann, M.; Kanuri, G.; Spruss, A.; Stahl, C.; Bergheim, I. Oral intake of chicoric acid reduces acute alcohol-induced hepatic steatosis in mice. Nutrition 2014, 30, 882–889. [Google Scholar] [CrossRef]
- Diao, Z.; Li, J.; Liu, Q.; Wang, Y. In-vivo metabolite profiling of chicoric acid in rat plasma, urine and feces after oral administration using liquid chromatography quadrupole time of flight mass spectrometry. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2018, 1081–1082, 8–14. [Google Scholar] [CrossRef]
- Gan, C.; Liu, L.; Du, Y.; Wang, L.; Gao, M.; Wu, L.; Yang, C. Simultaneous determination and pharmacokinetic study of four phenol compounds in rat plasma by ultra-high performance liquid chromatography with tandem mass spectrometry after oral administration of Echinacea purpurea extract. J. Sep. Sci. 2016, 39, 1628–1637. [Google Scholar] [CrossRef]
- Schlernitzauer, A.; Oiry, C.; Hamad, R.; Galas, S.; Cortade, F.; Chabi, B.; Casas, F.; Pessemesse, L.; Fouret, G.; Feillet-Coudray, C.; et al. Chicoric acid is an antioxidant molecule that stimulates AMP kinase pathway in L6 myotubes and extends lifespan in Caenorhabditis elegans. PLoS ONE 2013, 8, e78788. [Google Scholar] [CrossRef] [PubMed]
- Sloley, B.D.; Urichuk, L.J.; Tywin, C.; Coutts, R.T.; Pang, P.K.; Shan, J.J. Comparison of chemical components and antioxidants capacity of different Echinacea species. J. Pharm. Pharmacol. 2001, 53, 849–857. [Google Scholar] [CrossRef] [PubMed]
- Pellati, F.; Benvenuti, S.; Magro, L.; Melegari, M.; Soragni, F. Analysis of phenolic compounds and radical scavenging activity of Echinacea spp. J. Pharm. Biomed. Anal. 2004, 35, 289–301. [Google Scholar] [CrossRef]
- Tsai, Y.L.; Chiu, C.C.; Yi-Fu Chen, J.; Chan, K.C.; Lin, S.D. Cytotoxic effects of Echinacea purpurea flower extracts and cichoric acid on human colon cancer cells through induction of apoptosis. J. Ethnopharmacol. 2012, 143, 914–919. [Google Scholar] [CrossRef]
- Facino, R.M.; Carini, M.; Aldini, G.; Saibene, L.; Pietta, P.; Mauri, P. Echinacoside and caffeoyl conjugates protect collagen from free radical-induced degradation: A potential use of Echinacea extracts in the prevention of skin photodamage. Planta Med. 1995, 61, 510–514. [Google Scholar] [CrossRef] [PubMed]
- Chang-Chien, J.; Yen, Y.C.; Li, S.Y.; Hsu, T.C.; Yang, J.J. Ferulic acid-mediated protection against neomycin-induced hair cell loss in transgenic zebrafish. J. Funct. Foods 2017, 28, 157–167. [Google Scholar] [CrossRef]
- Zong, Y.; Chen, F.; Li, S.; Zhang, H. (-)-Epigallocatechin-3-gallate (EGCG) prevents aminoglycosides-induced ototoxicity via anti-oxidative and anti-apoptotic pathways. Int. J. Pediatr. Otorhinolaryngol. 2021, 150, 110920. [Google Scholar] [CrossRef]
- Zheng, Z.; Tang, D.; Zhao, L.; Li, W.; Han, J.; Hu, B.; Nie, G.; He, Y. Liproxstatin-1 Protects Hair Cell-Like HEI-OC1 Cells and Cochlear Hair Cells against Neomycin Ototoxicity. Oxid. Med. Cell Longev. 2020, 2020, 1782659. [Google Scholar] [CrossRef]
- Gu, R.; Longenecker, R.J.; Homan, J.; Kil, J. Ebselen attenuates tobramycin-induced ototoxicity in mice. J. Cyst. Fibros. 2021, 20, 271–277. [Google Scholar] [CrossRef] [Green Version]
- Manzanilla, B.; Robles, J. Antiradical properties of curcumin, caffeic acid phenethyl ester, and chicoric acid: A DFT study. J. Mol. Model 2022, 28, 68. [Google Scholar] [CrossRef]
- Babich, O.; Sukhikh, S.; Prosekov, A.; Asyakina, L.; Ivanova, S. Medicinal Plants to Strengthen Immunity during a Pandemic. Pharmaceuticals 2020, 13, 313. [Google Scholar] [CrossRef] [PubMed]
- Sukhikh, S.; Noskova, S.; Ivanova, S.; Ulrikh, E.; Izgaryshev, A.; Babich, O. Chondroprotection and Molecular Mechanism of Action of Phytonutraceuticals on Osteoarthritis. Molecules 2021, 26, 2391. [Google Scholar] [CrossRef] [PubMed]
- Aarland, R.C.; Banuelos-Hernandez, A.E.; Fragoso-Serrano, M.; Sierra-Palacios, E.D.; Diaz de Leon-Sanchez, F.; Perez-Flores, L.J.; Rivera-Cabrera, F.; Mendoza-Espinoza, J.A. Studies on phytochemical, antioxidant, anti-inflammatory, hypoglycaemic and antiproliferative activities of Echinacea purpurea and Echinacea angustifolia extracts. Pharm. Biol. 2017, 55, 649–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langland, J.; Jacobs, B.; Wagner, C.E.; Ruiz, G.; Cahill, T.M. Antiviral activity of metal chelates of caffeic acid and similar compounds towards herpes simplex, VSV-Ebola pseudotyped and vaccinia viruses. Antivir. Res. 2018, 160, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Chicca, A.; Adinolfi, B.; Martinotti, E.; Fogli, S.; Breschi, M.C.; Pellati, F.; Benvenuti, S.; Nieri, P. Cytotoxic effects of Echinacea root hexanic extracts on human cancer cell lines. J. Ethnopharmacol. 2007, 110, 148–153. [Google Scholar] [CrossRef]
- Venuto, A.; Erickson, T. Evaluating the Death and Recovery of Lateral Line Hair Cells Following Repeated Neomycin Treatments. Life 2021, 11, 1180. [Google Scholar] [CrossRef] [PubMed]
- Kenyon, E.J.; Kirkwood, N.K.; Kitcher, S.R.; O′Reilly, M.; Derudas, M.; Cantillon, D.M.; Goodyear, R.J.; Secker, A.; Baxendale, S.; Bull, J.C.; et al. Identification of ion-channel modulators that protect against aminoglycoside-induced hair cell death. JCI Insight 2017, 2, 24. [Google Scholar] [CrossRef] [Green Version]
- Han, E.; Ho Oh, K.; Park, S.; Chan Rah, Y.; Park, H.C.; Koun, S.; Choi, J. Analysis of behavioral changes in zebrafish (Danio rerio) larvae caused by aminoglycoside-induced damage to the lateral line and muscles. Neurotoxicology 2020, 78, 134–142. [Google Scholar] [CrossRef]
- Broderick, M.T.; Prince, A.D.P.; Dhukhwa, A.; Mukherjea, D.; Jiang, P.; Campbell, K.C.M.; Rybak, L.P.; Brenner, M.J. Effects of Moringa Extract on Aminoglycoside-Induced Hair Cell Death and Organ of Corti Damage. Otol. Neurotol. 2021, 42, 1261–1268. [Google Scholar] [CrossRef]
- Liu, Q.; Hu, Y.; Cao, Y.; Song, G.; Liu, Z.; Liu, X. Chicoric Acid Ameliorates Lipopolysaccharide-Induced Oxidative Stress via Promoting the Keap1/Nrf2 Transcriptional Signaling Pathway in BV-2 Microglial Cells and Mouse Brain. J. Agric. Food Chem. 2017, 65, 338–347. [Google Scholar] [CrossRef]
- Hussein, O.E.; Hozayen, W.G.; Bin-Jumah, M.N.; Germoush, M.O.; Abd El-Twab, S.M.; Mahmoud, A.M. Chicoric acid prevents methotrexate hepatotoxicity via attenuation of oxidative stress and inflammation and up-regulation of PPARgamma and Nrf2/HO-1 signaling. Environ. Sci. Pollut. Res. Int. 2020, 27, 20725–20735. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xie, G.; Liu, Q.; Duan, X.; Liu, Z.; Liu, X. Pharmacokinetics, tissue distribution, and plasma protein binding study of chicoric acid by HPLC-MS/MS. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2016, 1031, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Simon, H.U.; Haj-Yehia, A.; Levi-Schaffer, F. Role of reactive oxygen species (ROS) in apoptosis induction. Apoptosis 2000, 5, 415–418. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, K.; Lei, Y.; Li, Q.; Nice, E.C.; Huang, C. Redox signaling: Potential arbitrator of autophagy and apoptosis in therapeutic response. Free Radic. Biol. Med. 2015, 89, 452–465. [Google Scholar] [CrossRef] [PubMed]
- Huth, M.E.; Ricci, A.J.; Cheng, A.G. Mechanisms of aminoglycoside ototoxicity and targets of hair cell protection. Int. J. Otolaryngol. 2011, 2011, 937861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, M.; Karasawa, T.; Steyger, P.S. Aminoglycoside-Induced Cochleotoxicity: A Review. Front. Cell Neurosci. 2017, 11, 308. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Liu, H.; Huang, Z.; Yang, R.; Gong, L. The Ameliorative Effect of JNK Inhibitor D-JNKI-1 on Neomycin-Induced Apoptosis in HEI-OC1 Cells. Front. Mol. Neurosci. 2022, 15, 824762. [Google Scholar] [CrossRef]
- Sahu, B.D.; Tatireddy, S.; Koneru, M.; Borkar, R.M.; Kumar, J.M.; Kuncha, M.; Srinivas, R.; Shyam Sunder, R.; Sistla, R. Naringin ameliorates gentamicin-induced nephrotoxicity and associated mitochondrial dysfunction, apoptosis and inflammation in rats: Possible mechanism of nephroprotection. Toxicol. Appl. Pharmacol. 2014, 277, 8–20. [Google Scholar] [CrossRef]
- Manner, T.; Kanto, J.; Scheinin, H.; Scheinin, M. Meptazinol and pentazocine: Plasma catecholamines and other effects in healthy volunteers. Br. J. Clin. Pharmacol. 1987, 24, 689–697. [Google Scholar] [CrossRef]
- Kenyon, E.J.; Kirkwood, N.K.; Kitcher, S.R.; Goodyear, R.J.; Derudas, M.; Cantillon, D.M.; Baxendale, S.; de la Vega de Leon, A.; Mahieu, V.N.; Osgood, R.T.; et al. Identification of a series of hair-cell MET channel blockers that protect against aminoglycoside-induced ototoxicity. JCI Insight 2021, 6, 7. [Google Scholar] [CrossRef]
- Kitcher, S.R.; Kirkwood, N.K.; Camci, E.D.; Wu, P.; Gibson, R.M.; Redila, V.A.; Simon, J.A.; Rubel, E.W.; Raible, D.W.; Richardson, G.P.; et al. ORC-13661 protects sensory hair cells from aminoglycoside and cisplatin ototoxicity. JCI Insight 2019, 4, 15. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros, A.; Swartz, K.J. Dextran Labeling and Uptake in Live and Functional Murine Cochlear Hair Cells. J. Vis. Exp. 2020, 156, e60769. [Google Scholar] [CrossRef] [PubMed]
- Sarasamma, S.; Audira, G.; Siregar, P.; Malhotra, N.; Lai, Y.H.; Liang, S.T.; Chen, J.R.; Chen, K.H.; Hsiao, C.D. Nanoplastics Cause Neurobehavioral Impairments, Reproductive and Oxidative Damages, and Biomarker Responses in Zebrafish: Throwing up Alarms of Wide Spread Health Risk of Exposure. Int. J. Mol. Sci. 2020, 21, 1410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todd, D.W.; Philip, R.C.; Niihori, M.; Ringle, R.A.; Coyle, K.R.; Zehri, S.F.; Zabala, L.; Mudery, J.A.; Francis, R.H.; Rodriguez, J.J.; et al. A Fully Automated High-Throughput Zebrafish Behavioral Ototoxicity Assay. Zebrafish 2017, 14, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Benvenutti, R.; Marcon, M.; Reis, C.G.; Nery, L.R.; Miguel, C.; Herrmann, A.P.; Vianna, M.R.M.; Piato, A. N-acetylcysteine protects against motor, optomotor and morphological deficits induced by 6-OHDA in zebrafish larvae. PeerJ 2018, 6, e4957. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lai, T.-W.; Cheng, H.-L.; Su, T.-R.; Yang, J.-J.; Su, C.-C. Cichoric Acid May Play a Role in Protecting Hair Cells from Ototoxic Drugs. Int. J. Mol. Sci. 2022, 23, 6701. https://doi.org/10.3390/ijms23126701
Lai T-W, Cheng H-L, Su T-R, Yang J-J, Su C-C. Cichoric Acid May Play a Role in Protecting Hair Cells from Ototoxic Drugs. International Journal of Molecular Sciences. 2022; 23(12):6701. https://doi.org/10.3390/ijms23126701
Chicago/Turabian StyleLai, Ting-Wei, Hsin-Lin Cheng, Tzu-Rong Su, Jiann-Jou Yang, and Ching-Chyuan Su. 2022. "Cichoric Acid May Play a Role in Protecting Hair Cells from Ototoxic Drugs" International Journal of Molecular Sciences 23, no. 12: 6701. https://doi.org/10.3390/ijms23126701
APA StyleLai, T.-W., Cheng, H.-L., Su, T.-R., Yang, J.-J., & Su, C.-C. (2022). Cichoric Acid May Play a Role in Protecting Hair Cells from Ototoxic Drugs. International Journal of Molecular Sciences, 23(12), 6701. https://doi.org/10.3390/ijms23126701