Abstract
The rhizospheric bacterium Pseudomonas protegens Pf-5 can colonize the seed and root surfaces of plants, and can protect them from pathogen infection. Secondary metabolites, including lipopeptides and polyketides produced by Pf-5, are involved in its biocontrol activity. We isolated a crude extract from Pf-5. It exhibited significant surface activity and strong antibacterial activity against Pantoea ananatis DZ-12, which causes maize brown rot on leaves. HPLC analysis combined with activity tests showed that the polyketide pyoluteorin in the crude extract participated in the suppression of DZ-12 growth, and that the lipopeptide orfamide A was the major biosurfactant in the crude extract. Further studies indicated that the pyoluteorin in the crude extract significantly suppressed the biofilm formation of DZ-12, and it induced the accumulation of reactive oxygen species in DZ-12 cells. Scanning electron microscopy and transmission electron microscopy observation revealed that the crude extract severely damaged the pathogen cells and caused cytoplasmic extravasations and hollowing of the cells. The pathogenicity of DZ-12 on maize leaves was significantly reduced by the crude extract from Pf-5 in a dose-dependent manner. The polyketide pyoluteorin had strong antibacterial activity against DZ-12, and it has the potential for development as an antimicrobial agent.
1. Introduction
The phytopathogenic Pantoea ananatis is a Gram-negative bacterial species that causes severe economic losses in onion, maize, pineapple, and honeydew melon [1,2]. P. ananatis can also cause brown rot in maize leaves and cause economic losses in maize production [2]. The disease, termed maize brown rot leaves, produces water-soaked lesions on the leaves, which eventually become necrotic and straw-colored. The disease can be disseminated over the entire leaf surface in its advanced stage [3]. This disease also causes premature leaf drying, shortening of the growth cycle, and the reduction of kernel length and weight [4].
The plant rhizosphere is the narrow zone of soil that is relatively rich in nutrients due to the accumulation of a variety of plant exudates [5]. The rhizosphere harbors beneficial bacteria named plant growth-promoting rhizobacteria (PGPR). These colonize the rhizosphere, root surfaces, and radicular tissues. They promote plant growth and suppress soil-borne plant pathogens [6]. These beneficial biological activities have led to the development of many PGPR strains as commercial biological control agents (BCAs). Among these PGPR strains, Pseudomonas species have received significant attention [7].
To survive and to successfully compete with other microorganisms, PGPR can effectively utilize seed and root exudates to produce a wide spectrum of bioactive metabolites such as antibiotics, siderophores, volatile organic compounds (VOCs), and growth-promoting substances [8]. The Pseudomonas biocontrol strains also can induce systemic resistance in plants [9]. Several Pseudomonas strains, such as Pseudomonas protegens Pf-5, are effective at controlling plant diseases caused by Pythium ultimum (damping-off of cotton) and Rhizoctonia solani [10]. The P. protegens CHA0 strain suppresses root rots of tobacco and tomato caused by Pythium sp., as well as the damping-off of cucumber and take-all of wheat [11]. In addition, Pseudomonas fluorescens 2P24 can protect plants against tomato bacterial wilt caused by Ralstonia solanacearum [12].
Among the Pseudomonas biocontrol strains, the ability to produce antibiotics is an effective method for controlling plant diseases. Phenazine-1-carboxylic acid (PCA) produced by Pseudomonas species provides the effective suppression of pathogens [13], such as Gaeumannomyces graminis which causes take-all disease of wheat [14]. PCA is the main active component of the benzimidazole-derived biopesticide “Shenqinmycin” [15]. The polyketide antibiotic 2,4-diacetylphloroglucinol (2,4-DAPG) is a phloroglucinol derivative compound from some Pseudomonas species that exhibits broad-spectrum antimicrobial activity [16] and suppresses black root rot of tobacco [8]. Other metabolites, such as pyoluteorin (Plt), pyrrolnitrin (Prn), siderophores, and lipopeptides are active substrates that are produced by Pseudomonas biocontrol agents and are reported as being associated with biocontrol activity [17,18].
The Pseudomonas commensal, P. protegens Pf-5, is notable as a BCA, for its efficient rhizosphere colonization and the ability to produce many secondary metabolites [19]. Pf-5 can produce the lipopeptides orfamide A, rhizoxin, pyrrolnitrin, pyoluteorin, 2,4-DAPG, and the siderophores pyochelin and pyoverdine [18,19,20]. These secondary metabolites contribute to the suppression of plant diseases. Pf-5 can also produce the VOC hydrogen cyanide (HCN), which is toxic to phytopathogens. In the present study, we explored the mechanism underlying the antibacterial activity of a crude extract prepared from Pf-5 against P. ananatis DZ-12.
2. Results
2.1. Antibacterial and Surface Activity of Crude Extract Prepared from P. protegens Pf-5
The bio-surfactant activity of crude extract prepared from the supernatant of P. protegens Pf-5 was measured and expelling oil was performed according to the previously reported method [21]. The crude extract showed significant surfactant activity (Figure 1A), indicating that the surfactant compound was present in the crude extract. The crude extract also exhibited antibacterial activity against P. ananatis DZ-12 (Figure 1B).

Figure 1.
Analysis of the bio-surfactant activity prepared from P. protegens Pf-5. (A) Antibacterial activity; (B) Crude extract from P. protegens Pf-5. The crude extract was directly used.
2.2. Pyoluteorin and Orfamide A produced by P. protegens Pf-5 Showed In Vitro Antibacterial and Surface Activity, Respectively
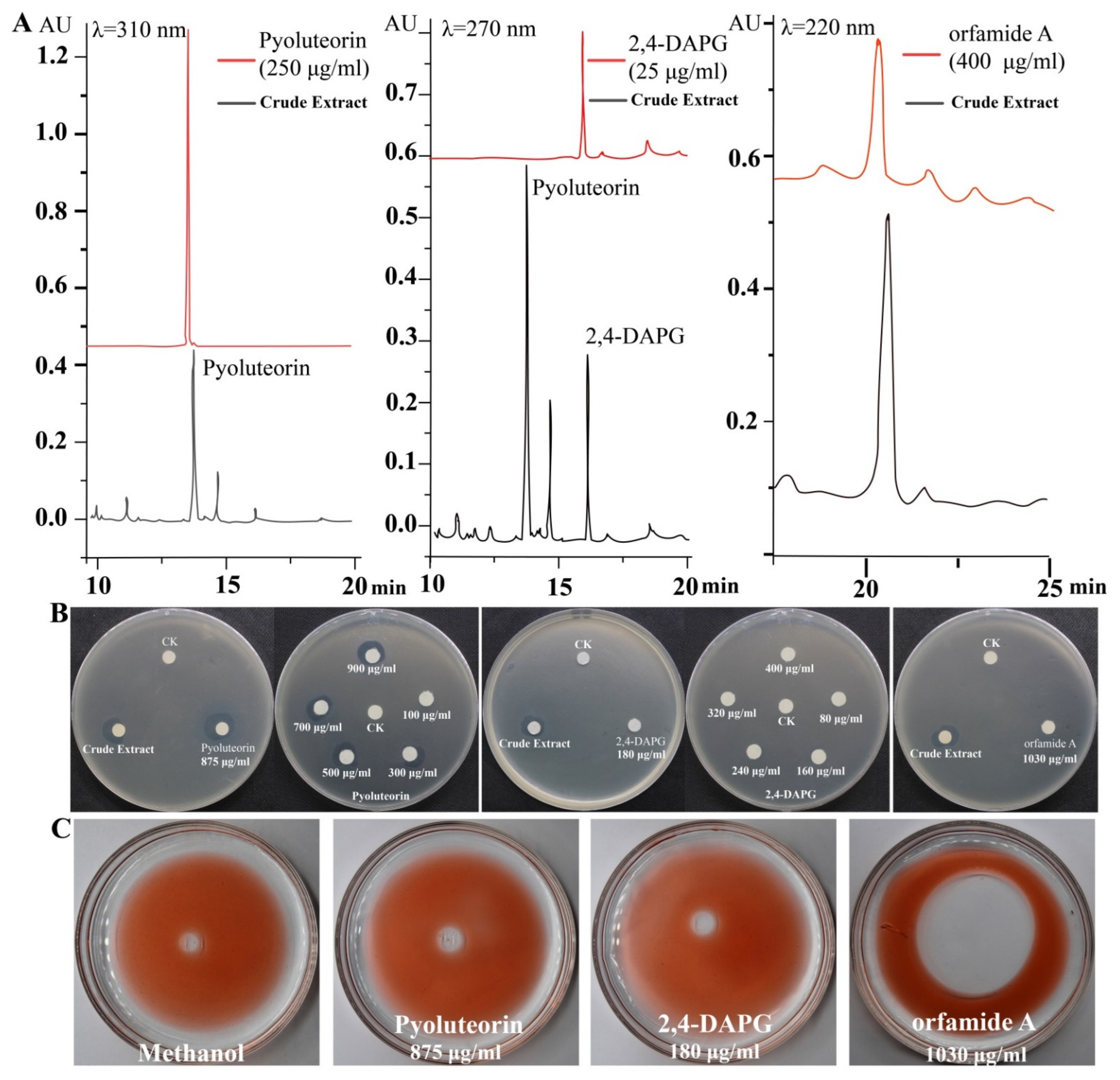
We identified the active compounds in the crude extract using HPLC analysis. Three different standard compounds, pyoluteorin, 2,4-DAPG, and orfamide A, were used as references. We found that these three compounds were present in the crude extract (Figure 2A). Standard curves were used to determine that the concentrations of pyoluteorin, 2,4-DAPG, and orfamide A in crude extract were 875 μg/mL, 180 μg/mL, and 1030 μg/mL, respectively. Pyoluteorin showed strong antibacterial activity among the selected compounds being treated with the same concentration (875 μg/mL) against P. ananatis DZ-12. However, 180 μg/mL 2,4-DAPG and 1030 μg/mL orfamide A had no significant antibacterial activity (Figure 2B). Based on these results, various doses of pyoluteorin and 2,4-DAPG were selected for further experiments. Pyoluteorin had antibacterial activity that was enhanced with increasing dose (100 to 900 μg/mL). The 2,4-DAPG had lower antibacterial activity; even at 400 μg/mL, inhibition zones were not observed (Figure 2B). To verify these results, seven mutants of Pf-5 were tested. The results showed that only mutant ΔpltB that cannot synthesize pyoluteorin loses its ability to suppress the growth of DZ-12 (Figure S1). These results showed that pyoluteorin was the main antibacterial compound in the crude extract. In previous findings, the crude extract also showed biosurfactant activity. We performed the same experiments and found that these three major compounds showed such activity. Orfamide A can produce expelling oil with surfactant activity. However, 2,4-DAPG and pyoluteorin did not have this activity (Figure 2C). These results demonstrate that orfamide A is the major biosurfactant in the crude extract.
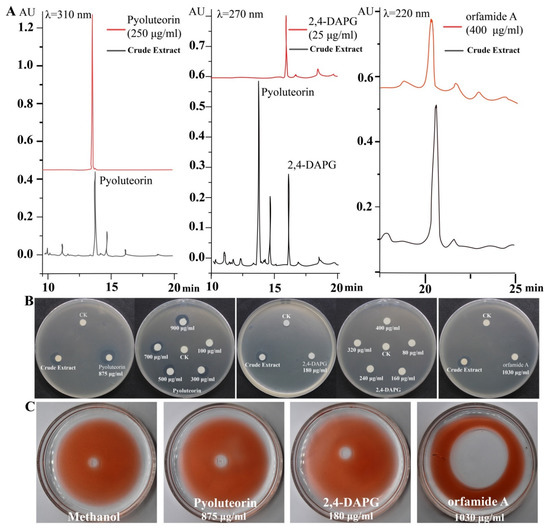
Figure 2.
HPLC analysis of the major components in the crude extract (A) and detection of their antibacterial activity (B) and biosurfactant activity (C).
2.3. Pyoluteorin Produced by P. protegens Pf-5 Contributes to the Inhibitory Effect on Biofilm Formation of P. ananatis DZ-12
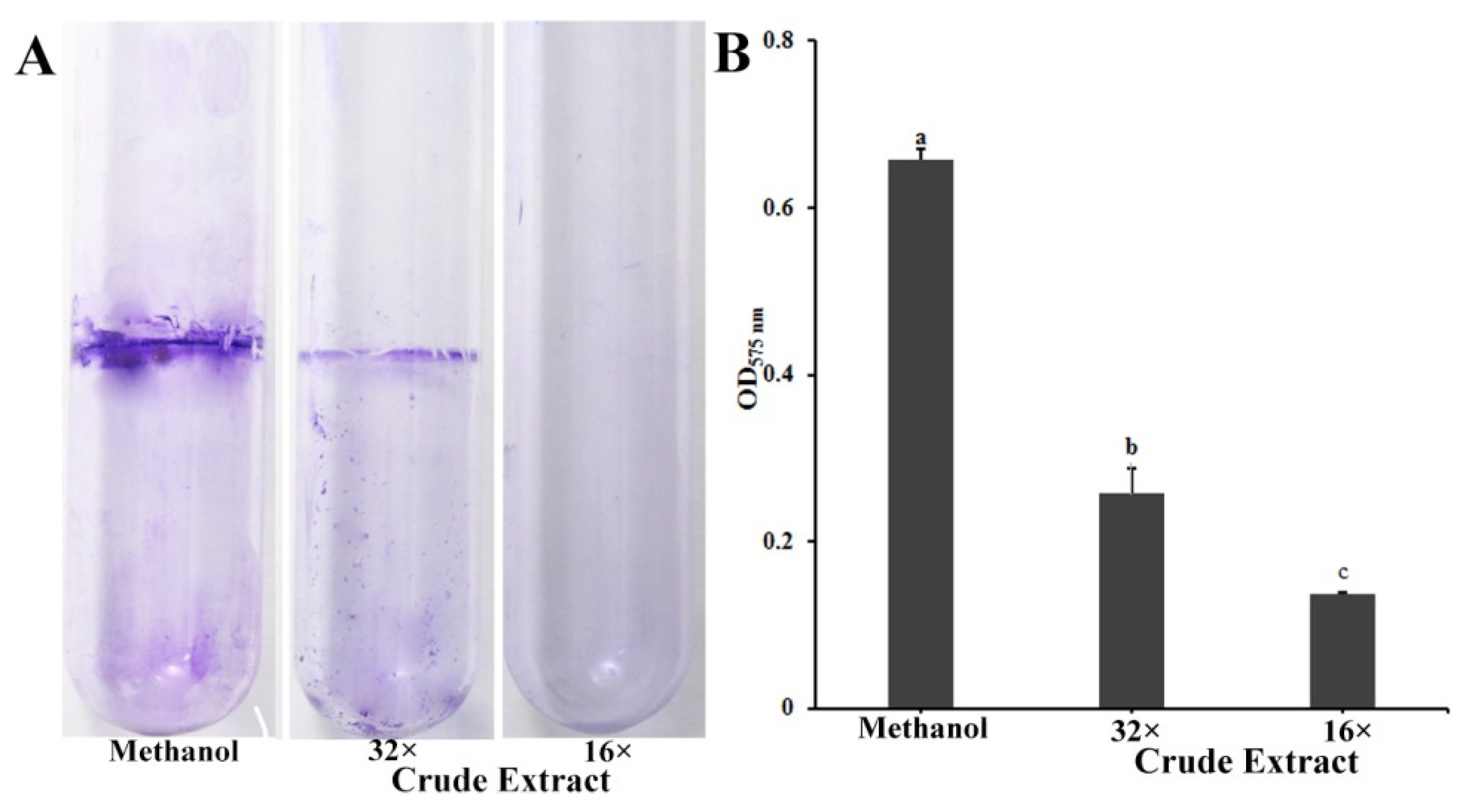
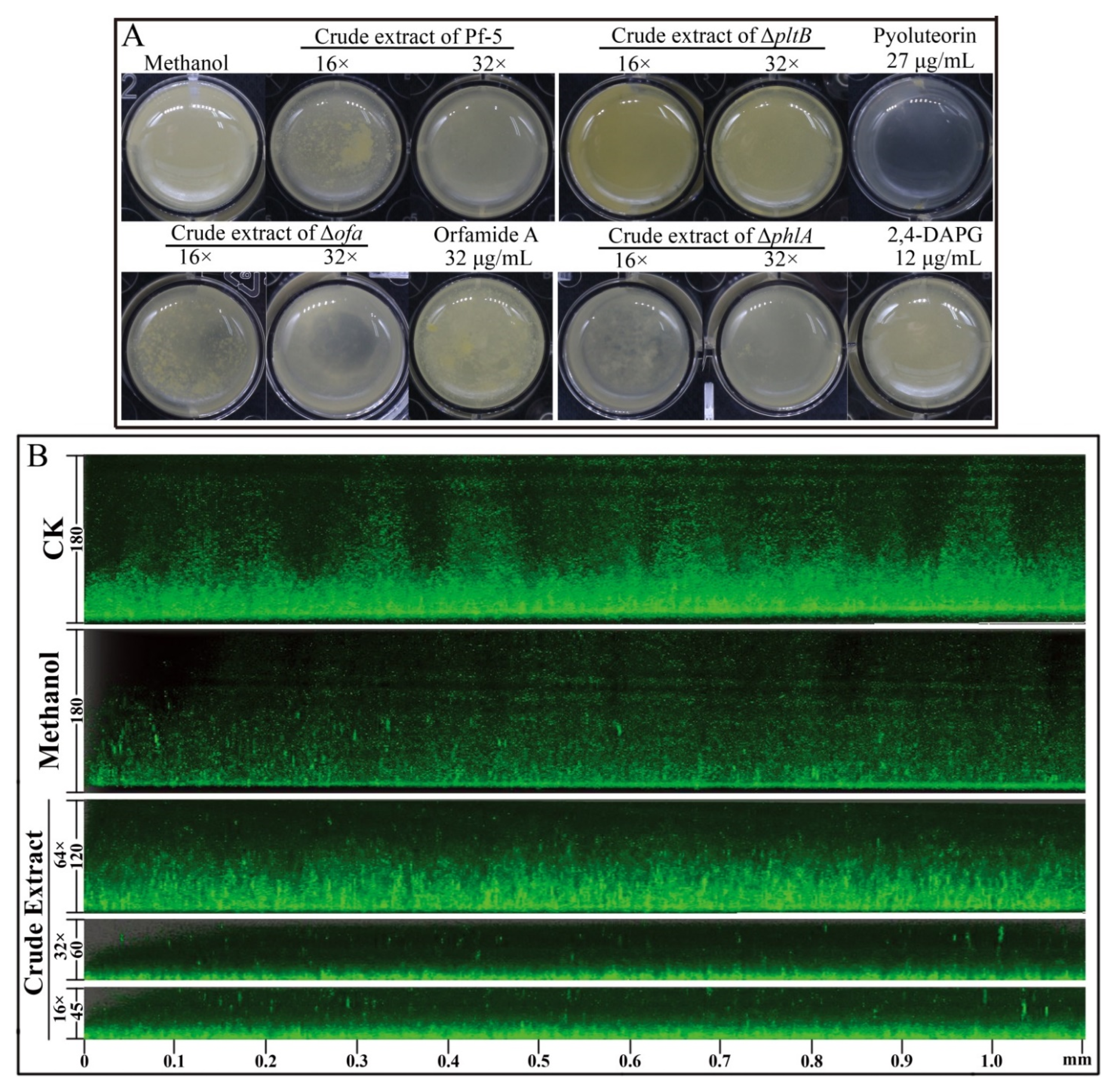
Biofilms have a major role in plants colonization and are virulence determinants in many bacterial pathogens [22]. Because crude extract inhibits the biofilm formation of P. ananatis DZ-12, the biofilm was measured using crystal violet staining. Dilutions of 16× and 32× of the crude extract had a significant inhibitory effect on the biofilm formation of P. ananatis DZ-12 (Figure 3).
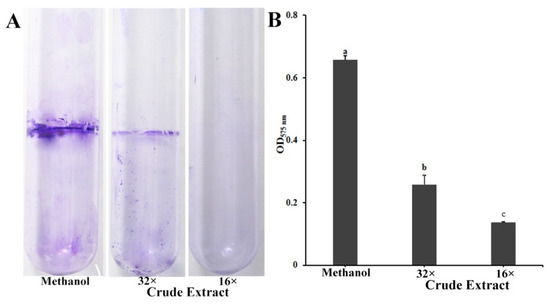
Figure 3.
Effect of crude extract prepared from P. protegens Pf-5 on the biofilm formation of P. ananatis DZ-12. (A) Biofilm on the inner surface of glass tubes were stained with crystal violet and photographed; (B) The graph data show the average OD575 nm value ± standard deviation (SD) of three replicates. A concentration of 32× and 16× dilutions of the crude extract from P. protegens Pf-5 were used. Different letters represent statistically significant differences according to the one-way ANOVA test (p < 0.05).
In order to determine which compounds play a key role in the suppression of the biofilm of DZ-12, three mutants, Δofa, ΔpltB, and ΔphlA which have respectively lost the ability to synthesize orfamide A, pyoluteorin, and 2,4-DAPG, were used. Observations of P. ananatis DZ-12 biofilm in a different dilution of the crude extract from wild type Pf-5 and the two mutants Δofa and ΔphlA all showed suppression of biofilm formation. Their crude extract reduced the growth of bacterial cells and also disrupted the thickness of biofilm structure compared to the control methanol treatment (Figure 4A). However, mutant ΔpltB, which cannot synthesize pyoluteorin, has lost this activity. To confirm this phenomenon, three pure commercial standards orfamide A, pyoluteorin, and 2,4-DAPG were used. The results had showed only pyoluteorin could suppress the biofilm formation of DZ-12. This indicated that the pyoluteorin present in the crude extract was the key factor for antibiofilm activity.
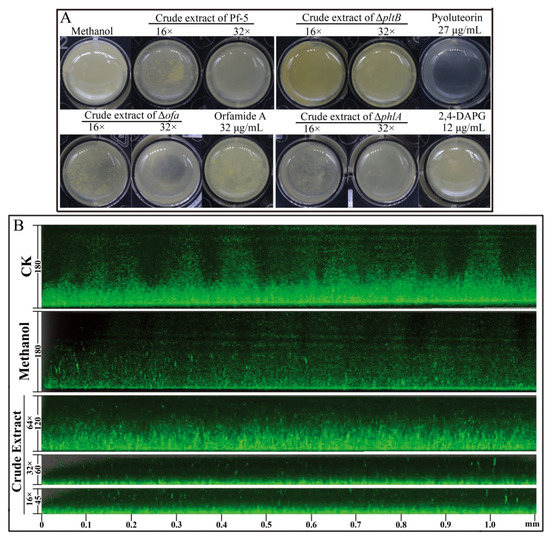
Figure 4.
Effect of crude extract prepared from P. protegens Pf-5 on the biofilm formation of P. ananatis DZ-12. Bacterial cells were statistically cultured on LB medium in a 24-well plate (A), and in chambers in slides (B), at 28 °C for 48 h. For the common observation (A), different dilutions of the crude extract (32×, 16×) from P. protegens Pf-5 and its mutants Δofa, ΔpltB, and ΔphlA were used. A concentration of 27 μg/mL pyoluteorin, 32 μg/mL orfamide A, and 12 μg/mL 2,4-DAPG were also used. For the fluorescence microscope observation (B), different dilutions of the crude extract (64×, 32×, and 16×) from P. protegens Pf-5 were used. Representative images of the Z-stack assembly in the biofilms of cells and biofilm thicknesses were measured using a Leica SP5 confocal microscope.
We also used P. ananatis DZ-12 labeled with the gfp gene to observe biofilm formation in detail. A similar inhibitory effect was seen under a fluorescence microscope. The thickness of biofilm was significantly reduced more than three times when P. ananatis DZ-12 was treated with a 16× dilution of crude extract (Figure 4B). These results further strengthen the evidence that the crude extract can inhibit the growth of P. ananatis DZ-12 and suppress biofilm formation.
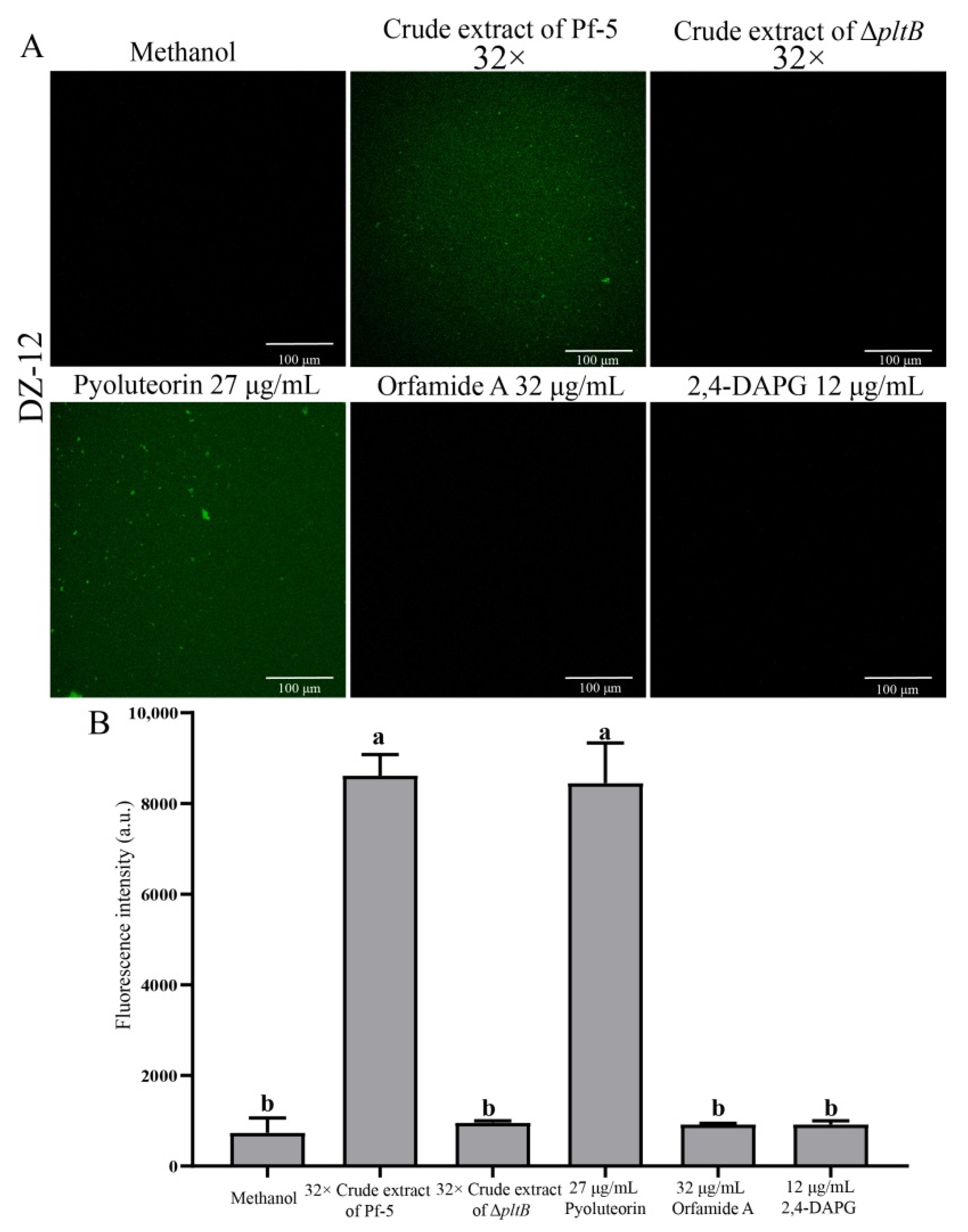
2.4. Induction of Reactive Oxygen Species in P. ananatis DZ-12 Exposed to Crude Extract Containing Pyoluteorin
High concentrations of reactive oxygen species (ROS) production are harmful to cells and can result in cell death [23]. Our results showed that crude extract had strong antibacterial activity and induced ROS in P. ananatis DZ-12. Green fluorescence microscopy observation revealed that the bacterial cells had strong green fluorescence after treatment with 32 × dilutions of crude extract from Pf-5 compared to the control (Figure 5A). A similar result was also observed when cells of DZ-12 were treated with 27 μg/mL pyoluteorin (Figure 5A). However, a crude extract from the ΔpltB mutant, as well as orfamide A and 2,4-DAPG, did not result in the ROS accumulation of DZ-12 (Figure 5A). The fluorescence intensity confirmed that the ROS accumulation level of DZ-12, when treated with 32× dilutions of crude extract from Pf-5 and 27 μg/mL pyoluteorin, was significantly higher than that of the control, ΔpltB, and two compounds, orfamide A and 2,4-DAPG (Figure 5B). These results indicated that the pyoluteorin presenting in the crude extract contributes to the ROS generation of DZ-12.
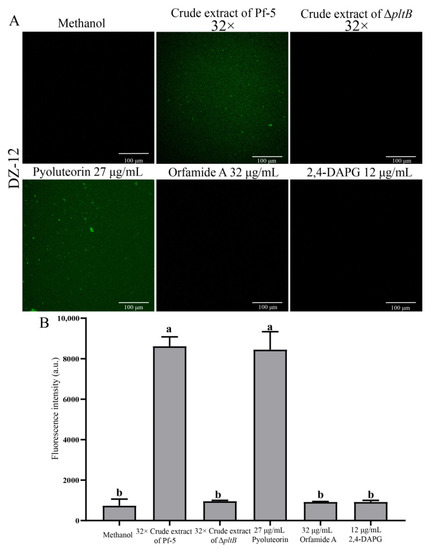
Figure 5.
The effects of crude extract on ROS production P. ananatis DZ-12 observed using a Leica SP5 confocal microscope (A), and analyzed by EnSight Multimode Plate Reader (B). A concentration of 32× dilutions of the crude extract from P. protegens Pf-5 and its mutant ΔpltB were used. A concentration of 27 μg/mL pyoluteorin, 32 μg/mL orfamide A, and 12 μg/mL 2,4-DAPG were also used. The graph data show the average fluorescence intensity value ± standard deviation of three replicates. Different letters represent statistically significant differences according to the one-way ANOVA test (p < 0.05).
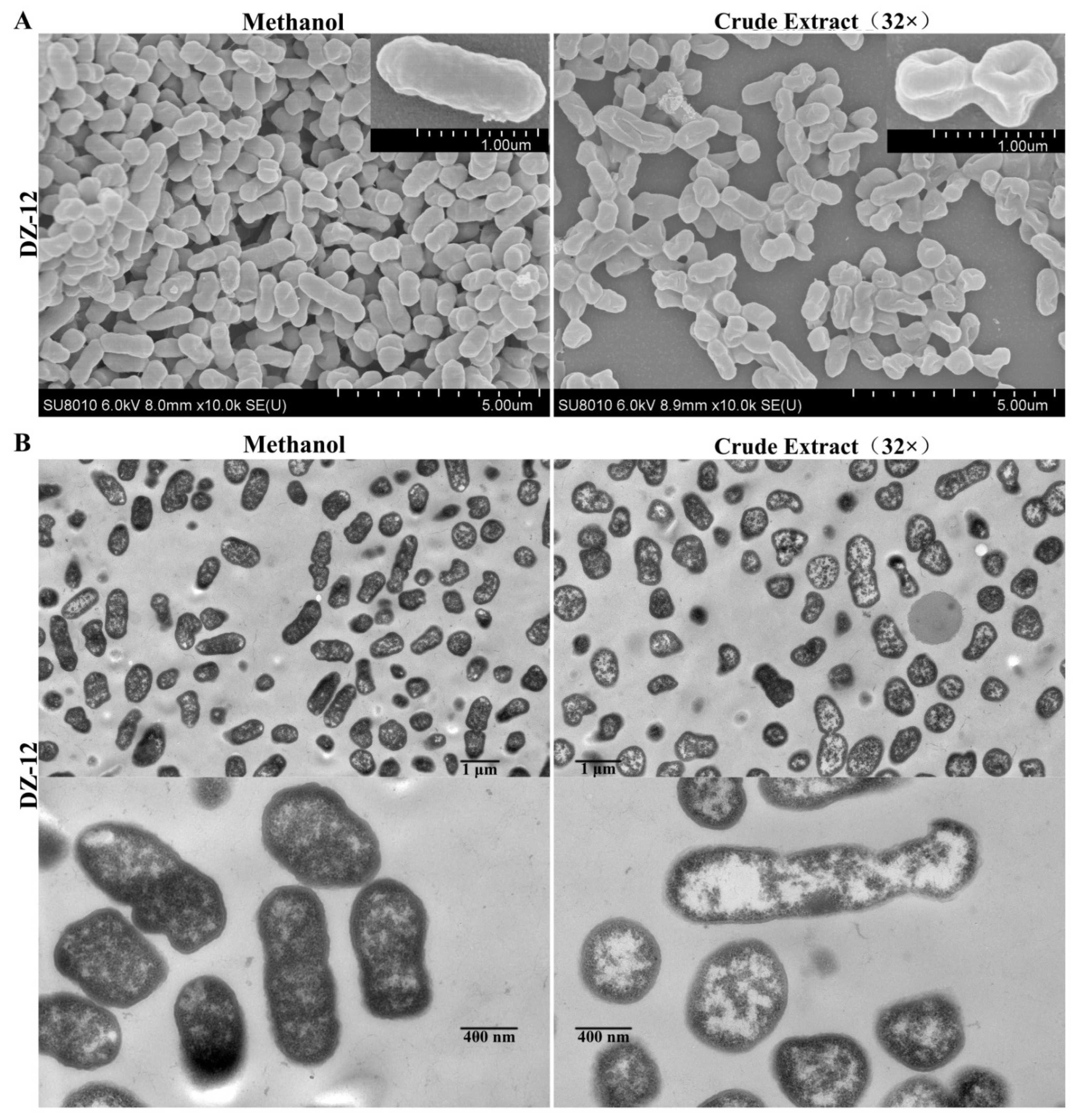
2.5. Ultrastructural Changes in P. ananatis DZ-12 Caused by Crude Extract
From the results, pyoluteorin presenting in crude extract contributes to anti-biofilm and ROS generation activities. Thus, the crude extract was directly used in the following experiments. Scanning electron microscopy (SEM) and transmission electron microscopy (TEM) were used to observe the ultrastructural and morphological changes in P. ananatis DZ-12 cells under the treatment of crude extract. The SEM results showed that control methanol-treated P. ananatis DZ-12 cells had a normal shape, complete and fully cylindrical. In contrast, after being treated with a 32 × dilution of crude extract, cells were depressed and shriveled (Figure 6A). These changes may be due to cytoplasm leakage. TEM was used to confirm these observations. In the methanol control, the structure of the bacterial cell was complete, with a uniform cytoplasmic structure. In contrast, the cell walls and cell membranes of the cells treated with crude extract were severely damaged, causing cytoplasmic extravasation and hollowing (Figure 6B). These results indicate that there were active substances in the crude extract that could damage the cell wall and plasma membrane and cause cytoplasm leakage.
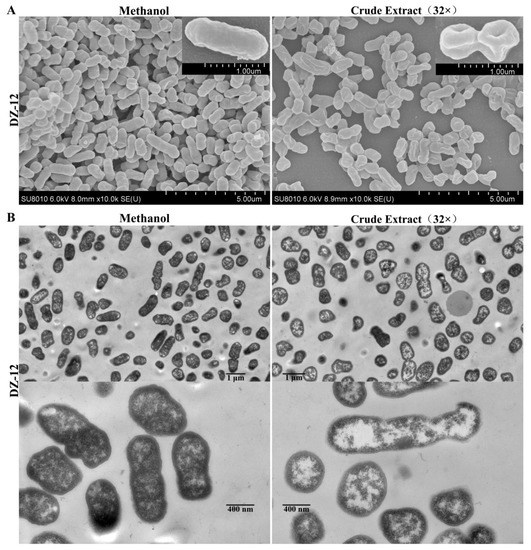
Figure 6.
Effect of crude extract on the micro- and ultra-structures of the P. ananatis DZ-12 cells, as determined by SEM (A) and TEM (B).
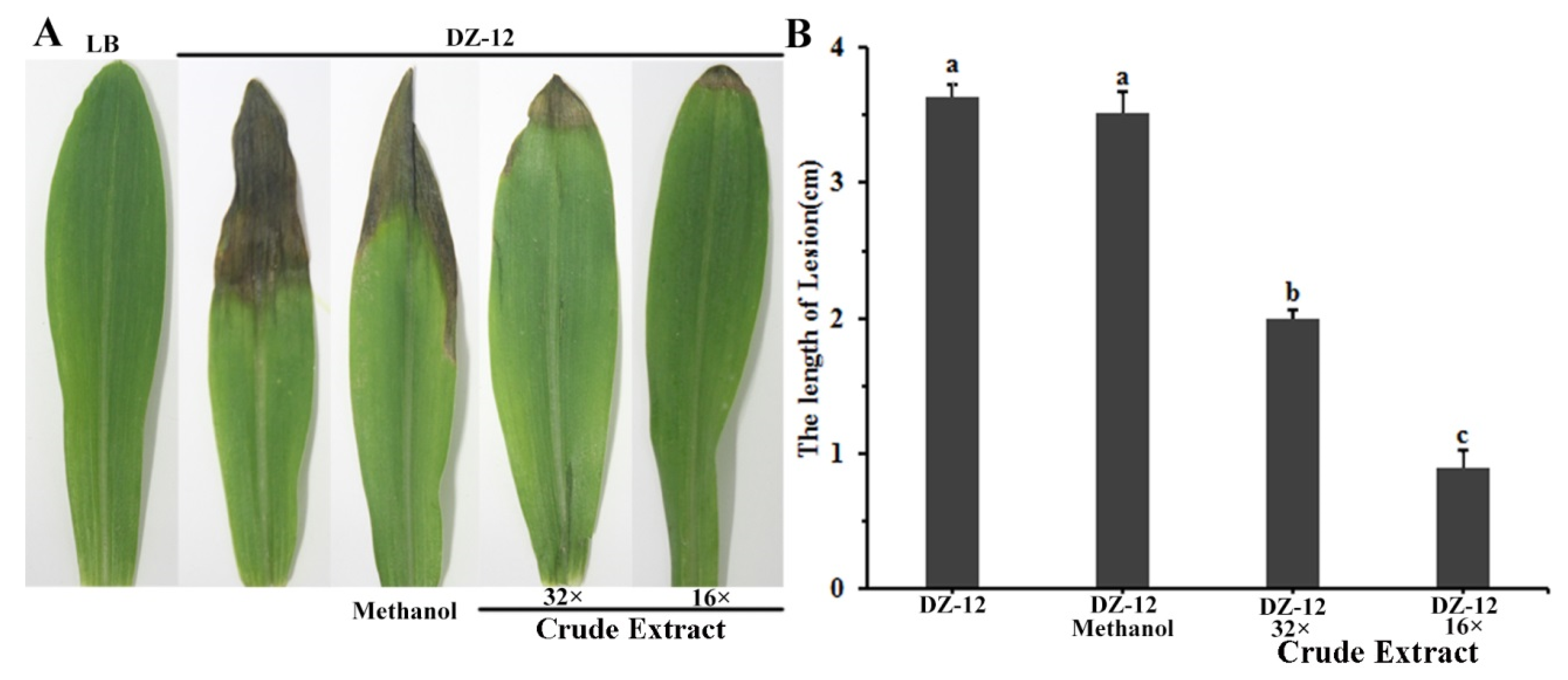
2.6. Crude Extract Suppresses the Virulence of Brown Rot of Maize Leaves Caused by P. ananatis DZ-12
To study the effect of crude extract on DZ-12 strain pathogenicity on maize seedling leaves, a plant infection experiment was conducted. Different dilutions of crude extract provided significant reductions in the lengths of yellow-brown lesions in maize leaves caused by P. ananatis DZ-12. The effect was enhanced with increasing dose concentration (Figure 7). The average lengths of the leaf lesions were 2 cm and 1.0 cm in leaves treated with 32× and 16× dilutions of the crude extract, which were, respectively, 1/2 and 1/4 of the control treatment. The results showed that the crude extract from P. protegens Pf-5 reduced the pathogenic effects of P. ananatis DZ-12 in maize leaves.
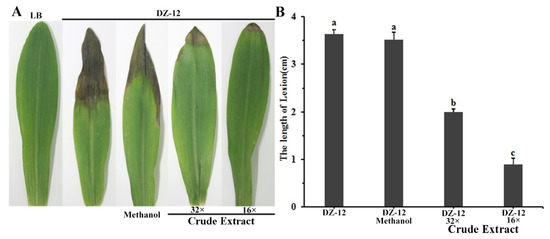
Figure 7.
Crude extract from P. protegens Pf-5 influenced the infection of P. ananatis DZ-12 on maize leaves. (A) Disease symptom of maize plants inoculated with the DZ-12 treated with different dilutions of crude extract. Pictures were taken 7 days post-inoculation (dpi) and indicate representative results; (B) The graph data show the average lengths of leaf lesions ± standard deviation of three replicates. A concentration of 32× and 16× dilutions of the crude extract from P. protegens Pf-5 were used. Different letters represent statistically significant differences according to the one-way ANOVA test (p < 0.05).
3. Discussion
P. ananatis causes disease in many important crop plants, including maize, onion, honeydew melon, and pineapple [2,24]. We previously isolated strain DZ-12 from the rotted leaves of maize, and genomic analysis revealed that DZ-12 belongs to P. ananatis [2]. In this study, we found that pyoluteorin that is present in the crude extract of P. protegens Pf-5 can suppress the growth of P. ananatis DZ-12 (Figure 1 and Figure 2). Pyoluteorin is composed of a resorcinol ring, biosynthesized by polyketide synthase/nonribosomal peptide synthase (PKS/NRPS) hybrid complexes involving the sequential addition and modification of simple carboxyl acids to a growing carbon chain [19,25]. A similar study found that pyoluteorin inhibits Oomycetes, including Pythium ultimum, and contributes to the biocontrol of Pythium damping-off in cotton [26]. Pyoluteorin also has anti-tumor activity, which can induce cell cycle arrest and apoptosis in human triple-negative breast cancer cells [27,28]. Recently, pyoluteorin was reported to have activity against Chlamydomonas reinhardtii and Heterobasidion species [29,30]. All of these results indicate that pyoluteorin has a broad spectrum of antimicrobial activity.
The biofilm of P. ananatis DZ-12 serves as a major determinant in its pathogenesis [2]. We found that the biofilm of P. ananatis DZ-12 was significantly inhibited by a crude extract from P. protegens Pf-5 (Figure 3 and Figure 4), and this was confirmed by results from confocal laser scanning microscopy (Figure 4). The further study indicates that pyoluteorin presenting in crude extract contributes to inhibitory effects on the biofilm formation of DZ-12 (Figure 4). The decreased pathogenicity of the cured strain may also be related to its impairment in biofilm formation. The inhibition or prevention of biofilm formation represents an emerging strategy in the war against antibiotic resistance, interfering with key players in bacterial virulence [31]. Pyoluteorin has a pyrrole ring and it belongs to nitrogenous heterocyclic compounds. Several compounds having a similar structure were reported for interesting anti-biofilm properties [32,33].
The crude extract of P. protegens Pf-5 induced a high ROS accumulation in DZ-12 cells (Figure 5). Other studies have found that low ROS concentrations act as intracellular messengers for molecular events, but that large amounts of ROS cause cell death [23]. The results also showed that the existence of pyoluteorin in the crude extract resulted in the ROS generation of DZ-12 (Figure 5). Pyoluteorin was reported to induce the generation of intracellular ROS in human cancer cells [28]. Other research showed that pyoluteorin induced cytosolic Ca2+ fluxes in C. reinhardtii and inhibited its growth [29]. In our previous study, we found that the lipopeptide bacillomycin D produced by Bacillus amyloliquefaciens could induce the ROS generation of F. graminearum [34].
Ultrastructural alterations in the morphologies of DZ-12 were observed using SEM and TEM to investigate the activities of crude extracts. SEM and TEM showed that untreated P. ananatis cells were in a normal state, complete with a cylindrical trunk (Figure 6). However, the cells treated with crude extract were depressed and shriveled. This may have been caused by the pyoluteorin present in the crude extract. Recently, a similar phenomenon was also reported. Pyoluteorin could cause a variety of morphological changes in C. reinhardtii, such as uniflagellar structures [29]. In human cancer cells, pyoluteorin induces cell cycle arrest, possibly via the reduction of the mitochondrial membrane [28].
Lipopeptide compounds are essential in the biocontrol of Pseudomonas BCA in plant pathogens. They may be useful for managing plant pathogen infestations because they are selectively nontoxic, stable, and environmentally benign [35]. Our study also showed that orfamide A had bio-surfactant activity (Figure 2). The bio-surfactant activity of cyclic lipopeptides (CLPs) has been reported to contribute to the swarming motility of Pseudomonas strains, and of biofilm formation [36] and colonization [37]. Orfamides are CLPs that consist of a saturated or unsaturated unbranched β-hydroxy fatty chain (12, 14, or 16 carbons in length) bound to a decapeptide, with eight of the amino acids forming a lactone ring structure [20]. Orfamide can inhibit the growth of the green algae, C. reinhardtii [38]. Orfamide can also act as an insecticidal metabolite against the aphid Myzus persicae [39]. However, in this study, orfamide A had no effect on the growth of DZ-12. This is in view of the fact that many pesticides also contain surfactants that increase the solubility of active substances and increase their surface adherence. Thus, the crude extract containing the bio-surfactant orfamide was directly used.
In conclusion, we demonstrated that the crude extract from P. protegens Pf-5 had strong antibacterial activity against the pathogen P. ananatis DZ-12 and exhibited significant surface activity. HPLC analysis showed that the polyketide pyoluteorin is present in the crude extract, and it plays an important role in the growth suppression of DZ-12, in vivo and in vitro. This compound has the potential to be developed as a pathogen control agent.
4. Materials and Methods
4.1. Bacterial Strains and Growth Conditions
The strain P. protegens Pf-5 and its mutants were cultured at 30 °C on King’s B (KB) medium B agar. The gfp-labeled strain P. ananatis DZ-12 was grown at 30 °C on Luria Bertani (LB) medium. Antibiotics ampicillin (100 μg/mL) and chloramphenicol (25 μg/mL) were added into the liquid medium or solid agar plate medium, when required. All of the strains used in this study are listed in Table S1.
4.2. Preparation of Crude Extraction from P. protegens Pf-5
The crude extraction from Pf-5 or its mutants was prepared according to the methods described by Hassan et al. [40] and Oni et al. [41] with some changes. Briefly, a single colony of Pf-5 or its mutants was inoculated into 10 mL of KB medium in a 50 mL conical flask, and cultured at 30 °C, 200 rpm for 24 h. Then, 2 mL of this culture was transferred to 200 mL KB medium in a 500 mL flask, followed by incubation for 48 h at 30 °C. The supernatant was collected by centrifugation at 10,000 rpm for 25 min at 4 °C, and the pH was maintained at 2.0 using HCl. The supernatant was stored at 4 °C overnight. The precipitates were collected by centrifugation at 4 °C for 10 min at 12,000× g. The collected precipitate was completely suspended by using an appropriate amount of methanol. Then, the pH of the precipitate was adjusted to 7.0 with NaOH, and allowed to stand for 12 h. Finally, the supernatant was collected by centrifugation at 12,000 rpm for 10 min, and was concentrated to 2 mL using a rotary evaporator. The crude extract was obtained by suspending samples in methanol and performing filtration (0.22 μm, Nylon).
4.3. Construction of the Gene Knockout Mutants of P. protegens Pf-5
The construction process of the ΔpltB mutant is used as the example. Briefly, 770 bp upstream and 770 bp downstream fragments were amplified by PCR with two pairs of primers, pltB-F1 (Xba I) and pltB-R1, as well as pltB-F2 and pltB-R2 (Hind III) (Supporting Information Tables S1 and S2), respectively, to obtain the deleted mutant of the pltB gene. The pltB-R1 primer was designed to possess the complementary fragment to the pltB-F2 primer. The upstream and downstream PCR products were infused by using overlap PCR with the primers pltB-F1 and pltB-R2. Subsequently, the 1540 bp fusing fragment was digested with Xba I and Hind III, and cloned into the same sites of the suicide vector pK18mobsacB. The recombinant plasmid pK18-pltB was transferred from E. coli Top10 to P. protegens Pf-5, with the help of E. coli Top10 (pRK-2013), through triparental mating. The single cross-over clones were first selected on KB agar, including Amp and Km. After a second homologous recombination event, the Km-sensitive, Amp-resistant, and sucrose-resistant mutants of pltB were then screened. The ΔpltB mutant with a 7260 bp deletion in the pltB ORF (from 56 bp to 7315 bp) was validated by PCR and sequenced using the primers pltB-outF and pltB-outR. In the same way, we also constructed the Δofa, ΔphlA, ΔhcnABC, ΔrzxB, Δpfl4656, and ΔprnA mutants (Supporting Information Tables S1 and S2).
4.4. Antibacterial Activity Assay
The antibacterial activities of Pf-5 and its mutants (Δofa, ΔphlA, ΔpltB, ΔhcnABC, ΔrzxB, Δpfl4656, and ΔprnA), the crude extract, and three standards (pyoluteorin, 2,4-DAPG, and orfamide A) were determined by the agar diffusion method described previously [42]. Briefly, the P. ananatis DZ-12 strain was grown in LB broth medium overnight at 30 °C, 200 rpm. The overnight culture of P. ananatis DZ-12 OD600 was maintained at 1.0, and 1 mL of dilute suspensions of DZ-12 was mixed into 100 mL melted LB agar medium, and poured into 9 cm plates. Sterilized filter paper disks 0.6 cm in diameter were placed in 1/4 Petri plates. Then, 5 μL of bacterial suspension (OD600 = 2), or corresponding secondary metabolites, was added onto the filter paper. The same amount of methanol was used as a control. The plates were incubated at 30 °C for 12 h, and the plates were then photographed, and the diameter of the inhibition zone was measured. Standard pyoluteorin (Cas No. 25683-07-2) and 2,4-DAPG (Cas No. 2161-86-6) were purchased from Apexbio (Houston, TX, USA). Orfamide A (Cas No. 939960-34-6) was purchased from Bioaustralis (Smithfield, NSW, Australia).
4.5. Bio-Surfactant Activity Assay
Bio-surfactant activity was analyzed by using the expelling oil method previously described by Liang et al. [21]. To obtain the red oil, a 10 mL light fraction of paraffin oil (CAS: 8042-47-5, Sangon Biotech Co., Ltd., Shanghai, China) was dyed red by using 10 mg Sudan III. The 1 mL volume of red oil was dropped onto the 9 cm plate with 20 mL double distilled water (ddH2O). After spreading the red oil across the whole surface, 1 μL crude extract was applied to the center of the plate, with 1 μL methanol being used as a control. A total of 1 μL of 875 μg/mL pyoluteorin, 180 μg/mL 2,4-DAPG, and 1030 orfamide A μg/mL were also detected by using the same method.
4.6. Identification of Compounds Using High-Performance Liquid Chromatography Analysis
The crude extract was added to semi-preparative high-performance liquid chromatography HPLC (Waters, Milford, MA, USA) equipped with a C18 column (ZORBAX SB-C18, 4.6 × 250 mm, 5 μm, Agilent, Palo Alto, CA, USA). The mobile phase comprising A was acetonitrile (ACN), with 0.1% (vol/vol) trifluoroacetic acid (TFA); Mobile B was Milli-Q water with 0.1% (vol/vol) TFA. The flow rate was 1 mL/min, and the injection volume for each sample was 5 μL. Pyoluteorin and 2,4-DAPG were detected by a gradient elution procedure: 5–95% ACN for 20 min, held at 95% CAN for 5 min. The UV absorptions at 310 nm and 270 nm were recorded for pyoluteorin and 2,4-DAPG, respectively. For the detection of orfamide A, the sample was eluted from 60–90% ACN in 10 min and held at 90% for 20 min. The detection was processed under UV absorption at 205 nm.
4.7. Observed Structural Changes in P. ananatis DZ-12 Caused by Crude Extract
Scanning electron microscopy (SEM) and transmission electron microscopy (TEM) were used to observe the ultrastructural and morphology changes of DZ-12 cells treated with crude extract according to the previous method [31]. For SEM, the cells of DZ-12 were prefixed with 2.5% glutaraldehyde and followed by three rounds of rinsing using 100 mM phosphate buffer. Subsequently, the samples were post-fixed for 3 h in 1% osmium tetroxide, and dehydrated using an ethanol gradient. After that, the samples were coated with gold particles and observed via Hitachi S-3000N SEM at a 5 kV voltage (Hitachi, Tokyo, Japan). For TEM analysis, the prefixed cells were embedded in Epon 812, sectioned using an ultra-microtome, and examined with a Hitachi H-600 transmission electron microscope.
4.8. Biofilm Formation
The biofilm formation of P. ananatis DZ-12 was initially quantified by staining with crystal violet. The overnight culture of P. ananatis DZ-12 OD600 reached 0.6, and 200 μL cells were transformed into 20 mL of LB medium. The LB medium contained 16 × or 32× dilutions of crude extract (final concentration), followed by methanol that was used as a control. The bacterial cells were grown at 30 °C, 200 rpm. When the cell OD600 reached 1.0, 4 mL of cultured bacterial cells were transferred to a sterilized glass test tube and incubated at 28 °C for 48 h. The bacterial cultures were carefully removed from the glass test tube, and the test tubes were washed gently with ddH2O 2–3 times. Finally, the biofilms on the inner surface of glass tubes were stained with 0.1% crystal violet and quantified at 570 nm. The experiment was repeated thrice.
For the common observation of biofilm formation, a 24-well plate were used. When the OD600 of the P. ananatis DZ-12 cells reached 0.6, 3 μL of bacterial cultures were, respectively, inoculated into the wells. Each well contained 1.5 mL, with a different dilution of crude extract (16×, 32×) from Pf-5 and its mutants Δofa, ΔpltB, and ΔphlA. Three pure commercial compounds were used. The final concentration of pyoluteorin, orfamide A, and 2,4-DAPG, were, respectively, 27 μg/mL, 32 μg/mL, and 12 μg/mL. The 24-well plate was incubated at 28 °C for 48 h. The biofilm was observed.
In addition, we used the gfp gene-labeled strain DZ-12 to understand detailed observations of the biofilm previously described by [43]. A 24-well plate and Nunc Lab-Tek™ II chamber slides were used. Briefly, when the OD600 of the P. ananatis DZ-12 cells reached 0.6, 3 μL of bacterial culture was inoculated into the wells. Each well contained 1.5 mL, using a different dilution of crude extract (16×, 32×, and 64×). In the control treatment, equal volumes of methanol were added. Then, the wells were gently mixed, and 200 μL of corresponding mixtures were transferred to the Nunc Lab-Tek™ II chamber slides. The 24-well plate and the chamber slides were incubated at 28 °C for 48 h. To photograph the biofilms in the chamber slides, 24-well plates was observed using a Leica SP5 confocal microscope (Leica Microsystems Vertrieb GmbH, Wetzlar, Germany) equipped with Leica LAS AF software.
4.9. Reactive Oxygen Species Detection
The detection of reactive oxygen species (ROS) was performed according to the method previously described [34], with some modifications. Briefly, P. ananatis DZ-12 were exposed to a crude extract or 27 μg/mL pyoluteorin, 32 μg/mL orfamide A, and 12 μg/mL 2,4-DAPG, followed by incubation for 4 h at 28 °C. The DZ-12 bacterial cells were collected using centrifugation at room temperature, and then washed two times using 10 mM sodium phosphate buffer (PBS) (pH 7.4). The collected bacterial cells were resuspended using the appropriate amount of PBS to obtain OD600 = 1.0 of the bacterial suspension. Then, the bacterial cells were incubated with 10 μM dichloro-dihydro-fluorescein diacetate (DCFH-DA) (JianCheng Bioengineering, Nanjing, China) for 30 min at 19 °C–21 °C. ROS were measured on the basis of the intracellular peroxide-dependent oxidation of DCFH-DA to form the fluorescent compound 2′,7′-dichlorofluorescein (DCF). The samples were observed under fluorescence microscopy (Olympus BX43 microscope excitation 488 nm, emission 535 nm), and analyzed using cell Sens Standard Software v.6.2 (Tokyo, Japan). Fluorescence intensity was determined using an EnSight Multimode Plate Reader (Perkinelmer, Waltham, MA, USA).
4.10. Plant Infection Assay
When the OD600 of overnight cultured P. ananatis DZ-12 cells reached 0.6, the cultured cells (1%) were transferred into 20 mL of LB broth medium with various dilutions (16× or 32×) of crude extract, and grown at 28 °C, 200 rpm for 12 h. The bacterial cells were harvested by centrifugation and washed three times with PBS, and they were resuspended in LB medium to 1 × 107 cells/mL. Then, the bacterial cell suspension was sprayed onto the 7 days old leaves of maize seedlings (c.v B73 cultivar), and PBS was used as a negative control. The plants were maintained in a greenhouse at 30 °C during the evaluation period, with natural light. The development of disease symptoms was monitored and photographs were taken 7 days post-inoculation (dpi). The pathogenicity test assays were repeated in triplicate, with 12 technical replicates.
4.11. Statistical Analysis
A completely randomized design was used to conduct all experiments in the present study, with each experiment repeated thrice. A statistical analysis was performed by using the statistical package SPSS. Tukey’s HSD test was applied for the separation of means at p ≤ 0.05, after conducting the analysis of variance (ANOVA) for all data sets.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms23126431/s1.
Author Contributions
Q.G. and H.W. planned and designed this research. R.W., J.L. and P.L. performed the research and methodology. Q.G. and Z.W. helped in the analysis and compiled the result and data of the manuscript. L.Z., Q.A. and A.R.K. helped with the experiments and improved the manuscript writing. X.G. contributed to the critical revision of the manuscript. H.W. and J.Q. wrote the whole manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Korea-China cooperation research project (Development of rice multi-stress resistance green technologies and applications, 2022YFE0198100), National Natural Science Foundation of China (grants 31972325, 32172490), Natural Science Foundation of Jiangsu Province (CN) (BK20201239), the Special Fund for the Fundamental Research Funds for Central Universities (grant KYZ201708).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- De Maayer, P.; Chan, W.Y.; Rubagotti, E.; Venter, S.N.; Toth, I.K.; Birch, P.R.J.; Coutinho, T.A. Analysis of the Pantoea ananatis pan-genome reveals factors underlying its ability to colonize and interact with plant, insect and vertebrate hosts. BMC Genom. 2014, 15, 404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Gao, L.; Huang, H.; Zhao, Y.; Hanif, A.; Wu, H.; Gu, Q.; Wu, L.; Gao, X. Exploring the pathogenic function of Pantoea ananatis endogenous plasmid by an efficient and simple plasmid elimination strategy. Microbiol. Res. 2021, 246, 126710. [Google Scholar] [CrossRef] [PubMed]
- Malagi, G.; Santos, I.D.; Camochena, R.C.; Moccellin, R. Diagrammatic scale for severity evaluation of maize white foliar spot. Rev. Cienc. Agron. 2011, 42, 797–804. [Google Scholar] [CrossRef]
- Mamede, M.C.; Tebaldi, N.D.; Mota, L.; Martins, O.M.; Coelho, L. Detection of Pantoea ananatis in corn seeds on semi-selective medium. Trop. Plant Pathol. 2018, 43, 254–256. [Google Scholar] [CrossRef] [Green Version]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The rhizosphere microbiome and plant health. Trends. Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Mhatre, P.H.; Karthik, C.; Kadirvelu, K.; Divya, K.L.; Venkatasalam, E.P.; Srinivasan, S.; Ramkumar, G.; Saranya, C.; Shanmuganathan, R. Plant growth promoting rhizobacteria (PGPR): A potential alternative tool for nematodes bio-control. Biocatal. Agric. Biotechnol. 2019, 17, 119–128. [Google Scholar] [CrossRef]
- Walsh, U.F.; Morrissey, J.P.; O’Gara, F. Pseudomonas for biocontrol of phytopathogens: From functional genomics to commercial exploitation. Curr. Opin. Biotechnol. 2001, 12, 289–295. [Google Scholar] [CrossRef]
- Weller, D.M. Pseudomonas Biocontrol Agents of Soilborne Pathogens: Looking Back Over 30 Years. Phytopathology 2007, 97, 250–256. [Google Scholar] [CrossRef] [Green Version]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.M.; Bakker, P.A.H.M. Induced Systemic Resistance by Beneficial Microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [Green Version]
- Howell, C.R.; Stipanovic, R.D. Control of Rhizoctonia solani on cotton seedlings with Pseudomonas fluorescens and with an antibiotic produced by the bacterium. Phytopathology 1979, 69, 480–482. [Google Scholar] [CrossRef] [Green Version]
- Shaukat, S.S.; Siddiqui, I.A. Impact of biocontrol agents Pseudomonas fluorescens CHA0 and its genetically modified derivatives on the diversity of culturable fungi in the rhizosphere of mungbean. J. Appl. Microbiol. 2003, 95, 1039–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Gu, G.; Chen, W.; Gao, L.; Wu, X.; Zhang, L. The outer membrane protein OprF and the sigma factor SigX regulate antibiotic production in Pseudomonas fluorescens 2P24. Microbiol. Res. 2018, 206, 159–167. [Google Scholar] [CrossRef]
- Yue, S.; Huang, P.; Li, S.; Jan, M.; Hu, H.; Wang, W.; Zhang, X. Enhanced Production of 2-Hydroxyphenazine from Glycerol by a Two-Stage Fermentation Strategy in Pseudomonas chlororaphis GP72AN. J. Agric. Food Chem. 2020, 68, 561–566. [Google Scholar] [CrossRef] [PubMed]
- Slininger, P.J.; Burkhead, K.D.; Schisler, D.A.; Bothast, R.J. Isolation, identification, and accumulation of 2-acetamidophenol in liquid cultures of the wheat take-all biocontrol agent Pseudomonas fluorescens 2–79. Appl. Microbiol. Biotechnol. 2000, 54, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Hu, H.; Wang, W.; Zhang, X. Genetic engineering of Pseudomonas chlororaphis GP72 for the enhanced production of 2-Hydroxyphenazine. Microb. Cell Fact. 2016, 15, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, S.L.F.; Everts, K.L.; Gardener, B.M.; Masler, E.P.; Abdelnabby, H.M.E.; Skantar, A.M. Assessment of DAPG-producing Pseudomonas fluorescens for Management of Meloidogyne incognita and Fusarium oxysporum on Watermelon. J. Nematol. 2016, 48, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Mishra, J.; Arora, N.K. Secondary metabolites of fluorescent pseudomonads in biocontrol of phytopathogens for sustainable agriculture. Appl. Soil Ecol. 2018, 125, 35–45. [Google Scholar] [CrossRef]
- Loper, J.E.; Henkels, M.D.; Shaffer, B.T.; Valeriote, F.A.; Gross, H. Isolation and identification of rhizoxin analogs from Pseudomonas fluorescens Pf-5 by using a genomic mining strategy. Appl. Environ. Microbiol. 2008, 74, 3085–3093. [Google Scholar] [CrossRef] [Green Version]
- Paulsen, I.T.; Press, C.M.; Ravel, J.; Kobayashi, D.Y.; Myers, G.S.A.; Mavrodi, D.V.; DeBoy, R.T.; Seshadri, R.; Ren, Q.; Madupu, R.; et al. Complete genome sequence of the plant commensal Pseudomonas fluorescens Pf-5. Nat. Biotechnol. 2005, 23, 873–878. [Google Scholar] [CrossRef]
- Gross, H.; Stockwell, V.O.; Henkels, M.D.; Nowak-Thompson, B.; Loper, J.E.; Gerwick, W.H. The Genomisotopic Approach: A Systematic Method to Isolate Products of Orphan Biosynthetic Gene Clusters. Chem. Biol. 2007, 14, 53–63. [Google Scholar] [CrossRef] [Green Version]
- Liang, Z.; Qiao, J.; Li, P.; Zhang, L.; Qiao, Z.; Lin, L.; Yu, C.; Yang, Y.; Zubair, M.; Gu, Q.; et al. novel Rap-Phr system in Bacillus velezensis NAU-B3 regulates surfactin production and sporulation via interaction with ComA. Appl. Microbiol. Biotechnol. 2020, 104, 10059–10074. [Google Scholar] [CrossRef] [PubMed]
- Danhorn, T.; Fuqua, C. Biofilm Formation by Plant-Associated Bacteria. Annu. Rev. Microbiol. 2007, 61, 401–422. [Google Scholar] [CrossRef] [PubMed]
- Cadenas, E.; Davies, K.J.A. Mitochondrial free radical generation, oxidative stress, and aging11This article is dedicated to the memory of our dear friend, colleague, and mentor Lars Ernster (1920–1998), in gratitude for all he gave to us. Free Radical Bio. Med. 2000, 29, 222–230. [Google Scholar] [CrossRef]
- Smith, D.D.N.; Kirzinger, M.W.B.; Stavrinides, J. Draft Genome Sequence of the Antibiotic-Producing Epiphytic Isolate Pantoea ananatis BRT175. Genome Announc. 2013, 1, e00902-13. [Google Scholar] [CrossRef] [Green Version]
- Kraus, J.; Loper, J.E. Characterization of a Genomic Region Required for Production of the Antibiotic Pyoluteorin by the Biological Control Agent Pseudomonas fluorescens Pf-5. Appl. Environ. Microb. 1995, 61, 849–854. [Google Scholar] [CrossRef] [Green Version]
- Maurhofer, M.; Keel, C.; Haas, D.; Défago, G. Pyoluteorin production by Pseudomonas fluorescens strain CHA0 is involved in the suppression of Pythium damping-off of cress but not of cucumber. Eur. J. Plant Pathol. 1994, 100, 221–232. [Google Scholar] [CrossRef]
- Fan, D.; Yu, S.; Yang, Y.; Qu, S. Pyoluteorin Induces Apoptosis and Autophagy in NSCLC Cells. Biol. Pharm. Bull. 2021, 44, 976–983. [Google Scholar] [CrossRef]
- Ding, T.; Yang, L.; Zhang, W.; Shen, Y. Pyoluteorin induces cell cycle arrest and apoptosis in human triple-negative breast cancer cells MDA-MB-231. J. Pharm. Pharmacol. 2020, 72, 969–978. [Google Scholar] [CrossRef]
- Rose, M.M.; Scheer, D.; Hou, Y.; Hotter, V.S.; Komor, A.J.; Aiyar, P.; Scherlach, K.; Vergara, F.; Yan, Q.; Loper, J.E.; et al. The bacterium Pseudomonas protegens antagonizes the microalga Chlamydomonas reinhardtii using a blend of toxins. Environ. Microbiol. 2021, 23, 5525–5540. [Google Scholar] [CrossRef]
- Pellicciaro, M.; Padoan, E.; Lione, G.; Celi, L.; Gonthier, P. Pyoluteorin Produced by the Biocontrol Agent Pseudomonas protegens Is Involved in the Inhibition of Heterobasidion Species Present in Europe. Pathogens 2022, 11, 391. [Google Scholar] [CrossRef]
- Parrino, B.; Carbone, D.; Cascioferro, S.; Pecoraro, C.; Giovannetti, E.; Deng, D.; Di Sarno, V.; Musella, S.; Auriemma, G.; Cusimano, M.G.; et al. 1,2,4-Oxadiazole topsentin analogs as staphylococcal biofilm inhibitors targeting the bacterial transpeptidase sortase A. Eur. J. Med. Chem. 2021, 209, 112892. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.J.; Reed, C.S.; Melander, C. Effects of N-pyrrole substitution on the anti-biofilm activities of oroidin derivatives against Acinetobacter baumannii. Bioorg. Med. Chem. Lett. 2008, 18, 4325–4327. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Fang, F.; Li, Y. Synthesis and anti-biofilm activities of dihydro-pyrrol-2-one derivatives on Pseudomonas aeruginosa. Bioorg. Med. Chem. Lett. 2015, 25, 597–601. [Google Scholar] [CrossRef] [PubMed]
- Gu, Q.; Yang, Y.; Yuan, Q.; Shi, G.; Wu, L.; Lou, Z.; Huo, R.; Wu, H.; Borriss, R.; Gao, X.; et al. Bacillomycin D Produced by Bacillus amyloliquefaciens Is Involved in the Antagonistic Interaction with the Plant-Pathogenic Fungus Fusarium graminearum. Appl. Environ. Microb. 2017, 83, e01075-17. [Google Scholar] [CrossRef] [Green Version]
- Meena, K.R.; Kanwar, S.S.; Freire, D. Lipopeptides as the Antifungal and Antibacterial Agents: Applications in Food Safety and Therapeutics. Biomed Res. Int. 2015, 2015, 473050. [Google Scholar] [CrossRef] [Green Version]
- Raaijmakers, J.M.; De Bruijn, I.; Nybroe, O.; Ongena, M. Natural functions of lipopeptides from Bacillus and Pseudomonas: More than surfactants and antibiotics. FEMS Microbiol. Rev. 2010, 34, 1037–1062. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, T.H.; Nybroe, O.; Koch, B.; Hansen, M.; Sørensen, J. Genes involved in cyclic lipopeptide production are important for seed and straw colonization by Pseudomonas sp. strain DSS73. Appl. Environ. Microb. 2005, 71, 4112–4116. [Google Scholar] [CrossRef] [Green Version]
- Aiyar, P.; Schaeme, D.; García-Altares, M.; Carrasco Flores, D.; Dathe, H.; Hertweck, C.; Sasso, S.; Mittag, M. Antagonistic bacteria disrupt calcium homeostasis and immobilize algal cells. Nat. Commun. 2017, 8, 1756. [Google Scholar] [CrossRef]
- Jang, J.Y.; Yang, S.Y.; Kim, Y.C.; Lee, C.W.; Park, M.S.; Kim, J.C.; Kim, I.S. Identification of Orfamide A as an Insecticidal Metabolite Produced by Pseudomonas protegens F6. J. Agric. Food Chem. 2013, 61, 6786–6791. [Google Scholar] [CrossRef]
- Hassan, K.A.; Johnson, A.; Shaffer, B.T.; Ren, Q.; Kidarsa, T.A.; Elbourne, L.D.; Hartney, S.; Duboy, R.; Goebel, N.C.; Zabriskie, T.M.; et al. Inactivation of the GacA response regulator in Pseudomonas fluorescens Pf-5 has far-reaching transcriptomic consequences. Environ. Microbiol. 2010, 12, 899–915. [Google Scholar] [CrossRef]
- Oni, F.E.; Geudens, N.; Omoboye, O.O.; Bertier, L.; Hua, H.; Adiobo, A.; Sinnaeve, D.; Martins, J.C.; Höfte, M. Fluorescent Pseudomonas and cyclic lipopeptide diversity in the rhizosphere of cocoyam (Xanthosoma sagittifolium). Environ. Microbiol. 2019, 21, 1019–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Gu, Q.; Xie, Y.; Lou, Z.; Xue, P.; Fang, L.; Yu, C.; Jia, D.; Huang, G.; Zhu, B.; et al. Cold-adapted Bacilli isolated from the Qinghai–Tibetan Plateau are able to promote plant growth in extreme environments. Environ. Microbiol. 2019, 21, 3505–3526. [Google Scholar] [CrossRef] [PubMed]
- Banin, E.; Vasil, M.L.; Greenberg, E.P. Iron and Pseudomonas aeruginosa biofilm formation. Proc. Natl. Acad. Sci. USA 2005, 102, 11076. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).