
A Glutathione Peroxidase Gene from Litopenaeus vannamei Is Involved in Oxidative Stress Responses and Pathogen Infection Resistance

 ,
, 
Abstract
:1. Introduction
2. Results and Discussion
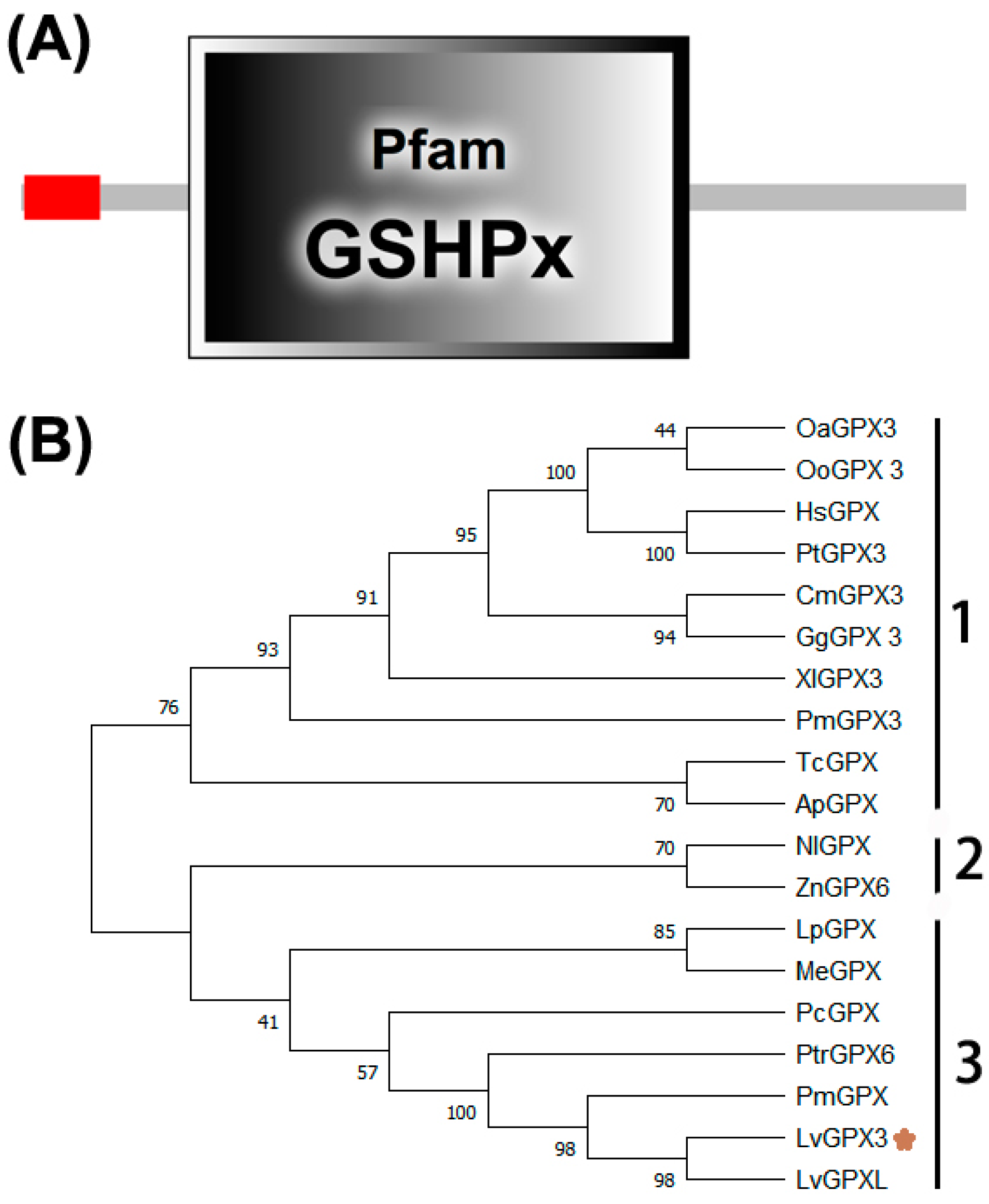
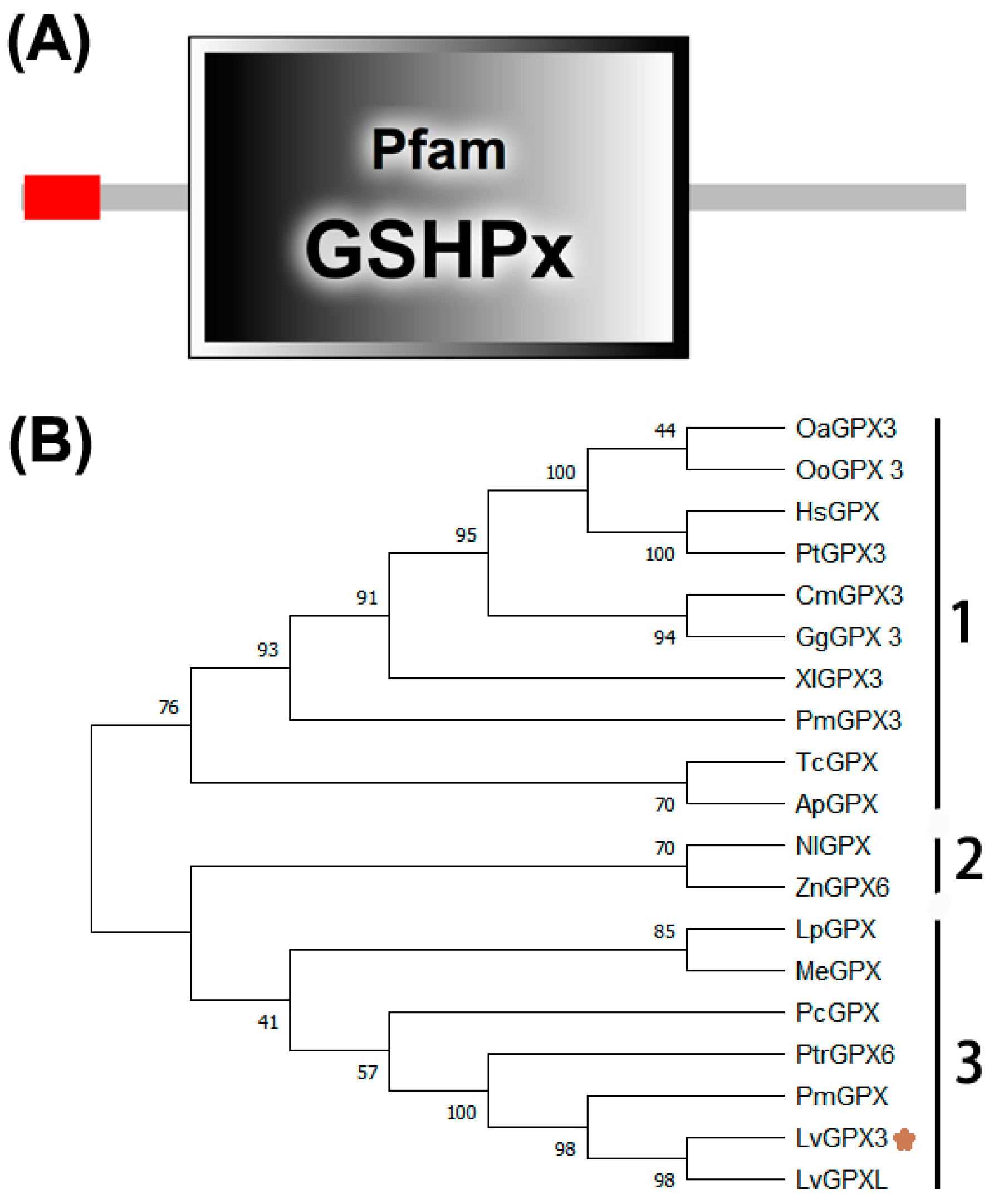
2.1. LvGPX3 Cloning and Sequence Analysis
2.2. Phylogenetic Analysis
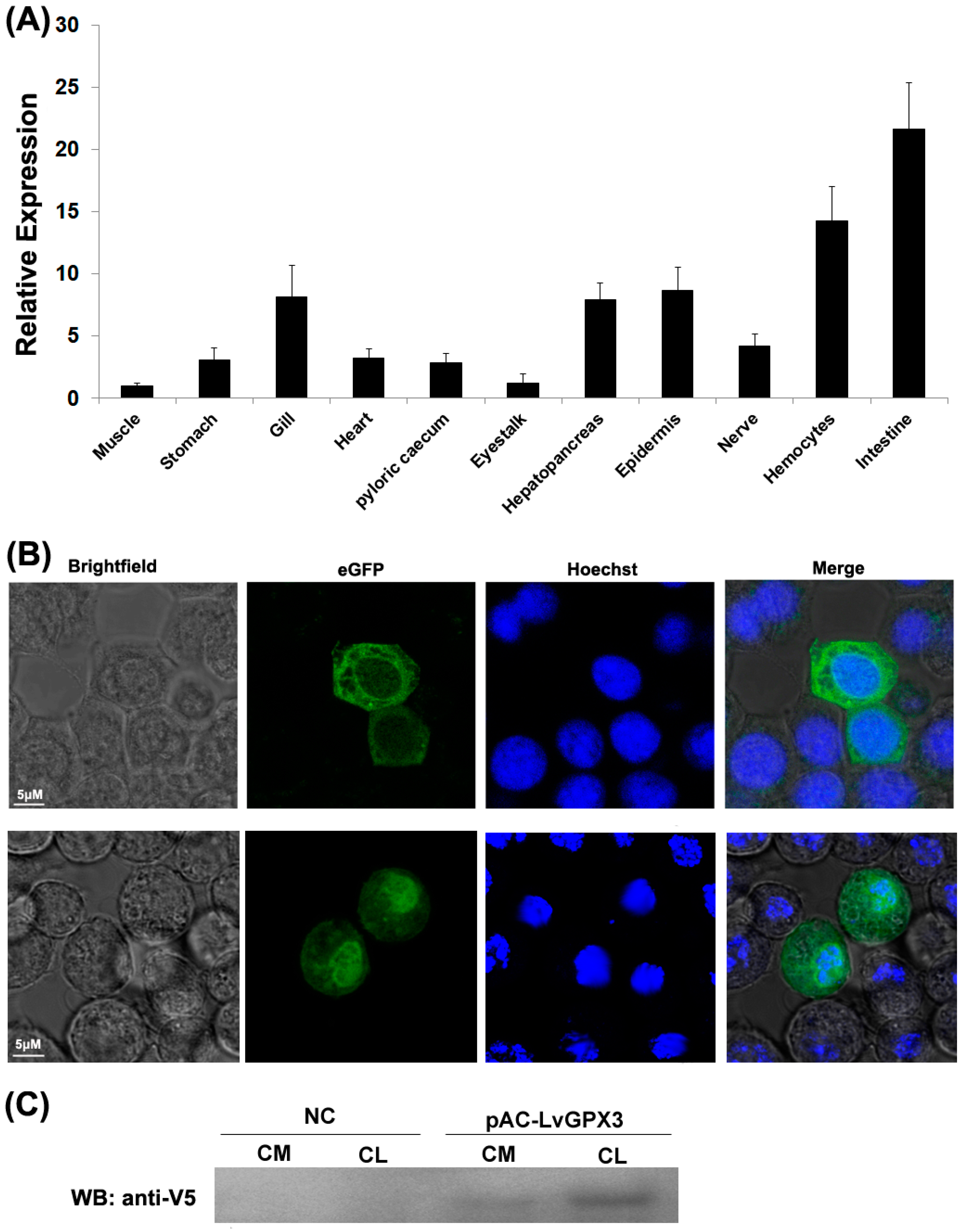
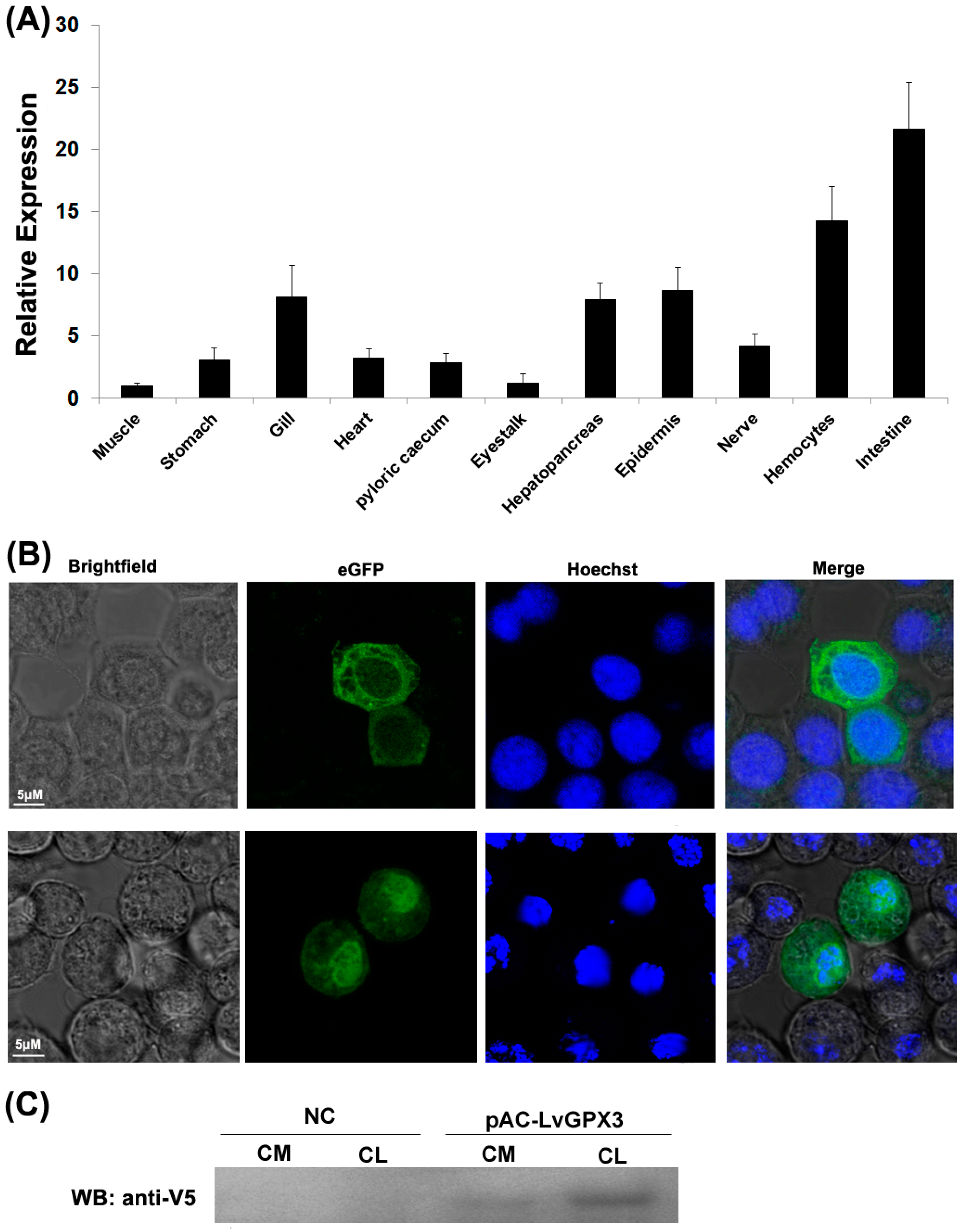
2.3. Constitutive Transcription and Protein Expression of LvGPX3
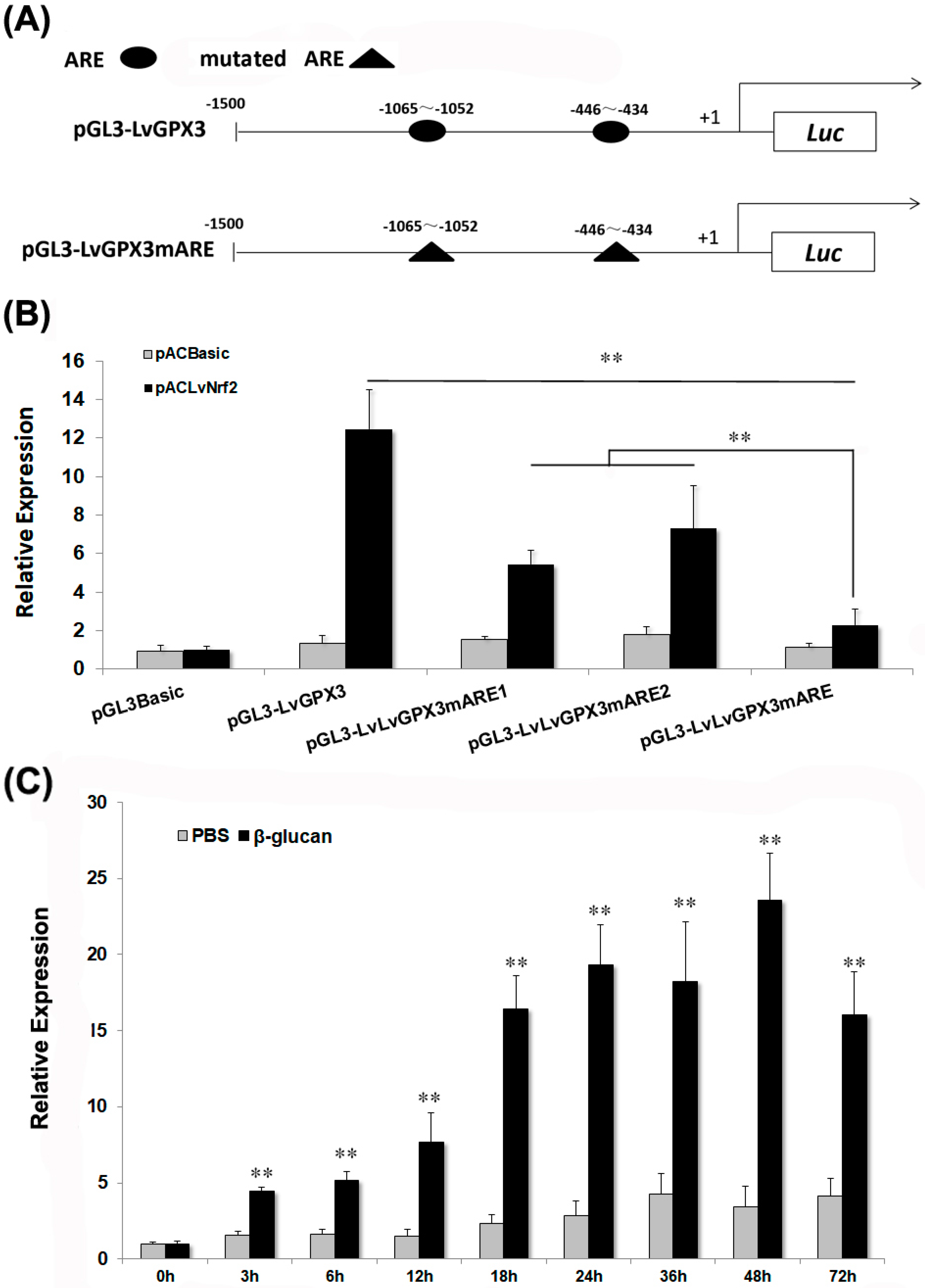
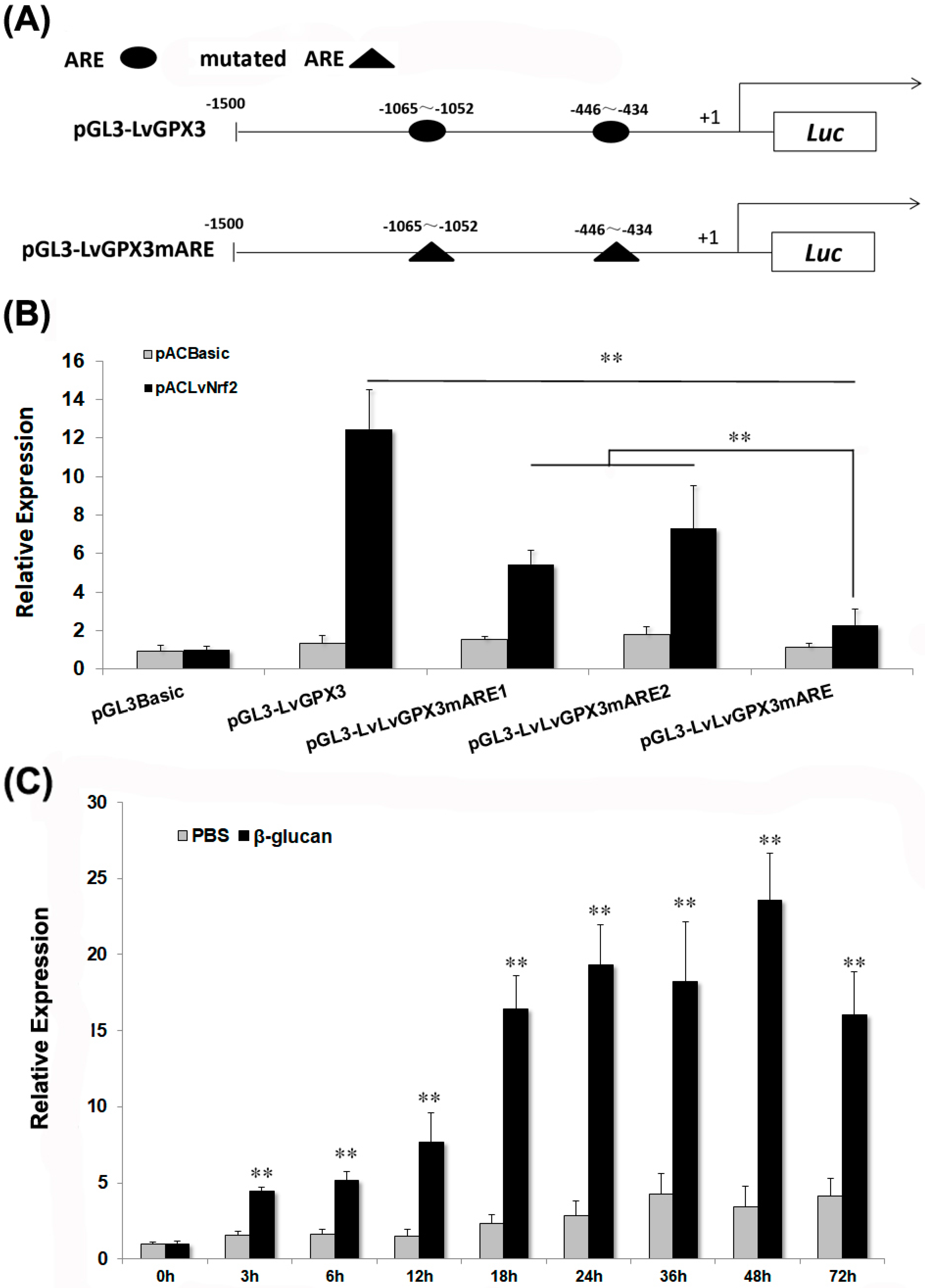
2.4. LvGPX3 was Induced with Oxidative Stress
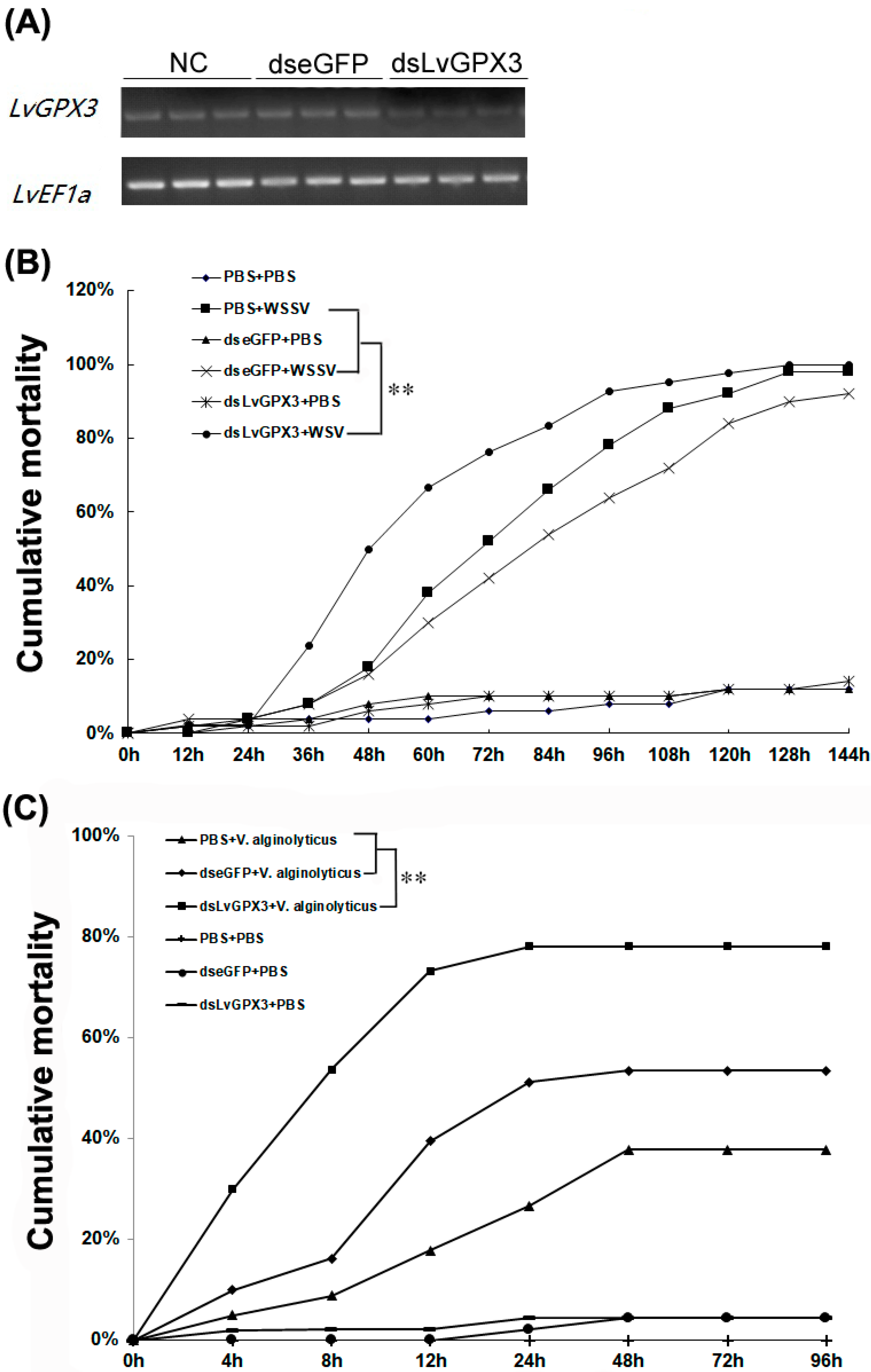
2.5. Knockdown Expression of LvGPX3 Depressed the Antioxidative Stress Response in Shrimp
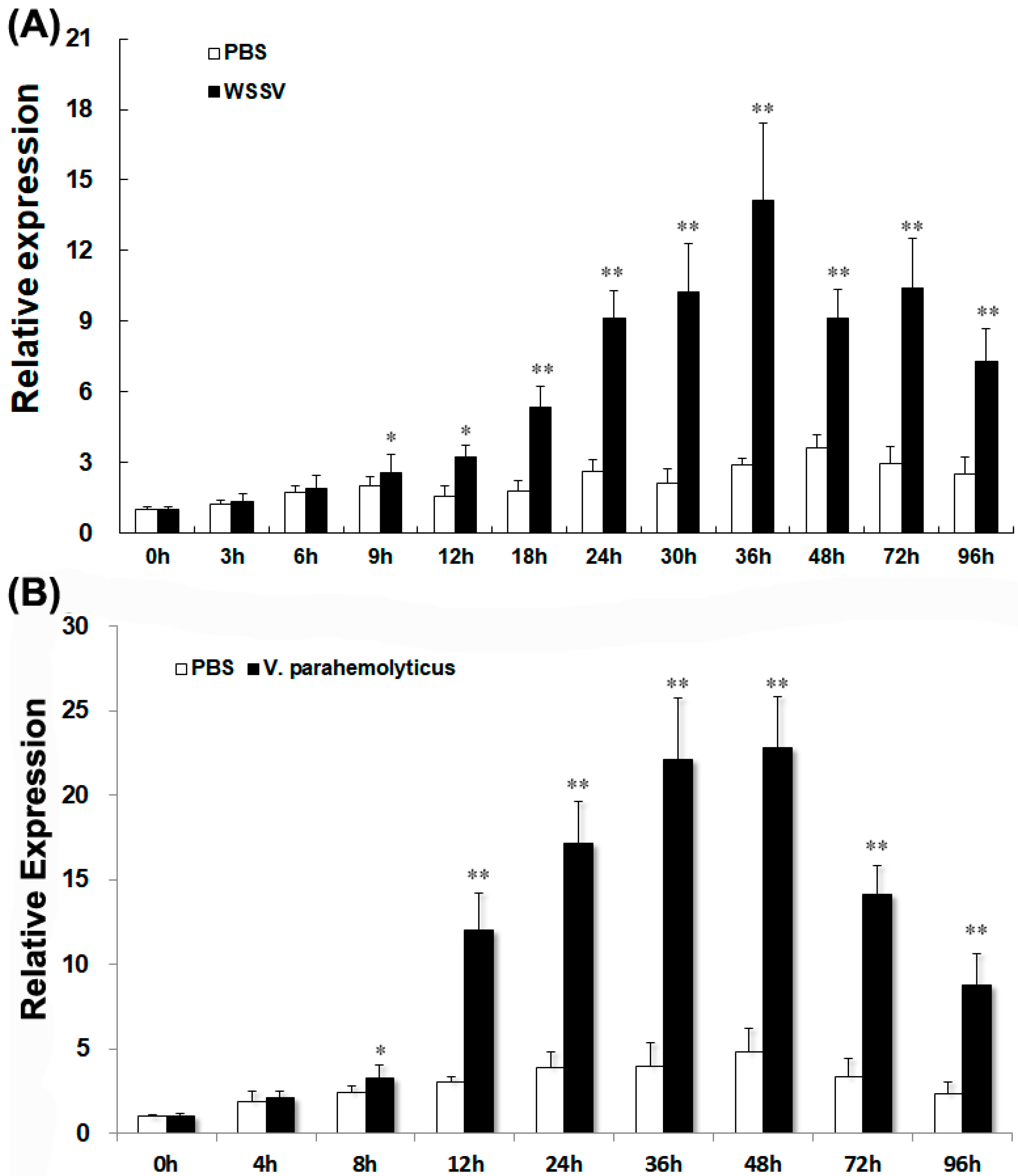
2.6. LvGPX3 Expression was Induced after Infection with WSSV or V. alginolyticus
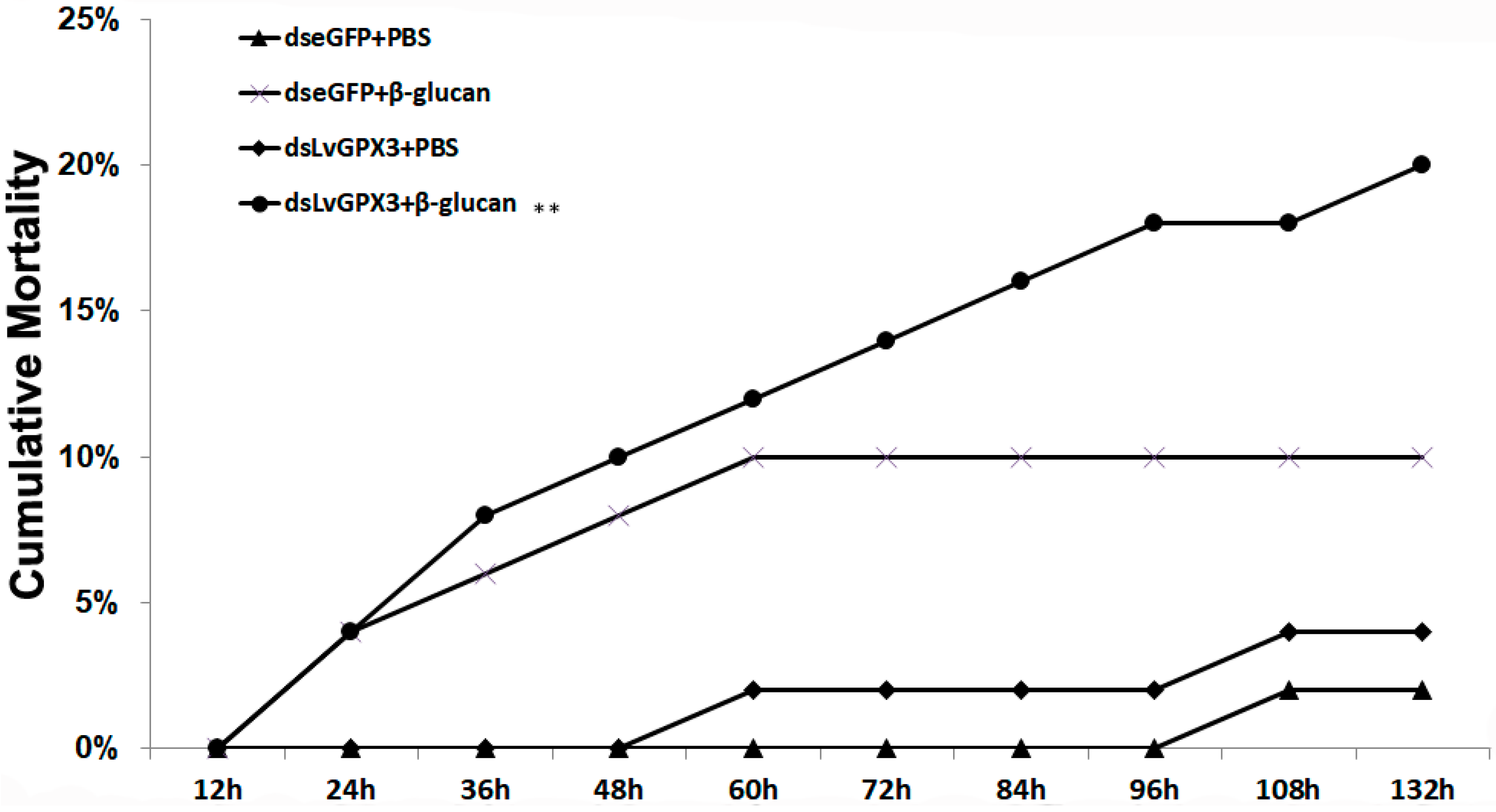
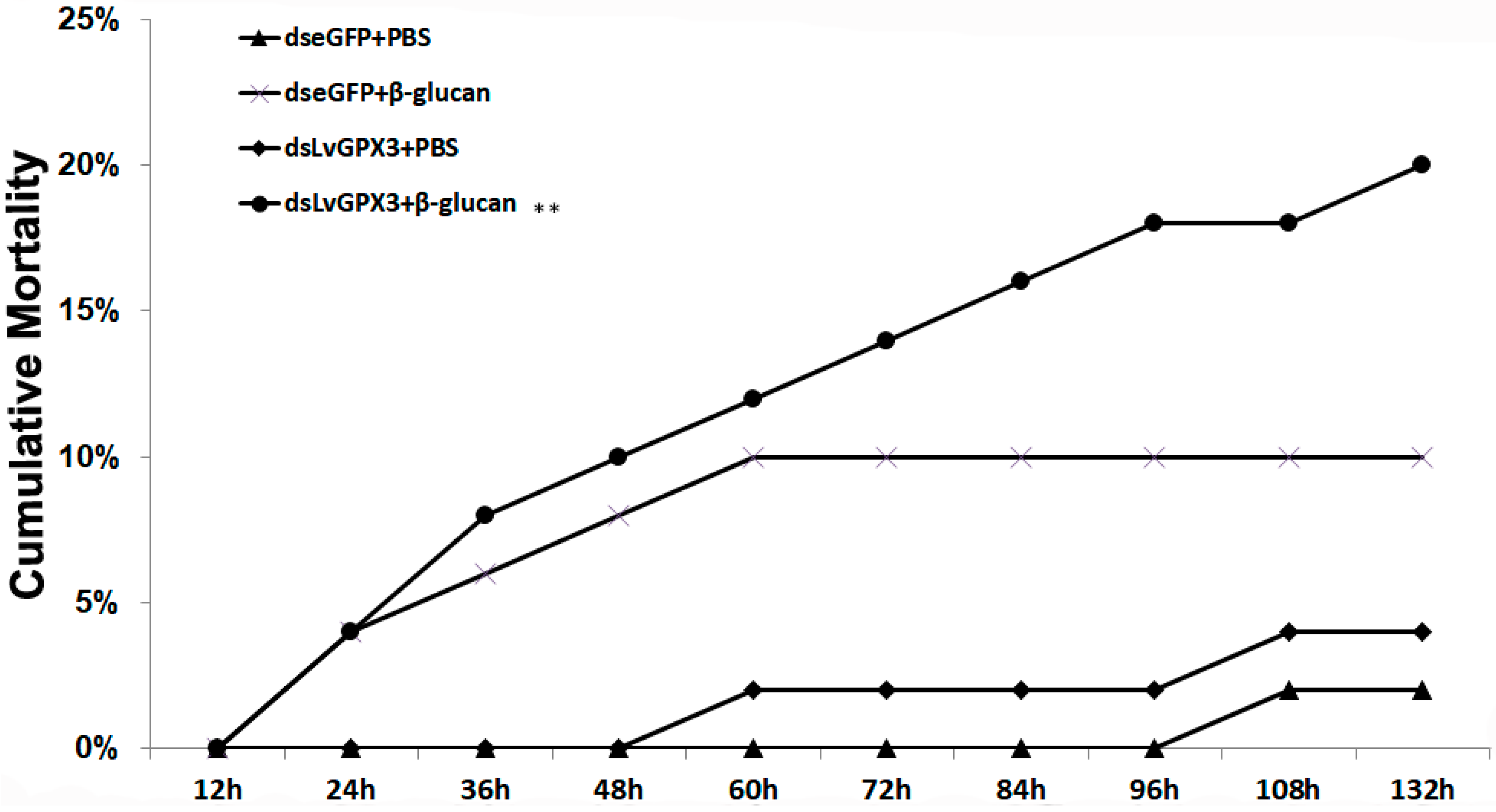
2.7. Knockdown of LvGPX3 Increased the Cumulative Mortality of WSSV- or V. alginolyticus-Infected Shrimps
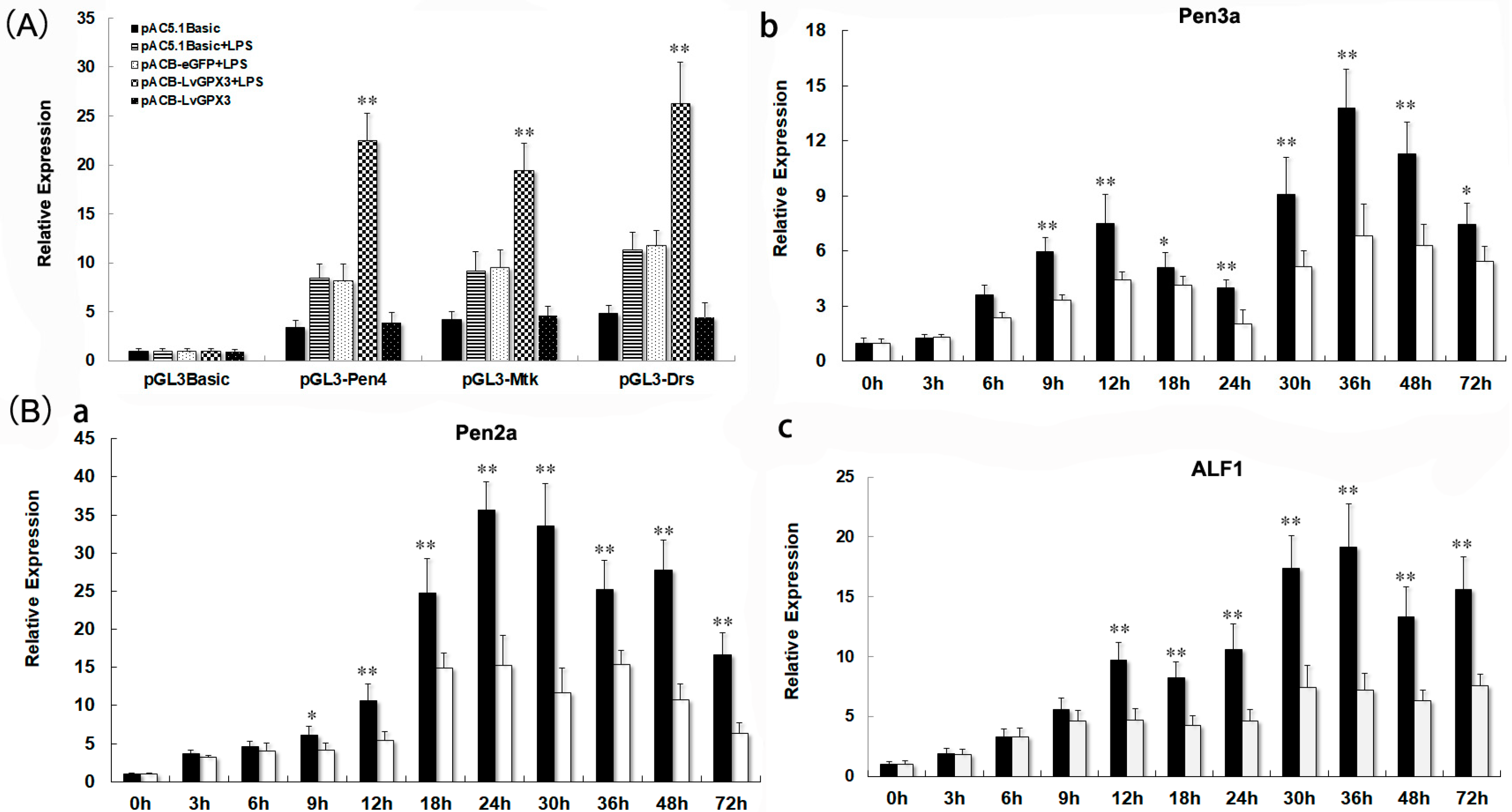
2.8. LvGPX3 Enhanced the Immune Response to Pathogens in Shrimp
3. Materials and Methods
3.1. Plasmid Vectors Construction
3.2. Cell Culture and Dual-Luciferase Reporter Gene Assay
3.3. Synthesis of Double-Stranded RNA
3.4. Oxidative Stress Induction and Preparation of Templates for Real-Time RT-PCR Assays
3.5. Pathogenic Challenge
3.6. Cumulative Mortality of LvGPX3-Knockdown Shrimp following Injection with WSSV, V. alginolyticus or β-Glucan
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Koeberle, S.C.; Gollowitzer, A.; Laoukili, J.; Kranenburg, O.; Werz, O.; Koeberle, A.; Kipp, A. Distinct and overlapping functions of glutathione peroxidases 1 and 2 in limiting NF-κB-driven inflammation through redox-active mechanisms. Redox Biol. 2020, 28, 101338. [Google Scholar] [CrossRef] [PubMed]
- Arthur, J.R. The glutathione peroxidases. Cell. Mol. Life Sci. 2001, 57, 1825–1835. [Google Scholar] [CrossRef]
- Borchert, A.; Kalms, J.; Roth, S.R.; Rademacher, M.; Schmidt, A.; Holzhutter, H.-G.; Kühn, H.; Scheerer, P. Crystal structure and functional characterization of selenocysteine-containing glutathione peroxidase 4 suggests an alternative mechanism of peroxide reduction. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2018, 1863, 1095–1107. [Google Scholar] [CrossRef]
- Rojas, J.; Díaz, A.A.B. Chapter 1-Antioxidant Activity of Phenolic Compounds Biosynthesized by Plants and Its Relationship With Prevention of Neurodegenerative Diseases—ScienceDirect. Bioact. Compd. 2019, 3–31. [Google Scholar] [CrossRef]
- Nirgude, S.; Choudhary, B. Insights into the role of GPX3, a highly efficient plasma antioxidant, in cancer. Biochem. Pharmacol. 2020, 184, 114365. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, Y. Role of Mitochondrial Phospholipid Hydroperoxide Glutathione Peroxidase (PHGPx) as a Antiapoptotic Factor. Biol. Pharm. Bull. 2004, 27, 956–960. [Google Scholar] [CrossRef] [Green Version]
- Coimbra, D.; Alva, N.; Duran, M.; Carbonell, T.; Rama, R. Oxidative stress and apoptosis after acute respiratory hypoxia and reoxygenation in rat brain. Redox Biol. 2017, 12, 216–225. [Google Scholar] [CrossRef]
- Huzayyin, A.; Andreazza, A.C.; Turecki, G.; Cruceanu, C.; Rouleau, G.A.; Alda, M.; Young, L.T. Decreased global methylation in patients with bipolar disorder who respond to lithium. Int. J. Neuropsychopharmacol. 2014, 17, 561–569. [Google Scholar] [CrossRef]
- Strycharz-Dudziak, M.; Kiełczykowska, M.; Drop, B.; Swiatek, L.; Kliszczewska, E.; Musik, I.; Polz-Dacewicz, M. Total Antioxidant Status (TAS), Superoxide Dismutase (SOD), and Glutathione Peroxidase (GPx) in Oropharyngeal Cancer Associated with EBV Infection. Oxidative Med. Cell. Longev. 2019, 6, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-H.; Tseng, M.-C.; Cheng, W. Identification and cloning of the antioxidant enzyme, glutathione peroxidase, of white shrimp, Litopenaeus vannamei, and its expression following Vibrio alginolyticus infection. Fish Shellfish Immunol. 2007, 23, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhong, X.; Shi, W.; Guo, B. Study of malondialdehyde (MDA) content, superoxide dismutase (SOD) and glutathione peroxidase (GSH-Px) activities in chickens infected with avian infectious bronchitis virus. Afr. J. Biotechnol. 2011, 10, 9213–9217. [Google Scholar]
- Alfaro, A.; Nguyen, T.V.; Bayot, B.; Rodríguez, J.; Borbor, C.L.D.; Sonnenholzner, S. Metabolic responses of whiteleg shrimp to white spot syndrome virus (WSSV). J. Invertebr. Pathol. 2021, 180, 107545. [Google Scholar] [CrossRef] [PubMed]
- Parrilla, P.; Zenteno-Savin, T.; Javier, M.B.F. Antioxidant enzyme activity in pacific whiteleg shrimp (Litopenaeus vannamei) in response to infection with white spot syndrome virus. Aquaculture 2013, 380, 41–46. [Google Scholar] [CrossRef]
- Wang, L.; Wu, J.; Wang, W.-N.; Cai, D.-X.; Liu, Y.; Wang, A.-L. Glutathione peroxidase from the white shrimp Litopenaeus vannamei: Characterization and its regulation upon pH and Cd exposure. Ecotoxicology 2012, 21, 1585–1592. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.-T.; Aoki, T.; Huang, Y.-T.; Hirono, I.; Chen, T.-C.; Huang, J.-Y.; Chang, G.-D.; Lo, C.F.; Wang, H.-C. White Spot Syndrome Virus Induces Metabolic Changes Resembling the Warburg Effect in Shrimp Hemocytes in the Early Stage of Infection. J. Virol. 2011, 85, 12919–12928. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y. Comparative transcriptome analysis reveals the different roles between hepatopancreas and intestine of Litopenaeus vannamei in immune response to aflatoxin B1 (AFB1) challenge. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2019, 222, 1–10. [Google Scholar] [CrossRef]
- Ma, Q. Role of Nrf2 in Oxidative Stress and Toxicity. Annu. Rev. Pharmacol. 2013, 53, 401–426. [Google Scholar] [CrossRef] [Green Version]
- Battin, E.E.; Brumaghim, J. Antioxidant Activity of Sulfur and Selenium: A Review of Reactive Oxygen Species Scavenging, Glutathione Peroxidase, and Metal-Binding Antioxidant Mechanisms. Cell Biochem. Biophys. 2009, 55, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Jia, M.; Qin, D.; Zhao, C.; Chai, L.; Yu, Z.; Wang, W.; Tong, L.; Lv, L.; Wang, Y.; Rehwinkel, J.; et al. Redox homeostasis maintained by GPX4 facilitates STING activation. Nat. Immunol. 2020, 21, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, S.; Lv, K.; Yin, B.; Xiao, B.; Li, S.; He, J.; Li, C. An invertebrate STING from shrimp activates an innate immune defense against bacterial infection. FEBS Lett. 2017, 591, 1010–1017. [Google Scholar] [CrossRef] [PubMed]
- Qiao, G.; Kim, S.-K.; Cho, Y.-R.; Kim, S.; Jang, I.K. Expression of c-Type Lysozyme from the Fleshy Shrimp Fenneropenaeus chinensis Is Upregulated Following Vibrio anguillarum and Lipopolysaccharide Injection. Fish. Aquat. Sci. 2013, 16, 267–272. [Google Scholar] [CrossRef] [Green Version]
- Yihong, C.; Zhao, L.; Pang, L.-R.; Li, X.-Y.; Weng, S.-P.; He, J.-G. Identification and characterization of Inositol-requiring enzyme-1 and X-box binding protein 1, two proteins involved in the unfolded protein response of Litopenaeus vannamei. Dev. Comp. Immunol. 2012, 38, 66–77. [Google Scholar]
- Imler, J.-L.; Tauszig-Delamasure, S.; Jouanguy, E.; Forestier, C.; Hoffmann, J.A. LPS-induced immune response in Drosophila. J. Endotoxin Res. 2000, 6, 459–462. [Google Scholar] [CrossRef]
- Yihong, C.; Yuan, F.; Bi, H.-T.; Zhang, Z.-Z.; Yue, H.-T.; Yuan, K.; Chen, Y.-G.; Wen, S.-P.; He, J.-G. Transcriptome analysis of the unfolded protein response in hemocytes of Litopenaeus vannamei. Fish Shellfish Immunol. 2016, 54, 153–163. [Google Scholar]
- Thitamadee, S.; Srisala, J.; Taengchaiyaphum, S.; Sritunyalucksana, K. Double-dose β-glucan treatment in WSSV-challenged shrimp reduces viral replication but causes mortality possibly due to excessive ROS production. Fish Shellfish Immunol. 2014, 40, 478–484. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequence (5′-3′) |
| For gene expression a | |
| pAC-LvGPX3-EcoRⅠ-F | CGGGAATTCTATGTTGTGGGCGGGGTTCG |
| pAC-LvgGPX3-XhoⅠ-R | CGGCTCGAGGCAAATTCCTGGCTCAGGAGGAAC |
| pAC-LvNrf2-EcoRⅤ-F | CGGGATATCATGGAAGGCCCTGTAATTGA |
| pAC-LvNrf2-XbaⅠ-R | ATATCTAGACTCTGCTTGGGGTCATCCTTCCC |
| For real-time RT-PCR | |
| QPCR-LvGPX3-F | GATCGTCAATGTGGCGACCTA |
| QPCR-LvGPX3-R | CTCTTGCTTCCCGAATTGGTT |
| QPCR-LvPen2a-F | GGTTACAGGCCCGATACCCA |
| QPCR-LvPen2a-R | GTGACAACAGCTTCCGAACTTG |
| QPCR-LvPen3a-F | GCTTGCGTGATATGAGTGAGTG |
| QPCR-LvPen3a-R | AATTACAACGAAAGGCAGATGG |
| QPCR-LvALF1-F | GACAGGCTTCCGAGCAACAC |
| QPCR-LvALF1-R | GTGGCACAAGAGCAATCAGG |
| QPCR-LvCrustin-F | CGACGACAATGACGCAACAG |
| QPCR-LvCrustin-R | AAGACCTCCACCCAATCCAAA |
| QPCR-LvEF1a-F | GCTGATTGCGCCGTACTCAT |
| QPCR-LvEF1a-R | TCACGGGTCTGTCCGTTCTT |
| For dsRNA template amplification | |
| DsRNA-LvTRIM32-485-T7-F1 | GGATCCTAATACGACTCACTATAGG TGAGCTTCAGTTTGCGTCCAG |
| DsRNA-LvTRIM32-485-R1 | TACAGCGGGTGTTCATTCTCG |
| DsRNA-LvTRIM32-485-F2 | TGAGCTTCAGTTTGCGTCCAG |
| DsRNA-LvTRIM32-485-T7-R2 | GGATCCTAATACGACTCACTATAGG TACAGCGGGTGTTCATTCTCG |
| For reporter gene assay b | |
| pGL3-LvGPX3-Kpn I-F | ATAGGTACCCGTACCAACAGGCAACAACA |
| pGL3-LvGPX3-BglⅡ-R | CCGCTCGAGGCTGAAAAATAAATATGAAAGGTG |
| pGL3-LvGPX3mARE-F1 | agtcatcggatcCACGCATAAACATGCAAAAAAAC |
| pGL3-LvGPX3mARE-R1 | TACTCCATGTGTTTACAGAAATGTAGAT |
| pGL3-LvGPX3mARE-F2 | agacatcgcatcCACGTATAAACATAAGCATAAAAC |
| pGL3-LvGPX3mARE-R2 | TGTTTTATGTTTATGTTTATGCGTG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, J.; Li, B.; Hong, Q.; Yan, Z.; Yang, X.; Lu, K.; Chen, G.; Wang, L.; Chen, Y. A Glutathione Peroxidase Gene from Litopenaeus vannamei Is Involved in Oxidative Stress Responses and Pathogen Infection Resistance. Int. J. Mol. Sci. 2022, 23, 567. https://doi.org/10.3390/ijms23010567
Fan J, Li B, Hong Q, Yan Z, Yang X, Lu K, Chen G, Wang L, Chen Y. A Glutathione Peroxidase Gene from Litopenaeus vannamei Is Involved in Oxidative Stress Responses and Pathogen Infection Resistance. International Journal of Molecular Sciences. 2022; 23(1):567. https://doi.org/10.3390/ijms23010567
Chicago/Turabian StyleFan, Jinquan, Binbin Li, Qianming Hong, Zeyu Yan, Xinjun Yang, Kecheng Lu, Guoliang Chen, Lei Wang, and Yihong Chen. 2022. "A Glutathione Peroxidase Gene from Litopenaeus vannamei Is Involved in Oxidative Stress Responses and Pathogen Infection Resistance" International Journal of Molecular Sciences 23, no. 1: 567. https://doi.org/10.3390/ijms23010567
APA StyleFan, J., Li, B., Hong, Q., Yan, Z., Yang, X., Lu, K., Chen, G., Wang, L., & Chen, Y. (2022). A Glutathione Peroxidase Gene from Litopenaeus vannamei Is Involved in Oxidative Stress Responses and Pathogen Infection Resistance. International Journal of Molecular Sciences, 23(1), 567. https://doi.org/10.3390/ijms23010567