
The Influence of Angiotensin Peptides on Survival and Motility of Human High-Grade Serous Ovarian Cancer Cells in Serum Starvation Conditions

 ,
,  ,
,  ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
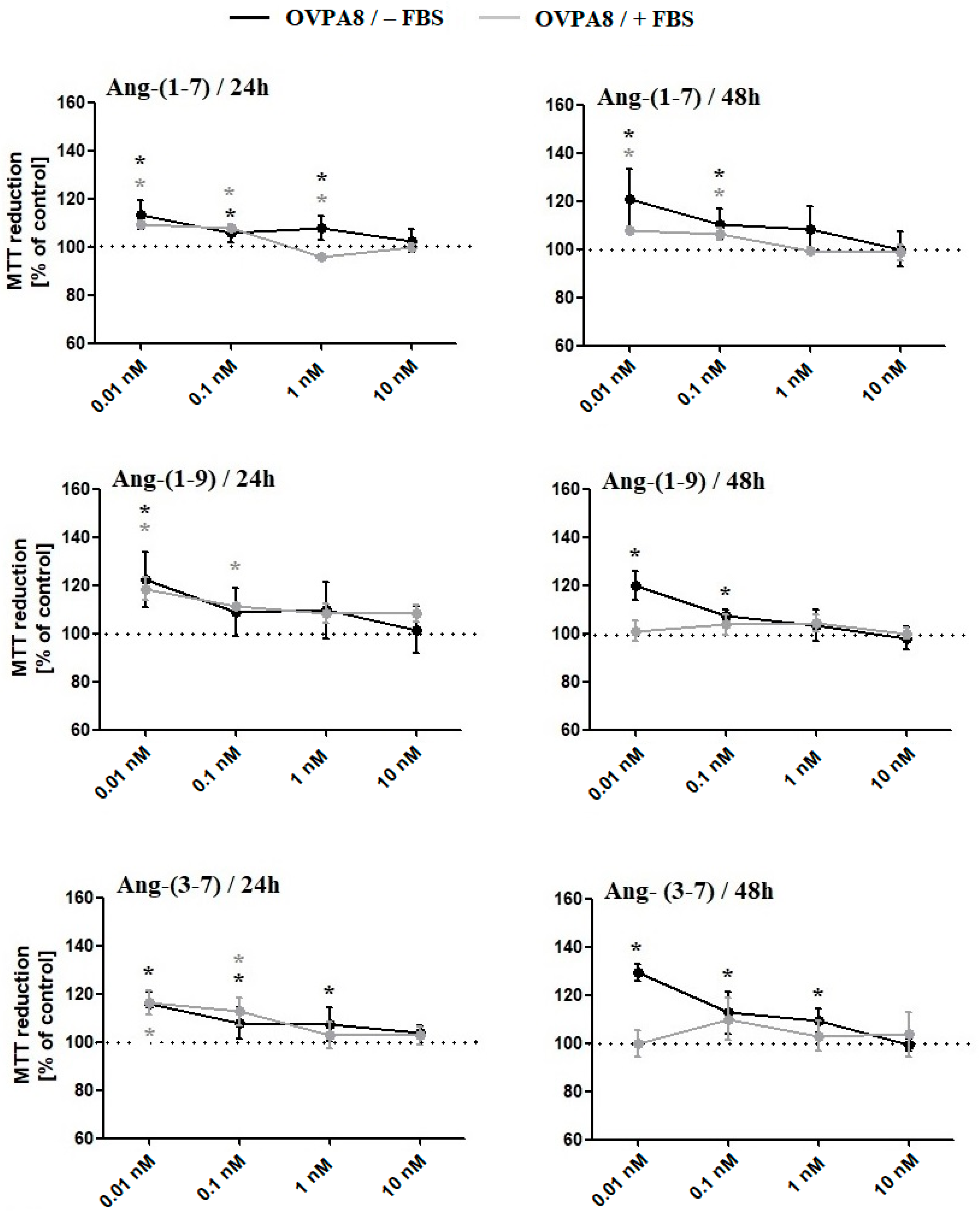
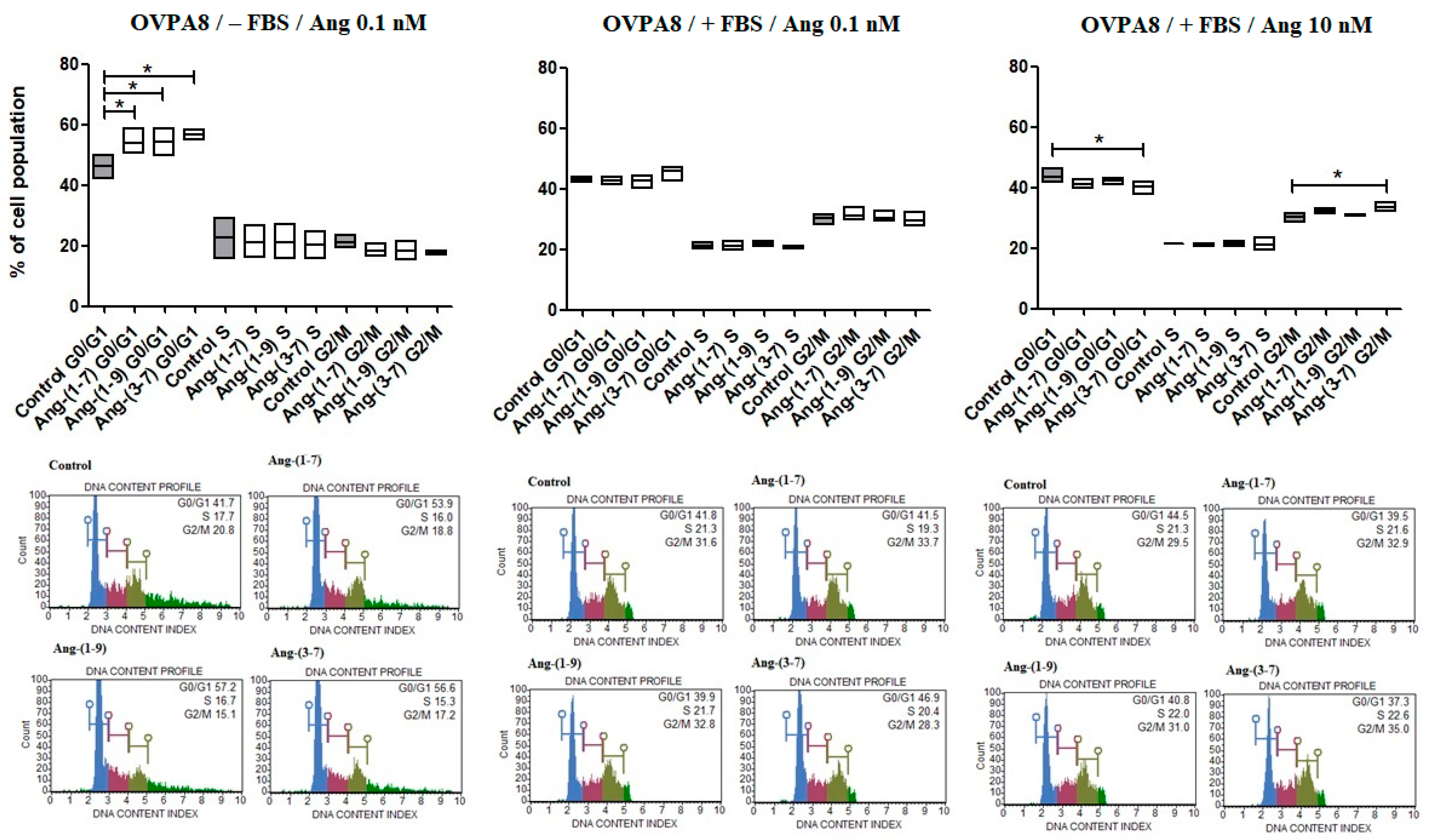
2.1. The Changes in OVPA8 Cells Viability, Metabolic Activity and Proliferation after Angiotensin Peptides Exposure
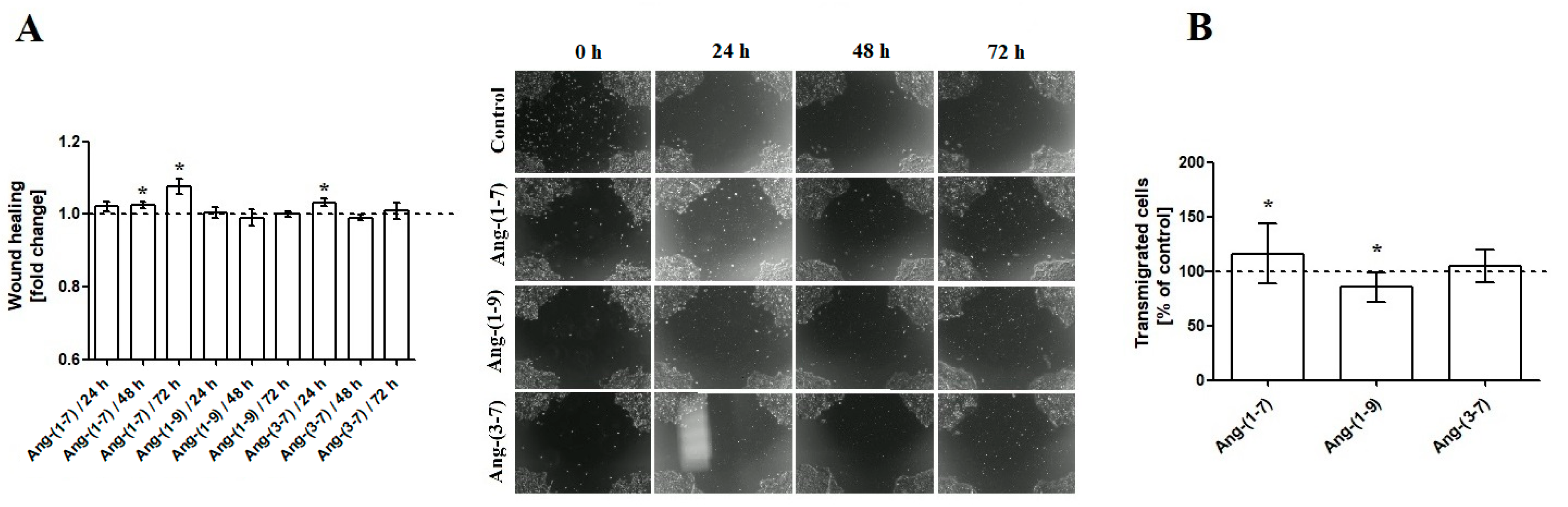
2.2. The Changes in OVPA8 Cell Mobility after Angiotensin Peptides Exposure
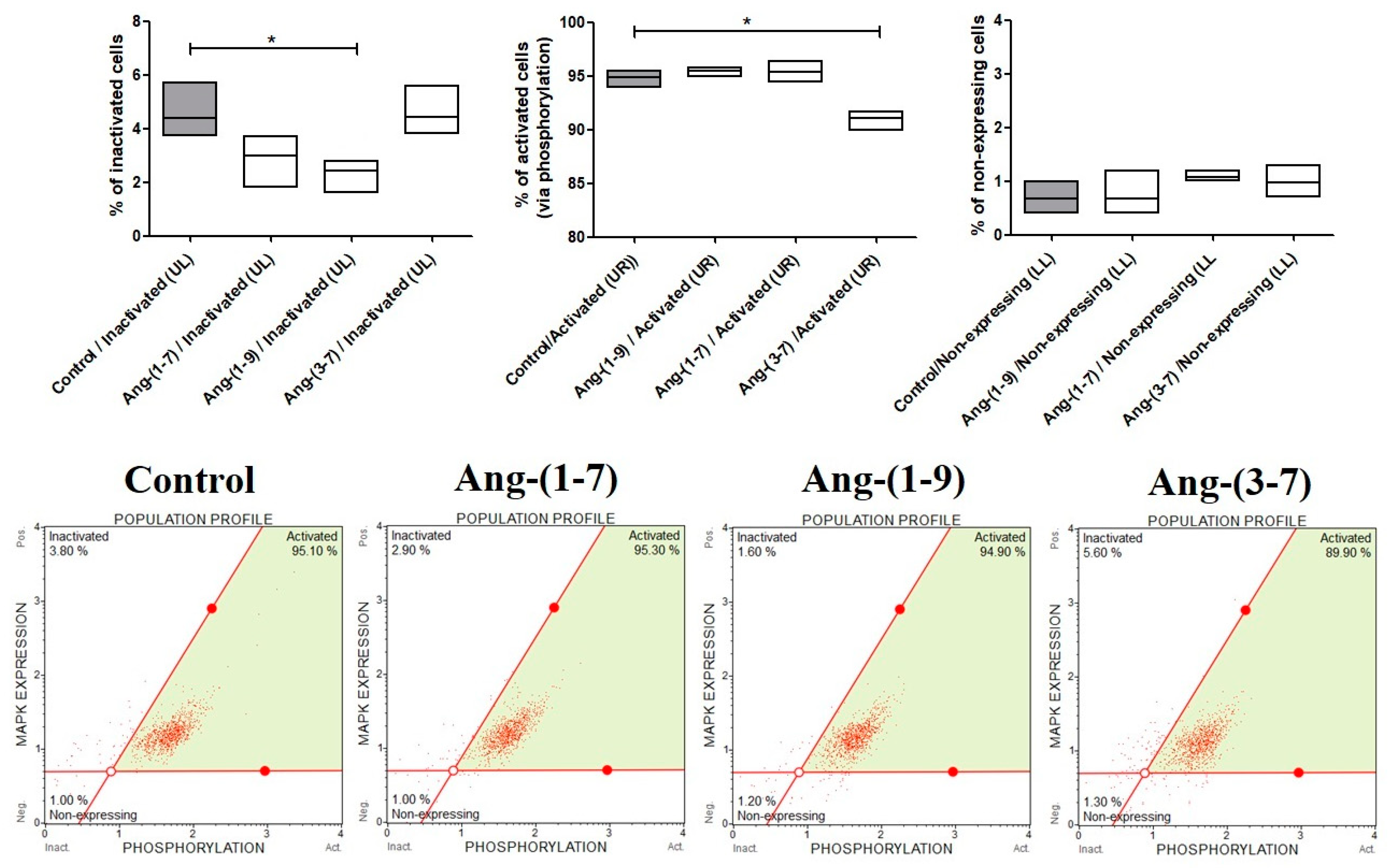
2.3. The Changes in MAPK Phosphorylation Relative to the Total MAPK Expression in OVPA8 Cells after Angiotensin Peptides Exposure
2.4. The Changes in the mRNA Expression of NFkB Family Members in OVPA8 Cells after Angiotensin Peptides Exposure
2.5. The Changes in the Level of Angiotensin Receptors after Angiotensin Peptides Exposure
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Reagents
4.2. MTT Assay and Alamar Blue Assay
4.3. Assay Muse® Cell Cycle Assay
4.4. Muse® Ki67 Proliferation Assay
4.5. Muse® MAPK Activation Dual Detection Assay
4.6. Wound Healing Assay
4.7. Boyden Chamber Assay
4.8. RT-qPCR
4.9. Western Blot
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Domińska, K. Involvement of ACE2/Ang-(1–7)/MAS1 Axis in the Regulation of Ovarian Function in Mammals. Int. J. Mol. Sci. 2020, 21, 4572. [Google Scholar] [CrossRef] [PubMed]
- Tudrej, P.; Kujawa, K.A.; Cortez, A.J.; Lisowska, K.M. Characteristics of in vitro model systems for ovarian cancer studies. Oncol. Clin. Pract. 2019, 15, 246–259. [Google Scholar] [CrossRef]
- Tudrej, P.; Olbryt, M.; Zembala-Nożyńska, E.; Kujawa, K.A.; Cortez, A.J.; Fiszer-Kierzkowska, A.; Wojciech Pigłowski, W.; Nikiel, B.; Głowala-Kosińska, M.; Bartkowska-Chrobok, A.; et al. Establishment and Characterization of the Novel High-Grade Serous Ovarian Cancer Cell Line OVPA8. Int. J. Mol. Sci. 2018, 19, 2080. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Fan, J.; Wu, F.; Huang, Q.; Guo, M.; Lv, Z.; Han, J.; Duan, L.; Hu, G.; Chen, L.; et al. The ACE2/Angiotensin-(1–7)/Mas Receptor Axis: Pleiotropic Roles in Cancer. Front. Physiol. 2017, 8, 276. [Google Scholar] [CrossRef]
- Ager, E.I.; Neo, J.; Christophi, C. The renin-angiotensin system and malignancy. Carcinogenesis 2008, 29, 1675–1684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domińska, K.; Lachowicz-Ochedalska, A. The involvement of the renin-angiotensin system (RAS) in cancerogenesis. Postepy Biochem. 2008, 54, 294–300. [Google Scholar]
- Bi, F.F.; Li, D.; Cao, C.; Li, C.Y.; Yang, Q. Regulation of angiotensin II type 1 receptor expression in ovarian cancer: A potential role for BRCA1. J. Ovarian Res. 2013, 6, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, Y.A.; Choi, C.H.; Do, I.G.; Song, S.Y.; Lee, J.K.; Cho, Y.J.; Choi, J.J.; Jeon, H.K.; Ryu, J.Y.; Lee, Y.Y.; et al. Dual targeting of angiotensin receptors (AGTR1 and AGTR2) in epithelial ovarian carcinoma. Gynecol. Oncol. 2014, 135, 108–117. [Google Scholar] [CrossRef]
- Ziaja, M.; Kinga Anna Urbanek, K.A.; Kowalska, K.; Piastowska-Ciesielska, A.W. Angiotensin II and Angiotensin Receptors 1 and 2—Multifunctional System in Cells Biology, What Do We Know? Cells 2021, 10, 381. [Google Scholar] [CrossRef] [PubMed]
- Haulica, I.; Bild, W.; Serban, D.N. Angiotensin peptides and their pleiotropic actions. J. Renin. Angiotensin Aldosterone Syst. 2005, 6, 121–131. [Google Scholar] [CrossRef] [Green Version]
- Suganuma, T.; Ino, K.; Shibata, K.; Kajiyama, H.; Nagasaka, T.; Mizutani, S.; Kikkawa, F. Functional expression of the angiotensin II type 1 receptor in human ovarian carcinoma cells and its blockade therapy resulting in suppression of tumor invasion, angiogenesis, and peritoneal dissemination. Clin. Cancer Res. 2005, 11, 2686–2694. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Yu, S.; Lam, M.M.T.; Poon, T.C.W.; Sun, L.; Jiao, Y.; Wong, A.S.T.; Lee, L.T.O. Angiotensin II promotes ovarian cancer spheroid formation and metastasis by upregulation of lipid desaturation and suppression of endoplasmic reticulum stress. J. Exp. Clin. Cancer Res. 2019, 38, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ino, K.; Shibata, K.; Kajiyama, H.; Yamamoto, E.; Nagasaka, T.; Nawa, A.; Nomura, S.; Kikkawa, F. Angiotensin II type 1 receptor expression in ovarian cancer and its correlation with tumour angiogenesis and patient survival. Br. J. Cancer 2006, 94, 552–560. [Google Scholar] [CrossRef] [Green Version]
- Chappell, M.C. Biochemical evaluation of the renin-angiotensin system: The good, bad, and absolute? Am. J. Physiol. Heart Circ. Physiol. 2016, 310, H137–H152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domińska, K.; Kowalska, K.; Habrowska-Górczyńska, D.E.; Urbanek, K.A.; Ochędalski, T.; Piastowska-Ciesielska, A.W. The opposite effects of angiotensin 1–9 and angiotensin 3–7 in prostate epithelial cells. Biochem. Biophys. Res. Commun. 2019, 519, 868–873. [Google Scholar] [CrossRef] [PubMed]
- Domińska, K.; Kowalska, K.; Urbanek, K.A.; Habrowska-Górczyńska, D.E.; Ochędalski, T.; Piastowska-Ciesielska, A.W. The Impact of Ang-(1–9) and Ang-(3–7) on the Biological Properties of Prostate Cancer Cells by Modulation of Inflammatory and Steroidogenesis Pathway Genes. Int. J. Mol. Sci. 2020, 21, 6227. [Google Scholar] [CrossRef]
- Domińska, K.; Okła, P.; Kowalska, K.; Habrowska-Górczyńska, D.; Urbanek, K.A.; Ochędalski, T.; Piastowska-Ciesielska, A.W. Angiotensin 1–7 modulates molecular and cellular processes central to the pathogenesis of prostate cancer. Sci. Rep. 2018, 8, 15772. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.S.; Hsu, P.Y.; Lai, M.D.; Chang, H.Y.; Ho, C.L.; Cheng, H.L.; Chen, H.T.; Lin, Y.J.; Wu, T.J.; Tzai, T.S.; et al. An unusual function of RON receptor tyrosine kinase as a transcriptional regulator in cooperation with EGFR in human cancer cells. Carcinogenesis 2010, 31, 1456–1464. [Google Scholar] [CrossRef] [Green Version]
- Sobecki, M.; Mrouj, K.; Colinge, J.; Gerbe, F.; Jay, P.; Krasinska, L.; Dulic, V.; Fisher, D. Cell-Cycle Regulation Accounts for Variability in Ki-67 Expression Levels. Cancer Res. 2017, 77, 2722–2734. [Google Scholar] [CrossRef] [Green Version]
- Miller, I.; Min, M.; Yang, C.; Tian, C.; Gookin, S.; Carter, D.; Spencer, S.L. Ki67 is a Graded Rather than a Binary Marker of Proliferation versus Quiescence. Cell Rep. 2018, 24, 1105–1112.e5. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Kaufman, P.D. Ki-67: More than a proliferation marker. Chromosoma 2018, 127, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Lim, W.; Song, G. Inhibitory effects of delphinidin on the proliferation of ovarian cancer cells via PI3K/AKT and ERK 1/2 MAPK signal transduction. Oncol. Lett. 2017, 14, 810–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kielbik, M.; Krzyzanowski, D.; Pawlik, B.; Klink, M. Cisplatin-induced ERK1/2 activity promotes G1 to S phase progression which leads to chemoresistance of ovarian cancer cells. Oncotarget 2018, 9, 19847–19860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, S.; Yang, Y.; Song, R.; Yang, X.; Liu, H.; Ma, Q.; Yang, L.; Meng, R.; Tao, T.; Wang, S.; et al. Ang-(1-7) promotes the migration and invasion of human renal cell carcinoma cells via Mas-mediated AKT signaling. Biochem. Biophys. Res. Commun. 2015, 460, 333–340. [Google Scholar] [CrossRef]
- Hoffmann, B.R.; Stodola, T.J.; Wagner, J.R.; Didier, D.N.; Exner, E.C.; Lombard, J.H.; Greene, A.S. Mechanisms of Mas1 Receptor-Mediated Signaling in the Vascular Endothelium. Arter. Thromb. Vasc. Biol. 2017, 37, 433–445. [Google Scholar] [CrossRef] [Green Version]
- Domińska, K.; Okła, P.; Kowalska, K.; Habrowska-Górczyńska, D.; Urbanek, K.A.; Ochędalski, T.; Piastowska-Ciesielska, A.W. Influence and mechanism of Angiotensin 1–7 on biological properties of normal prostate epithelial cells. Biochem. Biophys. Res. Commun. 2018, 502, 152–159. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene/Protein | Ang-(1-7) | Ang-(1-9) | Ang-(3-7) |
|---|---|---|---|
| AGTR1 (AT1R) | 0.38 (±0.04) ↓ * | 1.26 (±0.17) | 1.7 (±0.3) |
| AGT2R (AT2R) | 0.74 (±0.16) | 1.4 (±0.32) | 2.1 (±0.58) ↑ * |
| MAS1 (MAS1R) | 0.65 (±0.08) ↓ * | 1.7 (±0.32) | 1.9 (±0.33) ↑ * |
| AGT4R (AT4R) | 1.00 (±0.07) | 1.06 (±0.05) | 1.18 (±0.06) |
| AGTR1/AGTR2 | 0.55 (±0.05) ↓ * | 0.98 (±0.05) | 0.80 (±0.06) ↓ * |
| AR (AR) | 0.64 (±0.04) ↓ * | 1.40 (±0.18) | 1.16 (±0.12) |
| ESR1 (ERα) | 1.13 (±0.2) | 1.02 (±0.18) | 1.11 (±0.20) |
| ESR2 (ERβ) | 0.84 (±0.16) | 2.56 (±0.81) | 2.91 (±0.97) ↑ * |
| ESR1/ESR2 | 1.32 (±0.19) | 0.38 (±0.11) ↓ * | 0.38 (±0.13) ↓ * |
| PGR (PRβ) | 0.37 (±0.09) | 2.75 (±0.80) | 1.43 (±0.58) |
| BIRC5 (Survivin) | 1.2 (±0.08) | 1.2 (±0.06) | 1.08 (±0.05) |
| BCL2 (Bcl-2) | 1.11 (±0.12) | 1.8 (±0.43) | 3.12 (±0.70) ↑ * |
| BAX (Bax) | 1.07 (±0.05) | 1.03 (±0.04) | 1.15 (±0.06) |
| BCL2/BAX | 1.05 (±0.11) | 1.7 (±0.43) | 2.74 (±0.61) ↑ * |
| MKI67 (Ki-67) | 1.12 (±0.05) | 1.04 (±0.05) | 1.07 (±0.04) |
| NFKB1 (NF-κB1) | 0.85 (±0.06) ↓ * | 0.90 (±0.06) | 0.92 (±0.05) |
| NFKB2 (NF-κB2) | 1.19 (±0.03) | 1.03 (±0.04) | 1.11 (±0.06) |
| NFKB1/NFKB2 | 0.70 (±0.02) ↓ * | 0.86 (±0.02) ↓ * | 0.83 (±0.03) ↓ * |
| RELA (RelA) | 1.11 (±0.05) | 1.06 (±0.01) | 1.05 (±0.08) |
| RELB (RelB) | 1.65 (±0.06) ↑ * | 1.56 (±0.06) | 1.57 (±0.15) |
| REL (c-Rel) | 0.99 (±0.04) | 0.91 (±0.02) | 0.93 (±0.04) |
| VIM (Vimetin) | 1.2 (±0.16) | 0.8 (±0.06) | 0.8 (±0.05) |
| HIF1A (Hif1a) | 0.99 (±0.12) | 0.95 (±0.01) | 0.95 (±0.04) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Domińska, K.; Urbanek, K.A.; Kowalska, K.; Habrowska-Górczyńska, D.E.; Kozieł, M.J.; Ochędalski, T.; Piastowska-Ciesielska, A.W. The Influence of Angiotensin Peptides on Survival and Motility of Human High-Grade Serous Ovarian Cancer Cells in Serum Starvation Conditions. Int. J. Mol. Sci. 2022, 23, 52. https://doi.org/10.3390/ijms23010052
Domińska K, Urbanek KA, Kowalska K, Habrowska-Górczyńska DE, Kozieł MJ, Ochędalski T, Piastowska-Ciesielska AW. The Influence of Angiotensin Peptides on Survival and Motility of Human High-Grade Serous Ovarian Cancer Cells in Serum Starvation Conditions. International Journal of Molecular Sciences. 2022; 23(1):52. https://doi.org/10.3390/ijms23010052
Chicago/Turabian StyleDomińska, Kamila, Kinga Anna Urbanek, Karolina Kowalska, Dominika Ewa Habrowska-Górczyńska, Marta Justyna Kozieł, Tomasz Ochędalski, and Agnieszka Wanda Piastowska-Ciesielska. 2022. "The Influence of Angiotensin Peptides on Survival and Motility of Human High-Grade Serous Ovarian Cancer Cells in Serum Starvation Conditions" International Journal of Molecular Sciences 23, no. 1: 52. https://doi.org/10.3390/ijms23010052
APA StyleDomińska, K., Urbanek, K. A., Kowalska, K., Habrowska-Górczyńska, D. E., Kozieł, M. J., Ochędalski, T., & Piastowska-Ciesielska, A. W. (2022). The Influence of Angiotensin Peptides on Survival and Motility of Human High-Grade Serous Ovarian Cancer Cells in Serum Starvation Conditions. International Journal of Molecular Sciences, 23(1), 52. https://doi.org/10.3390/ijms23010052