
Mycobacterium tuberculosis H37Rv Strain Increases the Frequency of CD3+TCR+ Macrophages and Affects Their Phenotype, but Not Their Migration Ability

 , , , ,
, , , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
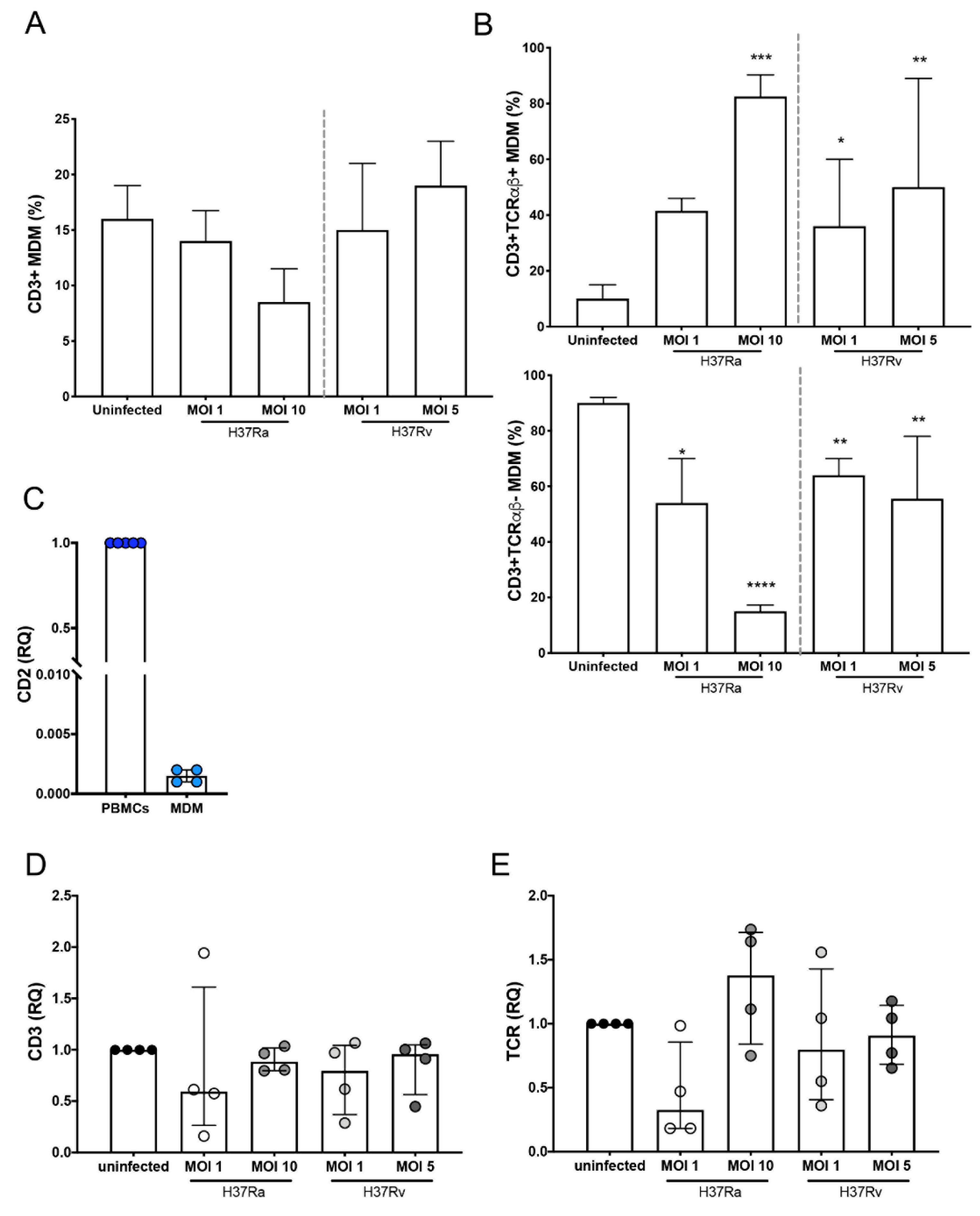
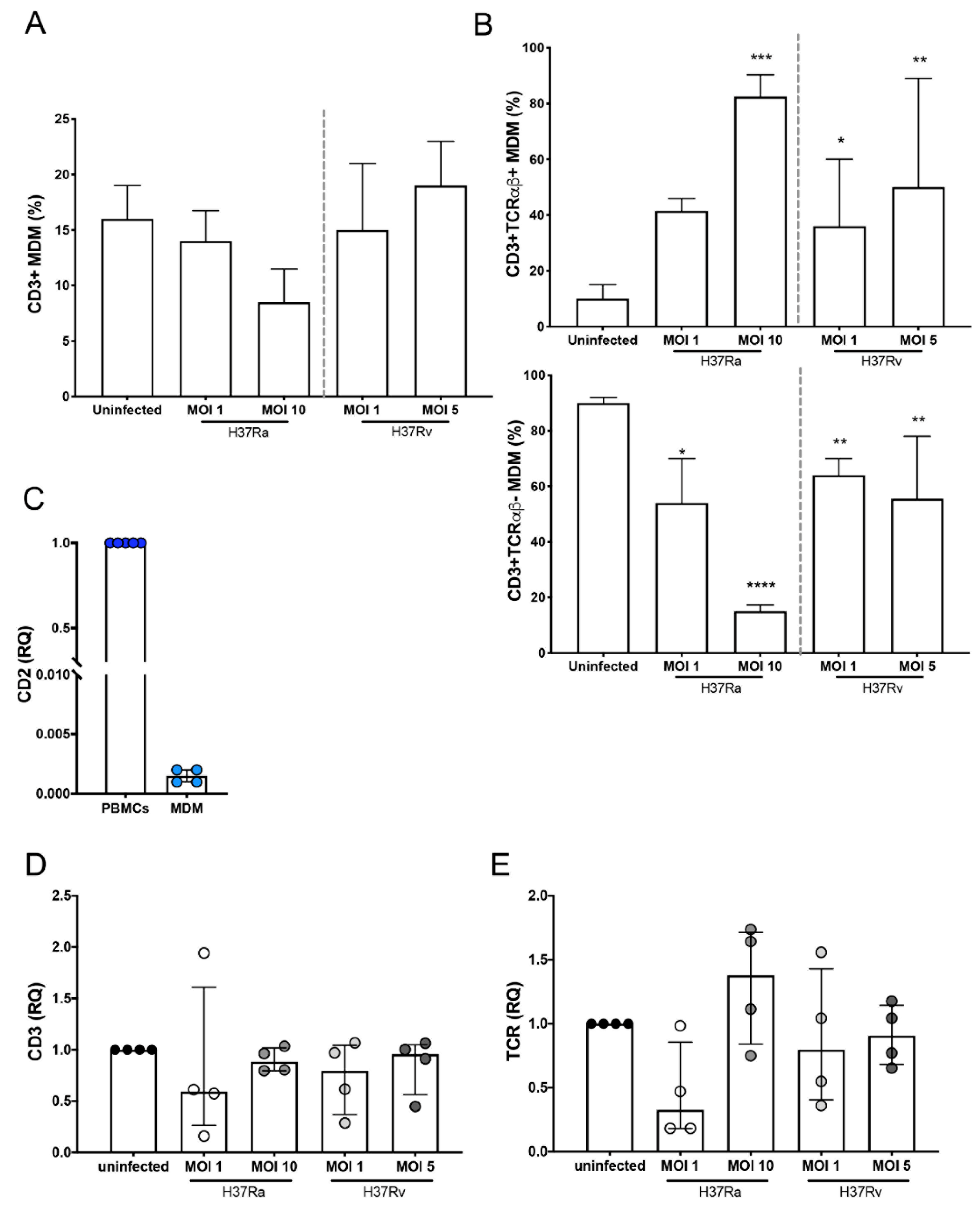
2.1. Mtb H37Rv Virulent Strain Increases the Expression of TCR on the CD3+ MDMs
2.2. Antigen-Presenting Molecules Are Differentially Modified on the Surface of CD3+TCRαβ− and CD3+TCRαβ+ MDMs
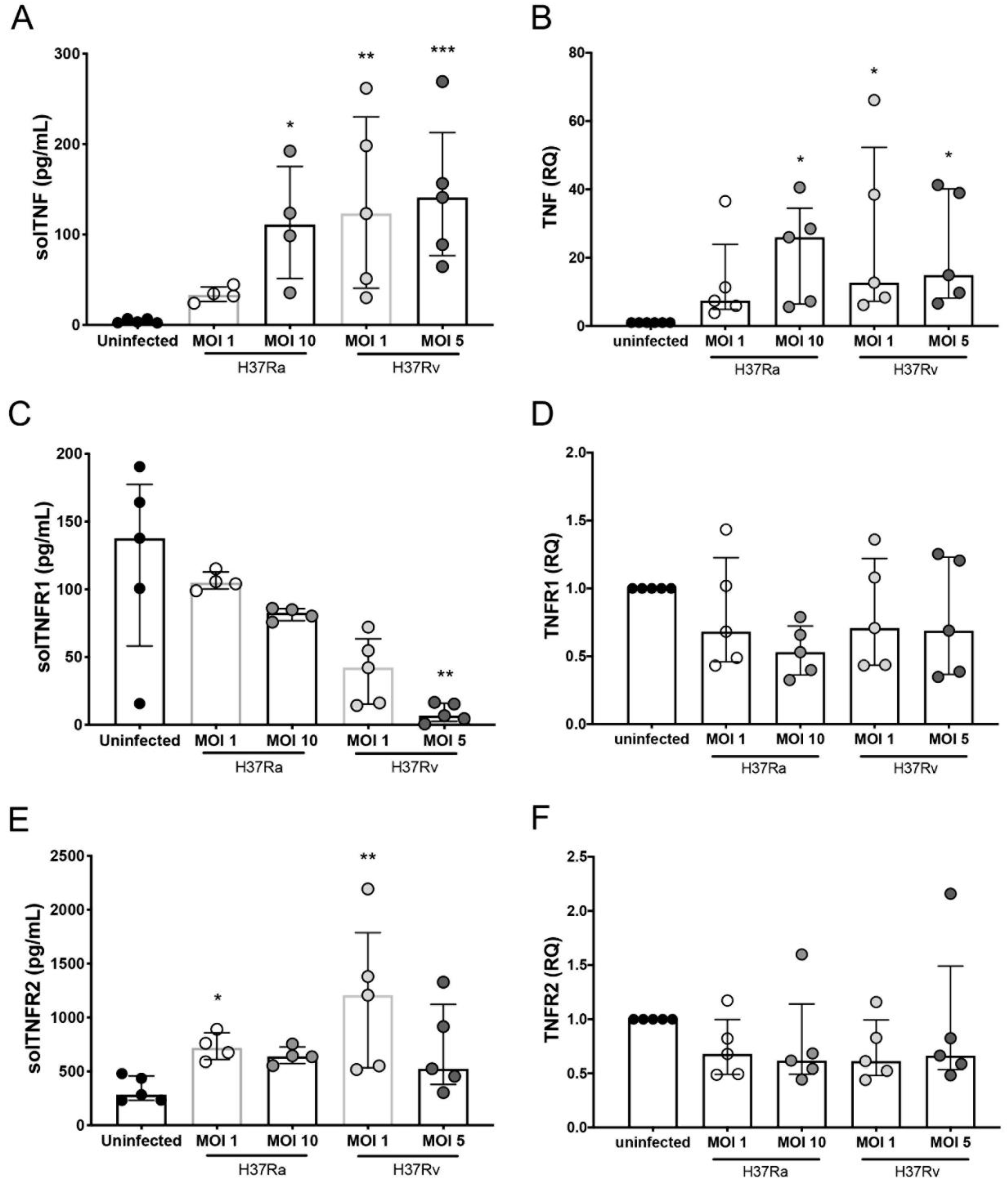
2.3. H37Rv Virulent Mtb Strain Decreases the Expression of tmTNF, and H37Ra Avirulent Mtb Strain Increases the Expression of tmTNF and tmTNFR1 on CD3+TCRαβ− and CD3+TCRαβ+ MDMs
2.4. Infection with the Virulent Mtb Strain Increases the Level of solTNF and solTNFR2, but Not solTNFR1, in MDM Subpopulations That Favor a Proinflammatory Microenvironmen
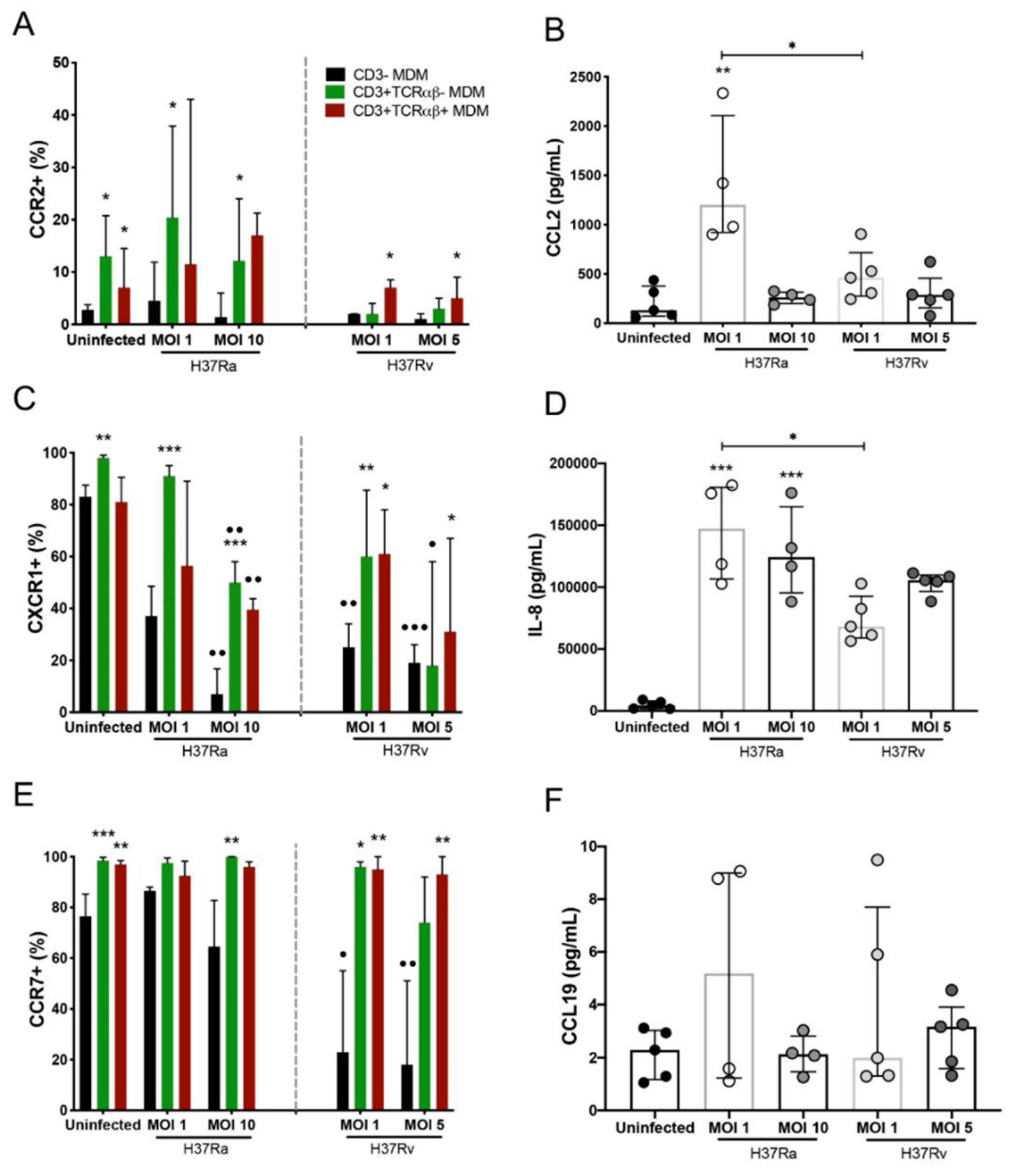
2.5. Virulence of Mtb Affects the Expression of Chemokine Receptors on CD3− and CD3+ MDM Subpopulations
2.6. Virulence of Mtb affects the Migration Capacity of MDMs, but the CD3+ MDM Subpopulation Maintains Its Migration Ability
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Enrichment of CD14+ Cells and Generation of MDMs
4.3. In Vitro MDM-Mtb Infection Assays
4.4. Assay of Cell Surface Markers by Flow Cytometry
4.5. Analysis of Gene Expression by Quantitative Real-Time PCR
4.6. Quantitation of Molecules by ELISA
4.7. Transwell Cell Migration Assay
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization (WHO). Global Tuberculosis Report. 2021. Available online: https://www.who.int/teams/global-tuberculosis-programme/tb-reports/global-tuberculosis-report-2021 (accessed on 28 November 2021).
- Cho, S.J.; Stout-Delgado, H.W. Aging and Lung Disease. Annu. Rev. Physiol. 2020, 82, 433–459. [Google Scholar] [CrossRef] [Green Version]
- Sica, A.; Mantovani, A. Macrophage Plasticity and Polarization: In Vivo Veritas. J. Clin. Investig. 2012, 122, 787–795. [Google Scholar] [CrossRef]
- Mantovani, S. Mphage_M1-M2_rev_JCI2012. J. Clin. Investig. 2012, 122, 787–795. [Google Scholar] [CrossRef]
- Beham, A.W.; Puellmann, K.; Laird, R.; Fuchs, T.; Streich, R.; Breysach, C.; Raddatz, D.; Oniga, S.; Peccerella, T.; Findeisen, P.; et al. A TNF-Regulated Recombinatorial Macrophage Immune Receptor Implicated in Granuloma Formation in Tuberculosis. PLoS Pathog. 2011, 7, e1002375. [Google Scholar] [CrossRef]
- Ruiz, A.; Palacios, Y.; Garcia, I.; Chavez-Galan, L. Transmembrane TNF and Its Receptors TNFR1 and TNFR2 in Mycobacterial Infections. Int. J. Mol. Sci. 2021, 22, 5461. [Google Scholar] [CrossRef]
- Rodriguez-Cruz, A.; Vesin, D.; Ramon-Luing, L.; Zuñiga, J.; Quesniaux, V.F.J.; Ryffel, B.; Lascurain, R.; Garcia, I.; Chávez-Galán, L. CD3+ Macrophages Deliver Proinflammatory Cytokines by a CD3- and Transmembrane TNF-Dependent Pathway and Are Increased at the BCG-Infection Site. Front. Immunol. 2019, 10, 2550. [Google Scholar] [CrossRef]
- Chavez-Galan, L.; Vesin, D.; Blaser, G.; Uysal, H.; Benmerzoug, S.; Rose, S.; Ryffel, B.; Quesniaux, V.F.J.; Garcia, I. Myeloid Cell TNFR1 Signaling Dependent Liver Injury and Inflammation upon BCG Infection. Sci. Rep. 2019, 9, 5297. [Google Scholar] [CrossRef]
- Ocaña-Guzmán, R.; Téllez-Navarrete, N.A.; Ramón-Luing, L.A.; Herrera, I.; de Ita, M.; Carrillo-Alduenda, J.L.; Choreño-Parra, J.A.; Medina-Quero, K.; Zúñiga, J.; Chávez-Galán, L. Leukocytes from Patients with Drug-Sensitive and Multidrug-Resistant Tuberculosis Exhibit Distinctive Profiles of Chemokine Receptor Expression and Migration Capacity. J. Immunol. Res. 2021, 2021, 6654220. [Google Scholar] [CrossRef]
- Ordway, D.; Henao-Tamayo, M.; Harton, M.; Palanisamy, G.; Troudt, J.; Shanley, C.; Basaraba, R.J.; Orme, I.M. The Hypervirulent Mycobacterium Tuberculosis Strain HN878 Induces a Potent TH1 Response Followed by Rapid Down-Regulation. J. Immunol. 2007, 179, 522–531. [Google Scholar] [CrossRef] [Green Version]
- Park, H.-S.; Back, Y.W.; Jang, I.-T.; Lee, K.-I.; Son, Y.-J.; Choi, H.-G.; Dang, T.B.; Kim, H.-J. Mycobacterium Tuberculosis Rv2145c Promotes Intracellular Survival by STAT3 and IL-10 Receptor Signaling. Front. Immunol. 2021, 12, 6662931. [Google Scholar] [CrossRef]
- Refai, A.; Gritli, S.; Barbouche, M.R.; Essafi, M. Mycobacterium Tuberculosis Virulent Factor ESAT-6 Drives Macrophage Differentiation toward the pro-Inflammatory M1 Phenotype and Subsequently Switches It to the Anti-Inflammatory M2 Phenotype. Front. Cell. Infect. Microbiol. 2018, 8, 327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, S.; Hong, Z.; Zhang, G.; Li, J.; Tian, G.B.; Zhou, H.; Huang, X. Mycobacterium PPE31 Contributes to Host Cell Death. Front. Cell. Infect. Microbiol. 2021, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Moody, D.B.; Guy, M.R.; Grant, E.; Cheng, T.Y.; Brenner, M.B.; Besra, G.S.; Porcelli, S.A. CD1b-Mediated T Cell Recognition of a Glycolipid Antigen Generated from Mycobacterial Lipid and Host Carbohydrate during Infection. J. Exp. Med. 2000, 192, 965–976. [Google Scholar] [CrossRef] [PubMed]
- de Libero, G.; Mori, L. The T-Cell Response to Lipid Antigens of Mycobacterium Tuberculosis. Front. Immunol. 2014, 5, 219. [Google Scholar] [CrossRef] [Green Version]
- Flynn, J.L.; Chan, J. Immunology of Tuberculosis. Annu. Rev. Immunol. 2001, 19, 93–129. [Google Scholar] [CrossRef]
- Domingo-Gonzalez, R.; Prince, O.; Cooper, A.; Khader, S.A. Cytokines and Chemokines in Mycobacterium Tuberculosis Infection. Microbiol. Spectr. 2016, 4, 4–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Méndez-Samperio, P. Expression and Regulation of Chemokines in Mycobacterial Infection. J. Infect. 2008, 57, 374–384. [Google Scholar] [CrossRef]
- Fuchs, T.; Puellmann, K.; Hahn, M.; Dollt, C.; Pechlivanidou, I.; Ovsiy, I.; Kzhyshkowska, J.; Gratchev, A.; Fleig, J.; Emmert, A.; et al. A Second Combinatorial Immune Receptor in Monocytes/Macrophages Is Based on the TCRγδ. Immunobiology 2013, 218, 960–968. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, T.; Puellmann, K.; Emmert, A.; Fleig, J.; Oniga, S.; Laird, R.; Heida, N.M.; Schäfer, K.; Neumaier, M.; Beham, A.W.; et al. The Macrophage-TCRαβ Is a Cholesterol-Responsive Combinatorial Immune Receptor and Implicated in Atherosclerosis. Biochem. Biophys. Res. Commun. 2015, 456, 59–65. [Google Scholar] [CrossRef] [PubMed]
- McClean, C.M.; Tobin, D.M. Macrophage Form, Function, and Phenotype in Mycobacterial Infection: Lessons from Tuberculosis and Other Diseases. Pathog. Dis. 2016, 74, ftw068. [Google Scholar] [CrossRef] [Green Version]
- Lopez, K.; Iwany, S.K.; Suliman, S.; Reijneveld, J.F.; Ocampo, T.A.; Jimenez, J.; Calderon, R.; Lecca, L.; Murray, M.B.; Moody, D.B.; et al. CD1b Tetramers Broadly Detect T Cells That Correlate with Mycobacterial Exposure but Not Tuberculosis Disease State. Front. Immunol. 2020, 11, 199. [Google Scholar] [CrossRef]
- Layton, E.D.; Yu, K.K.Q.; Smith, M.T.; Scriba, T.J.; de Rosa, S.C.; Seshadri, C. Validation of a CD1b Tetramer Assay for Studies of Human Mycobacterial Infection or Vaccination. J. Immunol. Methods 2018, 458, 44–52. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, R.; Jiang, J.; Cao, Z.; Zhai, F.; Sun, W.; Cheng, X. A Subset of CD1c + Dendritic Cells Is Increased in Patients with Tuberculosis and Promotes Th17 cell Polarization. Tuberculosis 2018, 113, 189–199. [Google Scholar] [CrossRef]
- To, K.W.; Reino, J.J.G.; Yoo, D.H.; Tam, L.S. Tumour Necrosis Factor Antagonist and Tuberculosis in Patients with Rheumatoid Arthritis: An Asian Perspective. Respirology 2013, 18, 765–773. [Google Scholar] [CrossRef]
- Keeton, R.; Allie, N.; Dambuza, I.; Abel, B.; Hsu, N.J.; Sebesho, B.; Randall, P.; Burger, P.; Fick, E.; Quesniaux, V.F.J.; et al. Soluble TNFRp75 Regulates Host Protective Immunity against Mycobacterium Tuberculosis. J. Clin. Investig. 2014, 124, 1537–1551. [Google Scholar] [CrossRef]
- Cooper, A.M.; Solache, A.; Khader, S.A. Interleukin-12 and Tuberculosis: An Old Story Revisited. Curr. Opin. Immunol. 2007, 19, 441–447. [Google Scholar] [CrossRef] [Green Version]
- Juffermans, N.P.; Dekkers, P.E.P.; Peppelenbosch, M.P.; Speelman, P.; van Deventer, S.J.H.; van der Poll, T. Expression of the Chemokine Receptors CXCR1 and CXCR2 on Granulocytes in Human Endotoxemia and Tuberculosis: Involvement of the P38 Mitogen-Activated Protein Kinase Pathway. J. Infect. Dis. 2000, 182, 888–894. [Google Scholar] [CrossRef]
- Cui, K.; Ardell, C.L.; Podolnikova, N.P.; Yakubenko, V.P. Distinct Migratory Properties of M1, M2, and Resident Macrophages Are Regulated by Adβ2and Amβ2integrin-Mediated Adhesion. Front. Immunol. 2018, 9, 2650. [Google Scholar] [CrossRef]
- Chávez-Galán, L.; Ocaña-Guzmán, R.; Torre-Bouscoulet, L.; García-De-Alba, C.; Sada-Ovalle, I. Exposure of Monocytes to Lipoarabinomannan Promotes Their Differentiation into Functionally and Phenotypically Immature Macrophages. J. Immunol. Res. 2015, 2015, 984973. [Google Scholar] [CrossRef] [Green Version]
- Carranza, C.; Juárez, E.; Torres, M.; Ellner, J.J.; Sada, E.; Schwander, S.K. Mycobacterium Tuberculosis Growth Control by Lung Macrophages and CD8 Cells from Patient Contacts. Am. J. Respir. Crit. Care Med. 2006, 173, 238–245. [Google Scholar] [CrossRef] [Green Version]
- Al-haidari, A.A.; Syk, I.; Jirström, K.; Thorlacius, H. CCR4 Mediates CCL17 (TARC)-Induced Migration of Human Colon Cancer Cells via RhoA/Rho-Kinase Signaling. Int. J. Colorectal Dis. 2013, 28, 1479–1487. [Google Scholar] [CrossRef] [Green Version]
- Campanella, G.S.v.; Luster, A.D. A Chemokine-Mediated In Vivo T-Cell Recruitment Assay Gabriele. Methods Enzymol. 2009, 461, 397–412. [Google Scholar] [CrossRef] [PubMed]
- Carr, M.W.; Roth, S.J.; Luther, E.; Rose, S.S.; Springer, T.A. Monocyte Chemoattractant Protein 1 Acts as a T-Lymphocyte Chemoattractant. Proc. Natl. Acad. Sci. USA 1994, 91, 3652–3656. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramon-Luing, L.A.; Carranza, C.; Téllez-Navarrete, N.A.; Medina-Quero, K.; Gonzalez, Y.; Torres, M.; Chavez-Galan, L. Mycobacterium tuberculosis H37Rv Strain Increases the Frequency of CD3+TCR+ Macrophages and Affects Their Phenotype, but Not Their Migration Ability. Int. J. Mol. Sci. 2022, 23, 329. https://doi.org/10.3390/ijms23010329
Ramon-Luing LA, Carranza C, Téllez-Navarrete NA, Medina-Quero K, Gonzalez Y, Torres M, Chavez-Galan L. Mycobacterium tuberculosis H37Rv Strain Increases the Frequency of CD3+TCR+ Macrophages and Affects Their Phenotype, but Not Their Migration Ability. International Journal of Molecular Sciences. 2022; 23(1):329. https://doi.org/10.3390/ijms23010329
Chicago/Turabian StyleRamon-Luing, Lucero A., Claudia Carranza, Norma A. Téllez-Navarrete, Karen Medina-Quero, Yolanda Gonzalez, Martha Torres, and Leslie Chavez-Galan. 2022. "Mycobacterium tuberculosis H37Rv Strain Increases the Frequency of CD3+TCR+ Macrophages and Affects Their Phenotype, but Not Their Migration Ability" International Journal of Molecular Sciences 23, no. 1: 329. https://doi.org/10.3390/ijms23010329
APA StyleRamon-Luing, L. A., Carranza, C., Téllez-Navarrete, N. A., Medina-Quero, K., Gonzalez, Y., Torres, M., & Chavez-Galan, L. (2022). Mycobacterium tuberculosis H37Rv Strain Increases the Frequency of CD3+TCR+ Macrophages and Affects Their Phenotype, but Not Their Migration Ability. International Journal of Molecular Sciences, 23(1), 329. https://doi.org/10.3390/ijms23010329