
Cord Blood T Cells Expressing High and Low PKCζ Levels Develop into Cells with a Propensity to Display Th1 and Th9 Cytokine Profiles, Respectively

 , ,
, ,
Abstract
1. Introduction
2. Results
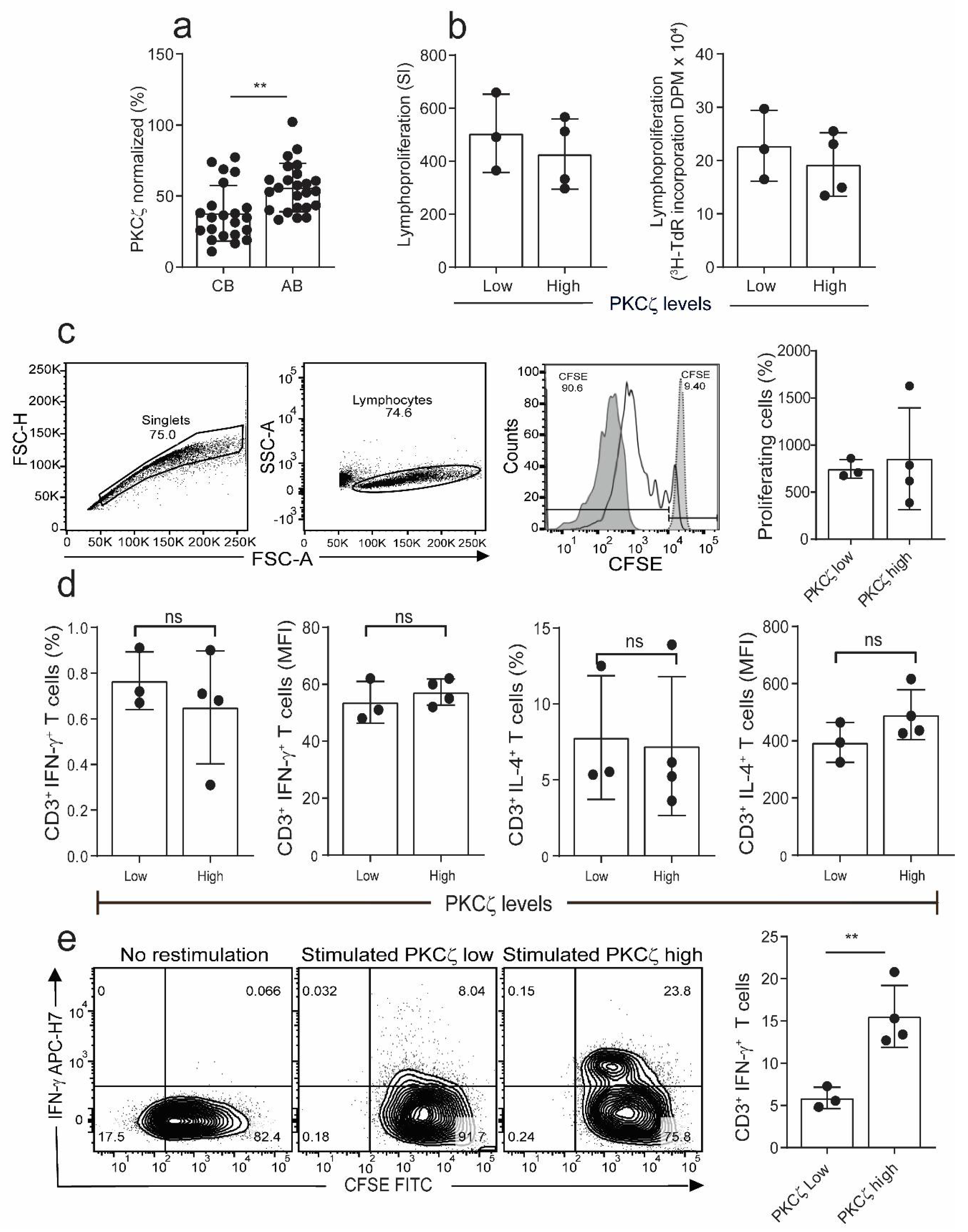
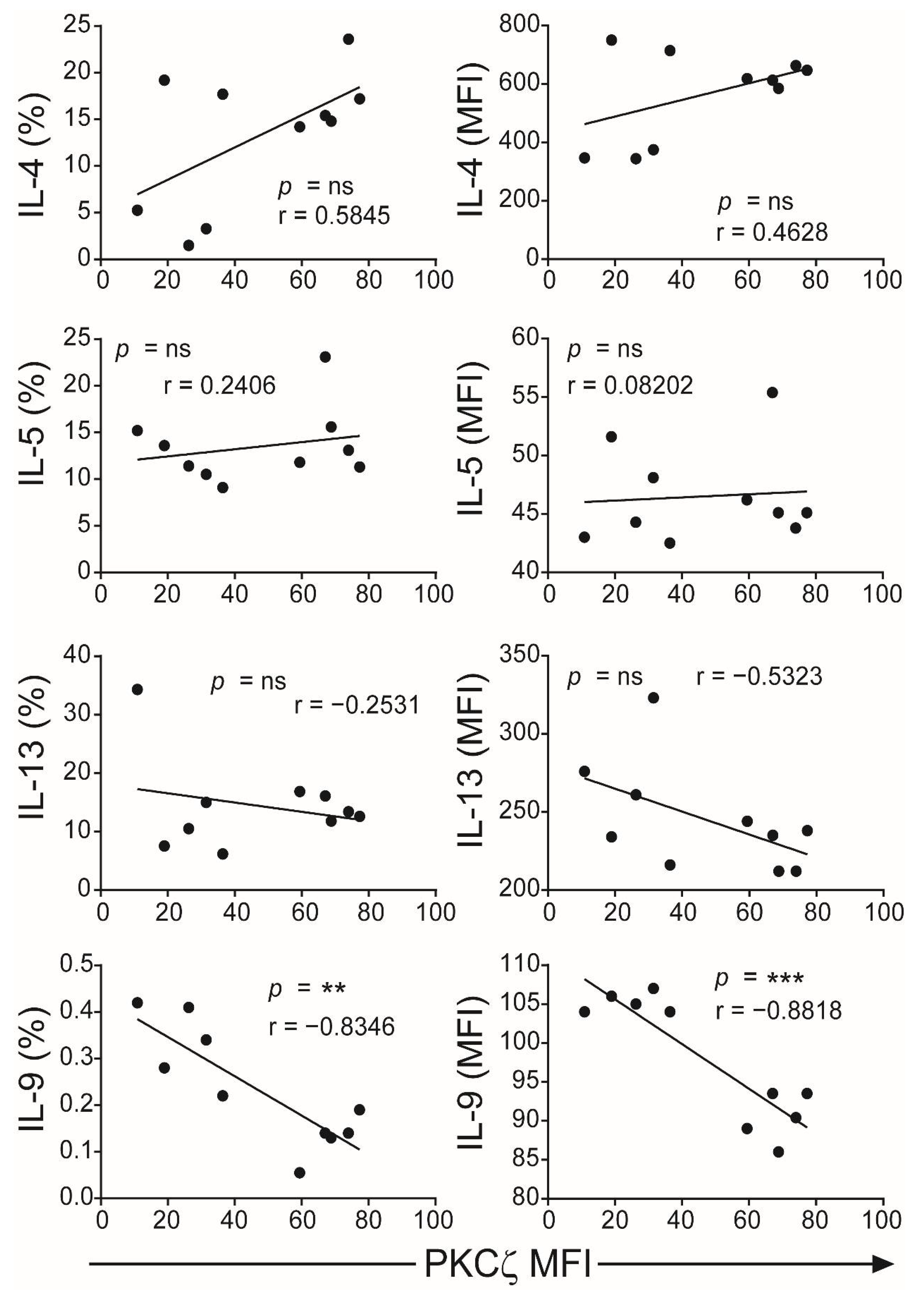
2.1. The Levels of PKCζ in Immature CBTC Does Not Affect Cytokine Pattern Expressed and Lymphoproliferation
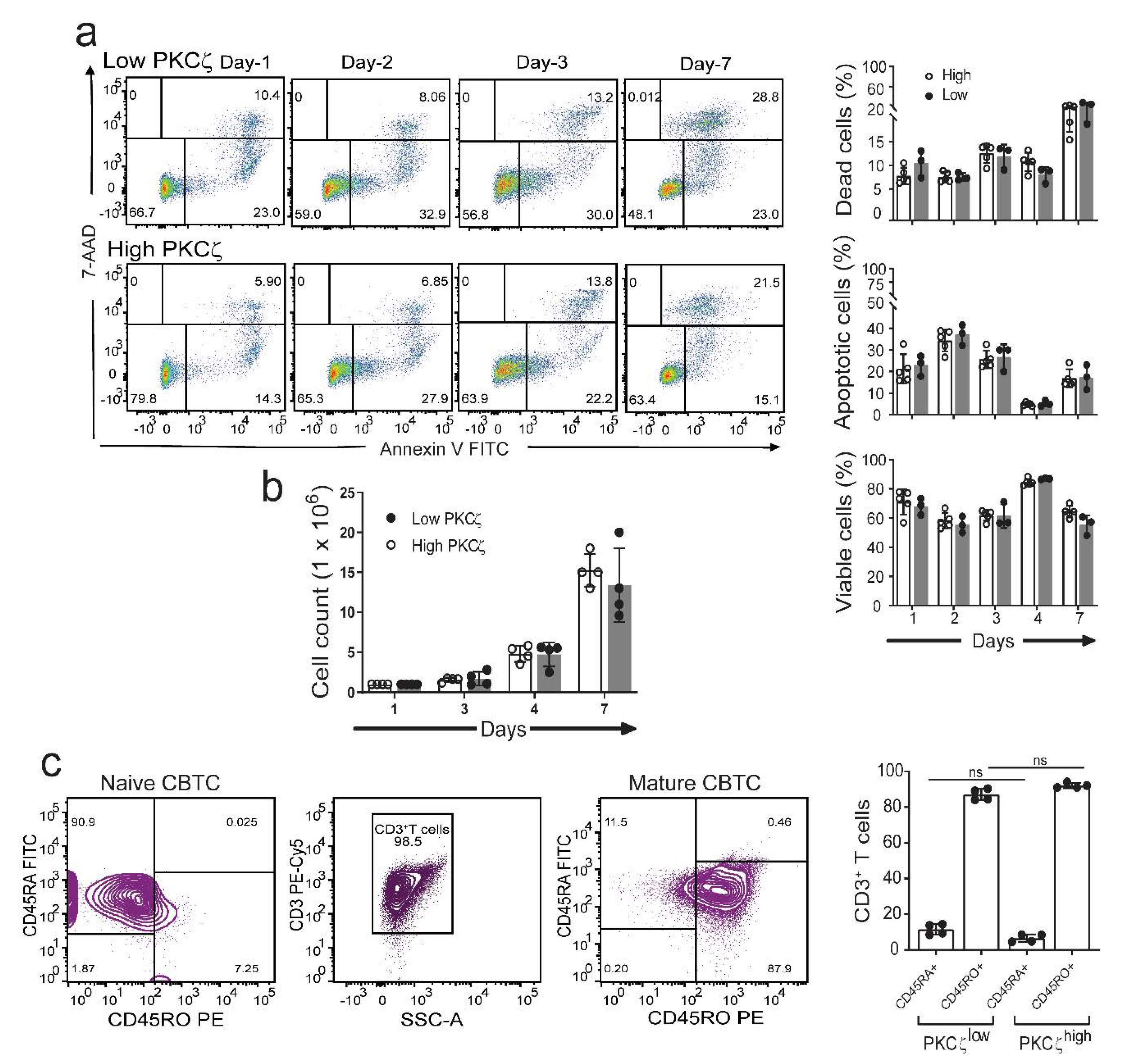
2.2. CBTC PKCζ Levels and T Cell Survival during Maturation
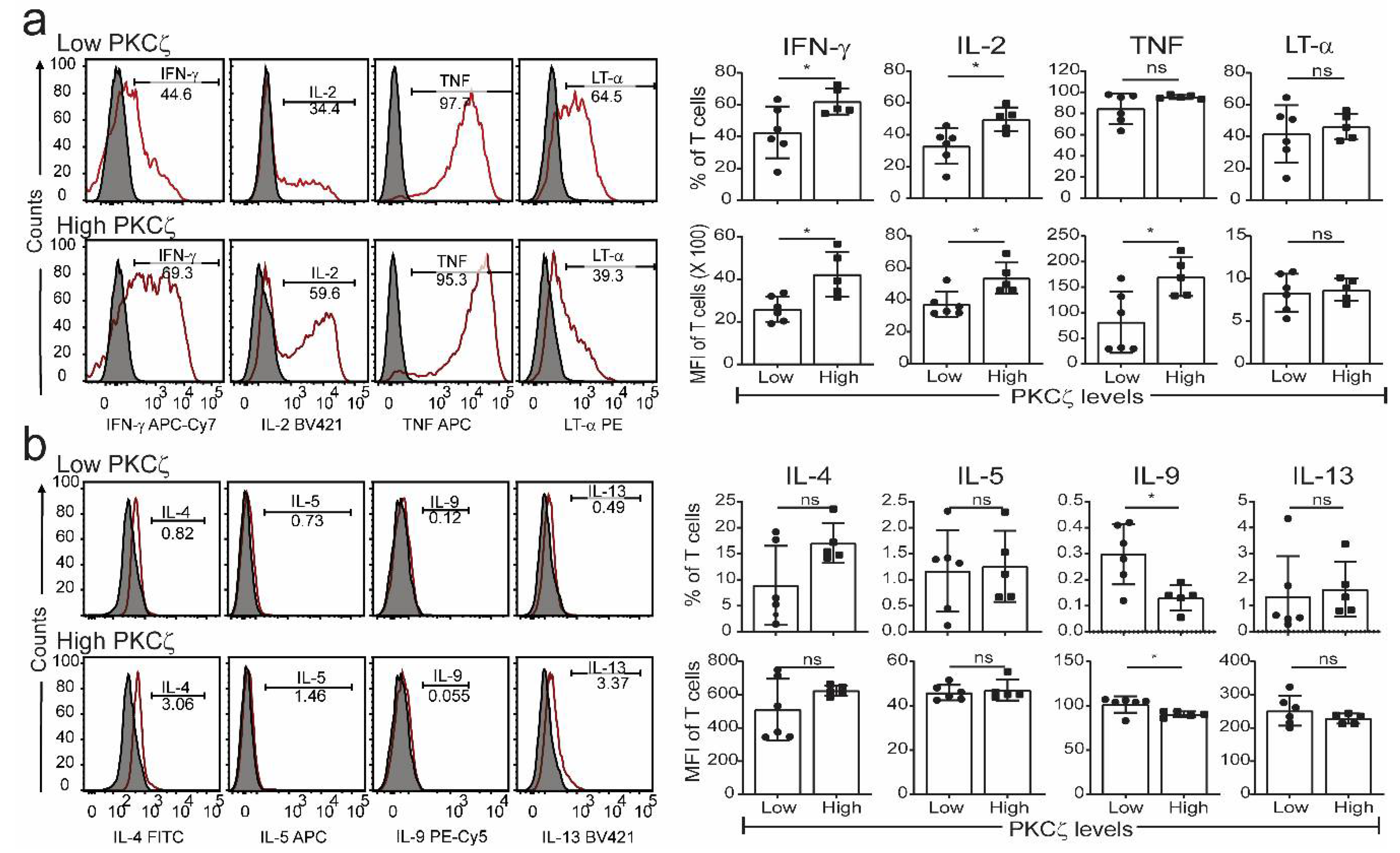
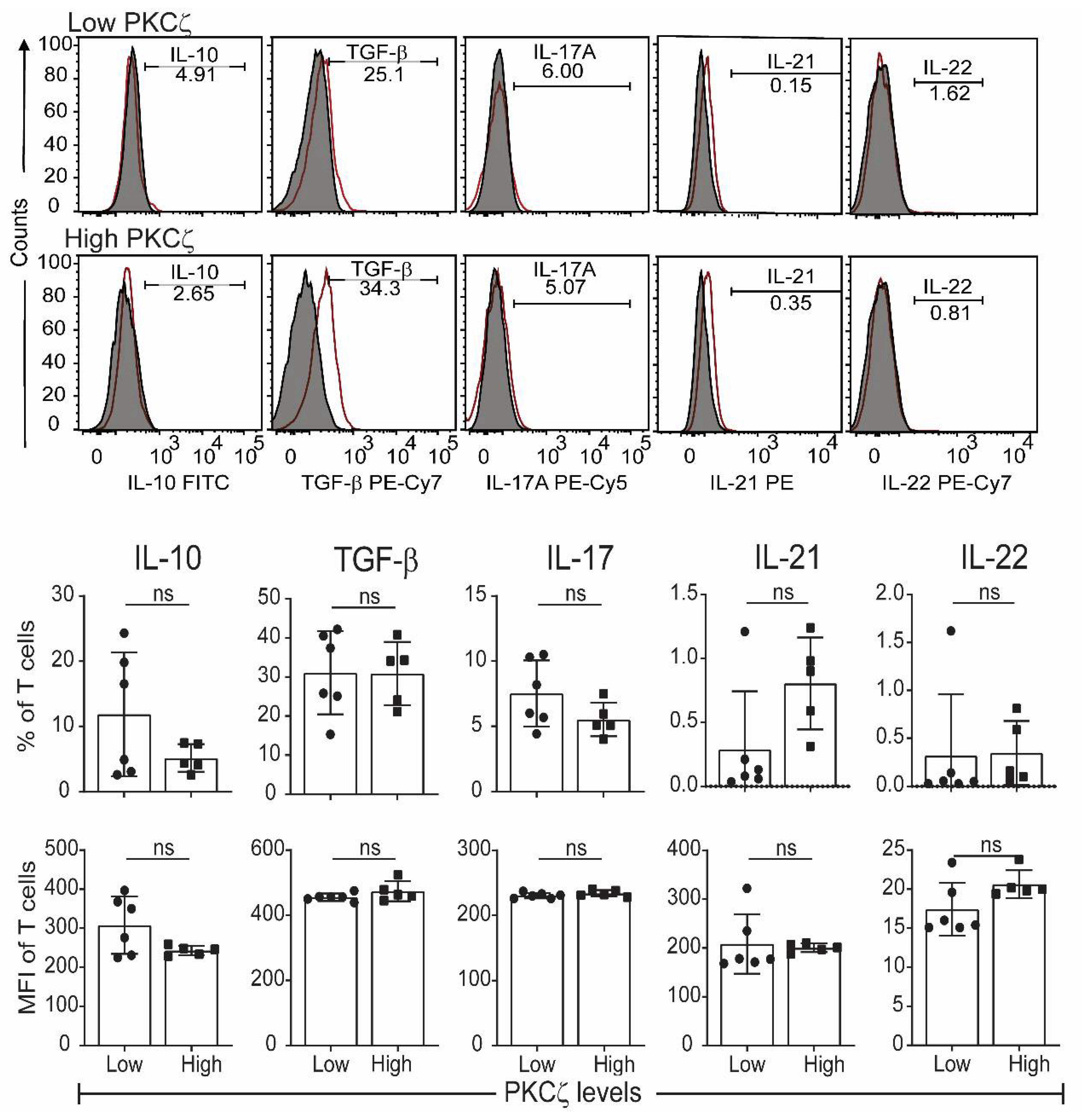
2.3. Low PKCζ Expression Is Associated with Decreased Development towards a Th1 Cytokine Profile
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Ethics Statement
4.3. Preparation of Cord Blood and Peripheral Blood Mononuclear Cells
4.4. Isolation of T Cells
4.5. CBTC Maturation
4.6. Apoptosis and Cell Viability Assays
4.7. T Cell Proliferation Assays
4.8. T Cell Maturation
4.9. Detection of Intracellular Cytokines
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CBTC | cord blood T cells |
| CBMC | CB mononuclear cells |
| PBMC | peripheral blood MC |
| AB | adult blood |
| PKC | Protein Kinase C |
| PHA | Phytohaemagglutinin |
| PMA | Phorbol myristate acetate |
| IFN-γ | Interferon-gamma |
| rhIL-2 | recombinant human interleukin 2 |
| TNF | tumour necrosis factor-alpha |
| LT-α | Lymphotoxin α |
| TGF-β | Transforming growth factor-beta |
| CFSE | carboxyfluorescein succinimidyl ester |
| MFI | Median Fluorescence Intensity |
| [3H]-TdR | tritiated thymidine |
| 7-AAD | 7-aminoactinomycin D |
| IgE | Immunoglobulin E |
| FcεR1 | High-affinity IgE receptor |
| RPMI 1640 | Roswell Park Memorial Institute 1640 |
| IL-5R | IL-5 receptor |
| FCS | foetal calf serum |
References
- Prescott, S.L.; Tang, M.L. The Australasian Society of Clinical Immunology and Allergy position statement: Summary of allergy prevention in children. Med. J. Aust. 2005, 182, 464–467. [Google Scholar] [CrossRef]
- Loh, W.; Tang, M.L.K. The epidemiology of food allergy in the global context. Int. J. Environ. Res. Public Health 2018, 15, 2043. [Google Scholar] [CrossRef]
- Chu, D.K.; Mohammed-Ali, Z.; Jiménez-Saiz, R.; Walker, T.D.; Goncharova, S.; Llop-Guevara, A.; Kong, J.; Gordon, M.E.; Barra, N.G.; Gillgrass, A.E.; et al. T helper cell IL-4 drives intestinal Th2 priming to oral peanut antigen, under the control of OX40L and independent of innate-like lymphocytes. Mucosal. Immunol. 2014, 7, 1395–1404. [Google Scholar] [CrossRef]
- Angkasekwinai, P.; Dong, C. IL-9-producing T cells: Potential players in allergy and cancer. Nat. Rev. Immunol. 2021, 21, 37–48. [Google Scholar] [CrossRef]
- Goswami, R.; Kaplan, M.H. A brief history of IL-9. J. Immunol. 2011, 186, 3283–3288. [Google Scholar] [CrossRef] [PubMed]
- Angkasekwinai, P. Allergic inflammation and atopic disease: Role of Th9 cells. In Th9 Cells: Methods and Protocols; Goswami, R., Ed.; Springer: New York, NY, USA, 2017; pp. 189–199. [Google Scholar]
- Renauld, J.-C. New insights into the role of cytokines in asthma. J. Clin. Pathol. 2001, 54, 577–589. [Google Scholar] [CrossRef] [PubMed]
- Hauber, H.P.; Bergeron, C.; Hamid, Q. IL-9 in allergic inflammation. Int. Arch. Allergy Immunol. 2004, 134, 79–87. [Google Scholar] [CrossRef]
- Soussi-Gounni, A.; Kontolemos, M.; Hamid, Q. Role of IL-9 in the pathophysiology of allergic diseases. J. Allergy Clin. Immunol. 2001, 107, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Coquet, J.M. A singular role for interleukin-9 in the development of asthma. Sci. Immunol. 2020, 5. [Google Scholar] [CrossRef]
- Koch, S.; Sopel, N.; Finotto, S. Th9 and other IL-9-producing cells in allergic asthma. Semin. Immunopathol. 2017, 39, 55–68. [Google Scholar] [CrossRef]
- Jones, C.P.; Gregory, L.G.; Causton, B.; Campbell, G.A.; Lloyd, C.M. Activin A and TGF-β promote Th9 cell–mediated pulmonary allergic pathology. J. Allergy Clin. Immunol. 2012, 129, 1000–1010.e3. [Google Scholar] [CrossRef] [PubMed]
- Shik, D.; Tomar, S.; Lee, J.B.; Chen, C.Y.; Smith, A.; Wang, Y.H. IL-9-producing cells in the development of IgE-mediated food allergy. Semin. Immunopathol. 2017, 39, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Lotoski, L.C.; Chooniedass, R.; Su, R.C.; Simons, F.E.; Liem, J.; Becker, A.B.; Uzonna, J.; HayGlass, K.T. Elevated antigen-driven IL-9 responses are prominent in peanut allergic humans. PLoS ONE 2012, 7, e45377. [Google Scholar] [CrossRef] [PubMed]
- Brough, H.A.; Cousins, D.J.; Munteanu, A.; Wong, Y.F.; Sudra, A.; Makinson, K.; Stephens, A.C.; Arno, M.; Ciortuz, L.; Lack, G.; et al. IL-9 is a key component of memory Th cell peanut-specific responses from children with peanut allergy. J. Allergy Clin. Immunol. 2014, 134, 1329–1338.e10. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Wang, Y.; Li, J.; Li, S.; Zhang, Y.; Shen, J.; Tan, W.; Wu, C. Detection of IL-9 producing T cells in the PBMCs of allergic asthmatic patients. BMC Immunol. 2017, 18, 38. [Google Scholar] [CrossRef]
- Schmitt, E.; Germann, T.; Goedert, S.; Hoehn, P.; Huels, C.; Koelsch, S.; Kühn, R.; Müller, W.; Palm, N.; Rüde, E. IL-9 production of naive CD4+ T cells depends on IL-2, is synergistically enhanced by a combination of TGF-beta and IL-4, and is inhibited by IFN-gamma. J. Immunol. 1994, 153, 3989–3996. [Google Scholar] [PubMed]
- Dardalhon, V.; Awasthi, A.; Kwon, H.; Galileos, G.; Gao, W.; Sobel, R.A.; Mitsdoerffer, M.; Strom, T.B.; Elyaman, W.; Ho, I.C.; et al. IL-4 inhibits TGF-beta-induced Foxp3+ T cells and, together with TGF-beta, generates IL-9+ IL-10+ Foxp3− effector T cells. Nat. Immunol. 2008, 9, 1347–1355. [Google Scholar] [CrossRef] [PubMed]
- Veldhoen, M.; Uyttenhove, C.; van Snick, J.; Helmby, H.; Westendorf, A.; Buer, J.; Martin, B.; Wilhelm, C.; Stockinger, B. Transforming growth factor-beta ‘reprograms’ the differentiation of T helper 2 cells and promotes an interleukin 9-producing subset. Nat. Immunol. 2008, 9, 1341–1346. [Google Scholar] [CrossRef] [PubMed]
- Petit-Frere, C.; Dugas, B.; Braquet, P.; Mencia-Huerta, J. Interleukin-9 potentiates the interleukin-4-induced IgE and IgG1 release from murine B lymphocytes. Immunology 1993, 79, 146. [Google Scholar]
- Temann, U.A.; Geba, G.P.; Rankin, J.A.; Flavell, R.A. Expression of interleukin 9 in the lungs of transgenic mice causes airway inflammation, mast cell hyperplasia, and bronchial hyperresponsiveness. J. Exp. Med. 1998, 188, 1307–1320. [Google Scholar] [CrossRef]
- Louahed, J.; Toda, M.; Jen, J.; Hamid, Q.; Renauld, J.-C.; Levitt, R.C.; Nicolaides, N.C. Interleukin-9 upregulates mucus expression in the airways. Am. J. Respir. Cell Mol. Biol. 2000, 22, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Matsuzawa, S.; Sakashita, K.; Kinoshita, T.; Ito, S.; Yamashita, T.; Koike, K. IL-9 enhances the growth of human mast cell progenitors under stimulation with stem cell factor. J. Immunol. 2003, 170, 3461–3467. [Google Scholar] [CrossRef]
- Sehra, S.; Yao, W.; Nguyen, E.T.; Glosson-Byers, N.L.; Akhtar, N.; Zhou, B.; Kaplan, M.H. Th9 cells are required for tissue mast cell accumulation during allergic inflammation. J. Allergy Clin. Immunol. 2015, 136, 433–440.e1. [Google Scholar] [CrossRef]
- Louahed, J.; Kermouni, A.; Van Snick, J.; Renauld, J.-C. IL-9 induces expression of granzymes and high-affinity IgE receptor in murine T helper clones. J. Immunol. 1995, 154, 5061–5070. [Google Scholar]
- Dong, Q.; Louahed, J.; Vink, A.; Sullivan, C.D.; Messler, C.J.; Zhou, Y.; Haczku, A.; Huaux, F.; Arras, M.; Holroyd, K.J. IL-9 induces chemokine expression in lung epithelial cells and baseline airway eosinophilia in transgenic mice. Eur. J. Immunol. 1999, 29, 2130–2139. [Google Scholar] [CrossRef]
- Gounni, A.S.; Gregory, B.; Nutku, E.; Aris, F.; Latifa, K.; Minshall, E.; North, J.; Tavernier, J.; Levit, R.; Nicolaides, N. Interleukin-9 enhances interleukin-5 receptor expression, differentiation, and survival of human eosinophils. Blood J. Am. Soc. Hematol. 2000, 96, 2163–2171. [Google Scholar]
- Stone, K.D.; Prussin, C.; Metcalfe, D.D. IgE, mast cells, basophils, and eosinophils. J. Allergy Clin. Immunol. 2010, 125, S73–S80. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.K.; Herbert, C.; Yang, M.; Koskinen, A.M.; McKenzie, A.N.; Foster, P.S. Role of interleukin-13 in eosinophil accumulation and airway remodelling in a mouse model of chronic asthma. Clin. Exp. Allergy 2002, 32, 1104–1111. [Google Scholar] [CrossRef]
- Gour, N.; Wills-Karp, M. IL-4 and IL-13 signaling in allergic airway disease. Cytokine 2015, 75, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Hii, C.S.; Costabile, M.; Mayne, G.C.; Der, C.J.; Murray, A.W.; Ferrante, A. Selective deficiency in protein kinase C isoenzyme expression and inadequacy in mitogen-activated protein kinase activation in cord blood T cells. Biochem. J. 2003, 370, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Perveen, K.; Quach, A.; McPhee, A.; Prescott, S.L.; Barry, S.C.; Hii, C.S.; Ferrante, A. Validation of monoclonal antiPKC isozyme antibodies for flow cytometry analyses in human T cell subsets and expression in cord blood T cells. Sci. Rep. 2019, 9, 9263. [Google Scholar] [CrossRef] [PubMed]
- Prescott, S.L.; Irvine, J.; Dunstan, J.A.; Hii, C.; Ferrante, A. Protein kinase C zeta: A novel protective neonatal T-cell marker that can be upregulated by allergy prevention strategies. J. Allergy Clin. Immunol. 2007, 120, 200–206. [Google Scholar] [CrossRef] [PubMed]
- D’Vaz, N.; Ma, Y.; Dunstan, J.A.; Lee-Pullen, T.F.; Hii, C.; Meldrum, S.; Zhang, G.; Metcalfe, J.; Ferrante, A.; Prescott, S.L. Neonatal protein kinase C zeta expression determines the neonatal T-cell cytokine phenotype and predicts the development and severity of infant allergic disease. Allergy 2012, 67, 1511–1518. [Google Scholar] [CrossRef]
- Harb, H.; Irvine, J.; Amarasekera, M.; Hii, C.S.; Kesper, D.A.; Ma, Y.; D’Vaz, N.; Renz, H.; Potaczek, D.P.; Prescott, S.L.; et al. The role of PKCζ in cord blood T-cell maturation towards Th1 cytokine profile and its epigenetic regulation by fish oil. Biosci. Rep. 2017, 37. [Google Scholar] [CrossRef]
- Acevedo, N.; Alashkar Alhamwe, B.; Caraballo, L.; Ding, M.; Ferrante, A.; Garn, H.; Garssen, J.; Hii, C.S.; Irvine, J.; Llinás-Caballero, K.; et al. Perinatal and early-life nutrition, epigenetics, and allergy. Nutrients 2021, 13, 724. [Google Scholar] [CrossRef]
- Leroy, I.; de Thonel, A.; Laurent, G.; Quillet-Mary, A. Protein kinase C zeta associates with death inducing signaling complex and regulates Fas ligand-induced apoptosis. Cell Signal. 2005, 17, 1149–1157. [Google Scholar] [CrossRef] [PubMed]
- Rainsford, E.; Reen, D.J. Interleukin 10, produced in abundance by human newborn T cells, may be the regulator of increased tolerance associated with cord blood stem cell transplantation. Br. J. Haematol. 2002, 116, 702–709. [Google Scholar] [CrossRef]
- Ohshima, Y.; Yang, L.-P.; Avice, M.-N.; Kurimoto, M.; Nakajima, T.; Sergerie, M.; Demeure, C.E.; Sarfati, M.; Delespesse, G. Naive human CD4+T cells are a major source of lymphotoxin α. J. Immunol. 1999, 162, 3790–3794. [Google Scholar]
- Bullens, D.M.; Rafiq, K.; Kasran, A.; Van Gool, S.W.; Ceuppens, J.L. Naive human T cells can be a source of IL-4 during primary immune responses. Clin. Exp. Immunol. 1999, 118, 384–391. [Google Scholar] [CrossRef]
- Kloosterboer, F.M.; van Luxemburg-Heijs, S.A.; Willemze, R.; Falkenburg, J.H. Similar potential to become activated and proliferate but differential kinetics and profiles of cytokine production of umbilical cord blood T cells and adult blood naive and memory T cells. Hum. Immunol. 2006, 67, 874–883. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target/Antibody | Fluorochrome | Clone | Catalogue | Company |
|---|---|---|---|---|
| Annexin V | FITC | 556420 | BD | |
| Anti-CD3 | APC-CY7 | SK7(Leu-4) | 557832 | BD |
| - | 7-AAD | 51-68981E | BD |
| Antibody | Fluorochrome | Clone | Catalogue | Company |
|---|---|---|---|---|
| Anti-CD45RA | FITC | HI100 | 555488 | BD |
| Anti-CD45RO | PE | UCHL1 | 555493 | BD |
| Anti-CD3 | PE-CY5 | HIT3a | 555341 | BD |
| Anti-CD45 | APC-H7 | 2D1 | 641399 | BD |
| Antibody | Fluorochrome | Clone | Catalogue | Company |
|---|---|---|---|---|
| Anti-IFN-γ | FITC | 4S.B3 | 554551 | BD |
| Anti-IL-4 | PE | 8D4-8 | 12-7049-42 | eBioscience |
| Anti-CD3 | PE-CY5 | HIT3a | 555341 | BD |
| Anti-CD45 | APC-H7 | 2D1 | 641399 | BD |
| Mouse-IgG1k | FITC | MOPC-21 | 555748 | BD |
| Mouse-IgG1k | PE | MOPC-21 | 556650 | BD |
| Antibody | Fluorochrome | Clone | Catalogue | Company |
|---|---|---|---|---|
| Anti-IL-2 | BV421 | 5344.111 | 562914 | BD |
| Anti-IL-10 | AF488 | JES3-9D7 | 501413 | BioLegend |
| Anti-LT-α | PE | 359-81-11 | 554556 | BD |
| Anti-IL-17A | PerCP-Cy™5.5 | N49-653 | 560799 | BD |
| Anti-TGF-βI | PE/Cy7 | TW4-2F8 | 349610 | BioLegend |
| Anti-TNF | APC | MAb11 | 554514 | BD |
| Anti-IFN-γ | APC/CY7 | 4S.B3 | 502530 | BioLegend |
| BV510(viability stain) | - | 564406 | BD |
| Target | Fluorochrome | Clone | Catalogue | Company |
|---|---|---|---|---|
| Anti-IL-13 | BV421 | JES10-5A2 | 563580 | BD |
| Anti-IL-4 | FITC | MP4-25D2 | 562047 | BD |
| Anti-IL-21 | PE | 3A3-N2.1 | 562042 | BD |
| Anti-IL-9 | PerCP-Cy™5.5 | MH9A3 | 561461 | BD |
| Anti-IL-5 | APC | TRFK5 | 562048 | BD |
| Anti-IL-22 | PE/Cy7 | 2G12A41 | 366707 | BioLegend |
| Anti-IFN-γ | APC/CY7 | 4S.B3 | 502530 | BioLegend |
| BV510 (viability stain) | - | 564406 | BD |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perveen, K.; Quach, A.; McPhee, A.; Prescott, S.L.; Barry, S.C.; Hii, C.S.; Ferrante, A. Cord Blood T Cells Expressing High and Low PKCζ Levels Develop into Cells with a Propensity to Display Th1 and Th9 Cytokine Profiles, Respectively. Int. J. Mol. Sci. 2021, 22, 4907. https://doi.org/10.3390/ijms22094907
Perveen K, Quach A, McPhee A, Prescott SL, Barry SC, Hii CS, Ferrante A. Cord Blood T Cells Expressing High and Low PKCζ Levels Develop into Cells with a Propensity to Display Th1 and Th9 Cytokine Profiles, Respectively. International Journal of Molecular Sciences. 2021; 22(9):4907. https://doi.org/10.3390/ijms22094907
Chicago/Turabian StylePerveen, Khalida, Alex Quach, Andrew McPhee, Susan L. Prescott, Simon C. Barry, Charles S. Hii, and Antonio Ferrante. 2021. "Cord Blood T Cells Expressing High and Low PKCζ Levels Develop into Cells with a Propensity to Display Th1 and Th9 Cytokine Profiles, Respectively" International Journal of Molecular Sciences 22, no. 9: 4907. https://doi.org/10.3390/ijms22094907
APA StylePerveen, K., Quach, A., McPhee, A., Prescott, S. L., Barry, S. C., Hii, C. S., & Ferrante, A. (2021). Cord Blood T Cells Expressing High and Low PKCζ Levels Develop into Cells with a Propensity to Display Th1 and Th9 Cytokine Profiles, Respectively. International Journal of Molecular Sciences, 22(9), 4907. https://doi.org/10.3390/ijms22094907