
Mast Cells in the Skin: Defenders of Integrity or Offenders in Inflammation?
by
 , ,
, ,
Martin Voss
1, 
Johanna Kotrba
1 ,
,
Evelyn Gaffal
2,
Konstantinos Katsoulis-Dimitriou
1,† and
Anne Dudeck
1,3,*,† 1
Medical Faculty, Institute for Molecular and Clinical Immunology, Otto-Von-Guericke-University Magdeburg, 39120 Magdeburg, Germany
2
Laboratory for Experimental Dermatology, Department of Dermatology, University Hospital Magdeburg, 39120 Magdeburg, Germany
3
Health Campus Immunology, Infectiology and Inflammation, Otto-Von-Guericke-University Magdeburg, 39120 Magdeburg, Germany
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this paper.
Int. J. Mol. Sci. 2021, 22(9), 4589; https://doi.org/10.3390/ijms22094589
Submission received: 31 March 2021
/
Revised: 23 April 2021
/
Accepted: 25 April 2021
/
Published: 27 April 2021
(This article belongs to the Special Issue Mast Cells: From Host Defense to Pathology)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Mast cells (MCs) are best-known as key effector cells of immediate-type allergic reactions that may even culminate in life-threatening anaphylactic shock syndromes. However, strategically positioned at the host–environment interfaces and equipped with a plethora of receptors, MCs also play an important role in the first-line defense against pathogens. Their main characteristic, the huge amount of preformed proinflammatory mediators embedded in secretory granules, allows for a rapid response and initiation of further immune effector cell recruitment. The same mechanism, however, may account for detrimental overshooting responses. MCs are not only detrimental in MC-driven diseases but also responsible for disease exacerbation in other inflammatory disorders. Focusing on the skin as the largest immune organ, we herein review both beneficial and detrimental functions of skin MCs, from skin barrier integrity via host defense mechanisms to MC-driven inflammatory skin disorders. Moreover, we emphasize the importance of IgE-independent pathways of MC activation and their role in sustained chronic skin inflammation and disease exacerbation.
1. Introduction
The skin is one of the body’s largest organs and serves as a barrier to the outer environment [1,2]. It consists of a complex network of various immune cell types and was, therefore, very early on identified as an immune organ [3]. Within the skin, immune cells are mainly located in the dermis [2] and are important for maintaining skin homeostasis and protecting against mechanical, chemical and pathogenic insults [2,4,5]. The skin is the organ richest in Mast cells (MC), with MCs constituting 10% of leukocytes in the mouse ear skin [6,7]. In healthy adult skin, more MCs are found in distal areas (arms and legs) compared to proximal areas [8]. MCs populate tissues as progenitors derived from the bone marrow or the yolk sac and differentiate locally into mature MCs in response to the existing cytokine micromilieu. Particularly in the skin, recent research in mice has shown that MCs are mainly of yolk sac origin [5,9,10,11,12]. Under physiologic conditions, MCs are located in the dermis, near blood vessels, nerves and hair follicles [6,13]. Perivascular MCs display an elongated cell shape compared to interstitial MCs and, in the case of arterioles, even line the vessel in parallel to the vessel axis [14,15]. Additionally, we recently demonstrated that a distinct number of perivascular MCs were not only attached to the vessel exterior. They formed intraluminal sheets, even under physiologic conditions, and constituted a part of the vascular unit directly in alignment with endothelial cells (ECs) [15].
MCs were first described in 1878 by Paul Ehrlich based on their unique staining with metachromatic dyes [16]. Indeed, MCs contain a large amount of secretory granules that are composed of a proteoglycan structure and in which, due to electrostatic interactions, a plethora of mediators is embedded, including MC-specific proteases, non-specific proteases, cytokines, chemokines, and growth factors [16]. Later on, it was recognized that MCs (1) express a high-density of the high-affinity IgE receptor, FcεRI, (2) bind soluble immunoglobulin E (IgE) antibodies, which are produced by B cells after sensitization to a specific allergen, and (3) are the main deposit of histamine, the bioactive amine, driving allergic reactions and anaphylaxis [17,18]. After reexposure, the same allergen is recognized by antigen-specific IgE, leading to crosslinking of the FcεR and intracellular Ca2+ release. This signaling cascade results in immediate degranulation of the secretory granules, from which, after exocytosis, mediators like histamine, cytokines and proteases extrude sequentially [17,18,19,20]. MC degranulation is followed by secretion of lipid mediators, including leukotrienes and prostaglandins, and de novo synthesis of a broad spectrum of cytokines, chemokines and growth factors [13]. Interestingly, IgE production by B cells is stimulated by MC degranulation, and, at the same time, FcεR expression on the MC plasma membrane is enhanced by IgE in a positive feedback loop [21].
Consequently, skin MCs are strategically positioned and equipped for the defense against invading pathogens, response to allergen encounters, and interaction with a resident or infiltrating immune effector cells. However, these interactions may be beneficial or detrimental, and MCs are heavily involved in inflammatory skin disorders. The purpose of this review is to delineate the fascinating role of MCs in skin barrier function, all the way from maintenance of skin homeostasis to infections and inflammatory disorders.
2. Role of Mast Cells in Barrier Integrity and Host Defense
2.1. Mast Cells in Skin Homeostasis
MCs have been reported to be very important as regulators of epidermal barrier function and skin homeostasis and play important roles in wound healing and skin aging [6,22,23,24,25,26,27]. Their role in skin homeostasis has been often reported, but so far, the underlying mechanisms could not be identified in detail. A common hypothesis is that MCs display their homeostatic effects through interactions with neighboring immune and non-immune-cells. Several publications underline the importance of complex intercellular communication in maintaining barrier function and immune homeostasis [4,6,28,29,30].
MCs are an important source of cytokines, chemokines and growth factors, which can play distinct roles in the skin barrier [31,32]. MCs do not only release mediators during degranulation in response to certain stimuli but they are also reported to secrete them constitutively, which serve as an important tool to communicate with neighboring cells [31].
Especially the communication with fibroblasts (FB) was of particular focus in the last decades [6,30]. FBs are also located in the dermis near MCs and are important for structural skin integrity [5,33]. MCs induce FB proliferation via interleukin (IL)-4 [34,35], IL-13 [36], vascular endothelial growth factor (VEGF) and basic fibroblast growth factor (bFGF) [6,37,38]. Moreover, bidirectional communication is necessary for maintaining skin-barrier homeostasis. The expression and secretion of stem cell factor (SCF), the MC growth factor, by FB promotes MC differentiation and controls MC activation [39,40]. Moreover, FBs inhibit MC activation by secreting the enzyme Cyp26b1, which locally downregulates P2X7 expression on skin MCs. This mechanism serves as a unique skin-barrier homeostatic network inhibiting ATP-dependent MC activation [30]. Additionally, MCs continuously secrete tumor necrosis factor (TNF) [36], IL-1β [41,42], IL-4 [43,44,45,46,47], bFGF/FGF-2 [48,49,50], transforming growth factor β1 (TGF-β1) [51] and VEGF/VPF [52,53]. These mediators have recently been also reported to influence FB functions [6,34,36,54,55], but the direct MC–FB interaction via these cytokines remains to be proven. In addition to their interactions with FBs, MCs were also reported to contribute to skin homeostasis via interactions with keratinocytes (KC) [5,28]. KCs play an important role in MC maturation since they also produce SCF [56,57]. On the other hand, MCs are reported to have both inhibitory and activating effects on KCs. For example, they can express keratinocyte growth factor (KGF) [58] and platelet-activating factor (PAF) [28,59] that activate KCs, while MC release of histamine, heparin and other MC mediators inhibits KC proliferation and, therefore, controls epidermal regeneration [60,61]. MC tryptase and chymase are reported to promote FB proliferation while inhibiting KC proliferation [62]. Additionally, Sehra et al. could show that MCs can regulate epidermal differentiation complex (EDC) genes, suggesting a protective role of MCs in regulating epidermal barrier integrity. Mice lacking MCs exhibited decreased levels of EDC gene expression, which was associated with higher permeability for environmental antigens [63].
Finally, MCs can act on ECs to maintain skin homeostasis [5,28]. Angiogenesis is an important process for normal skin development, homeostasis, and remodeling [64,65]. Skin MCs can spontaneously secrete several angiogenesis-related factors and, therefore, exhibit an intrinsic role in vascular development [28,66,67]. MC-derived tryptase additionally promotes angiogenesis by degrading the basement membrane [68]. Among all vasoactive mediators that are released by MCs [59], it was reported that MCs impact on blood ECs (BECs) via histamine [69], TNF [70,71], leukotrienes [72], prostaglandin D2 (PGD2) [73] PAF [74], VEGF-A and VEGF-B [65,75], IL-13 [76] and IL-1β [77] and on lymphatic ECs (LECs) via histamine [78] VEGF-C and VEGF-D [68,75]. However, these interactions are also characterized by a bidirectional mode. MCs are not only a source but also a target of angiogenic and lymphangiogenic factors [68,75]. VEGF-A that is expressed by ECs can regulate MC proliferation and maturation within the skin [79].
Collectively, the homeostatic environment of the skin is regulated very precisely, and MCs play a crucial role in maintaining skin barrier homeostasis and integrity by interacting with neighboring non-immune cells, like FBs, KCs and ECs [58].
2.2. Mast Cells as a Link between Innate and Adaptive Immunity
MCs are mainly known for their key effector functions in type I allergy, where they are activated by crosslinking of cell-surface-bound FcεRI–IgE complexes by specific antigen [80,81,82]. Additionally, MCs are known to exhibit important innate and adaptive immunity functions, as we previously discussed [83]. Given their response repertoire, consisting of a wide range of surface receptors and proinflammatory mediators, and their strategic positioning, MCs contribute to the first line of host defense against invading pathogens [80,82,84,85,86]. MCs express a broad spectrum of pattern recognition receptors, including Toll-like receptors (TLRs), although very little TLR expression was observed in human skin MCs [87,88], Fc receptors and complement receptors [89]. Additionally, MCs can sense cell stress and tissue damage through alarmin and purinergic receptors and be activated or modulated by binding cytokines, growth factors, chemokines and neuropeptides [84,90,91,92,93].
One of the most characteristic features of MCs is their high amount of intracellular secretory granules, which contain a plethora of preformed mediators [16]. Upon external stimuli, MCs can degranulate within seconds, which allows a faster response than other tissue-resident immune cells [94,95,96,97]. Therefore, in many cases, MCs act as initiators of immune responses. As explained above, MC granules contain a wide range of preformed mediators, including histamine, cytokines, chemokines and proteases [16,98]. They immediately release bioactive amines, histamine and serotonin [99] and trigger blood vessel dilatation and permeabilization, finally causing edema formation. These vascular responses are further enhanced by TNF, proteases and eicosanoids that activate vascular ECs [100]. Subsequently, MCs initiate early neutrophil (Nph) recruitment, for example, by TNF, particularly by direct degranulation of TNF into the bloodstream leading to priming of circulating Nph, by secretion of Nph attractants, such as CXCL-1 (KC) and CXCL-2 (MIP-2), and by the release of IL-33 [70,101,102,103]. Moreover, MCs have been reported to enhance Nph effector functions [104,105]. Due to causing increased vascular permeability and edema formation, MCs may also impact on recruitment of other innate and adaptive immune cells to the site of infection or inflammation [67].
In addition to their innate functions that foster adaptive immune responses, MCs can indirectly affect adaptive immunity by modulating dendritic cell (DC) functions [106,107]. MCs and DCs reside near environmental interfaces, allowing for intense intercellular communication [106,108]. This communication can be based on soluble MC mediators, such as histamine and TNF, or on uptake of intact MC granules by DCs and promotes DC migration, DC maturation and T cell priming capacity [109,110]. Moreover, direct MC–DC interactions, including synapse formation, modulate DC functions and thereby fine-tune adaptive immunity [111,112]. We could recently show that MC–DC synapse formation culminates in MHC class II transfer from DCs to MCs, thereby equipping MCs with antigen-presentation capacities that may contribute to effector T cell activation [112].
Inline, the antigen-presenting capacity of MCs has been reported in several studies [91,113,114]. Direct MC–T cell interaction and synapse formation included MHC class II and costimulatory molecules (CD80 and CD86) but can also be mediated by endothelial cell protein C receptor (EPCR) or MHC class I [114,115,116,117]. Additionally, MCs modulate T cell functions by releasing exosomes and soluble mediators. The mode of MC stimulation can either promote T cell polarization towards TH1, TH2 or TH17 or control immune responses by Treg activation via IL-2 or by direct inhibition of effector T cells via IL-10 [117,118,119,120,121,122,123,124].
Collectively, as discussed in more detail in Katsoulis-Dimitriou et al. [83], MCs critically contribute to innate host defense but also link the innate and adaptive immune response.
2.3. The Role of Mast Cells in Venom Detoxification
In 1991 already, Margie Profet hypothesized that allergic responses might be beneficial in the defense against venoms [125]. However, it took more than 20 years until this theory could be confirmed. By studying mice sensitized with a sub-lethal dose of honeybee venom, Marichal et al. showed that IgE antibody binding to FcεRI was responsible for conferring protection against subsequent lethal challenge [126]. Consequently, IgE-dependent MC activation, the mode of allergic reactions [81], has a protective role against noxious substances [127]. Importantly, a protective effect of MCs against venoms from honeybees and snakes, as well as wasps, scorpions, and the Gila monster, has been reported [126,128]. The response to poison from the Thalassophryne natteteri toadfish has also been recently shown to be IgE mediated [129]. Moreover, Palm et al. provided evidence that, in mice, phospholipase A2 (PLA2), a conserved component of many insect venoms, induced an IL-33-driven TH2 response, protecting against subsequent challenge with a lethal dose of PLA2 [130]. This is of particular interest since MCs express the ST2 receptor and are activated strongly by IL-33 [122]. However, MC response to envenomation is not only driven by IgE and IL-33, but they also possess the Mas-related G-protein coupled receptor MRGPRX2 (or its mouse orthologue Mrgprb2) that directly binds venom components, such as wasp venom peptides [131]. Mechanistically, MCs protect from intoxication by directly degrading venoms via proteases or by regulating the immune response [127]. Akahoshi et al. proved that MC chymase is responsible for enhancing resistance to Gila monster venom by directly degrading helodermin, a component of it [132]. Also, protection from two types of scorpion poisons was attributed to MC chymase [132]. Moreover, MC secreted carboxypeptidase A has been shown to protect from snake and bee venoms by a similar mechanism of direct degradation [133]. Importantly, Anderson et al. have recently shown that human MC tryptase can degrade six different snake venoms and have suggested it as a possible therapeutic strategy for treating snakebites [134].
Importantly, although MCs respond to and degrade venoms in general, they can also have a detrimental role. A massive MC reaction to a small amount of poison, such as a single bee sting, may overshoot and lead to anaphylaxis, a complication that may be exacerbated in patients suffering from mast cell activation syndromes (MCAS) [135].
2.4. The Role of Mast Cells in Bacterial Infections
MCs have been shown to exhibit a protective role in the host defense against a spectrum of bacterial infections [136]. Such infections include Mycoplasma pneumonia [137], Escherichia coli [138], Citrobacter rodentium [139], Francisella tularensis [140], Helicobacter sp. [141], and Mycobacterium tuberculosis [142]. MCs recognize bacteria through TLR signaling and contribute to bacterial clearance by inducing immune cell recruitment and by linking the innate and adaptive immune response through promoting DC maturation (Figure 1A) [83]. For example, Siebenhaar et al. have shown that MC-dependent control of Pseudomonas aeruginosa skin infections involves Nph recruitment by the MCs [143]. More recently, Zimmermann et al. delved deeper into P. aeruginosa skin wound infection, showing that wound healing was delayed in the absence of MCs. This was attributed to impaired bacterial clearance due to the lack of MC-derived IL-6, which was enhancing the bactericidal properties of KCs [144]. In another case, it was shown that MCs could be activated by co-culture with Staphylococcus aureus, a predominantly problematic skin bacterium that can cause serious infections of the skin and lungs and possibly leads to sepsis [145]. Arifuzzaman et al. showed that activation of MCs through the MRGPRX2 receptor, by using the wasp venom component mastoparan, enhanced the clearance of S. aureus from infected mouse skin in a process involving Nph recruitment. In addition, MC activation on this occasion led to more pronounced DC migration and stronger protection against re-infection [146].
However, MCs also directly control bacterial infection through phagocytosis, although they are not professional phagocytes, as well as the production of extracellular traps [147] and the release of antimicrobial peptides [148], such as lipocalin 2 and cathelicidin (Figure 1A) [149,150]. For example, Lei et al. have reported that MCs in S. aureus infected skin abscess were activated, and MC-derived tryptase was responsible for the inflammation [151]. Also, Nakamura et al. have shown that staphylococcal δ-toxin can activate MCs, leading to degranulation, which may contribute to the exacerbation of atopic dermatitis [152]. However, the release of the antimicrobial peptide with antibiotic properties cathelicidin by skin MCs has been reported to prevent skin infection by invasive S. aureus [150]. This is of particular interest when we take into account that IgE mediated mechanisms and MCs are involved in developing acquired immunity against S. aureus [153]
Intriguingly, by murine Mrgprb2, or its human ortholog MRGPRX2, MCs have been shown to detect and respond to quorum sensing signals, substances the bacteria use to communicate with each other and coordinate their behavior [154]. In addition, MCs play a significant role in the production of IL-12, which is needed for the protection of the host from polymicrobial infections [155].
Collectively, these reports highlight the important role of MCs in bacterial clearance, particularly in the context of wound healing.
2.5. The Role of Mast Cells in Virus Infections
MCs have been implicated with responses against a wide variety of viruses [106,156]. These include respiratory viruses, such as a respiratory syncytial virus (RSV) and parainfluenza, where MCs were associated with infection-driven asthma exacerbation [157,158,159], hepatitis C virus (HCV) [160] and even human immunodeficiency virus (HIV) [161]. Of note, when focusing in this review on MC functions in skin inflammation, MCs critically contribute also to viral infections that take place or start in the skin, such as vectorborne diseases (Figure 1B) [156].
Due to their strategic location and inherent nature as sensors of cell stress and inflammatory insult, MCs respond to inflammation caused by mosquito bite saliva and thus drive plasma leakage, Nph infiltration and draining lymph node (LN) hyperplasia [162]. However, MCs were also reported to downregulate antigen-specific responses to mosquito bites through an IL-10-dependent mechanism [163]. This is extremely important given that the local inflammation and innate response caused by the vector insect bite are often crucial for viral dissemination and infection severity, as shown by Pingen et al. in the context of arbovirus infection [164]. Dengue virus (DENV), which is a mosquito-borne flavivirus, can lead to vascular leakage and hemorrhagic fever, thus causing severe morbidity and mortality [165]. It is widely spread in tropical regions and can affect up to a hundred million people per year. Therefore, understanding its pathogenicity and developing new treatments is of paramount importance [166]. MCs have been shown to be infected by DENV, leading to their degranulation, which can be reduced by DENV neutralizing antibodies [167]. Troupin et al. were the first to report that skin MC infection by DENV is crucial for systemic virus dissemination (Figure 1B) since infectious viral particles localize in secretory granules, which are being trafficked to draining LNs [168]. Moreover, MC collaboration with macrophages (Mph) (Figure 1B) has been reported to control viral replication in the skin [169]. Similarly, MC degranulation-driven recruitment of natural killer (NK) cells and natural killer T cells (NKT) has been shown to promote viral clearance in the mouse model (Figure 1B) [170].
However, MCs are not only implicated with DENV dissemination and local skin clearance but also pathology since the innate response to the virus is underlying disease exacerbation [165]. A clinical study by Furuta et al. has shown that MC-derived VEGF, tryptase and chymase contribute to DENV shock syndrome [171]. Another clinical study reported that antibody-mediated MC activation leads to vascular leakage (Figure 1B) during DENV infection [100]. This is further supported by the fact that a high chymase serum level is a prognostic factor for DENV hemorrhagic fever [172]. Given that MCs mainly promote DENV infection, MCs have been proposed as potential therapeutic targets [173]. However, a recent study by Mantri et al. has reported a beneficial role for MCs in DENV infection. More specifically, MCs were shown to form immunological synapses with γδ T cells, leading to their activation and killing of infected DCs, which resulted in controlling the virus [115].
As another flavivirus, the Zika virus has been recognized as a threat to international health after the 2016 outbreak. Zika is a mosquito-borne virus that can lead to congenital defects when passed from the mother to the embryo through the placenta [174]. Similar to DENV, human placental MCs and a human MC cell line have been recently reported to be infected by Zika [175]. In addition, in an in vitro study, MCs have been associated with contributing to Zika virus pathology because MCs from a human cell line have been shown to be infected by the Zika virus and produce viral particles [176].
2.6. The Role of Mast Cells in Parasite Infections
MCs are implicated in cutaneous parasite infections, and, similarly to vectorborne viruses, in cases where parasites are disseminated by insect bites through the skin, such as infection with Leishmania sp., Plasmodium sp. and Trypanosoma sp. [177]. For example, in the case of malaria, when mice were infected with Plasmodium berghei, MCs were observed near sporozoites of the parasite at the site of the mosquito bite [178]. Moreover, a clinical study by Wilainam et al. showed that MC degranulation in the skin of patients with Plasmodium falciparum infection was a strong indicator of parasitemia (Figure 1C) and disease severity [179]. Another study has supported the notion that MCs promote Plasmodium sp. dissemination by showing that Plasmodium berghei infection of mice caused massive MC degranulation in the skin and draining LNs. In this context, disease severity was increased when mice were treated with the MC activator c48/80, while the disease was ameliorated upon treatment with the MC stabilizer DSCG [180].
In addition to vectorborne parasites, we herein focus on Leishmania sp. since they cause a predominantly cutaneous infection that can progress into more systemic forms, such as visceral leishmaniasis [181]. Leishmania promastigotes mainly infect Mph, but L. major and L. infantum have been reported to infect MCs directly [182]. For canine leishmaniosis, as well as human infection with Leishmania brasiliensis, MC accumulation at the site of infection has been reported [183,184]. Moreover, the number of MCs at the site of infection was associated with clinical disease progression [183]. MC numbers further seem to correlate with parasite infection since L. major susceptible BALB/c mice showed elevated MC numbers at the lesion site, in contrast to resistant C57BL/6 mice [181]. Furthermore, MCs seem to be crucial for controlling the infection since MC-deficient mice develop larger lesions with higher parasite loads and are prone to frequent dissemination of L. major to the spleen. In the absence of MCs, fewer DCs are recruited to the lesion (Figure 1C), leading to a deficit of IL-12, which is necessary for developing a healing TH1 response [185]. Another important study has shown that MCs can not only recruit DCs to the site of L. major infection but also directly interact with them (Figure 1C). Subsequent T cell activation by DCs that underwent this crosstalk led to the induction of a TH1 phenotype [14]. However, more recent evidence shows that MCs do not only have a protective role against Leishmania sp. infection through activating DCs but can directly kill the parasites by the production of NOS and the formation of extracellular traps (Figure 1C) [186]. Interestingly, Paul et al. reported in 2016 that MCs are dispensable for cutaneous leishmaniosis, introducing controversy on the topic [187]. Nevertheless, most published reports suggest a rather beneficial role of MCs in L. major infection.
2.7. The Role of Mast Cells in Fungi Infections
Fungi comprise a group of eukaryotic organisms, members of which cause serious infections, affecting approximately 300 million people a year. These infections are often initiated from environment-barrier interfaces, such as the skin and the lung mucosa [188]. MCs may contribute to host defense against fungi since they populate these interfaces in high densities and can sense pathogen products, for example, through TLR or C-type Lectin receptor signaling [89,156]. However, most studies have focused on Mph, monocytes (Mo) and Nph, as well as ECs, rather than on direct MC antifungal activity and orchestration of the immune response [189,190,191].
Nevertheless, MCs have been reported to play a crucial role in various fungal infections [188]. In the case of C. albicans, a fungus that can infect the skin but can also cause systemic infections, a recent study has shown that recognition of the fungi by rat bone marrow-derived MCs (BMMCs) through the C lectin type receptor (CLR) Dectin 1 led to MC degranulation and release of TNF-α, IL-6, IL-10, CCL2, CCL4 and NOS [192]. Moreover, Lopes et al. reported that, in vitro, human MCs could mount specific early, mid and late responses against C. albicans. More specifically, MCs were able to phagocytose the fungi and reduce its viability, followed by recruitment of Nph. In addition, infected MCs formed extracellular DNA traps suggesting, overall, a protective role of MCs against Candida infection [193]. Indeed, MCs were reported in vivo to kill extracellular but not ingested C. albicans in a process potentially involving degranulation [194].
However, MCs do not always have a protective role in fungi infections. In lung infections with Aspergillus fumigatus, resulting in IgE-mediated allergic bronchopulmonary aspergillosis (ABPA), MC proteases are responsible for releasing growth factors from ECs, promoting lesions and fibrosis [195]. This is further supported by the fact that omalizumab, a monoclonal antibody against IgE, is an effective treatment for ABPA [196], but A. fumigatus can also activate MCs in an IgE independent manner [197]. Moreover, extracts of Malassezia sporodialis, a species usually associated with cutaneous diseases and considered to contribute to developing atopic dermatitis [198], have been proven to cause MC activation. Mechanistically, binding of the CLR Dectin 1 to the fungi component curdlan leads to MC degranulation and release of Leukotriene C4, IL-6 and CCL2 [199]. In vitro, Ribbing et al. have also shown that human MCs can detect and respond to Malassezia sp. by engagement of the CLRs Dectin 1 and Mincle [200].
3. Mast Cell Contribution to Inflammatory Skin Disorders
3.1. IgE-Mediated Acute Allergic Cutaneous Responses
MCs are well-known as key effector cells in type I allergic responses. As described above, MCs respond to IgE/FcεRI crosslinking by immediate degranulation of secretory granules. Intriguingly, perivascular MCs can sample IgE from the bloodstream by generating processes through the vessel wall [201]. Granule-bound histamine, which becomes soluble almost immediately, is a key driver of vasodilatation and increased vessel permeability, ultimately resulting in rapid local edema formation. Granule-embedded cytokines and chemokines, including TNF, IL-6, CXCL-1 and eotaxins, account for subsequent effector cell recruitment, including Nph and eosinophil granulocytes (Eos) and Mo [16]. The orchestration of cell recruitment by MCs is further amplified by subsequent de novo synthesis of lipid mediators and additional cytokines and chemokines. This, consequently, manifests clinically as locally restricted early edema, compared to more disseminated and hardened skin edema at late time points. Importantly, due to the plethora of released mediators and multiple overlapping pathways, even local allergen encounters and MC activation may culminate in life-threatening systemic anaphylaxis. The severity of anaphylaxis was shown to increase with high plasma heparin levels resulting in factor XII autoactivation and bradykinin formation [202]. Importantly, the mechanisms converting a local reaction into a systemic anaphylactic response and defining the severity of an anaphylactic reaction remain unknown [203].
3.2. The Role of Mast Cells in Atopic Dermatitis
Atopic diseases are a family of IgE-mediated type I hypersensitivities, including atopic dermatitis (AD), allergic rhinitis, allergic asthma, food allergy and life-threatening anaphylaxis. AD is a chronic recurrent inflammatory skin disease affecting about 30% of children and is characterized by a TH2-cell dominated immune response, itching and impaired skin barrier (Figure 2A) [204]. Importantly, children with an early onset and persistent disease have a high risk of developing allergic asthma, a phenomenon known as “atopic march” [203,205]. AD comprises several endotypes between age groups and ethnicities, characterized by IgE levels and filaggrin mutation status, making traditional non-targeted therapies tricky [206]. Despite the elevated IgE serum levels, the role of MCs in chronic AD is still incompletely understood. Given the expression of FcεRI on Langerhans cells and Mph in chronic AD, an important role for IgE in allergen presentation and expansion of allergen-specific IL-4/IL-5-producing TH2 cells, promoting the subsequent infiltration of Eos (Figure 2A), has been proposed [207,208,209]. However, MCs accumulate in chronic AD lesions and even migrate into the epidermis [210]. Since most papillary and epidermal MCs were localized close to ECs, they may be involved in neoangiogenesis by expressing proangiogenic factors [211]. In addition, MCs have been demonstrated as a source of IL-4 [46], IL-5 [209] and IL-13 [212], thereby likely involved in the IgE/TH2/Eos vicious cycle (Figure 2). Importantly, IL-4 induced MC expansion and functionally re-shaped human skin MCs towards increased FcεRI expression and boosted histamine synthesis and release [213] while, in turn, repeated FcεRI triggering modified the MC transcriptome [214]. In line with this, even progenitor-derived MCs from AD patients differ from healthy controls by having enhanced levels of granule mediators and IL-6 responsiveness, indicating a link to the genetic predisposition of atopic disorders [200].
Depending on the route of activation, MCs can bridge innate and adaptive immunity and thereby contribute to allergic sensitization in the skin and lung [215]. For example, MC-derived IL-13 has been demonstrated to downregulate IL-12 production by skin DCs, thereby inhibiting the TH1 cell response to cutaneous antigen exposure [216]. Likewise, MCs counter-regulate IFN-γ expression in sensitized skin [217]. The pattern of cytokine expression in AD depends on the acuity or duration of the skin lesion. The acute onset of skin inflammation is associated with a predominance of TH2 cell infiltration and IL-4 expression (Figure 2A). In contrast, macrophage and Eos activation are dominant in chronic AD, where MC-derived cytokines, chemokines and MC interaction with the vessel endothelium contribute to monocyte recruitment (Figure 2B) [218,219]. The ongoing cytokine expression, local expansion of Th2 cells and pathologic keratinocyte damage caused by scratching or microbial agents (e.g., Staphylococcus aureus) amplify tissue inflammation. KCs release a spectrum of cytokines, such as IL-1β, IL-25, IL-33, thymic stromal lymphopoietin (TSLP), as well as the alarmins ATP and HMGB1 that skew DC towards type 2 immunity, further boosting MC effector functions [220]. Specifically, in AD, TLSP stimulates MC accumulation and Th2 cytokine production, which directs epithelial cell-mediated, IgE-independent MC activation that exacerbates disease severity (Figure 2B) [221,222,223,224]. Furthermore, it has been recently shown that TSLP can activate human MCs directly through cooperating with MRGPRX2, shedding some light on its IgE-independent mode of action [225]. Notably, TSLP is also produced by MCs in a caspase-1/NFκB-dependent way [226]. In line with TSLP, IL-33 release by KCs acts on TH2 cells, MCs and Eos, the key drivers of AD (Figure 2B), via the specific receptor ST2 [227]. IL-33 affects several MC functions, including growth, survival, and mediator release (as reviewed in [228]). In vitro, IL-33 was thought to fail direct induction of MC degranulation but to amplify the release of de novo synthesized lipid mediators and cytokines. However, we demonstrated that IL-33 initiates MC degranulation and MC-mediated edema formation and Nph recruitment in vivo [90,229], which may result from concomitant signaling of extracellular ATP via the purinergic receptor P2X7. Indeed, transgenic expression of IL-33 by KCs resulted in MC accumulation, increased blood histamine and total IgE levels, and increased levels of IL-5, IL-13, CCL5, and eotaxin 1 in blood and lesional skin, thereby closely resembling the AD features [230]. Notably, in a murine model, MCs were found to express MHC class II after prolonged exposure to IL-33, suggesting a possible role for MCs in promoting the vicious cycle of response to/induction of type 2 immunity [231].
AD exacerbation via IgE-independent MC activation is also triggered by the Mas-related G-protein coupled receptor MRGPRX2 (or its murine orthologue Mrgprb2) (Figure 2B). For example, as a model for AD, imiquimod application induced dermatitis with inflammatory cell infiltrates MC activation and increased histamine and cytokine serum levels in wt mice, but not in MRGPRB2−/− mice [232]. In addition, antimicrobial peptides, neuropeptides, major basic protein, eosinophil peroxidase, and many FDA-approved peptidergic drugs activate human MRGPRX2 and may result in pseudo-allergic responses [233,234]. Recent work on MRGPRX2-mediated signaling of neuropeptides in MCs demonstrated MC/nerve communication (Figure 2B), which is amplified in AD due to MC accumulation, MC/nerve-connections and increased Substance P levels (reviewed in detail in [235,236,237]. Serhan et al. demonstrated that house dust mite (HDM)-activated skin peptidergic nociceptors drive type 2 skin inflammation by induction of MC degranulation through the release of substance P [238]. Thus, MC mediators play a key role in linking nociception to skin inflammation and pruritus [239,240]. In AD, itch, one of the most burdensome hallmarks, is triggered by histaminergic routes [241,242,243] but also by non-histaminergic, but tryptase-dependent, mechanisms (Figure 2B), including MRGPRX2 activation [233,244,245,246,247]. Importantly, itch and scratching facilitate the encounter of external triggers, such as S. aureus, partially activating MRGPRX2 themselves and consequently further promoting the vicious cycle towards disease exacerbation (Figure 2B). For example, S. aureus δ-toxin and S. aureus enterotoxin B (SEB) enhance allergic skin inflammation by activating MCs, the latter via increased expression of IL-33 and ST2 [152,248,249].
Furthermore, pruritus as a main characteristic of AD explains the link between AD and food allergies. Leyva-Castillo et al. showed recently that mechanical skin injury causes expansion and activation of intestinal MCs, increases intestinal permeability and thereby promotes food anaphylaxis in sensitized mice [250]. Notably, the remote association between AD and food allergies again includes IL-33 and TLSP effects on MC degranulation [251].
Recent knowledge may be helpful for identifying potential biomarkers for disease progression and targets for therapeutic strategies, particularly to intervene in the atopic march. Indeed, there is already considerable effort to not only improve the treatment of histaminergic effects [252] but also to include biological therapies targeting the type 2 immune response, involving Th2 cells, MCs and others, such as IL-4, IL-5, IL-13, IL-31, TNF, TLSP [253,254] and IL-33 [255].
3.3. Mast Cell Functions in Contact Hypersensitivity
Affecting about 11% of women and 5% of men in industrialized countries, allergic contact dermatitis (ACD) is one of the most common occupational diseases [256,257,258]. ACD is a chronic skin inflammatory disorder driven by a T cell-mediated delayed-type hypersensitivity (DTH) response (Figure 3) to low-molecular-weight organic chemicals or metal ions (reviewed in [259,260]). During sensitization, these compounds, referred to as “haptens”, penetrate into the skin and bind to self-proteins in the dermis, thereby rendering them antigenic. Skin resident DCs take up the “haptenized” proteins and prime allergen-specific T cells in skin-draining LNs (Figure 3). Upon every following hapten encounter, referred to as “elicitation”, hapten-specific T cells infiltrate the allergen-exposed skin and mediate a Th1, and CD8+ T cell dominated skin inflammatory response [261].
The immune events underlying ACD have been widely studied with the help of experimental mouse models for contact sensitization (contact hypersensitivity, CHS), which is the DTH response to small sensitizing organic haptens, such as DNFB or TNCB [262]. Research of the last decades revealed that an innate immune response involving pattern recognition receptor-mediated sensing and infiltration and effector function of various immune cell types precedes the T cell-mediated inflammation [263,264]. Potently sensitizing haptens evoke a local inflammatory response within the first hours after administration (Figure 3A), providing adjuvant effects that promote the induction of the subsequent allergen-specific T cell response [265,266].
In the early 1990s, MCs have been suspected of promoting DTH elicitation by vasoactive mediators. However, the analysis of CHS responses in Kit mutant mice, as a model for MC deficiency, revealed controversial data ranging from defective [267], normal [268,269] to even enhanced [269] skin inflammation. However, as early as 1987, very early onset of MC degranulation, ear swelling and serum histamine levels in the first hours after hapten treatment have been reported [270]. Demonstrating that MC functions in sensing and initiation of innate responses can be easily overlooked by analyzing late time points, which in CHS are classically 24 h or 48 h after hapten challenge. Nevertheless, there is increasing evidence not only of critical MC relevance in skin inflammation upon hapten elicitation but also of contribution to sensitization (Figure 3A) (reviewed in [258,262,263,271]). Using a novel transgenic mouse model of MC deficiency, independent of kit mutations, we have demonstrated that MCs are key promoters of CHS and mediate the early adjuvant effects of several haptens [94]. Importantly, MC deficiency resulted in diminished edema formation and Nph and T cell skin infiltration upon elicitation, but also impaired migration of skin DCs to the LNs and reduced T cell priming upon hapten sensitization. Given the rapid MC response, we questioned the mode of activation. We found that MCs sense cell stress and tissue damage via concomitant IL-33/ST2 and ATP/P2X7 signaling, while MyD88-dependent TLR signaling is not involved [228]. Efficient MC activation in CHS responses also depends on FcεRI α-chain/ITAM signaling [272], is enhanced by antigen-independent effects of IgE [273], and may include inflammasome activation [274]. Hence, epidermal responses to hapten encounter, e.g., by keratinocytes and dendritic epidermal T cells (DETC) [259,264,275], seem to precede MC activation. In vitro, the formation and internalization of hapten–protein complexes by human keratinocytes has been shown to result in neoepitope formation [276,277,278]. While in vivo, the sequence of events is less understood, epidermal stress responses include inflammasome activation and release of IL-1β and IL-18 [279,280], production of reactive oxygen species (ROS) [281] and alarmins, e.g., ATP [282,283] and IL-33 [284,285,286,287]. The subsequent MC activation rapidly elicits skin blood vessel vasodilatation and increased vessel permeability, resulting in biphasic edema formation, peaking on the first 2 h after hapten encounter and followed by a second peak after 24 to 48 h in sensitized mice, that is predominantly driven by histamine effects [228]. Histamine has also been shown to impact sensitization to nickel [274] and to contribute to chronic ACD [288]. In addition to histamine-driven dysregulation of endothelial barrier integrity, we and others demonstrated a crucial role of MCs, particularly of MC-derived TNF (Figure 3), in the initiation of Nph infiltration to hapten-challenged ear skin [70,94,289]. Biedermann and colleagues demonstrated that MCs promote Nph recruitment by the release of MIP-2 and TNF [70], where, mechanistically, TNF may directly impact TNFR1-expressing ECs [71]. In a recent report, we could show in a mouse model of conditional TNF inactivation in CTMCs, that MC-derived TNF is dispensable for the induction of endothelial cell adhesion molecules. In contrast, we determined that perivascular MCs have the capacity to pass the vessel wall and directionally degranulate into the vessel lumen. After being infused into the bloodstream, MC-TNF directly primes circulating Nph via TNFR1 on the Nph surface and thereby allows their efficient extravasation to the inflamed skin [15]. Importantly, MC-mediated early recruitment of Nph is not only the driving force behind the amplitude of skin inflammation upon elicitation but is also required for efficient sensitization. Weber et al. showed a crucial role for Nph in DC migration to skin draining LNs and priming of allergen-specific T cells [288]. Besides controlling skin Nph numbers, MCs have been shown to contribute to hapten sensitization in various modes of action. Wang et al. reported that upon hapten sensitization, MCs degranulate rapidly, within 30 min, in the affected skin but later on accumulate in the skin draining LNs where they may contribute to T cell priming [290]. Moreover, a direct interaction between MCs and DCs in the skin upon hapten sensitization has been demonstrated by analyzing skin biopsies [291], as well as by dynamic in vivo analysis using intravital multiphoton microscopy [112]. Importantly, in vitro and in vivo data indicate an important role for MCs, and MC-derived TNF, in promoting the maturation of DCs and their subsequent migration to skin draining LNs (Figure 3A) [14,290,292]. Of note, MC-derived TNF amplifies predominantly the migration and function of the cDC1 subtype (e.g., CD103+ skin DCs reflecting the CD8+ LN DCs) and thereby the priming and expansion of CD8+ T cells [293], which have been previously shown to be dominant effector cells in hapten-induced skin inflammation [259]. This specification may be related to the mechanistic impact of MC secretory granules. We could show, that both upon LPS-induced and hapten-driven MC degranulation, the exocytosed intact granules are actively engulfed by neighboring DCs and shuttled to skin draining LNs [106]. Importantly, this MC granule uptake accelerated DC migration, facilitated DC maturation and therefore, boosted T cell priming (Figure 3A). Given the fact, that CD103+ skin cDC1 were most efficient in MC granule uptake, but that this advantage was undone in the absence of MC-TNF, MC-TNF effects on cDC1 functions may occur due to enhanced MC granule uptake. Most importantly, the impact of MCs on DC functionality is simultaneously mirrored by a mutual influence of DCs on MC functions. The hapten-induced dynamic interaction between MCs and DCs observed by intravital imaging, culminated in innate synapses and MHCII transfer from DCs to MCs. Consequently, MCs acquired antigen presenting capacity, which may contribute to effector T cell activation and T cell-mediated skin inflammation upon hapten elicitation [112].
Notably, MC-derived TNF has been additionally shown to promote nerve fiber elongation during CHS, as well as closer MC proximity to nerves (Figure 3B) [294]. Since ACD is manifested as a pruritic inflammatory skin disorder, MC’s contribution to both histaminergic and non-histaminergic itch might be considered [295]. Moreover, models of repeated antigen challenge have been used to study MC effects in chronic ACD. Here, MCs accumulate in an L-selectin and ICAM-1 dependent manner and shift the delayed-type to an immediate-type response [296]. Interestingly, Gimenez-Rivera demonstrated that MCs limit the exacerbation of chronic ACD by controlling TH1 and TH17 cytokines, and CD8+ tissue-resident memory T (TRM) cells, probably by degradation of the CD8+ T cell mitogen IL-15 [297]. In vitro, persistent exposure of human MCs to IL-33, as occurring in chronic inflammatory skin disorders, attenuated degranulation and FcεRI expression but induced amplified histamine production [298].
Of note, NF-κB-induced proinflammatory cytokine production by MCs, upon CHS, is counter-regulated by iTreg cells via TGF-β (Figure 3B) [299]. Moreover, MCs seem to elicit not only proinflammatory but also immunoregulatory functions in ACD (reviewed in [300,301,302]). While in low-dose CHS responses, MCs promote the early onset and magnitude of skin inflammation, MCs may regulate more severe CHS responses and cytokine patterns upon high-dose hapten challenge [303,304]. Mechanistically, MCs suppress severe CHS by the release of IL-10 [245,305], by maintaining IL-10 producing regulatory B cells, through the secretion of IL-5 [306], and by the PD-L1-mediated suppression of CD8+ effector T cell activation [307].
3.4. Mast Cells in Psoriasis
Psoriasis is considered to be an autoimmune disorder that is characterized by massive immune cell infiltration into the dermis and epidermis, followed by an abnormal proliferation of KCs [308]. The skin disease is perpetuated by both innate and adaptive mechanisms (Figure 4A) and, although the inflammation appears locally in the skin, it involves detrimental effects on patient quality of life and may be even accompanied by systemic manifestations [309]. MCs have been investigated early in the context of psoriasis, and it was demonstrated that MC degranulation is a constant and early feature in disease development [310]. Another study showed that MCs in psoriatic skin lesions were functionally hyperactive and suspected that histamine is involved in disease pathophysiology [311]. Indeed, MCs have been implicated in the onset of neuropathic pain, itching and pruritus (Figure 4A), the latter being some of the main symptoms of psoriasis [245]. This is triggered by a plethora of MC mediators, including histamine, tryptase, cytokines and growth factors [240]. In a recent study, Nobuo et al. reported the relationship between MCs and pruritus in a mouse model of imiquimod-induced psoriasis, showing that self-scratching behavior during the onset of psoriasis led to increased MC numbers, which, in turn, induce pruritus through the release of nerve growth factor (NGF) (Figure 4A) [312]. Notably, a recent study has employed transcriptional profiling to establish a common signature for itchy psoriatic and AD skin, showing that many MC genes, such as tryptase, phospholipase A2 (PLA2), IL-6, IL-17, IL-22 and MRGPRX2, were commonly upregulated between the two types of pruritus [313].
In addition to the onset of pruritus, MCs play an active role in driving disease pathogenesis [314]. Mechanistically, IL-33 levels were found to be increased in human psoriatic skin (Figure 4A), and, by stimulating MCs to release VEGF, contributed to the inflammation [315]. This is also supported by evidence that IL-33 in psoriatic plaques is secreted by KCs in response to inflammatory stimuli [316]. Most importantly, MCs have been shown to be major producers of IL-17 and IL-22 in human psoriatic skin, which drive the disease by inducing uncontrolled keratinocyte proliferation and psoriatic plaque formation (Figure 4A), suggesting MC-driven disease exacerbation [317,318]. On this note, a study by Lun et al. reported that PLA2, released by MCs through degranulation, is taken up by CD1a-expressing antigen-presenting cells in psoriatic skin. This promotes the generation of neolipid antigens, which are specifically recognized by CD1a-reactive T cells, leading to the release of IL-17 and IL-22 [123]. Therefore, MCs are not only driving the disease by direct production of IL-17 and IL-22 but also indirectly through the perpetuation of T cell activation (Figure 4A). Considering that MCs promote both the pathomechanism of psoriasis and psoriasis-associated pruritus, MCs represent potential therapeutic targets.
4. Mast Cell-Driven Skin Diseases
4.1. The Role of Mast Cells in Urticaria
Urticaria is a common skin disease that is characterized by transient erythematous swelling of the skin. It can be categorized into acute and chronic urticaria, or according to the clinical course of the disease, into spontaneous, physical (inducible) and other types of urticaria [319]. MCs are key effector cells in the pathogenesis of urticaria, mainly via the release of high amounts of the vasoactive mediator histamine (Figure 4B). Both MC degranulation in lesional skin and increased plasma concentrations of histamine, were observed in patients with chronic spontaneous urticaria (CSU) [320,321], symptomatic dermographism [322,323], cold-induced urticaria [324,325], heat-induced urticaria [326,327], solar urticaria [328], cholinergic urticaria [325,329] and delayed pressure urticaria [322,330]. MC-derived histamine promotes vasodilatation and a local increase in vascular permeability, leading to the prominent symptoms of urticaria, such as wheal formation and angioedema (Figure 4B). Therefore, histamine receptor antagonists, antihistamines, are used as an often inefficient first-line therapy in acute and chronic urticaria [331]. Besides histamine, MC-derived TNF was shown to play an important role in the pathogenesis of CSU by inducing the expression of EC adhesion molecules, including ICAM-1, VCAM-1 and E-Selectin, consequently promoting the recruitment of other immune cells to lesional skin (Figure 4B) [236]. The triggers of MC activation and subsequent degranulation in urticaria are heterogeneous and still not completely understood. In CSU, IgE antibodies against local autoallergens (type I autoimmunity), as well as IgG autoantibodies against IgE or the high-affinity IgE receptor FcεRI (type II autoimmunity) are considered to be the main causes for MC degranulation (Figure 4B) [332]. Based on this, the anti-IgE antibody Omalizumab is an effective therapeutic tool in severe CSU, especially for patients in whom antihistamine treatment is insufficient [333]. Intriguingly, omalizumab also reduced symptoms in patients with symptomatic dermographism, cold urticaria and solar urticaria, indicating a role for IgE-dependent pathomechanisms in physical urticaria [334]. However, recent studies have also revealed IgE-independent mechanisms of MC activation in physical urticaria. Boyden et al. could show, that patients with vibratory urticaria carry a specific mutation in the auto inhibitory subunit of the ADGRE2 receptor, causing hyperresponsiveness of MCs to vibratory stimuli [335]. This finding highlights the involvement of the ADGRE2 receptor in mechanical sensing by MCs. Apart from this, the neuropeptide substance P (SP) might play a key role in the pathogenesis of urticaria (Figure 4B). SP was shown to be increased in the skin of urticaria patients and to induce wheal and flare formation [336,337]. In this line, Fujisawa et al. could recently show, that MRGPRX2, which recognizes SP, is upregulated in skin MCs of CSU patients. Moreover, SP, as well as the Eos-derived mediators major basic protein (MBP) and eosinophil peroxidase, induced the MRGPRX2-dependent release of histamine by human skin MCs (Figure 4B) [338]. Based on this the authors concluded, that MRGPRX2 may be a potential target for therapeutic strategies aimed at alleviating urticaria.
4.2. Mast Cell-Driven Mechanisms in Mastocytosis
Mastocytosis represents a heterogeneous group of primary MC disorders, which result from a clonal expansion and accumulation of MCs in multiple organs (systemic mastocytosis, SM) or predominantly in the skin (cutaneous mastocytosis, CM). The main causes of mastocytosis are hypermorphic mutations in the proto-oncogene KIT (also referred to as c-kit). Whereas the KIT D816V mutation is present in nearly all SM cases (>80%) and can be targeted for therapy [339], the mutation pattern in CM patients varies [340]. KIT encodes for the receptor tyrosine kinase c-kit (CD117), which is the receptor for the MC growth factor SCF [341]. Consequently, c-kit constitutively induces MC proliferation and promotes MC survival mainly via the upregulation of anti-apoptotic molecules [341]. While MCs in the bone marrow of mastocytosis patients show enhanced expression of MCL-1 and Bcl-xL, cutaneous MCs exclusively upregulate Bcl-2, suggesting differential regulation of MC survival in this disease [342,343]. CM can be categorized, according to the characteristics and distribution of skin lesions, into maculopapular CM (also referred to as urticaria pigmentosa), diffuse CM and mastocytoma of the skin [344]. The lesions in CM typically consist of red-brown, itchy macules of varying sizes, which result from local MC accumulation and consequent MC degranulation. Therefore, oral antihistamines and topical corticosteroid treatment are used as first-line therapy to alleviate symptoms [345]. Moreover, tyrosine kinase inhibitors, including imatinib and midostaurin, are now approved for the treatment of systemic mastocytosis and may also be promising for the therapy of CM [346]. However, until now, no cure for mastocytosis has been found.
4.3. Mast Cell Activation Syndrome
Mast cell activation syndrome (MCAS) encompasses the second group of primary MC disorders, which result from chronic aberrant constitutive and reactive MC activation, without MC neoplasia, as observed in mastocytosis. While KIT-mutation-based clonal MC expansion is referred to as a primary MC disorder, MCAS can be further categorized as (1) secondary MCAS with an underlying IgE-dependent allergy or other reactive MCA-triggering pathology; or (2) idiopathic MCAS, where neither a triggering reactive state nor KIT-mutated MCs is identified [347]. While the exact pathomechanism is still unknown, Molderings et al. could show a familial occurrence of MCAS, suggesting a genetic component in this disease [348]. MCAS patients show an extreme heterogeneity of clinical symptoms, including dermatologic, lymphatic, pulmonary, cardiovascular, gastrointestinal, musculoskeletal, neurologic or constitutional disorders, which complicates the exact diagnosis [349]. Skin symptoms of MCAS, including itching, wheeling and flushing, are predominantly caused by the release of histamine and prostaglandin D2 [350]. Therefore, H1 antihistamines and MC-stabilizing agents, such as cromolyn sodium, are the first-line therapy in MCAS [351].
5. Conclusions
MCs are equipped with a plethora of receptors to sense invading pathogens or cell stress and tissue damage. In addition, skin MCs are strategically positioned beneath the epidermal barrier and attached to the endothelial barrier to translating danger signals into systemic signals, recruiting further immune effector cells. This communication axis is mediated by various mediators, including histamine, cytokines, chemokines, growth factors and proteases. The rapid response by degranulation, their capacity to degranulate into the bloodstream and the huge amount of preformed mediators make MCs be of crucial importance for the first-line defense against pathogens. However, when overshooting, the same features are responsible for detrimental effects in pathogen dissemination and disease exacerbation. Moreover, MCs are not only responsible for skin inflammation in the classic MC-driven skin disorders, such as mastocytosis and urticaria, but they also play an initiating and enhancing role in the vicious cycles of skin inflammation in atopic dermatitis, allergic contact dermatitis, and psoriasis. Conclusively, MCs represent the “Jekyll and Hyde” of the immune system, being beneficial in host defense while, at the same time, the “bad guy” driving inflammatory disorders. Both sides, however, highlight MCs as a potential target of therapeutic strategies.
Author Contributions
M.V., J.K., K.K.-D. and A.D. performed bibliographic search and analysis and prepared distinct sections conceiving the concepts expressed in figures. E.G. contributed to the discussion of clinic data. M.V. prepared the figures. K.K.-D. and A.D. conceived the topic and organization of the review and were responsible for the final format of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Deutsche Forschungsgemeinschaft (DFG, Germany) grant 361210922/RTG2408/TP4 and DU1172/8-1 to A.D.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tsepkolenko, A.; Tsepkolenko, V.; Dash, S.; Mishra, A.; Bader, A.; Melerzanov, A.; Giri, S. The regenerative potential of skin and the immune system. Clin. Cosmet. Investig. Dermatol. 2019, 12, 519–532. [Google Scholar] [CrossRef] [Green Version]
- Kabashima, K.; Honda, T.; Ginhoux, F.; Egawa, G. The immunological anatomy of the skin. Nat. Rev. Immunol. 2019, 19, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Salmon, J.K.; Armstrong, C.A.; Ansel, J.C. The skin as an immune organ. West. J. Med. 1994, 160, 146–152. [Google Scholar] [PubMed]
- Sumpter, T.L.; Balmert, S.C.; Kaplan, D.H. Cutaneous immune responses mediated by dendritic cells and mast cells. JCI Insight 2019. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, A.V.; Soulika, A.M. The Dynamics of the Skin’s Immune System. Int. J. Mol. Sci. 2019, 20, 1811. [Google Scholar] [CrossRef] [Green Version]
- Komi, D.E.A.; Khomtchouk, K.; Santa Maria, P.L. A Review of the Contribution of Mast Cells in Wound Healing: Involved Molecular and Cellular Mechanisms. Clin. Rev. Allergy Immunol. 2020, 58, 298–312. [Google Scholar] [CrossRef] [PubMed]
- Dwyer, D.F.; Barrett, N.A.; Austen, K.F.; Kim, E.Y.; Brenner, M.B.; Shaw, L.; Yu, B.; Goldrath, A.; Mostafavi, S.; Regev, A.; et al. Expression profiling of constitutive mast cells reveals a unique identity within the immune system. Nat. Immunol. 2016, 17, 878–887. [Google Scholar] [CrossRef]
- Janssens, A.S. Mast cell distribution in normal adult skin. J. Clin. Pathol. 2005, 58, 285–289. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Liu, Z.; Li, Z.; Wu, Y. Molecular regulation of mast cell development and maturation. Mol. Biol. Rep. 2010, 37, 1993–2001. [Google Scholar] [CrossRef]
- Gentek, R.; Ghigo, C.; Hoeffel, G.; Bulle, M.J.; Msallam, R.; Gautier, G.; Launay, P.; Chen, J.; Ginhoux, F.; Bajénoff, M. Hemogenic Endothelial Fate Mapping Reveals Dual Developmental Origin of Mast Cells. Immunity 2018, 48, 1160–1171.e5. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Liu, S.; Xu, J.; Zhang, X.; Han, D.; Liu, J.; Xia, M.; Yi, L.; Shen, Q.; Xu, S.; et al. Adult Connective Tissue-Resident Mast Cells Originate from Late Erythro-Myeloid Progenitors. Immunity 2018, 49, 640–653.e5. [Google Scholar] [CrossRef] [Green Version]
- Weitzmann, A.; Naumann, R.; Dudeck, A.; Zerjatke, T.; Gerbaulet, A.; Roers, A. Mast Cells Occupy Stable Clonal Territories in Adult Steady-State Skin. J. Investig. Dermatol. 2020, 140, 2433–2441.e5. [Google Scholar] [CrossRef] [PubMed]
- Elieh Ali Komi, D.; Wöhrl, S.; Bielory, L. Mast Cell Biology at Molecular Level: A Comprehensive Review. Clin. Rev. Allergy Immunol. 2020, 58, 342–365. [Google Scholar] [CrossRef]
- Dudeck, A.; Suender, C.A.; Kostka, S.L.; von Stebut, E.; Maurer, M. Mast cells promote Th1 and Th17 responses by modulating dendritic cell maturation and function. Eur. J. Immunol. 2011, 41, 1883–1893. [Google Scholar] [CrossRef] [PubMed]
- Dudeck, J.; Kotrba, J.; Immler, R.; Hoffmann, A.; Voss, M.; Alexaki, V.I.; Morton, L.; Jahn, S.R.; Katsoulis-Dimitriou, K.; Winzer, S.; et al. Directional mast cell degranulation of tumor necrosis factor into blood vessels primes neutrophil extravasation. Immunity 2021, 54. [Google Scholar] [CrossRef]
- Wernersson, S.; Pejler, G. Mast cell secretory granules: Armed for battle. Nat. Rev. Immunol. 2014, 14, 478–494. [Google Scholar] [CrossRef]
- Amin, K. The role of mast cells in allergic inflammation. Respir. Med. 2012, 106, 9–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellman, L.T.; Akula, S.; Thorpe, M.; Fu, Z. Tracing the origins of IgE, mast cells, and allergies by studies of wild animals. Front. Immunol. 2017, 8, 1749. [Google Scholar] [CrossRef] [PubMed]
- Shumilina, E.; Lam, R.S.; Wölbing, F.; Matzner, N.; Zemtsova, I.M.; Sobiesiak, M.; Mahmud, H.; Sausbier, U.; Biedermann, T.; Ruth, P.; et al. Blunted IgE-Mediated Activation of Mast Cells in Mice Lacking the Ca2+—Activated K+ Channel KCa3.1. J. Immunol. 2008, 180, 8040–8047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunder, C.A.; St. John, A.L.; Li, G.; Leong, K.W.; Berwin, B.; Staats, H.F.; Abraham, S.N. Mast cell-derived particles deliver peripheral signals to remote lymph nodes. J. Exp. Med. 2009, 206, 2455–2467. [Google Scholar] [CrossRef] [Green Version]
- Bax, H.J.; Keeble, A.H.; Gould, H.J. Cytokinergic IgE action in mast cell activation. Front. Immunol. 2012, 3, 229. [Google Scholar] [CrossRef] [Green Version]
- Weller, C.L.; Collington, S.J.; Williams, T.; Lamb, J.R. Mast cells in health and disease. Clin. Sci. 2011, 120, 473–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douaiher, J.; Succar, J.; Lancerotto, L.; Gurish, M.F.; Orgill, D.P.; Hamilton, M.J.; Krilis, S.A.; Stevens, R.L. Development of mast cells and importance of their tryptase and chymase serine proteases in inflammation and wound healing. Adv. Immunol. 2014, 122, 211–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang-lin, H.; Wei, G.; Hong-ying, L.; Jian, T. The Role of the Mast Cell in Skin Aging. J. Dermatol. Res. Ther. 2016, 2. [Google Scholar] [CrossRef]
- Abdel Hafez, S.M.N. Age related changes in the dermal mast cells and the associated changes in the dermal collagen and cells: A histological and electron microscopy study. Acta Histochem. 2019, 121, 619–627. [Google Scholar] [CrossRef]
- Pilkington, S.M.; Barron, M.J.; Watson, R.E.B.; Griffiths, C.E.M.; Bulfone-Paus, S. Aged human skin accumulates mast cells with altered functionality that localize to macrophages and vasoactive intestinal peptide-positive nerve fibres. Br. J. Dermatol. 2019, 180, 849–858. [Google Scholar] [CrossRef]
- Ribatti, D.; Tamma, R.; Annese, T.; Crivellato, E. The role of mast cells in human skin cancers. Clin. Exp. Med. 2021. [Google Scholar] [CrossRef] [PubMed]
- Varricchi, G.; Rossi, F.W.; Galdiero, M.R.; Granata, F.; Criscuolo, G.; Spadaro, G.; de Paulis, A.; Marone, G. Physiological Roles of Mast Cells: Collegium Internationale Allergologicum Update 2019. Int. Arch. Allergy Immunol. 2019, 247–261. [Google Scholar] [CrossRef]
- Galli, S.J.; Tsai, M. Mast cells: Versatile regulators of inflammation, tissue remodeling, host defense and homeostasis. J. Dermatol. Sci. 2008, 7–19. [Google Scholar] [CrossRef] [Green Version]
- Kurashima, Y.; Amiya, T.; Fujisawa, K.; Shibata, N.; Suzuki, Y.; Kogure, Y.; Hashimoto, E.; Otsuka, A.; Kabashima, K.; Sato, S.; et al. The enzyme Cyp26b1 mediates inhibition of mast cell activation by fibroblasts to maintain skin-barrier homeostasis. Immunity 2014, 530–541. [Google Scholar] [CrossRef] [Green Version]
- Mukai, K.; Tsai, M.; Saito, H.; Galli, S.J. Mast cells as sources of cytokines, chemokines, and growth factors. Immunol. Rev. 2018, 282, 121–150. [Google Scholar] [CrossRef]
- Tan, S.-Y.; Roediger, B.; Weninger, W. The role of chemokines in cutaneous immunosurveillance. Immunol. Cell Biol. 2015, 93, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Woodley, D.T. Distinct Fibroblasts in the Papillary and Reticular Dermis: Implications for Wound Healing. Dermatol. Clin. 2017, 35, 95–100. [Google Scholar] [CrossRef]
- Richter, A.; Puddicombe, S.M.; Lordan, J.L.; Bucchieri, F.; Wilson, S.J.; Djukanovic, R.; Dent, G.; Holgate, S.T.; Davies, D.E. The contribution of interleukin (IL)-4 and IL-13 to the epithelial-mesenchymal trophic unit in asthma. Am. J. Respir. Cell Mol. Biol. 2001, 25, 385–391. [Google Scholar] [CrossRef] [Green Version]
- Trautmann, A.; Toksoy, A.; Engelhardt, E.; Bröcker, E.B.; Gillitzer, R. Mast cell involvement in normal human skin wound healing: Expression of monocyte chemoattractant protein-1 is correlated with recruitment of mast cells which synthesize interleukin-4 in vivo. J. Pathol. 2000, 190, 100–106. [Google Scholar] [CrossRef]
- Brown Lobbins, M.L.; Shivakumar, B.R.; Postlethwaite, A.E.; Hasty, K.A. Chronic exposure of interleukin-13 suppress the induction of matrix metalloproteinase-1 by tumour necrosis factor α in normal and scleroderma dermal fibroblasts through protein kinase B/Akt. Clin. Exp. Immunol. 2018, 191, 84–95. [Google Scholar] [CrossRef] [Green Version]
- Qu, Z.; Huang, X.; Ahmadi, P.; Stenberg, P.; Liebler, J.M.; Le, A.C.; Planck, S.R.; Rosenbaum, J.T. Synthesis of basic fibroblast growth factor by murine mast cells. Regulation by transforming growth factor beta, tumor necrosis factor alpha, and stem cell factor. Int. Arch. Allergy Immunol. 1998, 115, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Tellechea, A.; Leal, E.C.; Kafanas, A.; Auster, M.E.; Kuchibhotla, S.; Ostrovsky, Y.; Tecilazich, F.; Baltzis, D.; Zheng, Y.; Carvalho, E.; et al. Mast Cells Regulate Wound Healing in Diabetes. Diabetes 2016, 65, 2006–2019. [Google Scholar] [CrossRef] [Green Version]
- Leist, M.; Sünder, C.A.; Drube, S.; Zimmermann, C.; Geldmacher, A.; Metz, M.; Dudeck, A.; Maurer, M. Membrane-bound stem cell factor is the major but not only driver of fibroblast-induced murine skin mast cell differentiation. Exp. Dermatol. 2017, 26, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Smrž, D.; Jung, M.-Y.; Bandara, G.; Desai, A.; Smržová, Š.; Kuehn, H.S.; Beaven, M.A.; Metcalfe, D.D.; Gilfillan, A.M. Stem Cell Factor Programs the Mast Cell Activation Phenotype. J. Immunol. 2012, 188, 5428–5437. [Google Scholar] [CrossRef]
- Nigrovic, P.A.; Binstadt, B.A.; Monach, P.A.; Johnsen, A.; Gurish, M.; Iwakura, Y.; Benoist, C.; Mathis, D.; Lee, D.M. Mast cells contribute to initiation of autoantibody-mediated arthritis via IL-1. Proc. Natl. Acad. Sci. USA 2007, 104, 2325–2330. [Google Scholar] [CrossRef] [Green Version]
- Reber, L.L.; Marichal, T.; Sokolove, J.; Starkl, P.; Gaudenzio, N.; Iwakura, Y.; Karasuyama, H.; Schwartz, L.B.; Robinson, W.H.; Tsai, M.; et al. Contribution of mast cell-derived interleukin-1β to uric acid crystal-induced acute arthritis in mice. Arthritis Rheumatol. 2014, 66, 2881–2891. [Google Scholar] [CrossRef] [PubMed]
- Hu-Li, J.; Shevach, E.M.; Mizuguchi, J.; Ohara, J.; Mosmann, T.; Paul, W.E. B cell stimulatory factor 1 (interleukin 4) is a potent costimulant for normal resting T lymphocytes. J. Exp. Med. 1987, 165, 157–172. [Google Scholar] [CrossRef]
- Bradding, P.; Feather, I.H.; Howarth, P.H.; Mueller, R.; Roberts, J.A.; Britten, K.; Bews, J.P.; Hunt, T.C.; Okayama, Y.; Heusser, C.H. Interleukin 4 is localized to and released by human mast cells. J. Exp. Med. 1992, 176, 1381–1386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradding, P.; Feather, I.H.; Wilson, S.; Bardin, P.G.; Heusser, C.H.; Holgate, S.T.; Howarth, P.H. Immunolocalization of cytokines in the nasal mucosa of normal and perennial rhinitic subjects. The mast cell as a source of IL-4, IL-5, and IL-6 in human allergic mucosal inflammation. J. Immunol. 1993, 151, 3853–3865. [Google Scholar] [PubMed]
- Horsmanheimo, L.; Harvima, I.T.; Järvikallio, A.; Harvima, R.J.; Naukkarinen, A.; Horsmanheimo, M. Mast cells are one major source of interleukin-4 in atopic dermatitis. Br. J. Dermatol. 1994, 131, 348–353. [Google Scholar] [CrossRef]
- Bradding, P.; Feather, I.H.; Wilson, S.; Holgate, S.T.; Howarth, P.H. Cytokine immunoreactivity in seasonal rhinitis: Regulation by a topical corticosteroid. Am. J. Respir. Crit. Care Med. 1995, 151, 1900–1906. [Google Scholar] [CrossRef] [PubMed]
- Artuc, M.; Steckelings, U.M.; Henz, B.M. Mast cell-fibroblast interactions: Human mast cells as source and inducers of fibroblast and epithelial growth factors. J. Investig. Dermatol. 2002, 118, 391–395. [Google Scholar] [CrossRef] [Green Version]
- Reed, J.A.; Albino, A.P.; McNutt, N.S. Human cutaneous mast cells express basic fibroblast growth factor. Lab. Investig. 1995, 72, 215–222. [Google Scholar]
- Qu, Z.; Liebler, J.M.; Powers, M.R.; Galey, T.; Ahmadi, P.; Huang, X.N.; Ansel, J.C.; Butterfield, J.H.; Planck, S.R.; Rosenbaum, J.T. Mast cells are a major source of basic fibroblast growth factor in chronic inflammation and cutaneous hemangioma. Am. J. Pathol. 1995, 147, 564–573. [Google Scholar]
- Pennington, D.W.; Lopez, A.R.; Thomas, P.S.; Peck, C.; Gold, W.M. Dog mastocytoma cells produce transforming growth factor beta1. J. Clin. Investig. 1992, 90, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Grützkau, A.; Krüger-Krasagakes, S.; Baumeister, H.; Schwarz, C.; Kögel, H.; Welker, P.; Lippert, U.; Henz, B.M.; Möller, A. Synthesis, storage, and release of vascular endothelial growth factor/vascular permeability factor (VEGF/VPF) by human mast cells: Implications for the biological significance of VEGF206. Mol. Biol. Cell 1998, 9, 875–884. [Google Scholar] [CrossRef] [Green Version]
- Heissig, B.; Rafii, S.; Akiyama, H.; Ohki, Y.; Sato, Y.; Rafael, T.; Zhu, Z.; Hicklin, D.J.; Okumura, K.; Ogawa, H.; et al. Low-dose irradiation promotes tissue revascularization through VEGF release from mast cells and MMP-9-mediated progenitor cell mobilization. J. Exp. Med. 2005, 202, 739–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Q.; Fang, L.; Tang, X.; Lu, S.; Tamm, M.; Stolz, D.; Roth, M. TGF-β Upregulated Mitochondria Mass through the SMAD2/3→C/EBPβ→PRMT1 Signal Pathway in Primary Human Lung Fibroblasts. J. Immunol. 2019, 202, 37–47. [Google Scholar] [CrossRef]
- Lacy, S.H.; Woeller, C.F.; Thatcher, T.H.; Pollock, S.J.; Small, E.M.; Sime, P.J.; Phipps, R.P. Activated Human Lung Fibroblasts Produce Extracellular Vesicles with Antifibrotic Prostaglandins. Am. J. Respir. Cell Mol. Biol. 2019, 60, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.-A.; Park, M.; Kim, Y.-H.; Woo, S.-Y. Th17 cell-mediated immune responses promote mast cell proliferation by triggering stem cell factor in keratinocytes. Biochem. Biophys. Res. Commun. 2017, 487, 856–861. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Mascarenhas, N.; Eckmann, L.; Miyamoto, Y.; Sun, X.; Kawakami, T.; Di Nardo, A. Skin microbiome promotes mast cell maturation by triggering stem cell factor production in keratinocytes. J. Allergy Clin. Immunol. 2017, 139, 1205–1216.e6. [Google Scholar] [CrossRef] [Green Version]
- Cho, K.-A.; Kim, H.J.; Kim, Y.-H.; Park, M.; Woo, S.-Y. Dexamethasone Promotes Keratinocyte Proliferation by Triggering Keratinocyte Growth Factor in Mast Cells. Int. Arch. Allergy Immunol. 2019, 179, 53–61. [Google Scholar] [CrossRef]
- Feuerherm, A.J.; Jørgensen, K.M.; Sommerfelt, R.M.; Eidem, L.E.; Lægreid, A.; Johansen, B. Platelet-activating factor induces proliferation in differentiated keratinocytes. Mol. Cell. Biochem. 2013, 384, 83–94. [Google Scholar] [CrossRef]
- Huttunen, M.; Hyttinen, M.; Nilsson, G.; Butterfield, J.H.; Horsmanheimo, M.; Harvima, I.T. Inhibition of keratinocyte growth in cell culture and whole skin culture by mast cell mediators. Exp. Dermatol. 2001, 10, 184–192. [Google Scholar] [CrossRef]
- Gschwandtner, M.; Mildner, M.; Mlitz, V.; Gruber, F.; Eckhart, L.; Werfel, T.; Gutzmer, R.; Elias, P.M.; Tschachler, E. Histamine suppresses epidermal keratinocyte differentiation and impairs skin barrier function in a human skin model. Allergy 2013, 68, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Artuc, M.; Steckelings, U.M.; Grützkau, A.; Smorodchenko, A.; Henz, B.M. A long-term coculture model for the study of mast cell-keratinocyte interactions. J. Investig. Dermatol. 2002, 119, 411–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sehra, S.; Serezani, A.P.M.; Ocaña, J.A.; Travers, J.B.; Kaplan, M.H. Mast Cells Regulate Epidermal Barrier Function and the Development of Allergic Skin Inflammation. J. Investig. Dermatol. 2016, 136, 1429–1437. [Google Scholar] [CrossRef] [Green Version]
- Swerlick, R.A. Angiogenesis. J. Dermatol. 1995, 22, 845–852. [Google Scholar] [CrossRef] [PubMed]
- Marone, G.; Varricchi, G.; Loffredo, S.; Granata, F. Mast cells and basophils in inflammatory and tumor angiogenesis and lymphangiogenesis. Eur. J. Pharmacol. 2016, 146–151. [Google Scholar] [CrossRef]
- McHale, C.; Mohammed, Z.; Gomez, G. Human Skin-Derived Mast Cells Spontaneously Secrete Several Angiogenesis-Related Factors. Front. Immunol. 2019, 1445. [Google Scholar] [CrossRef] [PubMed]
- Kunder, C.A.; St John, A.L.; Abraham, S.N. Mast cell modulation of the vascular and lymphatic endothelium. Blood 2011, 118, 5383–5393. [Google Scholar] [CrossRef]
- Hiromatsu, Y.; Toda, S. Mast cells and angiogenesis. Microsc. Res. Tech. 2003, 60, 64–69. [Google Scholar] [CrossRef]
- Ashina, K.; Tsubosaka, Y.; Nakamura, T.; Omori, K.; Kobayashi, K.; Hori, M.; Ozaki, H.; Murata, T. Histamine Induces Vascular Hyperpermeability by Increasing Blood Flow and Endothelial Barrier Disruption In Vivo. PLoS ONE 2015, 10, e0132367. [Google Scholar] [CrossRef]
- Biedermann, T.; Kneilling, M.; Mailhammer, R.; Maier, K.; Sander, C.A.; Kollias, G.; Kunkel, S.L.; Hültner, L.; Röcken, M. Mast cells control neutrophil recruitment during T cell-mediated delayed-type hypersensitivity reactions through tumor necrosis factor and macrophage inflammatory protein 2. J. Exp. Med. 2000, 192, 1441–1452. [Google Scholar] [CrossRef] [Green Version]
- Kneilling, M.; Mailhammer, R.; Hültner, L.; Schönberger, T.; Fuchs, K.; Schaller, M.; Bukala, D.; Massberg, S.; Sander, C.A.; Braumüller, H.; et al. Direct crosstalk between mast cell-TNF and TNFR1-expressing endothelia mediates local tissue inflammation. Blood 2009, 114, 1696–1706. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, T.M.; Zimmerman, G.A.; Prescott, S.M. Leukotrienes C4 and D4 stimulate human endothelial cells to synthesize platelet-activating factor and bind neutrophils. Proc. Natl. Acad. Sci. USA 1986, 83, 2204–2208. [Google Scholar] [CrossRef] [Green Version]
- Qu, W.M.; Huang, Z.L.; Xu, X.H.; Aritake, K.; Eguchi, N.; Nambu, F.; Narumiya, S.; Urade, Y.; Hayaishi, O. Lipocalin-type prostaglandin D syntase produces prostaglandin D2 involved in regulation of physiological sleep. Proc. Natl. Acad. Sci. USA 2006, 103, 17949–17954. [Google Scholar] [CrossRef] [Green Version]
- Brailoiu, E.; Barlow, C.L.; Ramirez, S.H.; Abood, M.E.; Brailoiu, G.C. Effects of Platelet-Activating Factor on Brain Microvascular Endothelial Cells. Neuroscience 2018, 377, 105–113. [Google Scholar] [CrossRef]
- Detoraki, A.; Staiano, R.I.; Granata, F.; Giannattasio, G.; Prevete, N.; de Paulis, A.; Ribatti, D.; Genovese, A.; Triggiani, M.; Marone, G. Vascular endothelial growth factors synthesized by human lung mast cells exert angiogenic effects. J. Allergy Clin. Immunol. 2009, 123, 1142–1149.e5. [Google Scholar] [CrossRef]
- Takagi, K.; Yamakuchi, M.; Matsuyama, T.; Kondo, K.; Uchida, A.; Misono, S.; Hashiguchi, T.; Inoue, H. IL-13 enhances mesenchymal transition of pulmonary artery endothelial cells via down-regulation of miR-424/503 in vitro. Cell. Signal. 2018, 42, 270–280. [Google Scholar] [CrossRef]
- Mohr, T.; Haudek-Prinz, V.; Slany, A.; Grillari, J.; Micksche, M.; Gerner, C. Proteome profiling in IL-1 β and VEGF-activated human umbilical vein endothelial cells delineates the interlink between inflammation and angiogenesis. PLoS ONE 2017, 12, e0179065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal, S.; Nath, S.; Meininger, C.J.; Gashev, A.A. Emerging Roles of Mast Cells in the Regulation of Lymphatic Immuno-Physiology. Front. Immunol. 2020, 11, 1234. [Google Scholar] [CrossRef]
- Koh, M.; Noguchi, S.; Araki, M.; Otsuka, H.; Yokosuka, M.; Soeta, S. Expressions of vascular endothelial growth factor receptors, Flk1 and Flt1, in rat skin mast cells during development. J. Vet. Med. Sci. 2020, 82, 745–753. [Google Scholar] [CrossRef] [PubMed]
- Metcalfe, D.D.; Baram, D.; Mekori, Y.A. Mast cells. Physiol. Rev. 1997, 77, 1033–1079. [Google Scholar] [CrossRef] [PubMed]
- Galli, S.J.; Tsai, M. IgE and mast cells in allergic disease. Nat. Med. 2012, 18, 693–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurish, M.F.; Austen, K.F. Developmental origin and functional specialization of mast cell subsets. Immunity 2012, 37, 25–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katsoulis-Dimitriou, K.; Kotrba, J.; Voss, M.; Dudeck, J.; Dudeck, A. Mast Cell Functions Linking Innate Sensing to Adaptive Immunity. Cells 2020, 9, 2538. [Google Scholar] [CrossRef] [PubMed]
- Marshall, J.S. Mast-cell responses to pathogens. Nat. Rev. Immunol. 2004, 4, 787–799. [Google Scholar] [CrossRef] [PubMed]
- St John, A.L.; Abraham, S.N. Innate immunity and its regulation by mast cells. J. Immunol. 2013, 190, 4458–4463. [Google Scholar] [CrossRef]
- Galli, S.J.; Nakae, S.; Tsai, M. Mast cells in the development of adaptive immune responses. Nat. Immunol. 2005, 6, 135–142. [Google Scholar] [CrossRef]
- Motakis, E.; Guhl, S.; Ishizu, Y.; Itoh, M.; Kawaji, H.; De Hoon, M.; Lassmann, T.; Carninci, P.; Hayashizaki, Y.; Zuberbier, T.; et al. Redefinition of the human mast cell transcriptome by deep-CAGE sequencing. Blood 2014, 123. [Google Scholar] [CrossRef]
- Plum, T.; Wang, X.; Rettel, M.; Krijgsveld, J.; Feyerabend, T.B.; Rodewald, H.R. Human Mast Cell Proteome Reveals Unique Lineage, Putative Functions, and Structural Basis for Cell Ablation. Immunity 2020, 52, 404–416.e5. [Google Scholar] [CrossRef] [PubMed]
- Espinosa-Riquer, Z.P.; Segura-Villalobos, D.; Ramírez-Moreno, I.G.; Pérez Rodríguez, M.J.; Lamas, M.; Gonzalez-Espinosa, C. Signal Transduction Pathways Activated by Innate Immunity in Mast Cells: Translating Sensing of Changes into Specific Responses. Cells 2020, 9, 2411. [Google Scholar] [CrossRef] [PubMed]
- Hoppe, A.; Katsoulis-Dimitriou, K.; Edler, H.J.; Dudeck, J.; Drube, S.; Dudeck, A. Mast cells initiate the vascular response to contact allergens by sensing cell stress. J. Allergy Clin. Immunol. 2020, 145, 1476–1479.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frandji, P.; Tkaczyk, C.; Oskeritzian, C.; David, B.; Desaymard, C.; Mécheri, S.; Leveson-Gower, D.B.; Sega, E.I.; Kalesnikoff, J.; Florek, M.; et al. Mast cells signal their importance in health and disease. Immunol. Rev. 2018, 282, 17949–17954. [Google Scholar] [CrossRef]
- Espinosa, E.; Valitutti, S. New roles and controls of mast cells. Curr. Opin. Immunol. 2018, 50, 39–47. [Google Scholar] [CrossRef]
- Redegeld, F.A.; Yu, Y.; Kumari, S.; Charles, N.; Blank, U. Non-IgE mediated mast cell activation. Immunol. Rev. 2018, 282, 87–113. [Google Scholar] [CrossRef]
- Dudeck, A.; Dudeck, J.; Scholten, J.; Petzold, A.; Surianarayanan, S.; Köhler, A.; Peschke, K.; Vöhringer, D.; Waskow, C.; Krieg, T.; et al. Mast cells are key promoters of contact allergy that mediate the adjuvant effects of haptens. Immunity 2011, 34, 973–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballesteros-Martinez, C.; Mendez-Barbero, N.; Montalvo-Yuste, A.; Jensen, B.M.; Gomez-Cardenosa, A.; Klitfod, L.; Garrido-Arandia, M.; Alvarez-Llamas, G.; Pastor-Vargas, C.; Vivanco, F.; et al. Endothelial Regulator of Calcineurin 1 Promotes Barrier Integrity and Modulates Histamine-Induced Barrier Dysfunction in Anaphylaxis. Front. Immunol. 2017, 8, 1323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendez-Barbero, N.; Yuste-Montalvo, A.; Nuñez-Borque, E.; Jensen, B.M.; Gutiérrez-Muñoz, C.; Tome-Amat, J.; Garrido-Arandia, M.; Díaz-Perales, A.; Ballesteros-Martinez, C.; Laguna, J.J.; et al. The TNF-like weak inducer of the apoptosis/fibroblast growth factor-inducible molecule 14 axis mediates histamine and platelet-activating factor-induced subcutaneous vascular leakage and anaphylactic shock. J. Allergy Clin. Immunol. 2020, 145, 583–596.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikelis, C.M.; Simaan, M.; Ando, K.; Fukuhara, S.; Sakurai, A.; Amornphimoltham, P.; Masedunskas, A.; Weigert, R.; Chavakis, T.; Adams, R.H.; et al. RhoA and ROCK mediate histamine-induced vascular leakage and anaphylactic shock. Nat. Commun. 2015, 6, 6725. [Google Scholar] [CrossRef] [Green Version]
- Rönnberg, E.; Melo, F.R.; Pejler, G. Mast cell proteoglycans. J. Histochem. Cytochem. 2012, 60, 950–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AhYoung, A.P.; Eckard, S.C.; Gogineni, A.; Xi, H.; Lin, S.J.; Gerhardy, S.; Cox, C.; Phung, Q.T.; Hackney, J.A.; Katakam, A.K.; et al. Neutrophil serine protease 4 is required for mast cell-dependent vascular leakage. Commun. Biol. 2020, 3, 687. [Google Scholar] [CrossRef]
- Syenina, A.; Jagaraj, C.J.; Aman, S.A.B.; Sridharan, A.; St John, A.L. Dengue vascular leakage is augmented by mast cell degranulation mediated by immunoglobulin Fcγ receptors. eLife 2015, 4. [Google Scholar] [CrossRef] [Green Version]
- De Filippo, K.; Dudeck, A.; Hasenberg, M.; Nye, E.; van Rooijen, N.; Hartmann, K.; Gunzer, M.; Roers, A.; Hogg, N. Mast cell and macrophage chemokines CXCL1/CXCL2 control the early stage of neutrophil recruitment during tissue inflammation. Blood 2013, 121, 4930–4937. [Google Scholar] [CrossRef] [Green Version]
- Nyekel, F.N.; Pacreau, E.; Benadda, S.; Msallam, R.; Abrink, M.; Pejler, G.; Davoust, J.; Benhamou, M.; Charles, N.; Launay, P.; et al. Mast cell degranulation exacerbates skin rejection by enhancing neutrophil recruitment. Front. Immunol. 2018, 9, 2690. [Google Scholar] [CrossRef]
- Schramm, R.; Schaefer, T.; Menger, M.D.; Thorlacius, H. Acute mast cell-dependent neutrophil recruitment in the skin is mediated by KC and LFA-1: Inhibitory mechanisms of dexamethasone. J. Leukoc. Biol. 2002, 72, 1122–1132. [Google Scholar] [PubMed]
- Sutherland, R.E.; Olsen, J.S.; McKinstry, A.; Villalta, S.A.; Wolters, P.J. Mast cell IL-6 improves survival from Klebsiella pneumonia and sepsis by enhancing neutrophil killing. J. Immunol. 2008, 181, 5598–5605. [Google Scholar] [CrossRef] [Green Version]
- Doener, F.; Michel, A.; Reuter, S.; Friedrich, P.; Böhm, L.; Relle, M.; Codarri, L.; Tenzer, S.; Klein, M.; Bopp, T.; et al. Mast cell-derived mediators promote murine neutrophil effector functions. Int. Immunol. 2013, 25, 553–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudeck, A.; Köberle, M.; Goldmann, O.; Meyer, N.; Dudeck, J.; Lemmens, S.; Rohde, M.; Roldán, N.G.; Dietze-Schwonberg, K.; Orinska, Z.; et al. Mast cells as protectors of health. J. Allergy Clin. Immunol. 2019, 144, S4–S18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gri, G.; Frossi, B.; D’Inca, F.; Danelli, L.; Betto, E.; Mion, F.; Sibilano, R.; Pucillo, C. Mast cell: An emerging partner in immune interaction. Front. Immunol. 2012, 3, 120. [Google Scholar] [CrossRef] [Green Version]
- Carroll-Portillo, A.; Surviladze, Z.; Cambi, A.; Lidke, D.S.; Wilson, B.S. Mast cell synapses and exosomes: Membrane contacts for information exchange. Front. Immunol. 2012, 3, 46. [Google Scholar] [CrossRef] [Green Version]
- Dudeck, J.; Froebel, J.; Kotrba, J.; Lehmann, C.H.K.K.; Dudziak, D.; Speier, S.; Nedospasov, S.A.; Schraven, B.; Dudeck, A. Engulfment of mast cell secretory granules on skin inflammation boosts dendritic cell migration and priming efficiency. J. Allergy Clin. Immunol. 2019, 143, 1849–1864.e4. [Google Scholar] [CrossRef]
- Caron, G.; Delneste, Y.; Roelandts, E.; Duez, C.; Herbault, N.; Magistrelli, G.; Bonnefoy, J.Y.; Pestel, J.; Jeannin, P. Histamine induces CD86 expression and chemokine production by human immature dendritic cells. J. Immunol. 2001, 166, 6000–6006. [Google Scholar] [CrossRef]
- Carroll-Portillo, A.; Cannon, J.L.; te Riet, J.; Holmes, A.; Kawakami, Y.; Kawakami, T.; Cambi, A.; Lidke, D.S. Mast cells and dendritic cells form synapses that facilitate antigen transfer for T cell activation. J. Cell Biol. 2015, 210, 851–864. [Google Scholar] [CrossRef]
- Dudeck, J.; Medyukhina, A.; Fröbel, J.; Svensson, C.-M.M.; Kotrba, J.; Gerlach, M.; Gradtke, A.-C.C.; Schröder, B.; Speier, S.; Figge, M.T.; et al. Mast cells acquire MHC II from dendritic cells during skin inflammation. J. Exp. Med. 2017, 214, 3791–3811. [Google Scholar] [CrossRef] [PubMed]
- Kambayashi, T.; Allenspach, E.J.; Chang, J.T.; Zou, T.; Shoag, J.E.; Reiner, S.L.; Caton, A.J.; Koretzky, G.A. Inducible MHC Class II Expression by Mast Cells Supports Effector and Regulatory T Cell Activation. J. Immunol. 2009, 182, 4686–4695. [Google Scholar] [CrossRef] [PubMed]
- Gaudenzio, N.; Espagnolle, N.; Mars, L.T.; Liblau, R.; Valitutti, S.; Espinosa, E. Cell-cell cooperation at the T helper cell/mast cell immunological synapse. Blood 2009, 114, 4979–4988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantri, C.K.; St. John, A.L. Immune synapses between mast cells and γδ T cells limit viral infection. J. Clin. Investig. 2019, 129, 1094–1108. [Google Scholar] [CrossRef] [Green Version]
- Stelekati, E.; Bahri, R.; D’Orlando, O.; Orinska, Z.; Mittrücker, H.-W.W.; Langenhaun, R.; Glatzel, M.; Bollinger, A.; Paus, R.; Bulfone-Paus, S. Mast cell-mediated antigen presentation regulates CD8+ T cell effector functions. Immunity 2009, 31, 665–676. [Google Scholar] [CrossRef]
- Schwartz, L.B.; Riedel, C.; Caulfield, J.P.; Wasserman, S.I.; Austen, K.F. Cell association of complexes of chymase, heparin proteoglycan, and protein after degranulation by rat mast cells. J. Immunol. 1981, 126, 2071–2078. [Google Scholar]
- Admyre, C.; Telemo, E.; Almqvist, N.; Lötvall, J.; Lahesmaa, R.; Scheynius, A.; Gabrielsson, S. Exosomes—Nanovesicles with possible roles in allergic inflammation. Allergy 2008, 63, 404–408. [Google Scholar] [CrossRef]
- Jutel, M.; Watanabe, T.; Klunker, S.; Akdis, M.; Thomet, O.A.R.; Malolepszy, J.; Zak-Nejmark, T.; Koga, R.; Kobayashi, T.; Blaser, K.; et al. Histamine regulates T-cell and antibody responses by differential expression of H1 and H2 receptors. Nature 2001, 413, 420–425. [Google Scholar] [CrossRef] [Green Version]
- Suurmond, J.; Habets, K.L.L.; Dorjée, A.L.; Huizinga, T.W.; Toes, R.E.M. Expansion of Th17 Cells by Human Mast Cells Is Driven by Inflammasome-Independent IL-1 β. J. Immunol. 2016, 197, 4473–4481. [Google Scholar] [CrossRef] [Green Version]
- Skokos, D.; Le Panse, S.; Villa, I.; Rousselle, J.-C.C.; Peronet, R.; David, B.; Namane, A.; Mécheri, S. Mast cell-dependent B and T lymphocyte activation is mediated by the secretion of immunologically active exosomes. J. Immunol. 2001, 166, 868–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salamon, P.; Shefler, I.; Moshkovits, I.; Munitz, A.; Horwitz Klotzman, D.; Mekori, Y.A.; Hershko, A.Y. IL-33 and IgE stimulate mast cell production of IL-2 and regulatory T cell expansion in allergic dermatitis. Clin. Exp. Allergy 2017, 47, 1409–1416. [Google Scholar] [CrossRef] [PubMed]
- Cheung, K.L.; Jarrett, R.; Subramaniam, S.; Salimi, M.; Gutowska-Owsiak, D.; Chen, Y.-L.L.; Hardman, C.; Xue, L.; Cerundolo, V.; Ogg, G. Psoriatic T cells recognize neolipid antigens generated by mast cell phospholipase delivered by exosomes and presented by CD1a. J. Exp. Med. 2016, 213, 2399–2412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leveson-Gower, D.B.; Sega, E.I.; Kalesnikoff, J.; Florek, M.; Pan, Y.; Pierini, A.; Galli, S.J.; Negrin, R.S. Mast cells suppress murine GVHD in a mechanism independent of CD4+CD25+ regulatory T cells. Blood 2013, 122, 3659–3665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Profet, M. The function of allergy: Immunological defense against toxins. Q. Rev. Biol. 1991, 66, 23–62. [Google Scholar] [CrossRef] [PubMed]
- Marichal, T.; Starkl, P.; Reber, L.L.; Kalesnikoff, J.; Oettgen, H.C.; Tsai, M.; Metz, M.; Galli, S.J. A beneficial role for immunoglobulin E in host defense against honeybee venom. Immunity 2013, 39, 963–975. [Google Scholar] [CrossRef] [Green Version]
- Tsai, M.; Starkl, P.; Marichal, T.; Galli, S.J. Testing the “toxin hypothesis of allergy”: Mast cells, IgE, and innate and acquired immune responses to venoms. Curr. Opin. Immunol. 2015, 36, 80–87. [Google Scholar] [CrossRef] [Green Version]
- Galli, S.J.; Starkl, P.; Marichal, T.; Tsai, M. Mast Cells and IgE can Enhance Survival During Innate and Acquired Host Responses to Venoms. Trans. Am. Clin. Climatol. Assoc. 2017, 128, 193–221. [Google Scholar]
- Bruni, F.M.; Coutinho, E.M.M.; Andrade-Barros, A.I.; Grund, L.Z.; Lopes-Ferreira, M.; Lima, C. Anaphylaxis induced by Thalassophryne nattereri venom in mice is an IgE/IgG1-mediated, IL-4-dependent phenomenon. Sci. Rep. 2020, 10, 1–15. [Google Scholar] [CrossRef]
- Palm, N.W.; Rosenstein, R.K.; Yu, S.; Schenten, D.D.; Florsheim, E.; Medzhitov, R. Bee venom phospholipase A2 induces a primary type 2 response that is dependent on the receptor ST2 and confers protective immunity. Immunity 2013, 39, 976–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNeil, B.D.; Pundir, P.; Meeker, S.; Han, L.; Undem, B.J.; Kulka, M.; Dong, X. Identification of a mast-cell-specific receptor crucial for pseudo-allergic drug reactions. Nature 2015, 519, 237–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akahoshi, M.; Song, C.H.; Piliponsky, A.M.; Metz, M.; Guzzetta, A.; Åbrink, M.; Schlenner, S.M.; Feyerabend, T.B.; Rodewald, H.R.; Pejler, G.; et al. Mast cell chymase reduces the toxicity of Gila monster venom, scorpion venom, and vasoactive intestinal polypeptide in mice. J. Clin. Investig. 2011, 121, 4180–4191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metz, M.; Piliponsky, A.M.; Chan, C.C.; Lammel, V.; Åbrink, M.; Pejler, G.; Tsai, M.; Galli, S.J. Mast cells can enhance resistance to snake and honeybee venoms. Science 2006, 313, 526–530. [Google Scholar] [CrossRef] [Green Version]
- Anderson, E.; Stavenhagen, K.; Kolarich, D.; Sommerhoff, C.P.; Maurer, M.; Metz, M. Human mast cell tryptase is a potential treatment for snakebite envenoming across multiple snake species. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef]
- Bonadonna, P.; Bonifacio, M.; Lombardo, C.; Zanotti, R. Hymenoptera Allergy and Mast Cell Activation Syndromes. Curr. Allergy Asthma Rep. 2016, 16, 1–9. [Google Scholar] [CrossRef]
- Johnzon, C.F.; Rönnberg, E.; Pejler, G. The Role of Mast Cells in Bacterial Infection. Am. J. Pathol. 2016, 186, 4–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Zhang, D.; Lyubynska, N.; Wolters, P.J.; Killeen, N.P.; Baluk, P.; McDonald, D.M.; Hawgood, S.; Caughey, G.H. Mast cells protect mice from mycoplasma pneumonia. Am. J. Respir. Crit. Care Med. 2006, 173, 219–225. [Google Scholar] [CrossRef] [Green Version]
- Kulka, M.; Fukuishi, N.; Rottem, M.; Mekori, Y.A.; Metcalfe, D.D. Mast cells, which interact with Escherichia coli, up-regulate genes associated with innate immunity and become less responsive to FcεRI-mediated activation. J. Leukoc. Biol. 2006, 79, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Wei, O.L.; Hilliard, A.; Kalman, D.; Sherman, M. Mast cells limit systemic bacterial dissemination but not colitis in response to Citrobacter rodentium. Infect. Immun. 2005, 73, 1978–1985. [Google Scholar] [CrossRef] [Green Version]
- Ketavarapu, J.M.; Rodriguez, A.R.; Yu, J.J.; Cong, Y.; Murthy, A.K.; Forsthuber, T.G.; Guentzel, M.N.; Klose, K.E.; Berton, M.T.; Arulanandam, B.P. Mast cells inhibit intramacrophage Francisella tularensis replication via contact and secreted products including IL-4. Proc. Natl. Acad. Sci. USA 2008, 105, 9313–9318. [Google Scholar] [CrossRef] [Green Version]
- Velin, D.; Bachmann, D.; Bouzourene, H.; Michetti, P. Mast cells are critical mediators of vaccine-induced Helicobacter clearance in the mouse model. Gastroenterology 2005, 129, 142–155. [Google Scholar] [CrossRef]
- Torres-Atencio, I.; Rosero, S.; Ordoñez, C.; Ruiz, M.; Goodridge, A. Mycobacterial lipids induce calcium mobilization and degranulation of mast cells. Am. J. Respir. Crit. Care Med. 2018, 198, 813–816. [Google Scholar] [CrossRef] [PubMed]
- Siebenhaar, F.; Syska, W.; Weller, K.; Magerl, M.; Zuberbier, T.; Metz, M.; Maurer, M. Control of Pseudomonas aeruginosa skin infections in mice is mast cell-dependent. Am. J. Pathol. 2007, 170, 1910–1916. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, C.; Troeltzsch, D.; Giménez-Rivera, V.A.; Galli, S.J.; Metz, M.; Maurer, M.; Siebenhaar, F. Mast cells are critical for controlling the bacterial burden and the healing of infected wounds. Proc. Natl. Acad. Sci. USA 2019, 116, 20500–20504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rönnberg, E.; Johnzon, C.-F.; Calounova, G.; Garcia Faroldi, G.; Grujic, M.; Hartmann, K.; Roers, A.; Guss, B.; Lundequist, A.; Pejler, G. Mast cells are activated by Staphylococcus aureus in vitro but do not influence the outcome of intraperitoneal S. aureus infection in vivo. Immunology 2014, 143, 155–163. [Google Scholar] [CrossRef]
- Arifuzzaman, M.; Mobley, Y.R.; Choi, H.W.; Bist, P.; Salinas, C.A.; Brown, Z.D.; Chen, S.L.; Staats, H.F.; Abraham, S.N. MRGPR-mediated activation of local mast cells clears cutaneous bacterial infection and protects against reinfection. Sci. Adv. 2019, 5, eaav0216. [Google Scholar] [CrossRef] [Green Version]
- Von Köckritz-Blickwede, M.; Goldmann, O.; Thulin, P.; Heinemann, K.; Norrby-Teglund, A.; Rohde, M.; Medina, E. Phagocytosis-independent antimicrobial activity of mast cells by means of extracellular trap formation. Blood 2008, 111, 3070–3080. [Google Scholar] [CrossRef] [PubMed]
- Möllerherm, H.; von Köckritz-Blickwede, M.; Branitzki-Heinemann, K. Antimicrobial activity of mast cells: Role and relevance of extracellular DNA traps. Front. Immunol. 2016, 7, 265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.L.; Wang, Z.; Igawa, S.; Choi, J.E.; Werbel, T.; Di Nardo, A. Lipocalin 2: A New Antimicrobial in Mast Cells. Int. J. Mol. Sci. 2019, 20, 2380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Nardo, A.; Yamasaki, K.; Dorschner, R.A.; Lai, Y.; Gallo, R.L. Mast Cell Cathelicidin Antimicrobial Peptide Prevents Invasive Group A Streptococcus Infection of the Skin. J. Immunol. 2008, 180, 7565–7573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, Z.; Zhang, D.; Lu, B.; Zhou, W.; Wang, D. Activation of mast cells in skin abscess induced by Staphylococcus aureus (S. aureus) infection in mice. Res. Vet. Sci. 2018, 118, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Oscherwitz, J.; Cease, K.B.; Chan, S.M.; Muñoz-Planillo, R.; Hasegawa, M.; Villaruz, A.E.; Cheung, G.Y.C.; McGavin, M.J.; Travers, J.B.; et al. Staphylococcus δ-toxin induces allergic skin disease by activating mast cells. Nature 2013, 503, 397–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starkl, P.; Watzenboeck, M.L.; Popov, L.M.; Zahalka, S.; Hladik, A.; Lakovits, K.; Radhouani, M.; Haschemi, A.; Marichal, T.; Reber, L.L.; et al. IgE Effector Mechanisms, in Concert with Mast Cells, Contribute to Acquired Host Defense against Staphylococcusaureus. Immunity 2020, 53, 793–804.e9. [Google Scholar] [CrossRef]
- Pundir, P.; Liu, R.; Vasavda, C.; Serhan, N.; Limjunyawong, N.; Yee, R.; Zhan, Y.; Dong, X.X.; Wu, X.; Zhang, Y.; et al. A Connective Tissue Mast-Cell-Specific Receptor Detects Bacterial Quorum-Sensing Molecules and Mediates Antibacterial Immunity. Cell Host Microbe 2019, 26, 114–122.e8. [Google Scholar] [CrossRef] [PubMed]
- Nakano, N.; Nishiyama, C.; Kanada, S.; Niwa, Y.; Shimokawa, N.; Ushio, H.; Nishiyama, M.; Okumura, K.; Ogawa, H. Involvement of mast cells in IL-12/23 p40 production is essential for survival from polymicrobial infections. Blood 2007, 109, 4846–4855. [Google Scholar] [CrossRef] [Green Version]
- Marshall, J.S.; Portales-Cervantes, L.; Leong, E. Mast cell responses to viruses and pathogen products. Int. J. Mol. Sci. 2019, 20, 4241. [Google Scholar] [CrossRef] [Green Version]
- Jolly, S.; Detilleux, J.; Desmecht, D. Extensive mast cell degranulation in bovine respiratory syncytial virus-associated paroxystic respiratory distress syndrome. Vet. Immunol. Immunopathol. 2004, 97, 125–136. [Google Scholar] [CrossRef]
- Shirato, K.; Taguchi, F. Mast cell degranulation is induced by A549 airway epithelial cell infected with respiratory syncytial virus. Virology 2009, 386, 88–93. [Google Scholar] [CrossRef]
- Graziano, F.M.; Tilton, R.; Hirth, T.; Segaloff, D.; Mullins, T.; Dick, E.; Buckner, C.K.; Busse, W.W. The effect of parainfluenza 3 infection on guinea pig basophil and lung mast cell histamine release. Am. Rev. Respir. Dis. 1989, 139, 715–720. [Google Scholar] [CrossRef]
- Xiong, L.; Zhen, S.; Yu, Q.; Gong, Z. HCV-E2 inhibits hepatocellular carcinoma metastasis by stimulating mast cells to secrete exosomal shuttle microRNAs. Oncol. Lett. 2017, 14, 2141–2146. [Google Scholar] [CrossRef] [Green Version]
- Sundstrom, J.B.; Ellis, J.E.; Hair, G.A.; Kirshenbaum, A.S.; Metcalfe, D.D.; Yi, H.; Cardona, A.C.; Lindsay, M.K.; Ansari, A.A. Human tissue mast cells are an inducible reservoir of persistent HIV infection. Blood 2007, 109, 5293–5300. [Google Scholar] [CrossRef] [PubMed]
- Demeure, C.E.; Brahimi, K.; Hacini, F.; Marchand, F.; Péronet, R.; Huerre, M.; St.-Mezard, P.; Nicolas, J.-F.; Brey, P.; Delespesse, G.; et al. Anopheles Mosquito Bites Activate Cutaneous Mast Cells Leading to a Local Inflammatory Response and Lymph Node Hyperplasia. J. Immunol. 2005, 174, 3932–3940. [Google Scholar] [CrossRef] [Green Version]
- Depinay, N.; Hacini, F.; Beghdadi, W.; Peronet, R.; Mécheri, S. Mast Cell-Dependent Down-Regulation of Antigen-Specific Immune Responses by Mosquito Bites. J. Immunol. 2006, 176, 4141–4146. [Google Scholar] [CrossRef] [Green Version]
- Pingen, M.; Bryden, S.R.; Pondeville, E.; Schnettler, E.; Kohl, A.; Merits, A.; Fazakerley, J.K.; Graham, G.J.; McKimmie, C.S. Host Inflammatory Response to Mosquito Bites Enhances the Severity of Arbovirus Infection. Immunity 2016, 44, 1455–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, S.W.; Wu-Hsieh, B.A.; Lin, Y.S.; Chen, W.Y.; Huang, Y.; Anderson, R. The monocyte-macrophage-mast cell axis in dengue pathogenesis. J. Biomed. Sci. 2018, 25, 77. [Google Scholar] [CrossRef]
- Messina, J.P.; Brady, O.J.; Golding, N.; Kraemer, M.U.G.; Wint, G.R.W.; Ray, S.E.; Pigott, D.M.; Shearer, F.M.; Johnson, K.; Earl, L.; et al. The current and future global distribution and population at risk of dengue. Nat. Microbiol. 2019, 4, 1508–1515. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.T.; Wan, S.W.; Chang, Y.C.; Lee, C.K.; Wu-Hsieh, B.A.; Anderson, R.; Lin, Y.S. Antibodies against nonstructural protein 1 protect mice from dengue virus-induced mast cell activation. Lab. Investig. 2017, 97, 602–614. [Google Scholar] [CrossRef] [Green Version]
- Troupin, A.; Shirley, D.; Londono-Renteria, B.; Watson, A.M.; McHale, C.; Hall, A.; Hartstone-Rose, A.; Klimstra, W.B.; Gomez, G.; Colpitts, T.M. A Role for Human Skin Mast Cells in Dengue Virus Infection and Systemic Spread. J. Immunol. 2016, 197, 4382–4391. [Google Scholar] [CrossRef] [Green Version]
- Chu, Y.T.; Wan, S.W.; Anderson, R.; Lin, Y.S. Mast cell-macrophage dynamics in modulation of dengue virus infection in skin. Immunology 2015, 146, 163–172. [Google Scholar] [CrossRef] [Green Version]
- John, A.L.S.; Rathore, A.P.S.; Yap, H.; Ng, M.L.; Metcalfe, D.D.; Vasudevan, S.G.; Abraham, S.N. Immune surveillance by mast cells during dengue infection promotes natural killer (NK) and NKT-cell recruitment and viral clearance. Proc. Natl. Acad. Sci. USA 2011, 108, 9190–9195. [Google Scholar] [CrossRef] [Green Version]
- Furuta, T.; Murao, L.A.; Lan, N.T.P.; Huy, N.T.; Huong, V.T.Q.; Thuy, T.T.; Tham, V.D.; Nga, C.T.P.; Ha, T.T.N.; Ohmoto, Y.; et al. Association of mast cell-derived VEGF and proteases in dengue shock syndrome. PLoS Negl. Trop. Dis. 2012, 6. [Google Scholar] [CrossRef] [Green Version]
- Tissera, H.; Rathore, A.P.S.; Leong, W.Y.; Pike, B.L.; Warkentien, T.E.; Farouk, F.S.; Syenina, A.; Ooi, E.E.; Gubler, D.J.; Wilder-Smith, A.; et al. Chymase level is a predictive biomarker of dengue hemorrhagic fever in pediatric and adult patients. J. Infect. Dis. 2017, 216, 1112–1121. [Google Scholar] [CrossRef] [Green Version]
- Morrison, J.; Rathore, A.P.S.; Mantri, C.K.; Aman, S.A.B.; Nishida, A.; John, A.L.S. Transcriptional Profiling Confirms the Therapeutic Effects of Mast Cell Stabilization in a Dengue Disease Model. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [Green Version]
- Sager, G.; Gabaglio, S.; Sztul, E.; Belov, G.A. Role of host cell secretory machinery in zika virus life cycle. Viruses 2018, 10, 559. [Google Scholar] [CrossRef] [Green Version]
- Rabelo, K.; Gonçalves, A.J.d.S.; de Souza, L.J.; Sales, A.P.; de Lima, S.M.B.; Trindade, G.F.; Ciambarella, B.T.; Amorim Tasmo, N.R.; Diaz, B.L.; de Carvalho, J.J.; et al. Zika Virus Infects Human Placental Mast Cells and the HMC-1 Cell Line, and Triggers Degranulation, Cytokine Release and Ultrastructural Changes. Cells 2020, 9, 975. [Google Scholar] [CrossRef] [PubMed]
- Coish, J.M.; Crozier, R.W.E.; Schieffelin, J.S.; Coorssen, J.R.; Hunter, F.F.; MacNeil, A.J. Mast Cell Infection by Zika Virus and Augmentation by Pre-existing Dengue Virus Immunity. FASEB J. 2020, 34, 1. [Google Scholar] [CrossRef]
- Lu, F.; Huang, S. The roles of mast cells in parasitic protozoan infections. Front. Immunol. 2017, 8, 363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choumet, V.; Attout, T.; Chartier, L.; Khun, H.; Sautereau, J.; Robbe-Vincent, A.; Brey, P.; Huerre, M.; Bain, O. Visualizing Non Infectious and Infectious Anopheles gambiae Blood Feedings in Naive and Saliva-Immunized Mice. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Wilainam, P.; Nintasen, R.; Viriyavejakul, P. Mast cell activation in the skin of Plasmodium falciparum malaria patients. Malar. J. 2015, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, B.; Huang, S.; Chen, X.; Liu, X.B.; Wu, Q.; Wang, Y.; Li, X.; Li, K.; Gao, H.; Cen, S.; et al. Activation of Mast Cells Promote Plasmodium berghei ANKA Infection in Murine Model. Front. Cell. Infect. Microbiol. 2019, 9, 322. [Google Scholar] [CrossRef] [PubMed]
- Saha, B.; Tonkal, A.M.D.J.; Croft, S.; Roy, S. Mast cells at the host-pathogen interface: Host-protection versus immune evasion in leishmaniasis. Clin. Exp. Immunol. 2004, 137, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Bidri, M.; Vouldoukis, I.; Djavad Mossalayi, M.; Debré, P.; Guillosson, J.J.; Mazier, D.; Arock, M. Evidence for direct interaction between mast cells and Leishmania parasites. Parasite Immunol. 1997, 19, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Tuon, F.F.; Amato, V.S.; Bacha, H.A.; AlMusawi, T.; Duarte, M.I.; Neto, V.A. Toll-like receptors and leishmaniasis. Infect. Immun. 2008, 76, 866–872. [Google Scholar] [CrossRef] [Green Version]
- Calabrese, K.S.; Cortada, V.M.C.L.; Dorval, M.E.C.; Souza Lima, M.A.A.; Oshiro, E.T.; Souza, C.S.F.; Silva-Almeida, M.; Carvalho, L.O.P.; Gonçalves da Costa, S.C.; Abreu-Silva, A.L. Leishmania (Leishmania) infantum/chagasi: Histopathological aspects of the skin in naturally infected dogs in two endemic areas. Exp. Parasitol. 2010, 124, 253–257. [Google Scholar] [CrossRef]
- Maurer, M.; Lopez Kostka, S.; Siebenhaar, F.; Moelle, K.; Metz, M.; Knop, J.; Stebut, E. Skin mast cells control T cell-dependent host defense in Leishmania major infections. FASEB J. 2006, 20, 2460–2467. [Google Scholar] [CrossRef]
- Naqvi, N.; Ahuja, K.; Selvapandiyan, A.; Dey, R.; Nakhasi, H.; Puri, N. Role of Mast Cells in clearance of Leishmania through extracellular trap formation. Sci. Rep. 2017, 7, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, C.; Wolff, S.; Zapf, T.; Raifer, H.; Feyerabend, T.B.; Bollig, N.; Camara, B.; Trier, C.; Schleicher, U.; Rodewald, H.R.; et al. Mast cells have no impact on cutaneous leishmaniasis severity and related Th2 differentiation in resistant and susceptible mice. Eur. J. Immunol. 2016, 46, 114–121. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Q.; Luo, Y.; Scheffel, J.; Zhao, Z.; Maurer, M. The complex role of mast cells in fungal infections. Exp. Dermatol. 2019, 28, 749–755. [Google Scholar] [CrossRef] [Green Version]
- Croft, C.A.; Culibrk, L.; Moore, M.M.; Tebbutt, S.J. Interactions of Aspergillus fumigatus conidia with airway epithelial cells: A critical review. Front. Microbiol. 2016, 7, 472. [Google Scholar] [CrossRef] [Green Version]
- Radsak, M.; Platzbecker, U.; Schmidt, C.S.; Hofmann, W.K.; Nolte, F. Infectious complications in patients with myelodysplastic syndromes: A review of the literature with emphasis on patients treated with 5-azacitidine. Eur. J. Haematol. 2017, 99, 112–118. [Google Scholar] [CrossRef] [Green Version]
- Margalit, A.; Kavanagh, K. The innate immune response to Aspergillus fumigatus at the alveolar surface. FEMS Microbiol. Rev. 2015, 39, 670–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nieto-Patlán, A.; Campillo-Navarro, M.; Rodríguez-Cortés, O.; Muñoz-Cruz, S.; Wong-Baeza, I.; Estrada-Parra, S.; Estrada-García, I.; Serafín-López, J.; Chacón-Salinas, R. Recognition of Candida albicans by Dectin-1 induces mast cell activation. Immunobiology 2015, 220, 1093–1100. [Google Scholar] [CrossRef] [PubMed]
- Lopes, J.P.; Stylianou, M.; Nilsson, G.; Urban, C.F. Opportunistic pathogen Candida albicans elicits a temporal response in primary human mast cells. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [Green Version]
- Trevisan, E.; Vita, F.; Medic, N.; Soranzo, M.R.; Zabucchi, G.; Borelli, V. Mast Cells Kill Candida albicans in the Extracellular Environment but Spare Ingested Fungi from Death. Inflammation 2014, 37, 2174–2189. [Google Scholar] [CrossRef]
- Carsin, A.; Romain, T.; Ranque, S.; Reynaud-Gaubert, M.; Dubus, J.C.; Mège, J.L.; Vitte, J. Aspergillus fumigatus in cystic fibrosis: An update on immune interactions and molecular diagnostics in allergic bronchopulmonary aspergillosis. Allergy Eur. J. Allergy Clin. Immunol. 2017, 72, 1632–1642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.X.; Fan, L.C.; Li, M.H.; Cao, W.J.; Xu, J.F. Beneficial effects of Omalizumab therapy in allergic bronchopulmonary aspergillosis: A synthesis review of published literature. Respir. Med. 2017, 122, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Urb, M.; Pouliot, P.; Gravelat, F.N.; Olivier, M.; Sheppard, D.C. Aspergillus fumigatus induces immunoglobulin E—Independent mast cell degranulation. J. Infect. Dis. 2009, 200, 464–472. [Google Scholar] [CrossRef] [Green Version]
- Nowicka, D.; Nawrot, U. Contribution of Malassezia spp. to the development of atopic dermatitis. Mycoses 2019, 62, 588–596. [Google Scholar] [CrossRef]
- Barbosa-Lorenzi, V.C.; Peyda, S.; Scheynius, A.; Nilsson, G.; Lunderius-Andersson, C. Curdlan induces selective mast cell degranulation without concomitant release of LTC4, IL-6 or CCL2. Immunobiology 2017, 222, 647–650. [Google Scholar] [CrossRef]
- Ribbing, C.; Engblom, C.; Lappalainen, J.; Lindstedt, K.; Kovanen, P.T.; Karlsson, M.A.; Lundeberg, L.; Johansson, C.; Nilsson, G.; Lunderius-Andersson, C.; et al. Mast cells generated from patients with atopic eczema have enhanced levels of granule mediators and an impaired Dectin-1 expression. Allergy 2011, 66, 110–119. [Google Scholar] [CrossRef]
- Cheng, L.E.; Hartmann, K.; Roers, A.; Krummel, M.F.; Locksley, R.M. Perivascular Mast Cells Dynamically Probe Cutaneous Blood Vessels to Capture Immunoglobulin E. Immunity 2013, 38, 166–175. [Google Scholar] [CrossRef] [Green Version]
- Sala-Cunill, A.; Björkqvist, J.; Senter, R.; Guilarte, M.; Cardona, V.; Labrador, M.; Nickel, K.F.; Butler, L.; Luengo, O.; Kumar, P.; et al. Plasma contact system activation drives anaphylaxis in severe mast cell-mediated allergic reactions. J. Allergy Clin. Immunol. 2015, 135, 1031–1043.e6. [Google Scholar] [CrossRef] [PubMed]
- Stone, S.F.; Brown, S.G.A. Mediators released during human anaphylaxis. Curr. Allergy Asthma Rep. 2012, 12, 33–41. [Google Scholar] [CrossRef]
- Yang, L.; Fu, J.; Zhou, Y. Research Progress in Atopic March. Front. Immunol. 2020, 11, 1907. [Google Scholar] [CrossRef]
- Matsumoto, K.; Iikura, K.; Morita, H.; Saito, H. Barrier dysfunction in the atopic march—How does atopic dermatitis lead to asthma in children? J. Allergy Clin. Immunol. 2020, 145, 1551–1553. [Google Scholar] [CrossRef]
- Czarnowicki, T.; He, H.; Krueger, J.G.; Guttman-Yassky, E. Atopic dermatitis endotypes and implications for targeted therapeutics. J. Allergy Clin. Immunol. 2019, 143, 1–11. [Google Scholar] [CrossRef]
- Leung, D.Y.M. Role of IgE in atopic dermatitis. Curr. Opin. Immunol. 1993, 5, 956–962. [Google Scholar] [CrossRef]
- Grabbe, J.; Haas, N.; Hamann, K.; Kolde, G.; Hakimi, J.; Czarnetzki, B.M. Demonstration of the high-affinity IgE receptor on human Langerhans cells in normal and diseased skin. Br. J. Dermatol. 1993, 129, 120–123. [Google Scholar] [CrossRef]
- Barata, L.T.; Ying, S.; Meng, Q.; Barkans, J.; Rajakulasingam, K.; Durham, S.R.; Kay, A.B. IL-4- and IL-5-positive T lymphocytes, eosinophils, and mast cells in allergen-induced late-phase cutaneous reactions in atopic subjects. J. Allergy Clin. Immunol. 1998, 101, 222–230. [Google Scholar] [CrossRef]
- Imayama, S.; Shibata, Y.; Hori, Y. Epidermal mast cells in atopic dermatitis. Lancet 1995, 346, 1559. [Google Scholar] [CrossRef]
- Groneberg, D.A.; Bester, C.; Grützkau, A.; Serowka, F.; Fischer, A.; Henz, B.M.; Welker, P. Mast cells and vasculature in atopic dermatitis—Potential stimulus of neoangiogenesis. Allergy Eur. J. Allergy Clin. Immunol. 2005, 60, 90–97. [Google Scholar] [CrossRef]
- Obara, W.; Kawa, Y.; Ra, C.; Nishioka, K.; Soma, Y.; Mizoguchi, M. T cells and mast cells as a major source of interleukin-13 in atopic dermatitis. Dermatology 2002, 205, 11–17. [Google Scholar] [CrossRef]
- Babina, M.; Guhl, S.; Artuc, M.; Zuberbier, T. IL-4 and human skin mast cells revisited: Reinforcement of a pro-allergic phenotype upon prolonged exposure. Arch. Dermatol. Res. 2016, 308, 665–670. [Google Scholar] [CrossRef]
- Suurmond, J.; Habets, K.L.L.; Tatum, Z.; Schonkeren, J.J.; Hoen, P.A.C.T.; Huizinga, T.W.J.; Laros, J.F.J.; Toes, R.E.M.; Kurreeman, F. Repeated FcεRI triggering reveals modified mast cell function related to chronic allergic responses in tissue. J. Allergy Clin. Immunol. 2016, 138, 869–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taube, C.; Stassen, M. Mast cells and mast cell-derived factors in the regulation of allergic sensitization. Chem. Immunol. Allergy 2008, 94, 58–66. [Google Scholar] [PubMed]
- Leyva-Castillo, J.M.; Das, M.; Artru, E.; Yoon, J.; Galand, C.; Geha, R.S. Mast cell–derived IL-13 downregulates IL-12 production by skin dendritic cells to inhibit the TH1 cell response to cutaneous antigen exposure. J. Allergy Clin. Immunol. 2021. [Google Scholar] [CrossRef]
- Alenius, H.; Laouini, D.; Woodward, A.; Mizoguchi, E.; Bhan, A.K.; Castigli, E.; Oettgen, H.C.; Geha, R.S. Mast cells regulate IFN-γ expression in the skin and circulating IgE levels in allergen-induced skin inflammation. J. Allergy Clin. Immunol. 2002, 109, 106–113. [Google Scholar] [CrossRef]
- Gordon, J.R. Monocyte chemoattractant peptide-1 expression during cutaneous allergic reactions in mice is mast cell dependent and largely mediates the monocyte recruitment response. J. Allergy Clin. Immunol. 2000, 106, 110–116. [Google Scholar] [CrossRef]
- Steinhoff, M.; Steinhoff, A.; Homey, B.; Luger, T.A.; Schneider, S.W. Role of vasculature in atopic dermatitis. J. Allergy Clin. Immunol. 2006, 118, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Hammad, H.; Lambrecht, B.N. Barrier Epithelial Cells and the Control of Type 2 Immunity. Immunity 2015, 43, 29–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holgate, S.T. The epithelium takes centre stage in asthma and atopic dermatitis. Trends Immunol. 2007, 28, 248–251. [Google Scholar] [CrossRef] [PubMed]
- Allakhverdi, Z.; Comeau, M.R.; Jessup, H.K.; Yoon, B.R.P.; Brewer, A.; Chartier, S.; Paquette, N.; Ziegler, S.F.; Sarfati, M.; Delespesse, G. Thymic stromal lymphopoietin is released by human epithelial cells in response to microbes, trauma, or inflammation and potently activates mast cells. J. Exp. Med. 2007, 204, 253–258. [Google Scholar] [CrossRef]
- Jariwala, S.P.; Abrams, E.; Benson, A.; Fodeman, J.; Zheng, T. The role of thymic stromal lymphopoietin in the immunopathogenesis of atopic dermatitis. Clin. Exp. Allergy 2011, 41, 1515–1520. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, N.; Sae-Wong, C.; Kangsanant, S.; Nabe, T.; Yoshino, S. Thymic stromal lymphopoietin-induced interleukin-17A is involved in the development of IgE-mediated atopic dermatitis-like skin lesions in mice. Immunology 2015, 146, 568–581. [Google Scholar] [CrossRef] [Green Version]
- Babina, M.; Wang, Z.; Franke, K.; Zuberbier, T. Thymic Stromal Lymphopoietin Promotes MRGPRX2-Triggered Degranulation of Skin Mast Cells in a STAT5-Dependent Manner with Further Support from JNK. Cells 2021, 10, 102. [Google Scholar] [CrossRef] [PubMed]
- Moon, P.D.; Kim, H.M. Thymic stromal lymphopoietin is expressed and produced by caspase-1/NF-κB pathway in mast cells. Cytokine 2011, 54, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Liew, F.Y.; Pitman, N.I.; McInnes, I.B. Disease-associated functions of IL-33: The new kid in the IL-1 family. Nat. Rev. Immunol. 2010, 10, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Saluja, R.; Zoltowska, A.; Ketelaar, M.E.; Nilsson, G. IL-33 and Thymic Stromal Lymphopoietin in mast cell functions. Eur. J. Pharmacol. 2016, 778, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Drube, S.; Kraft, F.; Dudeck, J.; Müller, A.-L.; Weber, F.; Göpfert, C.; Meininger, I.; Beyer, M.; Irmler, I.; Häfner, N.; et al. MK2/3 Are Pivotal for IL-33–Induced and Mast Cell–Dependent Leukocyte Recruitment and the Resulting Skin Inflammation. J. Immunol. 2016, 197, 3662–3668. [Google Scholar] [CrossRef]
- Imai, Y.; Yasuda, K.; Sakaguchi, Y.; Haneda, T.; Mizutani, H.; Yoshimoto, T.; Nakanishi, K.; Yamanishi, K. Skin-specific expression of IL-33 activates group 2 innate lymphoid cells and elicits atopic dermatitis-like inflammation in mice. Proc. Natl. Acad. Sci. USA 2013, 110, 13921–13926. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Egusa, C.; Maeda, T.; Numata, T.; Nakano, N.; Nishiyama, C.; Tsuboi, R. Il-33 promotes mhc class ii expression in murine mast cells. Immun. Inflamm. Dis. 2015, 3, 196–208. [Google Scholar] [CrossRef]
- Hao, Y.; Peng, B.; Che, D.; Zheng, Y.; Kong, S.; Liu, R.; Shi, J.; Han, H.; Wang, J.; Cao, J.; et al. Imiquimod-related dermatitis is mainly mediated by mast cell degranulation via Mas-related G-protein coupled receptor B2. Int. Immunopharmacol. 2020, 81. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, H.; Gupta, K.; Ali, H. Roles of Mas-related G protein–coupled receptor X2 on mast cell–mediated host defense, pseudoallergic drug reactions, and chronic inflammatory diseases. J. Allergy Clin. Immunol. 2016, 138, 700–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kühn, H.; Kolkhir, P.; Babina, M.; Düll, M.; Frischbutter, S.; Fok, J.S.; Jiao, Q.; Metz, M.; Scheffel, J.; Wolf, K.; et al. Mas-related G protein–coupled receptor X2 and its activators in dermatologic allergies. J. Allergy Clin. Immunol. 2021, 147, 456–469. [Google Scholar] [CrossRef]
- Thapaliya, M.; Chompunud Na Ayudhya, C.; Amponnawarat, A.; Roy, S.; Ali, H. Mast Cell-Specific MRGPRX2: A Key Modulator of Neuro-Immune Interaction in Allergic Diseases. Curr. Allergy Asthma Rep. 2021, 21, 3. [Google Scholar] [CrossRef] [PubMed]
- Kwang, H.L.; Ji, Y.K.; Kang, D.S.; Yoo, J.C.; Lee, W.J.; Jai, Y.R. Increased expression of endothelial cell adhesion molecules due to mediator release from human foreskin mast cells stimulated by autoantibodies in chronic urticaria sera. J. Investig. Dermatol. 2002, 118, 658–663. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Babina, M. MRGPRX2 signals its importance in cutaneous mast cell biology: Does MRGPRX2 connect mast cells and atopic dermatitis? Exp. Dermatol. 2020, 29, 1104–1111. [Google Scholar] [CrossRef]
- Serhan, N.; Basso, L.; Sibilano, R.; Petitfils, C.; Meixiong, J.; Bonnart, C.; Reber, L.L.; Marichal, T.; Starkl, P.; Cenac, N.; et al. House dust mites activate nociceptor–mast cell clusters to drive type 2 skin inflammation. Nat. Immunol. 2019, 20, 1435–1443. [Google Scholar] [CrossRef] [PubMed]
- Corbière, A.; Loste, A.; Gaudenzio, N. MRGPRX2 sensing of cationic compounds—A bridge between nociception and skin diseases? Exp. Dermatol. 2020, 30. [Google Scholar] [CrossRef]
- Siiskonen, H.; Harvima, I. Mast Cells and Sensory Nerves Contribute to Neurogenic Inflammation and Pruritus in Chronic Skin Inflammation. Front. Cell. Neurosci. 2019, 13, 422. [Google Scholar] [CrossRef]
- Ohsawa, Y.; Hirasawa, N. The role of histamine H1 and H4 receptors in atopic dermatitis: From basic research to clinical study. Allergol. Int. 2014, 63, 533–542. [Google Scholar] [CrossRef] [Green Version]
- Cowden, J.M.; Zhang, M.; Dunford, P.J.; Thurmond, R.L. The histamine H4 receptor mediates inflammation and pruritus in Th2-dependent dermal inflammation. J. Investig. Dermatol. 2010, 130, 1023–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Pan, W.H.; Wang, X.R.; Liu, Y.; Chen, M.; Xu, X.G.; Liao, W.Q.; Hu, J.H. Tryptase and protease-activated receptor-2 stimulate scratching behavior in a murine model of ovalbumin-induced atopic-like dermatitis. Int. Immunopharmacol. 2015, 28, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Steinhoff, M.; Buddenkotte, J.; Lerner, E.A. Role of mast cells and basophils in pruritus. Immunol. Rev. 2018, 282, 248–264. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.; Harvima, I.T. Mast cell-neural interactions contribute to pain and itch. Immunol. Rev. 2018, 282, 168–187. [Google Scholar] [CrossRef]
- Nakashima, C.; Ishida, Y.; Kitoh, A.; Otsuka, A.; Kabashima, K. Interaction of peripheral nerves and mast cells, eosinophils, and basophils in the development of pruritus. Exp. Dermatol. 2019, 28, 1405–1411. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.L.B.; Kim, B.S. Pruritus in allergy and immunology. J. Allergy Clin. Immunol. 2019, 144, 353–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ando, T.; Matsumoto, K.; Namiranian, S.; Yamashita, H.; Glatthorn, H.; Kimura, M.; Dolan, B.R.; Lee, J.J.; Galli, S.J.; Kawakami, Y.; et al. Mast cells are required for full expression of allergen/SEB-Induced skin inflammation. J. Investig. Dermatol. 2013, 133, 2695–2705. [Google Scholar] [CrossRef] [Green Version]
- Savinko, T.; Matikainen, S.; Saarialho-Kere, U.; Lehto, M.; Wang, G.; Lehtimäki, S.; Karisola, P.; Reunala, T.; Wolff, H.; Lauerma, A.; et al. IL-33 and ST2 in atopic dermatitis: Expression profiles and modulation by triggering factors. J. Investig. Dermatol. 2012, 132, 1392–1400. [Google Scholar] [CrossRef] [Green Version]
- Leyva-Castillo, J.M.; Galand, C.; Kam, C.; Burton, O.; Gurish, M.; Musser, M.A.; Goldsmith, J.D.; Hait, E.; Nurko, S.; Brombacher, F.; et al. Mechanical Skin Injury Promotes Food Anaphylaxis by Driving Intestinal Mast Cell Expansion. Immunity 2019, 50, 1262–1275.e4. [Google Scholar] [CrossRef] [PubMed]
- Galand, C.; Leyva-Castillo, J.M.; Yoon, J.; Han, A.; Lee, M.S.; McKenzie, A.N.J.; Stassen, M.; Oyoshi, M.K.; Finkelman, F.D.; Geha, R.S. IL-33 promotes food anaphylaxis in epicutaneously sensitized mice by targeting mast cells. J. Allergy Clin. Immunol. 2016, 138, 1356–1366. [Google Scholar] [CrossRef] [Green Version]
- Thangam, E.B.; Jemima, E.A.; Singh, H.; Baig, M.S.; Khan, M.; Mathias, C.B.; Church, M.K.; Saluja, R. The role of histamine and histamine receptors in mast cell-mediated allergy and inflammation: The hunt for new therapeutic targets. Front. Immunol. 2018, 9, 1873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, H.T.T.; Sugita, K.; Akdis, C.A. Novel Biologicals for the Treatment of Allergic Diseases and Asthma. Curr. Allergy Asthma Rep. 2016, 16, 70. [Google Scholar] [CrossRef] [PubMed]
- Boyman, O.; Kaegi, C.; Akdis, M.; Bavbek, S.; Bossios, A.; Chatzipetrou, A.; Eiwegger, T.; Firinu, D.; Harr, T.; Knol, E.; et al. EAACI IG Biologicals task force paper on the use of biologic agents in allergic disorders. Allergy Eur. J. Allergy Clin. Immunol. 2015, 70, 727–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, B.C.L.; Lam, C.W.K.; Tam, L.S.; Wong, C.K. IL33: Roles in allergic inflammation and therapeutic perspectives. Front. Immunol. 2019, 10, 364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peiser, M.; Tralau, T.; Heidler, J.; Api, A.M.; Arts, J.H.E.; Basketter, D.A.; English, J.; Diepgen, T.L.; Fuhlbrigge, R.C.; Gaspari, A.A.; et al. Allergic contact dermatitis: Epidemiology, molecular mechanisms, in vitro methods and regulatory aspects. Cell. Mol. Life Sci. 2012, 69, 763–781. [Google Scholar] [CrossRef] [Green Version]
- Ring, J. Occupational skin disease—A major health problem in Europe. J. Eur. Acad. Dermatol. Venereol. 2017, 31, 919–920. [Google Scholar] [CrossRef] [Green Version]
- Diepgen, T.L.; Ofenloch, R.F.; Bruze, M.; Bertuccio, P.; Cazzaniga, S.; Coenraads, P.J.; Elsner, P.; Goncalo, M.; Svensson, Å.; Naldi, L. Prevalence of contact allergy in the general population in different European regions. Br. J. Dermatol. 2016, 174, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.F.; Rustemeyer, T.; Thyssen, J.P. Recent advances in understanding and managing contact dermatitis. F1000Research 2018, 7, F1000. [Google Scholar] [CrossRef] [Green Version]
- Martin, S.F. Immunological mechanisms in allergic contact dermatitis. Curr. Opin. Allergy Clin. Immunol. 2015, 15, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Fujisawa, H.; Zhuang, L.; Freed, I.; Howell, B.G.; Shahid, S.; Shivji, G.M.; Mak, T.W.; Sauder, D.N. CD4 + Th1 and CD8 + Type 1 Cytotoxic T Cells Both Play a Crucial Role in the Full Development of Contact Hypersensitivity. J. Immunol. 2000, 165, 6783–6790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaspari, A.A.; Katz, S.I.; Martin, S.F. Contact Hypersensitivity. Curr. Protoc. Immunol. 2016, 113, 4.2.1–4.2.7. [Google Scholar] [CrossRef]
- Kaplan, D.H.; Igyártó, B.Z.; Gaspari, A.A. Early immune events in the induction of allergic contact dermatitis. Nat. Rev. Immunol. 2012, 12, 114–124. [Google Scholar] [CrossRef]
- Silvestre, M.C.; dos Reis, V.M.S.; Sato, M.N. Innate immunity and effector and regulatory mechanisms involved in allergic contact dermatitis. An. Bras. Dermatol. 2018, 93, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Vocanson, M.; Hennino, A.; Rozières, A.; Poyet, G.; Nicolas, J.F. Effector and regulatory mechanisms in allergic contact dermatitis. Allergy Eur. J. Allergy Clin. Immunol. 2009, 64, 1699–1714. [Google Scholar] [CrossRef]
- Bonneville, M.; Chavagnac, C.; Vocanson, M.; Rozieres, A.; Benetiere, J.; Pernet, I.; Denis, A.; Nicolas, J.F.; Hennino, A. Skin contact irritation conditions the development and severity of allergic contact dermatitis. J. Investig. Dermatol. 2007, 127, 1430–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Askenase, P.W.; Van Loveren, H.; Kraeuter-Kops, S.; Ron, Y.; Meade, R.; Theoharides, T.C.; Nordlund, J.J.; Scovern, H.; Gerhson, M.D.; Ptak, W. Defective elicitation of delayed-type hypersensitivity in W/Wv and SI/SId mast cell-deficient mice. J. Immunol. 1983, 131, 2687–2694. [Google Scholar]
- Thomas, W.R.; Schrader, J.W. Delayed hypersensitivity in mast-cell-deficient mice. J. Immunol. 1983, 130, 2565–2567. [Google Scholar] [PubMed]
- Mekori, Y.A.; Galli, S.J. Undiminished immunologic tolerance to contact sensitivity in mast cell-deficient W/Wv and Sl/Sld mice. J. Immunol. 1985, 135, 879–885. [Google Scholar]
- Kerdel, F.A.; Belsito, D.V.; Scotto-Chinnici, R.; Soter, N.A. Mast cell participation during the elicitation of murine allergic contact hypersensitivity. J. Investig. Dermatol. 1987, 88, 686–690. [Google Scholar] [CrossRef] [Green Version]
- Honda, T.; Egawa, G.; Grabbe, S.; Kabashima, K. Update of immune events in the murine contact hypersensitivity model: Toward the understanding of allergic contact dermatitis. J. Investig. Dermatol. 2013, 133, 303–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, M.; Nunomura, S.; Gon, Y.; Endo, D.; Kishiro, S.; Fukunaga, M.; Kitahata, Y.; Terui, T.; Ra, C. Abrogation of high-affinity IgE receptor-mediated mast cell activation at the effector phase prevents contact hypersensitivity to oxazolone. J. Investig. Dermatol. 2010, 130, 725–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryce, P.J.; Miller, M.L.; Miyajima, I.; Tsai, M.; Galli, S.J.; Oettgen, H.C. Immune sensitization in the skin is enhanced by antigen-independent effects of IgE. Immunity 2004, 20, 381–392. [Google Scholar] [CrossRef] [Green Version]
- Kambe, N.; Nakamura, Y.; Saito, M.; Nishikomori, R. The inflammasome, an innate immunity guardian, participates in skin urticarial reactions and contact hypersensitivity. Allergol. Int. 2010, 59, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Mraz, V.; Geisler, C.; Bonefeld, C.M. Dendritic Epidermal T Cells in Allergic Contact Dermatitis. Front. Immunol. 2020, 11, 874. [Google Scholar] [CrossRef] [PubMed]
- Bauer, B.; Andersson, S.I.; Stenfeldt, A.L.; Simonsson, C.; Bergström, J.; Ericson, M.B.; Jonsson, C.A.; Broo, K.S. Modification and expulsion of keratins by human epidermal keratinocytes upon hapten exposure in vitro. Chem. Res. Toxicol. 2011, 24, 737–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, A.; Nicholas, B.; Darley, R.; Parkinson, E.; Teo, Y.; Aleksic, M.; Maxwell, G.; Elliott, T.; Ardern-Jones, M.; Skipp, P. Characterization of the Class I MHC Peptidome Resulting From DNCB Exposure of HaCaT Cells. Toxicol. Sci. 2021, 180, 136–147. [Google Scholar] [CrossRef]
- Simonsson, C.; Andersson, S.I.; Stenfeldt, A.L.; Bergström, J.; Bauer, B.; Jonsson, C.A.; Ericson, M.B.; Broo, K.S. Caged fluorescent haptens reveal the generation of cryptic epitopes in allergic contact dermatitis. J. Investig. Dermatol. 2011, 131, 1486–1493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, H.; Gaide, O.; Pétrilli, V.; Martinon, F.; Contassot, E.; Roques, S.; Kummer, J.A.; Tschopp, J.; French, L.E. Activation of the IL-1β-processing inflammasome is involved in contact hypersensitivity. J. Investig. Dermatol. 2007, 127, 1956–1963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sebastião, A.I.; Ferreira, I.; Brites, G.; Silva, A.; Neves, B.M.; Teresa Cruz, M. NLRP3 Inflammasome and Allergic Contact Dermatitis: A Connection to Demystify. Pharmaceutics 2020, 12, 867. [Google Scholar] [CrossRef]
- Esser, P.R.; Wölfle, U.; Dürr, C.; von Loewenich, F.D.; Schempp, C.M.; Freudenberg, M.A.; Jakob, T.; Martin, S.F. Contact Sensitizers Induce Skin Inflammation via ROS Production and Hyaluronic Acid Degradation. PLoS ONE 2012, 7, e41340. [Google Scholar] [CrossRef] [Green Version]
- Weber, F.C.; Esser, P.R.; Müller, T.; Ganesan, J.; Pellegatti, P.; Simon, M.M.; Zeiser, R.; Idzko, M.; Jakob, T.; Martin, S.F. Lack of the purinergic receptor P2X7 results in resistance to contact hypersensitivity. J. Exp. Med. 2010, 207, 2609–2619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onami, K.; Kimura, Y.; Ito, Y.; Yamauchi, T.; Yamasaki, K.; Aiba, S. Nonmetal haptens induce ATP release from keratinocytes through opening of pannexin hemichannels by reactive oxygen species. J. Investig. Dermatol. 2014, 134, 1951–1960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucia Minciullo, P.; Guarneri, F.; MinciuLLo, P.; Calapai, F.; Cannavò, P. IL-31 and IL-33 circulating levels in allergic contact dermatitis. Eur. Ann. Allergy Clin. Immunol. 2015, 47, 156–158. [Google Scholar]
- Taniguchi, K.; Yamamoto, S.; Hitomi, E.; Inada, Y.; Suyama, Y.; Sugioka, T.; Hamasaki, Y. Interleukin 33 is induced by Tumor necrosis factor alpha and interferon gamma in keratinocytes and contributes to allergic contact dermatitis. J. Investig. Allergol. Clin. Immunol. 2013, 23, 428–434. [Google Scholar] [PubMed]
- Liu, B.; Tai, Y.; Achanta, S.; Kaelberer, M.M.; Caceres, A.I.; Shao, X.; Fang, J.; Jordt, S.E. IL-33/ST2 signaling excites sensory neurons and mediates itch response in a mouse model of Poison ivy contact allergy. Proc. Natl. Acad. Sci. USA 2016, 113, E7572–E7579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imai, Y.; Yasuda, K.; Sakaguchi, Y.; Futatsugi-Yumikura, S.; Yoshimoto, T.; Nakanishi, K.; Yamanishi, K. Immediate-type contact hypersensitivity is reduced in interleukin-33 knockout mice. J. Dermatol. Sci. 2014, 74, 159–161. [Google Scholar] [CrossRef] [PubMed]
- Seike, M.; Furuya, K.; Omura, M.; Hamada-Watanabe, K.; Matsushita, A.; Ohtsu, H. Histamine H4 receptor antagonist ameliorates chronic allergic contact dermatitis induced by repeated challenge. Allergy Eur. J. Allergy Clin. Immunol. 2010, 65, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Weber, F.C.; Németh, T.; Csepregi, J.Z.; Dudeck, A.; Roers, A.; Ozsvári, B.; Oswald, E.; Puskás, L.G.; Jakob, T.; Mócsai, A.; et al. Neutrophils are required for both the sensitization and elicitation phase of contact hypersensitivity. J. Exp. Med. 2015, 212, 15–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.W.; Tedla, N.; Lloyd, A.R.; Wakefield, D.; McNeil, H.P. Mast cell activation and migration to lymph nodes during induction of an immune response in mice. J. Clin. Investig. 1998, 102, 1617–1626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otsuka, A.; Kubo, M.; Honda, T.; Egawa, G.; Nakajima, S.; Tanizaki, H.; Kim, B.; Matsuoka, S.; Watanabe, T.; Nakae, S.; et al. Requirement of Interaction between Mast Cells and Skin Dendritic Cells to Establish Contact Hypersensitivity. PLoS ONE 2011, 6, e25538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suto, H.; Nakae, S.; Kakurai, M.; Sedgwick, J.D.; Tsai, M.; Galli, S.J. Mast Cell-Associated TNF Promotes Dendritic Cell Migration. J. Immunol. 2006, 176, 4102–4112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudeck, J.; Ghouse, S.M.; Lehmann, C.H.K.; Hoppe, A.; Schubert, N.; Nedospasov, S.A.; Dudziak, D.; Dudeck, A. Mast-Cell-Derived TNF Amplifies CD8+ Dendritic Cell Functionality and CD8+ T Cell Priming. Cell Rep. 2015, 13, 399–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakurai, M.; Monteforte, R.; Suto, H.; Tsai, M.; Nakae, S.; Galli, S.J. Mast cell-derived tumor necrosis factor can promote nerve fiber elongation in the skin during contact hypersensitivity in mice. Am. J. Pathol. 2006, 169, 1713–1721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meixiong, J.; Anderson, M.; Limjunyawong, N.; Sabbagh, M.F.; Hu, E.; Mack, M.R.; Oetjen, L.K.; Wang, F.; Kim, B.S.; Dong, X. Activation of Mast-Cell-Expressed Mas-Related G-Protein-Coupled Receptors Drives Non-histaminergic Itch. Immunity 2019, 50, 1163–1171.e5. [Google Scholar] [CrossRef]
- Shimada, Y.; Hasegawa, M.; Kaburagi, Y.; Hamaguchi, Y.; Komura, K.; Saito, E.; Takehara, K.; Steeber, D.A.; Tedder, T.F.; Sato, S. L-Selectin or ICAM-1 Deficiency Reduces an Immediate-Type Hypersensitivity Response by Preventing Mast Cell Recruitment in Repeated Elicitation of Contact Hypersensitivity. J. Immunol. 2003, 170, 4325–4334. [Google Scholar] [CrossRef] [Green Version]
- Gimenez-Rivera, V.-A.; Siebenhaar, F.; Zimmermann, C.; Siiskonen, H.; Metz, M.; Maurer, M. Mast Cells Limit the Exacerbation of Chronic Allergic Contact Dermatitis in Response to Repeated Allergen Exposure. J. Immunol. 2016, 197, 4240–4246. [Google Scholar] [CrossRef] [Green Version]
- Babina, M.; Wang, Z.; Franke, K.; Guhl, S.; Artuc, M.; Zuberbier, T. Yin-Yang of IL-33 in Human Skin Mast Cells: Reduced Degranulation, but Augmented Histamine Synthesis through p38 Activation. J. Investig. Dermatol. 2019, 139, 1516–1525.e3. [Google Scholar] [CrossRef] [PubMed]
- Su, W.; Fan, H.; Chen, M.; Wang, J.; Brand, D.; He, X.; Quesniaux, V.; Ryffel, B.; Zhu, L.; Liang, D.; et al. Induced CD4+ forkhead box protein-positive T cells inhibit mast cell function and established contact hypersensitivity through TGF-β1. J. Allergy Clin. Immunol. 2012, 130. [Google Scholar] [CrossRef] [PubMed]
- Gaudenzio, N.; Marichal, T.; Galli, S.J.; Reber, L.L. Genetic and imaging approaches reveal pro-inflammatory and immunoregulatory roles of mast cells in contact hypersensitivity. Front. Immunol. 2018, 9, 1275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galli, S.J.; Tsai, M. Mast cells in allergy and infection: Versatile effector and regulatory cells in innate and adaptive immunity. Eur. J. Immunol. 2010, 40, 1843–1851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galli, S.J.; Kalesnikoff, J.; Grimbaldeston, M.A.; Piliponsky, A.M.; Williams, C.M.M.; Tsai, M. Mast cells as “tunable” effector and immunoregulatory cells: Recent advances. Annu. Rev. Immunol. 2005, 23, 749–786. [Google Scholar] [CrossRef] [PubMed]
- Norman, M.U.; Hwang, J.; Hulliger, S.; Bonder, C.S.; Yamanouchi, J.; Santamaria, P.; Kubes, P. Mast cells regulate the magnitude and the cytokine microenvironment of the contact hypersensitivity response. Am. J. Pathol. 2008, 172, 1638–1649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reber, L.L.; Sibilano, R.; Starkl, P.; Roers, A.; Grimbaldeston, M.A.; Tsai, M.; Gaudenzio, N.; Galli, S.J. Imaging protective mast cells in living mice during severe contact hypersensitivity. JCI Insight 2017, 2. [Google Scholar] [CrossRef] [Green Version]
- Grimbaldeston, M.A.; Nakae, S.; Kalesnikoff, J.; Tsai, M.; Galli, S.J. Mast cell-derived interleukin 10 limits skin pathology in contact dermatitis and chronic irradiation with ultraviolet B. Nat. Immunol. 2007, 8, 1095–1104. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.W.H.S.; Lee, M.B.; Lee, D.; Min, K.Y.; Koo, J.; Kim, H.W.H.S.; Park, Y.H.; Kim, S.J.; Ikutani, M.; Takaki, S.; et al. The regulatory B cell–mediated peripheral tolerance maintained by mast cell IL-5 suppresses oxazolone-induced contact hypersensitivity. Sci. Adv. 2019, 5, 8152–8169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirano, T.; Honda, T.; Kanameishi, S.; Honda, Y.; Egawa, G.; Kitoh, A.; Nakajima, S.; Otsuka, A.; Nomura, T.; Dainichi, T.; et al. PD-L1 on mast cells suppresses effector CD8+ T-cell activation in the skin in murine contact hypersensitivity. J. Allergy Clin. Immunol. 2021. [Google Scholar] [CrossRef]
- Conti, P.; Gallenga, C.E.; Ronconi, G.; Caraffa, A.; Kritas, S.K. Activation of mast cells mediates inflammatory response in psoriasis: Potential new therapeutic approach with IL-37. Dermatol. Ther. 2019, 32, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Greb, J.E.; Goldminz, A.M.; Elder, J.T.; Lebwohl, M.G.; Gladman, D.D.; Wu, J.J.; Mehta, N.N.; Finlay, A.Y.; Gottlieb, A.B. Psoriasis. Nat. Rev. Dis. Prim. 2016, 2, 1–17. [Google Scholar] [CrossRef]
- Brody, I. Mast cell degranulation in the evolution of acute eruptive guttate psoriasis vulgaris. J. Investig. Dermatol. 1984, 82, 460–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, L.J.; Hansen, U.; Kristensen, J.K.; Nielsen, H.; Skov, P.S.; Nielsen, H.J. Studies on mast cells and histamine release in psoriasis: The effect of ranitidine. Acta Derm. Venereol. 1998, 78, 190–193. [Google Scholar] [CrossRef]
- Oishi, N.; Iwata, H.; Kambe, N.; Kobayashi, N.; Fujimoto, K.; Sato, H.; Hisaka, A.; Ueno, K.; Yamaura, K. Expression of precipitating factors of pruritus found in humans in an imiquimod-induced psoriasis mouse model. Heliyon 2019, 5, e01981. [Google Scholar] [CrossRef] [Green Version]
- Nattkemper, L.A.; Tey, H.L.; Valdes-Rodriguez, R.; Lee, H.; Mollanazar, N.K.; Albornoz, C.; Sanders, K.M.; Yosipovitch, G. The Genetics of Chronic Itch: Gene Expression in the Skin of Patients with Atopic Dermatitis and Psoriasis with Severe Itch. J. Investig. Dermatol. 2018, 138, 1311–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvima, I.T.; Nilsson, G.; Suttle, M.M.; Naukkarinen, A. Is there a role for mast cells in psoriasis? Arch. Dermatol. Res. 2008, 300, 461–478. [Google Scholar] [CrossRef] [Green Version]
- Theoharides, T.C.; Zhang, B.; Kempuraj, D.; Tagen, M.; Vasiadi, M.; Angelidou, A.; Alysandratos, K.D.; Kalogeromitros, D.; Asadi, S.; Stavrianeas, N.; et al. IL-33 augments substance P-induced VEGF secretion from human mast cells and is increased in psoriatic skin. Proc. Natl. Acad. Sci. USA 2010, 107, 4448–4453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balato, A.; Lembo, S.; Mattii, M.; Schiattarella, M.; Marino, R.; De Paulis, A.; Balato, N.; Ayala, F. IL-33 is secreted by psoriatic keratinocytes and induces pro-inflammatory cytokines via keratinocyte and mast cell activation. Exp. Dermatol. 2012, 21, 892–894. [Google Scholar] [CrossRef] [PubMed]
- Mashiko, S.; Bouguermouh, S.; Rubio, M.; Baba, N.; Bissonnette, R.; Sarfati, M. Human mast cells are major IL-22 producers in patients with psoriasis and atopic dermatitis. J. Allergy Clin. Immunol. 2015, 136, 351–359.e1. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.M.; Rubin, C.J.; Khandpur, R.; Wang, J.Y.; Riblett, M.; Yalavarthi, S.; Villanueva, E.C.; Shah, P.; Kaplan, M.J.; Bruce, A.T. Mast Cells and Neutrophils Release IL-17 through Extracellular Trap Formation in Psoriasis. J. Immunol. 2011, 187, 490–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuberbier, T.; Asero, R.; Bindslev-Jensen, C.; Walter Canonica, G.; Church, M.K.; Giménez-Arnau, A.; Grattan, C.E.H.; Kapp, A.; Merk, H.F.; Rogala, B.; et al. EAACI/GA2LEN/EDF/WAO guideline: Definition, classification and diagnosis of urticaria. Allergy Eur. J. Allergy Clin. Immunol. 2009, 64, 1417–1426. [Google Scholar] [CrossRef]
- Jacques, P.; Lavoie, A.; Bédard, P.M.; Brunet, C.; Hebert, J. Chronic idiopathic urticaria: Profiles of skin mast cell histamine release during active disease and remission. J. Allergy Clin. Immunol. 1992, 89, 1139–1143. [Google Scholar] [CrossRef]
- Guida, B.; De Martino, C.; De Martino, S.; Tritto, G.; Patella, V.; Trio, R.; D’Agostino, C.; Pecoraro, P.; D’Agostino, L. Histamine plasma levels and elimination diet in chronic idiopathic urticaria. Eur. J. Clin. Nutr. 2000, 54, 155–158. [Google Scholar] [CrossRef] [Green Version]
- Murphy, G.F.; Austen, K.F.; Fonferko, E.; Sheffer, A.L. Morphologically distinctive forms of cutaneous mast cell degranulation induced by cold and mechanical stimuli: An ultrastructural study. J. Allergy Clin. Immunol. 1987, 80, 603–611. [Google Scholar] [CrossRef]
- Garafalo, J.; Kaplan, A.P. Histamine release and therapy of severe dermatographism. J. Allergy Clin. Immunol. 1981, 68, 103–105. [Google Scholar] [CrossRef]
- Kring Tannert, L.; Stahl Skov, P.; Bjerremann Jensen, L.; Maurer, M.; Bindslev-Jensen, C. Cold urticaria patients exhibit normal skin levels of functional mast cells and histamine after tolerance induction. Dermatology 2012, 224, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, A.P.; Gray, L.; Shaff, R.E.; Horakova, Z.; Beaven, M.A. In vivo studies of mediator release in cold urticaria and cholinergic urticaria. J. Allergy Clin. Immunol. 1975, 55, 394–402. [Google Scholar] [CrossRef]
- Koh, Y.I.; Choi, I.S.; Lee, S.H.; Lee, J.B.; Park, C.H.; Hong, S.N. Localized heat urticaria associated with mast cell and eosinophil degranulation. J. Allergy Clin. Immunol. 2002, 109, 714–715. [Google Scholar] [CrossRef] [PubMed]
- Irwin, R.B.; Lieberman, P.; Friedman, M.M.; Kaliner, M.; Kaplan, R.; Bale, G.; Treadwell, G.; Yoo, T.J. Mediator release in local heat urticaria: Protection with combined H1 and H2 antagonists. J. Allergy Clin. Immunol. 1985, 76, 35–39. [Google Scholar] [CrossRef]
- Hawk, J.L.; Eady, R.A.; Challoner, A.V.; Kobza-Black, A.; Keahey, T.M.; Greaves, M.W. Elevated blood histamine levels and mast cell degranulation in solar urticaria. Br. J. Clin. Pharmacol. 1980, 9, 183–186. [Google Scholar] [CrossRef] [Green Version]
- Mayou, S.C.; Black, A.K.; Eady, R.A.J.; Greaves, M.W. Cholinergic dermographism. Br. J. Dermatol. 1986, 115, 371–377. [Google Scholar] [CrossRef]
- Mekori, Y.A.; Dobozin, B.S.; Schocket, A.L.; Kohler, P.F.; Clark, R.A.F. Delayed Pressure Urticaria Histologically Resembles Cutaneous Late-Phase Reactions. Arch. Dermatol. 1988, 124, 230–235. [Google Scholar] [CrossRef]
- Schaefer, P. Urticaria: Evaluation and Treatment. Am. Fam. Physician 2011, 83, 1078–1084. [Google Scholar] [PubMed]
- Kolkhir, P.; Church, M.K.; Weller, K.; Metz, M.; Schmetzer, O.; Maurer, M. Autoimmune chronic spontaneous urticaria: What we know and what we do not know. J. Allergy Clin. Immunol. 2017, 139, 1772–1781.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurer, M.; Rosén, K.; Hsieh, H.-J.; Saini, S.; Grattan, C.; Gimenéz-Arnau, A.; Agarwal, S.; Doyle, R.; Canvin, J.; Kaplan, A.; et al. Omalizumab for the Treatment of Chronic Idiopathic or Spontaneous Urticaria. N. Engl. J. Med. 2013, 368, 924–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurer, M.; Metz, M.; Brehler, R.; Hillen, U.; Jakob, T.; Mahler, V.; Pföhler, C.; Staubach, P.; Treudler, R.; Wedi, B.; et al. Omalizumab treatment in patients with chronic inducible urticaria: A systematic review of published evidence. J. Allergy Clin. Immunol. 2018, 141, 638–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyden, S.E.; Desai, A.; Cruse, G.; Young, M.L.; Bolan, H.C.; Scott, L.M.; Eisch, A.R.; Long, R.D.; Lee, C.-C.R.; Satorius, C.L.; et al. Vibratory Urticaria Associated with a Missense Variant in ADGRE2. N. Engl. J. Med. 2016, 374, 656–663. [Google Scholar] [CrossRef] [PubMed]
- Wallengren, J.; Möller, H.; Ekman, R. Occurrence of substance P, vasoactive intestinal peptide, and calcitonin gene-related peptide in dermographism and cold urticaria. Arch. Dermatol. Res. 1987, 279, 512–515. [Google Scholar] [CrossRef] [PubMed]
- Borici-Mazi, R.; Kouridakis, S.; Kontou-Fili, K. Cutaneous responses to substance P and calcitonin gene-related peptide in chronic urticaria: The effect of cetirizine and dimethindene. Allergy Eur. J. Allergy Clin. Immunol. 1999, 54, 46–56. [Google Scholar] [CrossRef]
- Fujisawa, D.; Kashiwakura, J.-I.I.; Kita, H.; Kikukawa, Y.; Fujitani, Y.; Sasaki-Sakamoto, T.; Kuroda, K.; Nunomura, S.; Hayama, K.; Terui, T.; et al. Expression of Mas-related gene X2 on mast cells is upregulated in the skin of patients with severe chronic urticaria. J. Allergy Clin. Immunol. 2014, 134, 622–633.e9. [Google Scholar] [CrossRef]
- Toledo, M.A.S.; Gatz, M.; Sontag, S.; Gleixner, K.V.; Eisenwort, G.; Feldberg, K.; Hamouda, A.E.I.; Kluge, F.; Guareschi, R.; Rossetti, G.; et al. Nintedanib targets KIT D816V neoplastic cells derived from induced pluripotent stem cells of systemic mastocytosis. Blood 2021, 137, 2070–2084. [Google Scholar] [CrossRef]
- Chatterjee, A.; Ghosh, J.; Kapur, R. Mastocytosis—A mutated KIT receptor induced myeloproliferative disorder. Oncotarget 2015, 6, 18250–18264. [Google Scholar] [CrossRef]
- Piao, X.; Bernstein, A. A point mutation in the catalytic domain of c-kit induces growth factor independence, tumorigenicity, and differentiation of mast cells. Blood 1996, 87, 3117–3123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, K.; Artuc, M.; Baldus, S.E.; Zirbes, T.K.; Hermes, B.; Thiele, J.; Mekori, Y.A.; Henz, B.M. Expression of Bcl-2 and Bcl-xL in Cutaneous and Bone Marrow Lesions of Mastocytosis. Am. J. Pathol. 2003, 163, 819–826. [Google Scholar] [CrossRef] [Green Version]
- Aichberger, K.J.; Gleixner, K.V.; Mirkina, I.; Cerny-Reiterer, S.; Peter, B.; Ferenc, V.; Kneidinger, M.; Baumgartner, C.; Mayerhofer, M.; Gruze, A.; et al. Identification of proapoptotic Bim as a tumor suppressor in neoplastic mast cells: Role of KIT D816V and effects of various targeted drugs. Blood 2009, 114, 5342–5351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, K.; Escribano, L.; Grattan, C.; Brockow, K.; Carter, M.C.; Alvarez-Twose, I.; Matito, A.; Broesby-Olsen, S.; Siebenhaar, F.; Lange, M.; et al. Cutaneous manifestations in patients with mastocytosis: Consensus report of the European Competence Network on Mastocytosis; The American Academy of Allergy, Asthma & Immunology; And the European Academy of Allergology and Clinical Immunology. J. Allergy Clin. Immunol. 2016, 137, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Siebenhaar, F.; Akin, C.; Bindslev-Jensen, C.; Maurer, M.; Broesby-Olsen, S. Treatment Strategies in Mastocytosis. Immunol. Allergy Clin. N. Am. 2014, 34, 433–447. [Google Scholar] [CrossRef] [PubMed]
- Jawhar, M.; Schwaab, J.; Naumann, N.; Horny, H.-P.; Sotlar, K.; Haferlach, T.; Metzgeroth, G.; Fabarius, A.; Valent, P.; Hofmann, W.-K.; et al. Response and progression on midostaurin in advanced systemic mastocytosis: KIT D816V and other molecular markers. Blood 2017, 130, 137–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valent, P.; Akin, C.; Nedoszytko, B.; Bonadonna, P.; Hartmann, K.; Niedoszytko, M.; Brockow, K.; Siebenhaar, F.; Triggiani, M.; Arock, M.; et al. Diagnosis, classification and management of mast cell activation syndromes (Mcas) in the era of personalized medicine. Int. J. Mol. Sci. 2020, 21, 9030. [Google Scholar] [CrossRef] [PubMed]
- Molderings, G.J.; Haenisch, B.; Bogdanow, M.; Fimmers, R.; Nöthen, M.M. Familial Occurrence of Systemic Mast Cell Activation Disease. PLoS ONE 2013, 8, e76241. [Google Scholar] [CrossRef] [PubMed]
- Afrin, L.B.; Ackerley, M.B.; Bluestein, L.S.; Brewer, J.H.; Brook, J.B.; Buchanan, A.D.; Cuni, J.R.; Davey, W.P.; Dempsey, T.T.; Dorff, S.R.; et al. Diagnosis of mast cell activation syndrome: A global “consensus-2”. Diagnosis 2020. [Google Scholar] [CrossRef] [PubMed]
- Butterfield, J.H. Survey of Mast Cell Mediator Levels from Patients Presenting with Symptoms of Mast Cell Activation. Int. Arch. Allergy Immunol. 2020, 181, 43–50. [Google Scholar] [CrossRef]
- Valent, P. Risk factors and management of severe life-threatening anaphylaxis in patients with clonal mast cell disorders. Clin. Exp. Allergy 2014, 44, 914–920. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The crucial role and dichotomy of mast cells (MCs) in cutaneous infections. (A) MCs critically contribute to host defenses against bacteria via direct killing, phagocytosis and the recruitment of neutrophils. (B) In viral infections, MCs control viral replication locally in the skin and promote the recruitment of natural killer cells (NK), CD4+ and CD8+ T cells, but also contribute to viral dissemination and viremia. (C) MCs control parasite infections via the release of nitric oxide species (NOS) and the formation of extracellular traps, as well as by promoting dendritic cell (DC) recruitment and activation, but can also contribute to parasite dissemination. Beneficial MC effects are displayed in green boxes; detrimental MC functions are shown in red boxes.
Figure 1.
The crucial role and dichotomy of mast cells (MCs) in cutaneous infections. (A) MCs critically contribute to host defenses against bacteria via direct killing, phagocytosis and the recruitment of neutrophils. (B) In viral infections, MCs control viral replication locally in the skin and promote the recruitment of natural killer cells (NK), CD4+ and CD8+ T cells, but also contribute to viral dissemination and viremia. (C) MCs control parasite infections via the release of nitric oxide species (NOS) and the formation of extracellular traps, as well as by promoting dendritic cell (DC) recruitment and activation, but can also contribute to parasite dissemination. Beneficial MC effects are displayed in green boxes; detrimental MC functions are shown in red boxes.
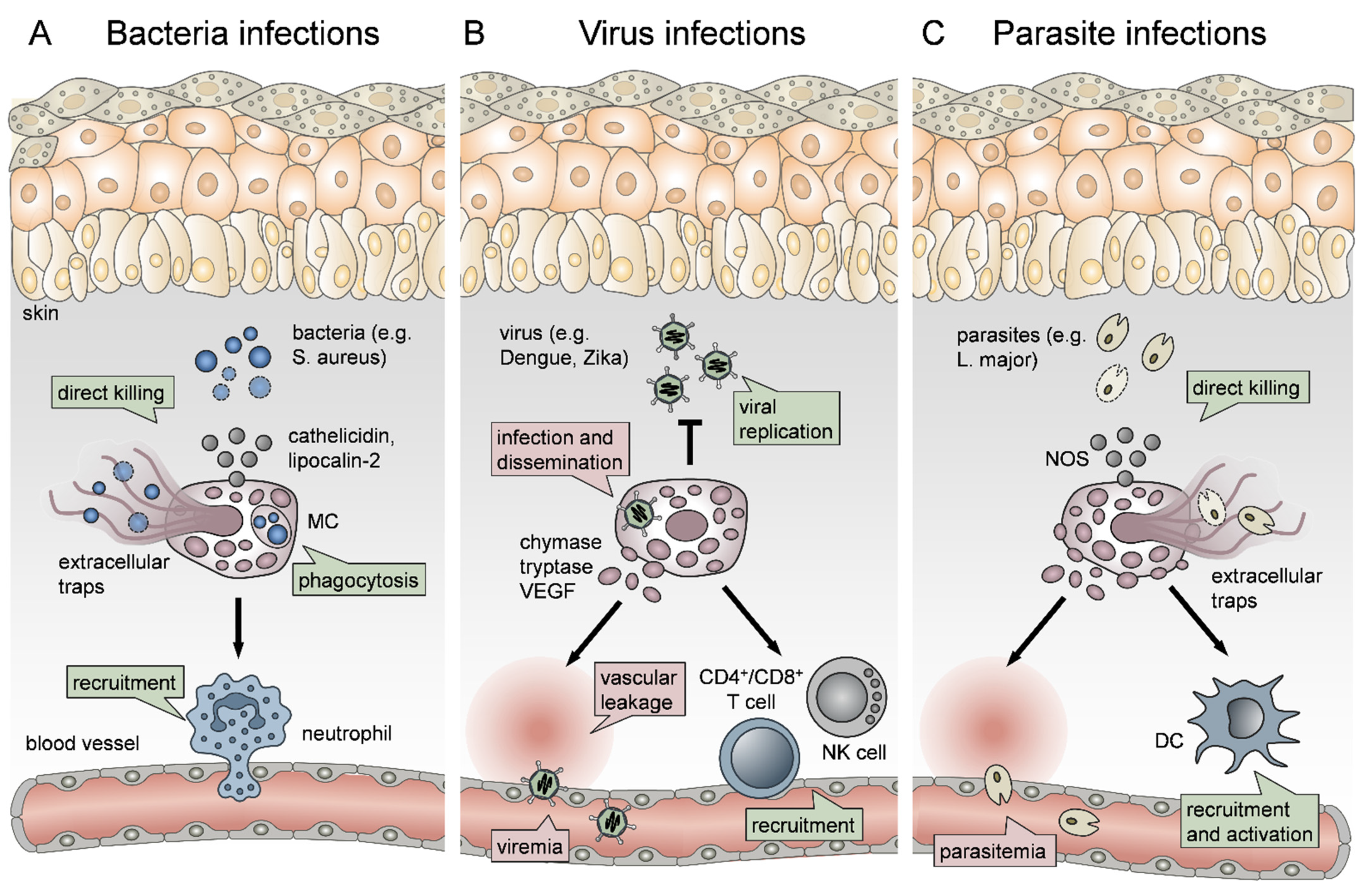
Figure 2.
The role of mast cells (MCs) in the vicious cycle of atopic dermatitis. (A) In the acute phase of atopic dermatitis (AD), allergens crosslink IgE antibodies bound to FcεRI on Langerhans cells (LC) and MCs. MC degranulation and cytokine release induce TH2 cell and eosinophil recruitment, thereby leading to a self-perpetuating cycle of TH2-driven inflammation. (B) The chronic phase of AD is characterized by a multifaceted TH response, which is perpetuated by an IgE-independent MC–nerve-skin axis. Itch and scratching promote keratinocyte (KC) activation and release of thymic stromal lymphopoietin (TSLP) and IL-33, thereby activating MCs through the TSLP receptor (TSPLR) and ST2 receptor, respectively. TSLP can also promote MRGPRX2 signaling. MC release of tryptase and histamine activates neurons, thus promoting itch and, in turn, activating MCs by neuropeptides, such as substance P (SP), through the Mas-related G-protein coupled receptor X2 (MRGPRX2). MC-derived macrophage inflammatory protein 1 (MIP-1) recruits monocytes and macrophages (Mph), which together with MCs promote T cell inflammation, finally leading to skin lichenification.
Figure 2.
The role of mast cells (MCs) in the vicious cycle of atopic dermatitis. (A) In the acute phase of atopic dermatitis (AD), allergens crosslink IgE antibodies bound to FcεRI on Langerhans cells (LC) and MCs. MC degranulation and cytokine release induce TH2 cell and eosinophil recruitment, thereby leading to a self-perpetuating cycle of TH2-driven inflammation. (B) The chronic phase of AD is characterized by a multifaceted TH response, which is perpetuated by an IgE-independent MC–nerve-skin axis. Itch and scratching promote keratinocyte (KC) activation and release of thymic stromal lymphopoietin (TSLP) and IL-33, thereby activating MCs through the TSLP receptor (TSPLR) and ST2 receptor, respectively. TSLP can also promote MRGPRX2 signaling. MC release of tryptase and histamine activates neurons, thus promoting itch and, in turn, activating MCs by neuropeptides, such as substance P (SP), through the Mas-related G-protein coupled receptor X2 (MRGPRX2). MC-derived macrophage inflammatory protein 1 (MIP-1) recruits monocytes and macrophages (Mph), which together with MCs promote T cell inflammation, finally leading to skin lichenification.
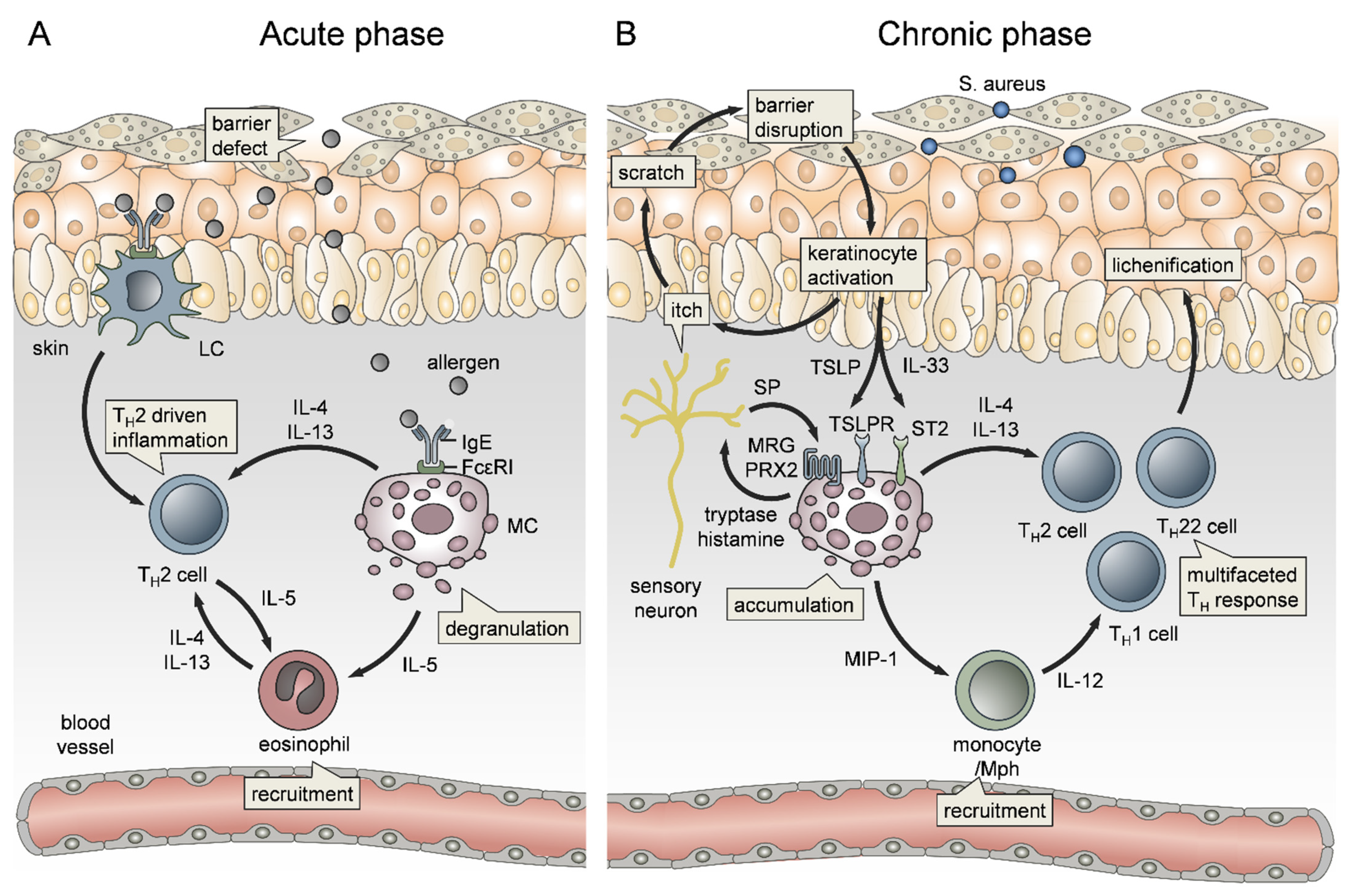
Figure 3.
Mast cell (MC) functions in allergic contact dermatitis. (A) In the sensitization phase of allergic contact dermatitis (ACD), or its mouse model of contact hypersensitivity (CHS), MCs sense keratinocyte (KC) cell stress due to their release of alarmins, such as IL-33 and ATP. Then, concomitant IL-33/ST2 and ATP/P2X7 signaling lead to the activation and degranulation of MCs. While vasodilation and vascular permeability are driven by histamine, MC-derived TNF primes and recruits neutrophils. Moreover, MC-derived TNF and MC granules promote dendritic cell (DC) maturation and migration to the skin draining lymph node (LN), thereby enhancing T cell priming. (B) Upon reexposure, MCs once more initiate vascular responses and neutrophil recruitment, but also nerve fiber elongation. Additionally, skin inflammation is amplified by infiltration of TH1 and cytotoxic T cells (TC1). Interferon γ (IFNγ) released by TC1 cells activates KCs, which in turn enhances the skin inflammation in a feedback loop by the production of the T cell recruiting chemokines Interferon γ-induced protein 10 (IP-10) and monokine-induced by gamma interferon (MIG). MC cytokine production is counter-regulated by regulatory T cells (Treg) via the release of transforming growth factor β (TGFβ).
Figure 3.
Mast cell (MC) functions in allergic contact dermatitis. (A) In the sensitization phase of allergic contact dermatitis (ACD), or its mouse model of contact hypersensitivity (CHS), MCs sense keratinocyte (KC) cell stress due to their release of alarmins, such as IL-33 and ATP. Then, concomitant IL-33/ST2 and ATP/P2X7 signaling lead to the activation and degranulation of MCs. While vasodilation and vascular permeability are driven by histamine, MC-derived TNF primes and recruits neutrophils. Moreover, MC-derived TNF and MC granules promote dendritic cell (DC) maturation and migration to the skin draining lymph node (LN), thereby enhancing T cell priming. (B) Upon reexposure, MCs once more initiate vascular responses and neutrophil recruitment, but also nerve fiber elongation. Additionally, skin inflammation is amplified by infiltration of TH1 and cytotoxic T cells (TC1). Interferon γ (IFNγ) released by TC1 cells activates KCs, which in turn enhances the skin inflammation in a feedback loop by the production of the T cell recruiting chemokines Interferon γ-induced protein 10 (IP-10) and monokine-induced by gamma interferon (MIG). MC cytokine production is counter-regulated by regulatory T cells (Treg) via the release of transforming growth factor β (TGFβ).
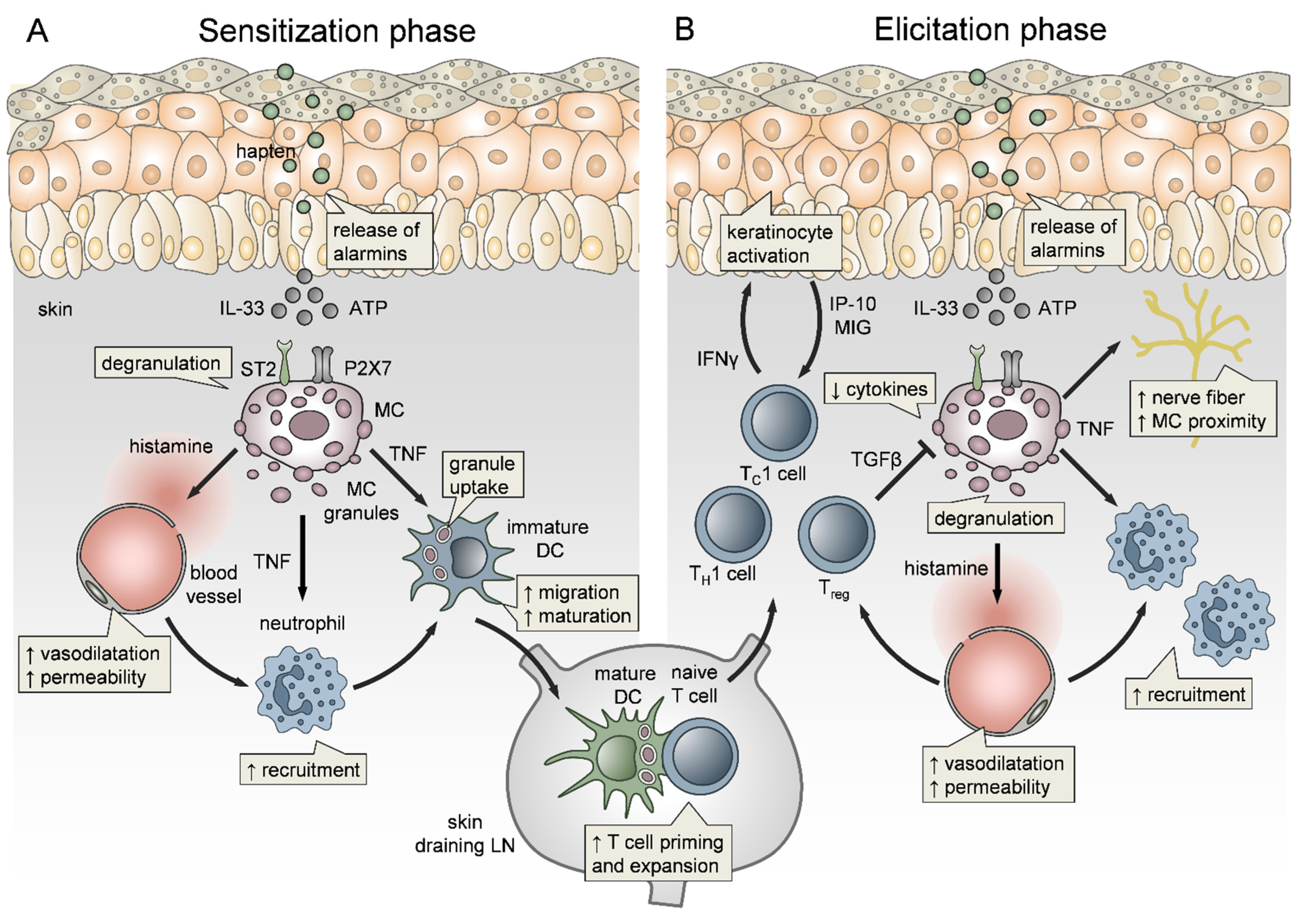
Figure 4.
The role of mast cells (MCs) in psoriasis and urticaria. (A) MCs directly contribute to psoriatic plaque formation via the release of the keratinocyte (KC) proliferation-inducing cytokines IL-17 and IL-22. Indirectly, MCs promote TH17 cell expansion via the production of phospholipase A2 (PLA2), which is taken up by dendritic cells (DC) and Langerhans cells (LC), leading to the presentation of neolipids through CD1a. Additionally, MCs promote pruritus through the release of nerve growth factor (NGF) that causes KC damage. Consequently, IL-33 production by KCs activates MCs through the ST2 receptor, thus ending in a self-perpetuating cycle. (B) In urticaria, MC activation and degranulation can be induced by numerous stimuli, including autoimmune and mechanical triggers. While histamine promotes vasodilation and vascular permeability, MC-derived TNF induces the expression of endothelial cell adhesion molecules. Moreover, tryptase and histamine are activating neurons, leading to itching and release of neuropeptides, such as substance P (SP). SP, in turn, activates MCs in an IgE-independent manner via the Mas-related G-protein coupled receptor X2 (MRGPRX2). Another IgE-independent mechanism of MC activation is through the mechanical sensing receptor ADGRE2.
Figure 4.
The role of mast cells (MCs) in psoriasis and urticaria. (A) MCs directly contribute to psoriatic plaque formation via the release of the keratinocyte (KC) proliferation-inducing cytokines IL-17 and IL-22. Indirectly, MCs promote TH17 cell expansion via the production of phospholipase A2 (PLA2), which is taken up by dendritic cells (DC) and Langerhans cells (LC), leading to the presentation of neolipids through CD1a. Additionally, MCs promote pruritus through the release of nerve growth factor (NGF) that causes KC damage. Consequently, IL-33 production by KCs activates MCs through the ST2 receptor, thus ending in a self-perpetuating cycle. (B) In urticaria, MC activation and degranulation can be induced by numerous stimuli, including autoimmune and mechanical triggers. While histamine promotes vasodilation and vascular permeability, MC-derived TNF induces the expression of endothelial cell adhesion molecules. Moreover, tryptase and histamine are activating neurons, leading to itching and release of neuropeptides, such as substance P (SP). SP, in turn, activates MCs in an IgE-independent manner via the Mas-related G-protein coupled receptor X2 (MRGPRX2). Another IgE-independent mechanism of MC activation is through the mechanical sensing receptor ADGRE2.
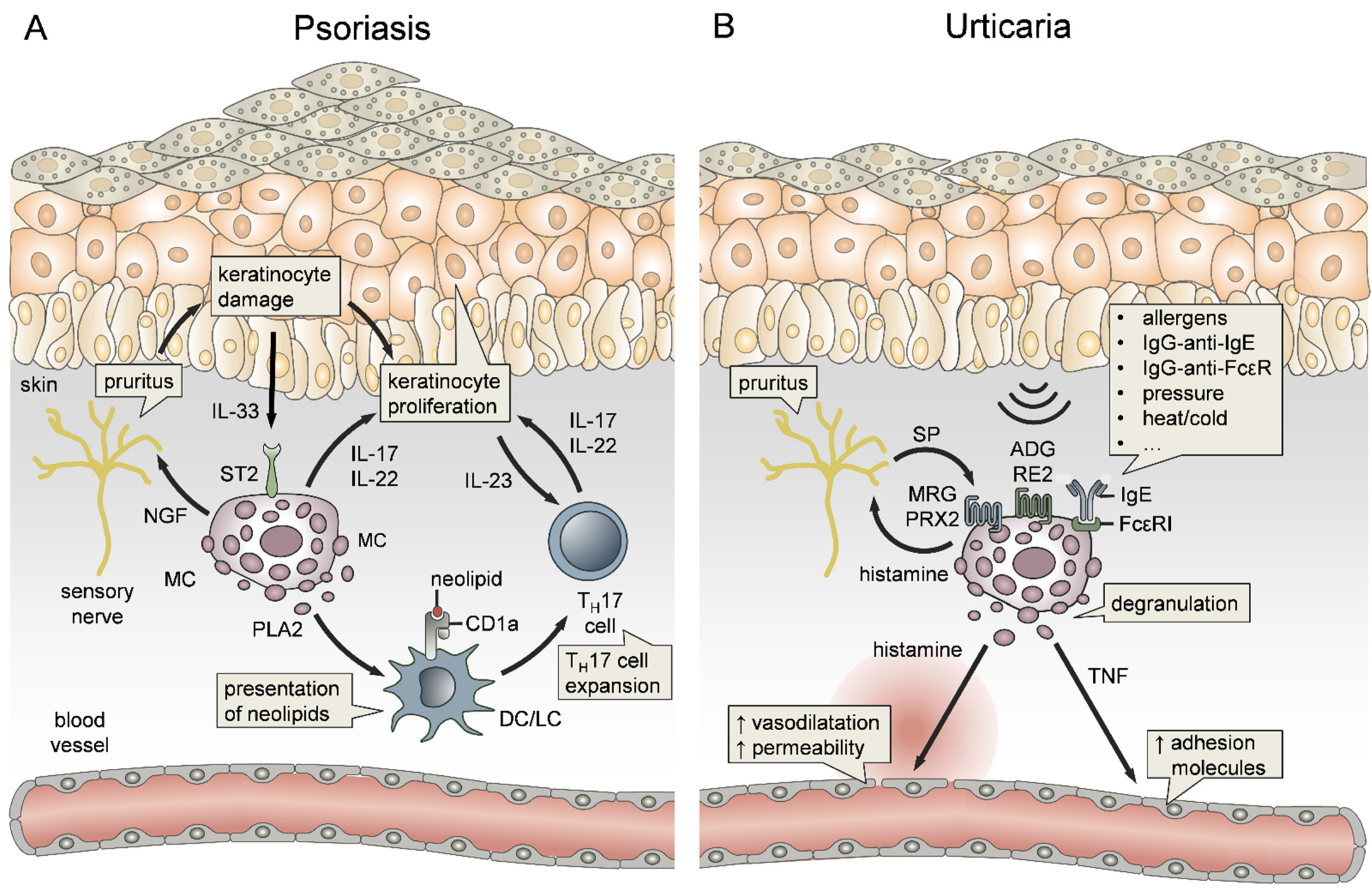
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Voss, M.; Kotrba, J.; Gaffal, E.; Katsoulis-Dimitriou, K.; Dudeck, A. Mast Cells in the Skin: Defenders of Integrity or Offenders in Inflammation? Int. J. Mol. Sci. 2021, 22, 4589. https://doi.org/10.3390/ijms22094589
AMA Style
Voss M, Kotrba J, Gaffal E, Katsoulis-Dimitriou K, Dudeck A. Mast Cells in the Skin: Defenders of Integrity or Offenders in Inflammation? International Journal of Molecular Sciences. 2021; 22(9):4589. https://doi.org/10.3390/ijms22094589
Chicago/Turabian StyleVoss, Martin, Johanna Kotrba, Evelyn Gaffal, Konstantinos Katsoulis-Dimitriou, and Anne Dudeck. 2021. "Mast Cells in the Skin: Defenders of Integrity or Offenders in Inflammation?" International Journal of Molecular Sciences 22, no. 9: 4589. https://doi.org/10.3390/ijms22094589
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.