
Sequence, Chromatin and Evolution of Satellite DNA

Abstract
1. Introduction
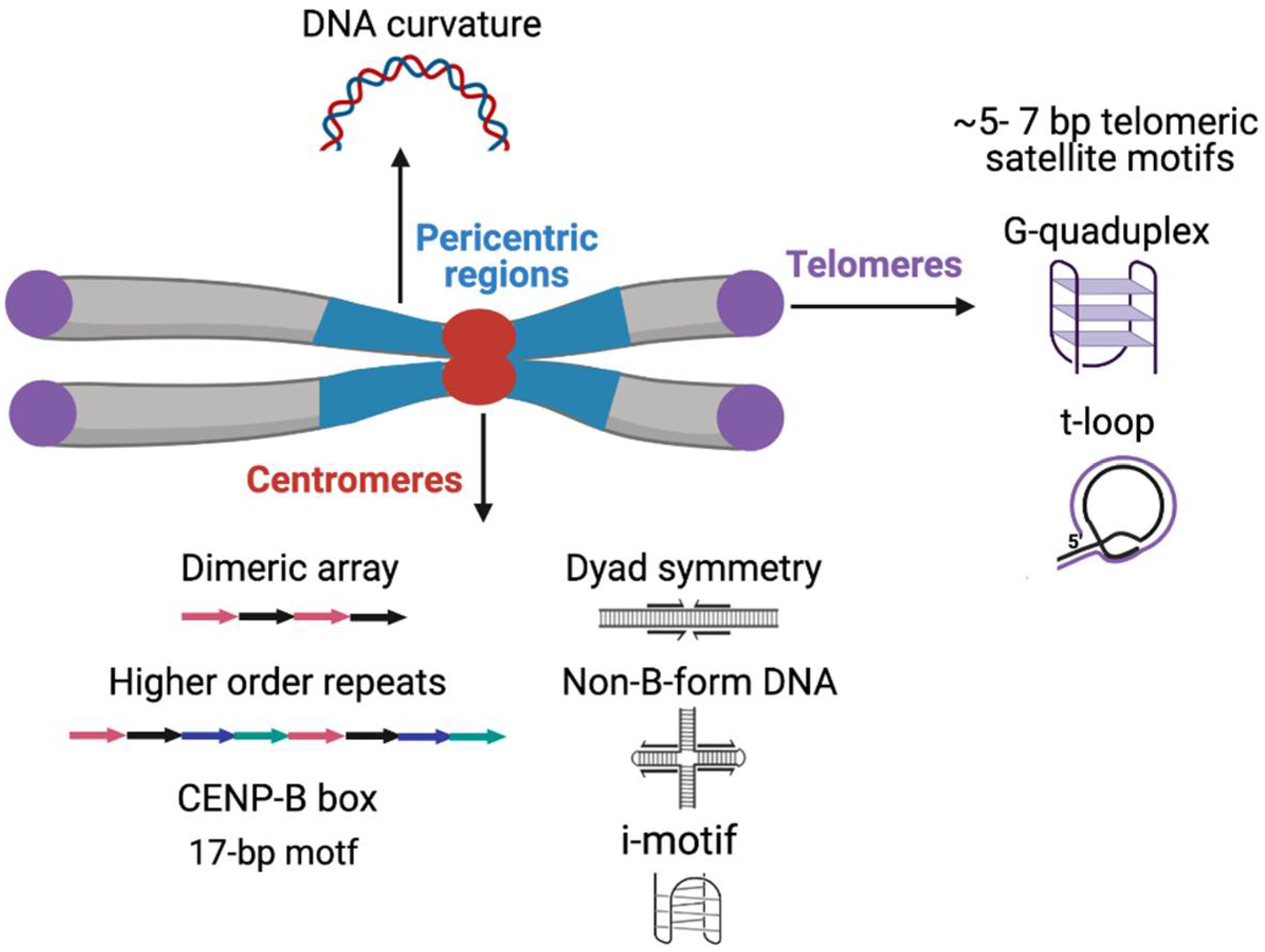
2. Satellite DNA Sequences
2.1. Functional Satellite Classes
2.1.1. Centromeric Satellites
Primate Centromeric Satellites
Mouse Centromeric Satellites
Plant Centromere Satellites
Drosophila Centromeric Satellites
2.1.2. Pericentromeric Satellites
Mammalian Pericentric Satellites
Plant Pericentric Satellites
Insect Pericentric Satellites
2.1.3. Telomeric and Subtelomeric Satellites
2.2. Roles of Satellite DNA Sequences
2.2.1. DNA Sequence Motifs
2.2.2. DNA Secondary Structures
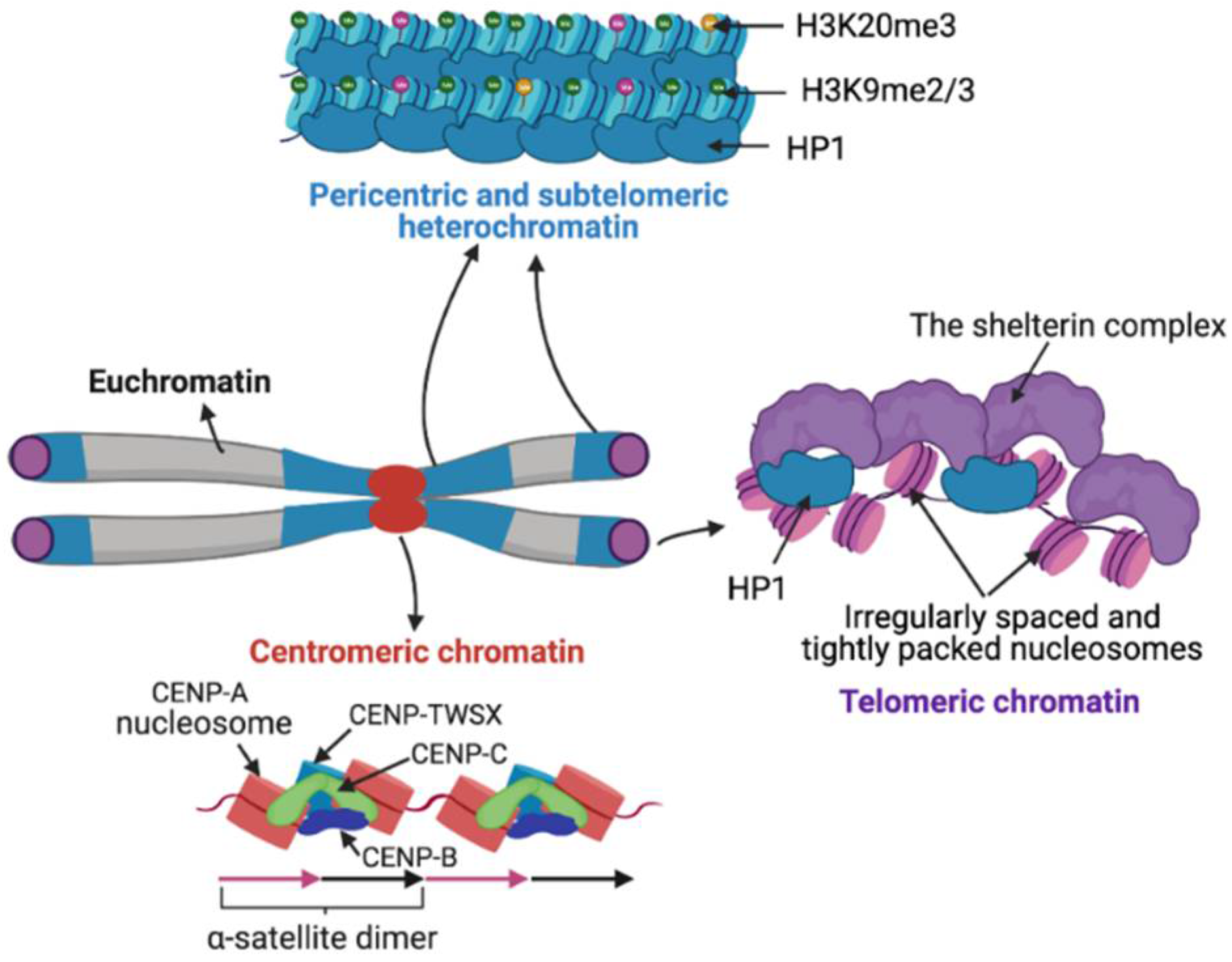
3. Satellite Chromatin
3.1. Centromeric Chromatin
3.2. Pericentric Heterochromatin
3.3. Telomeric and Subtelomeric Heterochromatin
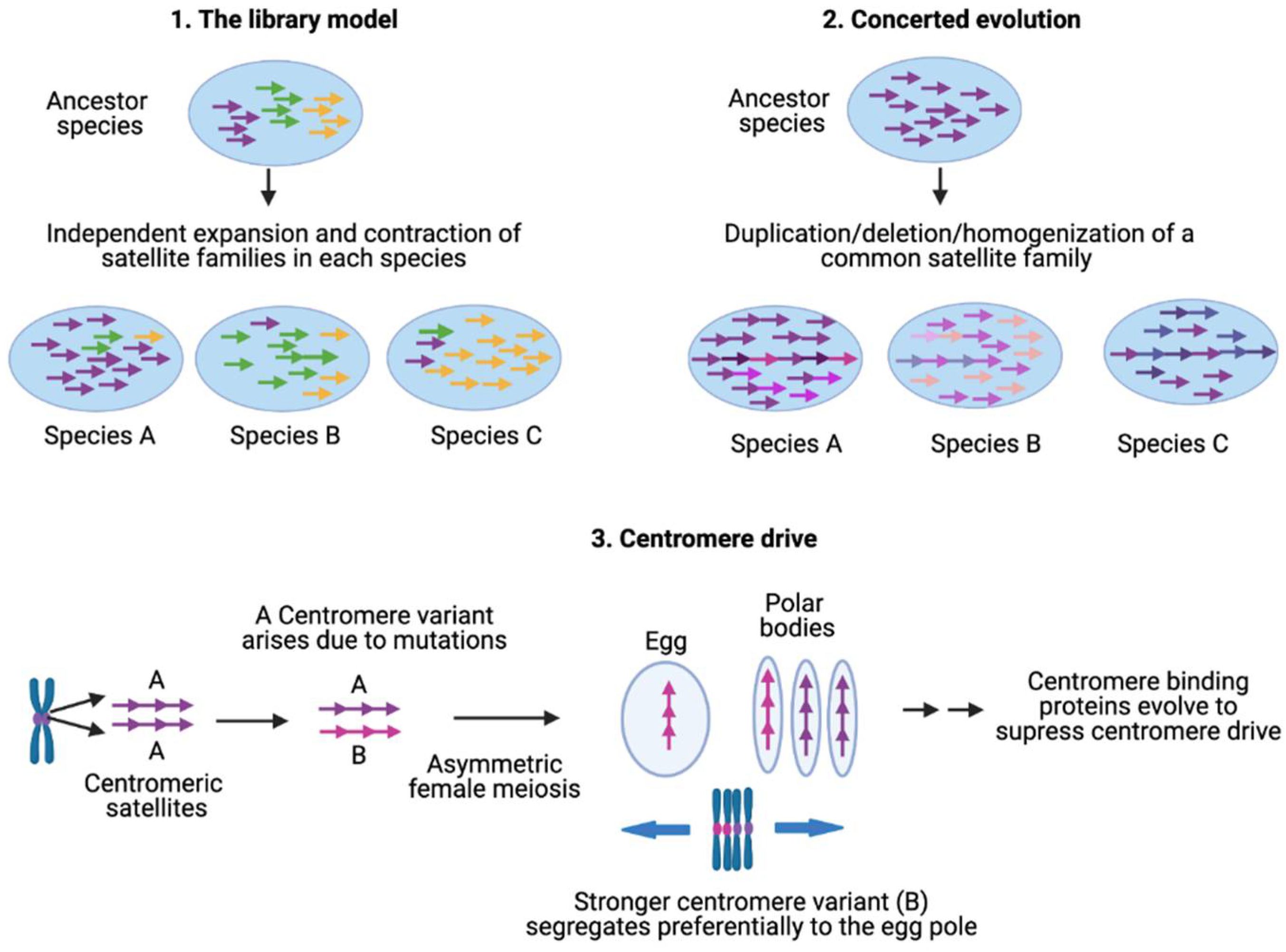
4. Satellite Evolution
4.1. Library Model for Satellite DNA Evolution
4.2. Concerted Evolution of Satellites
4.3. Centromere Drive
4.4. Repeat Evolution and Speciation
5. Conclusions and Future Perspective
Funding
Conflicts of Interest
References
- Kit, S. Equilibrium sedimentation in density gradients of DNA preparations from animal tissues. J. Mol. Biol. 1961, 3, 711–716. [Google Scholar] [CrossRef]
- Rudd, M.K.; Willard, H.F. Analysis of the centromeric regions of the human genome assembly. Trends Genet. 2004, 20, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Willard, H.F. Chromosome-specific organization of human alpha satellite DNA. Am. J. Hum. Genet. 1985, 37, 524–532. [Google Scholar]
- Schueler, M.G.; Higgins, A.W.; Rudd, M.K.; Gustashaw, K.; Willard, H.F. Genomic and genetic definition of a functional human centromere. Science 2001, 294, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Rudd, M.; Schueler, M.; Willard, H. Sequence Organization and Functional Annotation of Human Centromeres. Cold Spring Harb. Symp. Quant. Biol. 2003, 68, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Benson, G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Novák, P.; Neumann, P.; Pech, J.; Steinhaisl, J.; Macas, J. RepeatExplorer: A Galaxy-based web server for genome-wide characterization of eukaryotic repetitive elements from next-generation sequence reads. Bioinformatics 2013, 29, 792–793. [Google Scholar] [CrossRef] [PubMed]
- Novák, P.; Ávila Robledillo, L.; Koblížková, A.; Vrbová, I.; Neumann, P.; Macas, J. TAREAN: A computational tool for identification and characterization of satellite DNA from unassembled short reads. Nucleic Acids Res. 2017, 45, e111. [Google Scholar] [CrossRef]
- Subramanian, S.; Mishra, R.K.; Singh, L. Genome-wide analysis of microsatellite repeats in humans: Their abundance and density in specific genomic regions. Genome Biol. 2003, 4, R13. [Google Scholar] [CrossRef]
- International Human Genome Sequencing Consortium Initial sequencing and analysis of the human genome. Nat. Cell Biol. 2001, 409, 860–921. [CrossRef]
- Gomes, N.M.; Shay, J.W.; Wright, W.E. Telomere biology in Metazoa. FEBS Lett. 2010, 584, 3741–3751. [Google Scholar] [CrossRef] [PubMed]
- Kashi, Y.; King, D.; Soller, M. Simple sequence repeats as a source of quantitative genetic variation. Trends Genet. 1997, 13, 74–78. [Google Scholar] [CrossRef]
- Hearne, C.M.; Ghosh, S.; Todd, J.A. Microsatellites for linkage analysis of genetic traits. Trends Genet. 1992, 8, 288–294. [Google Scholar] [CrossRef]
- Zietkiewicz, E.; Rafalski, A.; Labuda, D. Genome Fingerprinting by Simple Sequence Repeat (SSR)-Anchored Polymerase Chain Reaction Amplification. Genomics 1994, 20, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Ramel, C. Mini- and microsatellites. Environ. Health Perspect. 1997, 105, 781–789. [Google Scholar] [CrossRef] [PubMed]
- Näslund, K.; Saetre, P.; Von Salomé, J.; Bergström, T.F.; Jareborg, N.; Jazin, E. Genome-wide prediction of human VNTRs. Genomics 2005, 85, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Jeffreys, A.J.; Wilson, V.; Thein, S.L. Individual-specific ‘fingerprints’ of human DNA. Nat. Cell Biol. 1985, 316, 76–79. [Google Scholar] [CrossRef]
- Azuma, C.; Kamiura, S.; Nobunaga, T.; Negoro, T.; Saji, F.; Tanizawa, O. Zygosity determination of multiple pregnancy by deoxyribonucleic acid fingerprints. Am. J. Obstet. Gynecol. 1989, 160, 734–736. [Google Scholar] [CrossRef]
- Alexandrov, A.; Kazakov, A.; Tumeneva, I.; Shepelev, V.; Yurov, Y. Alpha-satellite DNA of primates: Old and new families. Chromosoma 2001, 110, 253–266. [Google Scholar] [CrossRef]
- Wong, A.K.C.; Biddle, F.G.; Rattner, J.B. The chromosomal distribution of the major and minor satellite is not conserved in the genusMus. Chromosoma 1990, 99, 190–195. [Google Scholar] [CrossRef]
- Voullaire, L.E.; Slater, H.R.; Petrovic, V.; Choo, K.H. A functional marker centromere with no detectable alpha-satellite, satellite III, or CENP-B protein: Activation of a latent centromere? Am. J. Hum. Genet. 1993, 52, 1153–1163. [Google Scholar] [PubMed]
- Choo, K.; Earle, E.; McQuillan, C. A homologous subfamily of satellite III DNA on human chromosomes 14 and 22. Nucleic Acids Res. 1990, 18, 5641–5648. [Google Scholar] [CrossRef]
- Waye, J.S.; Willard, H.F. Human beta satellite DNA: Genomic organization and sequence definition of a class of highly repetitive tandem DNA. Proc. Natl. Acad. Sci. USA 1989, 86, 6250–6254. [Google Scholar] [CrossRef] [PubMed]
- Komissarov, A.S.; Gavrilova, E.V.; Demin, S.J.; Ishov, A.M.; Podgornaya, O.I. Tandemly repeated DNA families in the mouse genome. BMC Genom. 2011, 12, 531. [Google Scholar] [CrossRef] [PubMed]
- Dumbovic, G.; Forcales, S.-V.; Perucho, M. Emerging roles of macrosatellite repeats in genome organization and disease development. Epigenetics 2017, 12, 515–526. [Google Scholar] [CrossRef]
- Chadwick, B.P. Macrosatellite epigenetics: The two faces of DXZ4 and D4Z4. Chromosoma 2009, 118, 675–681. [Google Scholar] [CrossRef]
- Tautz, D.; Renz, M. Simple sequences are ubiquitous repetitive components of eukaryotic genomes. Nucleic Acids Res. 1984, 12, 4127–4138. [Google Scholar] [CrossRef]
- Kayser, M.; Schneider, P.M. DNA-based prediction of human externally visible characteristics in forensics: Motivations, scientific challenges, and ethical considerations. Forensic Sci. Int. Genet. 2009, 3, 154–161. [Google Scholar] [CrossRef]
- Henikoff, S.; Ahmad, K.; Malik, H.S. The Centromere Paradox: Stable Inheritance with Rapidly Evolving DNA. Science 2001, 293, 1098–1102. [Google Scholar] [CrossRef]
- Melters, D.P.; Bradnam, K.R.; Young, H.A.; Telis, N.; May, M.R.; Ruby, J.G.; Sebra, R.; Peluso, P.; Eid, J.; Rank, D.; et al. Comparative analysis of tandem repeats from hundreds of species reveals unique insights into centromere evolution. Genome Biol. 2013, 14, R10. [Google Scholar] [CrossRef]
- Fitzgerald-Hayes, M.; Clarke, L.; Carbon, J. Nucleotide sequence comparisons and functional analysis of yeast centromere DNAs. Cell 1982, 29, 235–244. [Google Scholar] [CrossRef]
- Round, E.K.; Flowers, S.K.; Richards, E.J. Arabidopsis thaliana Centromere Regions: Genetic Map Positions and Repetitive DNA Structure. Genome Res. 1997, 7, 1045–1053. [Google Scholar] [CrossRef] [PubMed]
- Kipling, D.; Ackford, H.E.; Taylor, B.A.; Cooke, H.J. Mouse minor satellite DNA genetically maps to the centromere and is physically linked to the proximal telomere. Genomics 1991, 11, 235–241. [Google Scholar] [CrossRef]
- Komissarov, A.S.; Kuznetsova, I.S.; Podgornaia, O.I. Mouse centromeric tandem repeats In Silico and In Situ. Genetika 2010, 46, 1217–1221. [Google Scholar] [CrossRef] [PubMed]
- Kuznetsova, I.S.; Prusov, A.N.; Enukashvily, N.I.; Podgornaya, O.I. New types of mouse centromeric satellite DNAs. Chromosom. Res. 2005, 13, 9–25. [Google Scholar] [CrossRef] [PubMed]
- Ananiev, E.V.; Phillips, R.L.; Rines, H.W. Chromosome-specific molecular organization of maize (Zea mays L.) centromeric regions. Proc. Natl. Acad. Sci. USA 1998, 95, 13073–13078. [Google Scholar] [CrossRef]
- Ananiev, E.V.; Phillips, R.L.; Rines, H.W. Complex structure of knobs and centromeric regions in maize chromosomes. TSitologiia Genet. 2000, 34, 11–15. [Google Scholar]
- Zhong, C.X.; Marshall, J.B.; Topp, C.; Mroczek, R.; Kato, A.; Nagaki, K.; Birchler, J.A.; Jiang, J.; Dawe, R.K. Centromeric Retroelements and Satellites Interact with Maize Kinetochore Protein CENH3. Plant Cell 2002, 14, 2825–2836. [Google Scholar] [CrossRef]
- Cheng, Z.; Dong, F.; Langdon, T.; Ouyang, S.; Buell, C.R.; Gu, M.; Blattner, F.R.; Jiang, J. Functional Rice Centromeres Are Marked by a Satellite Repeat and a Centromere-Specific Retrotransposon. Plant Cell 2002, 14, 1691–1704. [Google Scholar] [CrossRef]
- Haizel, T.; Lim, Y.; Leitch, A.R.; Moore, G. Molecular analysis of holocentric centromeres of Luzula species. Cytogenet. Genome Res. 2005, 109, 134–143. [Google Scholar] [CrossRef]
- Murata, M.; Ogura, Y.; Motoyoshi, F. Centromeric repetitive sequences in Arabidopsis thalitana. Jpn. J. Genet. 1994, 69, 361–370. [Google Scholar] [CrossRef]
- Simoens, C.; Gielen, J.; Van Montagu, M.; Inzé, D. Characterization of highly repetitive sequences ofArabidopsis thaliana. Nucleic Acids Res. 1988, 16, 6753–6766. [Google Scholar] [CrossRef]
- Grellet, F.; Delcasso, D.; Panabières, F.; Delseny, M. Organization and evolution of a higher plant alphoid-like satellite DNA sequence. J. Mol. Biol. 1986, 187, 495–507. [Google Scholar] [CrossRef]
- Chang, C.-H.; Chavan, A.; Palladino, J.; Wei, X.; Martins, N.M.C.; Santinello, B.; Chen, C.-C.; Erceg, J.; Beliveau, B.J.; Wu, C.-T.; et al. Islands of retroelements are major components of Drosophila centromeres. PLoS Biol. 2019, 17, e3000241. [Google Scholar] [CrossRef] [PubMed]
- Talbert, P.B.; Kasinathan, S.; Henikoff, S. Simple and Complex Centromeric Satellites in Drosophila Sibling Species. Genetics 2018, 208, 977–990. [Google Scholar] [CrossRef]
- Aldrup-MacDonald, M.E.; Sullivan, B.A. The Past, Present, and Future of Human Centromere Genomics. Genes 2014, 5, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Levy, S.; Sutton, G.; Ng, P.C.; Feuk, L.; Halpern, A.L.; Walenz, B.P.; Axelrod, N.; Huang, J.; Kirkness, E.F.; Denisov, G.; et al. The Diploid Genome Sequence of an Individual Human. PLoS Biol. 2007, 5, e254. [Google Scholar] [CrossRef] [PubMed]
- Alexandrov, I.A.; Medvedev, L.; Mashkova, T.; Kisselev, L.; Romanova, L.; Yurov, Y.; Aiexandrov, I. Definition of a new alpha satellite suprachromosomal family characterized by monomeric organization. Nucleic Acids Res. 1993, 21, 2209–2215. [Google Scholar] [CrossRef] [PubMed]
- Henikoff, J.G.; Thakur, J.; Kasinathan, S.; Henikoff, S. A unique chromatin complex occupies young α-satellite arrays of human centromeres. Sci. Adv. 2015, 1, e1400234. [Google Scholar] [CrossRef]
- Kipling, D.; Mitchell, A.R.; Masumoto, H.; E Wilson, H.; Nicol, L.; Cooke, H.J. CENP-B binds a novel centromeric sequence in the Asian mouse Mus caroli. Mol. Cell. Biol. 1995, 15, 4009–4020. [Google Scholar] [CrossRef]
- Kipling, D.; Warburton, P.E. Centromeres, CENP-B and Tigger too. Trends Genet. 1997, 13, 141–145. [Google Scholar] [CrossRef]
- Plohl, M.; Meštrović, N.; Mravinac, B. Satellite DNA Evolution. Genome Dyn. 2012, 7, 126–152. [Google Scholar] [CrossRef]
- Waye, J.S.; Durfy, S.J.; Pinkel, D.; Kenwrick, S.; Patterson, M.; Davies, K.E.; Willard, H.F. Chromosome-specific alpha satellite DNA from human chromosome 1: Hierarchical structure and genomic organization of a polymorphic domain spanning several hundred kilobase pairs of centromeric DNA. Genomics 1987, 1, 43–51. [Google Scholar] [CrossRef]
- Rudd, M.K.; Wray, G.A.; Willard, H.F. The evolutionary dynamics of α-satellite. Genome Res. 2005, 16, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Tyler-Smith, C.; Brown, W.R. Structure of the major block of alphoid satellite DNA on the human Y chromosome. J. Mol. Biol. 1987, 195, 457–470. [Google Scholar] [CrossRef]
- Rice, W.R. A Game of Thrones at Human Centromeres I. Multifarious structure necessitates a new molecular/evolutionary model. bioRxiv 2020. [Google Scholar] [CrossRef]
- Pike, L.M.; Carlisle, A.; Newell, C.; Hong, S.-B.; Musich, P.R. Sequence and evolution of rhesus monkey alphoid DNA. J. Mol. Evol. 1986, 23, 127–137. [Google Scholar] [CrossRef]
- Alves, G.; Seuánez, H.N.; Fanning, T. Alpha satellite DNA in neotropical primates (Platyrrhini). Chromosoma 1994, 103, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Sujiwattanarat, P.; Thapana, W.; Srikulnath, K.; Hirai, Y.; Hirai, H.; Koga, A. Higher-order repeat structure in alpha satellite DNA occurs in New World monkeys and is not confined to hominoids. Sci. Rep. 2015, 5, 10315. [Google Scholar] [CrossRef]
- Terada, S.; Hirai, Y.; Hirai, H.; Koga, A. Higher-order repeat structure in alpha satellite DNA is an attribute of hominoids rather than hominids. J. Hum. Genet. 2013, 58, 752–754. [Google Scholar] [CrossRef] [PubMed]
- Koga, A.; Hirai, Y.; Terada, S.; Jahan, I.; Baicharoen, S.; Arsaithamkul, V.; Hirai, H. Evolutionary Origin of Higher-Order Repeat Structure in Alpha-Satellite DNA of Primate Centromeres. DNA Res. 2014, 21, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Vance, G.H.; Curtis, C.A.; Heerema, N.A.; Schwartz, S.; Palmer, C.G. An apparently acentric marker chromosome originating from 9p with a functional centromere without detectable alpha and beta satellite sequences. Am. J. Med. Genet. 1997, 71, 436–442. [Google Scholar] [CrossRef]
- Depinet, T. Characterization of neo-centromeres in marker chromosomes lacking detectable alpha-satellite DNA. Hum. Mol. Genet. 1997, 6, 1195–1204. [Google Scholar] [CrossRef]
- Marshall, O.J.; Choo, K.H.A. Neocentromeres Come of Age. PLoS Genet. 2009, 5, e1000370. [Google Scholar] [CrossRef]
- Marshall, O.J.; Chueh, A.C.; Wong, L.H.; Choo, K.A. Neocentromeres: New Insights into Centromere Structure, Disease Development, and Karyotype Evolution. Am. J. Hum. Genet. 2008, 82, 261–282. [Google Scholar] [CrossRef]
- Maggert, K.A.; Karpen, G.H. The activation of a neocentromere in Drosophila requires proximity to an endogenous centromere. Genetics 2001, 158, 1615–1628. [Google Scholar]
- Thakur, J.; Sanyal, K. Efficient neocentromere formation is suppressed by gene conversion to maintain centromere function at native physical chromosomal loci in Candida albicans. Genome Res. 2013, 23, 638–652. [Google Scholar] [CrossRef]
- Williams, B.C.; Murphy, T.D.; Goldberg, M.L.; Karpen, G.H. Neocentromere activity of structurally acentric mini-chromosomes in Drosophila. Nat. Genet. 1998, 18, 30–38. [Google Scholar] [CrossRef]
- Shang, W.-H.; Hori, T.; Martins, N.M.; Toyoda, A.; Misu, S.; Monma, N.; Hiratani, I.; Maeshima, K.; Ikeo, K.; Fujiyama, A.; et al. Chromosome Engineering Allows the Efficient Isolation of Vertebrate Neocentromeres. Dev. Cell 2013, 24, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Garagna, S.; Redi, C.; Capanna, E.; Andayani, N.; Alfano, R.; Doi, P.; Viale, G. Genome distribution, chromosomal allocation, and organization of the major and minor satellite DNAs in 11 species and subspecies of the genus Mus. Cytogenet. Genome Res. 1993, 64, 247–255. [Google Scholar] [CrossRef]
- Kuznetsova, I.; Podgornaya, O.; Ferguson-Smith, M. High-resolution organization of mouse centromeric and pericentromeric DNA. Cytogenet. Genome Res. 2006, 112, 248–255. [Google Scholar] [CrossRef]
- Boursot, P.; Auffray, J.; Brittondavidian, J.; Bonhomme, F. The evolution of house mice. Annu. Rev. Ecol. Syst. 1993, 24, 119–152. [Google Scholar] [CrossRef]
- Cazaux, B.; Catalan, J.; Justy, F.; Escudé, C.; Desmarais, E.; Britton-Davidian, J. Evolution of the structure and composition of house mouse satellite DNA sequences in the subgenus Mus (Rodentia: Muridea): A cytogenomic approach. Chromosoma 2013, 122, 209–220. [Google Scholar] [CrossRef]
- Neumann, P.; Navrátilová, A.; Koblížková, A.; Kejnovský, E.; Hřibová, E.; Hobza, R.; Widmer, A.; Doležel, J.; Macas, J. Plant centromeric retrotransposons: A structural and cytogenetic perspective. Mob. DNA 2011, 2, 4. [Google Scholar] [CrossRef]
- Kordiš, D. A genomic perspective on the chromodomain-containing retrotransposons: Chromoviruses. Gene 2005, 347, 161–173. [Google Scholar] [CrossRef]
- Nasuda, S.; Hudakova, S.; Schubert, I.; Houben, A.; Endo, T.R. Stable barley chromosomes without centromeric repeats. Proc. Natl. Acad. Sci. USA 2005, 102, 9842–9847. [Google Scholar] [CrossRef] [PubMed]
- Bao, W.; Zhang, W.; Yang, Q.; Zhang, Y.; Han, B.; Gu, M.; Xue, Y.; Cheng, Z. Diversity of centromeric repeats in two closely related wild rice species, Oryza officinalis and Oryza rhizomatis. Mol. Genet. Genom. 2006, 275, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Talbert, P.B.; Zhang, W.; Wu, Y.; Yang, Z.; Henikoff, J.G.; Henikoff, S.; Jiang, J. The CentO satellite confers translational and rotational phasing on cenH3 nucleosomes in rice centromeres. Proc. Natl. Acad. Sci. USA 2013, 110, E4875–E4883. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhao, H.; Zhang, T.; Zeng, Z.; Zhang, P.; Zhu, B.; Han, Y.; Braz, G.T.; Casler, M.D.; Schmutz, J.; et al. Amplification and adaptation of centromeric repeats in polyploid switchgrass species. New Phytol. 2018, 218, 1645–1657. [Google Scholar] [CrossRef] [PubMed]
- Houben, A.; Schroeder-Reiter, E.; Nagaki, K.; Nasuda, S.; Wanner, G.; Murata, M.; Endo, T.R. CENH3 interacts with the centromeric retrotransposon cereba and GC-rich satellites and locates to centromeric substructures in barley. Chromosoma 2007, 116, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Hou, Y.; Ebina, H.; Levin, H.L.; Voytas, D.F. Chromodomains direct integration of retrotransposons to heterochromatin. Genome Res. 2008, 18, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Melo, J.R.; Nagaki, K.; Talbert, P.B.; Henikoff, S.; Dawe, R.K.; Jiang, J. Maize Centromeres: Organization and Functional Adaptation in the Genetic Background of Oat. Plant Cell 2004, 16, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Kaszás, E.; Birchler, J.A. Meiotic transmission rates correlate with physical features of rearranged centromeres in maize. Genetics 1998, 150, 1683–1692. [Google Scholar]
- Kaszás, E.; Birchler, J.A. Misdivision analysis of centromere structure in maize. EMBO J. 1996, 15, 5246–5255. [Google Scholar] [CrossRef] [PubMed]
- Dong, F.; Miller, J.T.; Jackson, S.A.; Wang, G.-L.; Ronald, P.C.; Jiang, J. Rice (Oryza sativa) centromeric regions consist of complex DNA. Proc. Natl. Acad. Sci. USA 1998, 95, 8135–8140. [Google Scholar] [CrossRef]
- Jiang, J.; Birchler, J.A.; Parrott, W.A.; Dawe, R.K. A molecular view of plant centromeres. Trends Plant Sci. 2003, 8, 570–575. [Google Scholar] [CrossRef]
- Birchler, J.A.; Gao, Z.; Han, F. Plant Centromeres. In Advanced Structural Safety Studies; Metzler, J.B., Ed.; Springer: New York, NY, USA, 2011; Volume 4, pp. 133–142. [Google Scholar]
- Lermontova, I.; Sandmann, M.; Demidov, D. Centromeres and kinetochores of Brassicaceae. Chromosom. Res. 2014, 22, 135–152. [Google Scholar] [CrossRef]
- Sun, X.; Le, H.D.; Wahlstrom, J.M.; Karpen, G.H. Sequence Analysis of a Functional Drosophila Centromere. Genome Res. 2003, 13, 182–194. [Google Scholar] [CrossRef]
- Sun, X.; Wahlstrom, J.; Karpen, G. Molecular Structure of a Functional Drosophila Centromere. Cell 1997, 91, 1007–1019. [Google Scholar] [CrossRef]
- Lohe, A.R.; Brutlag, D.L. Multiplicity of satellite DNA sequences in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 1986, 83, 696–700. [Google Scholar] [CrossRef]
- Schneider, K.L.; Xie, Z.; Wolfgruber, T.K.; Presting, G.G. Inbreeding drives maize centromere evolution. Proc. Natl. Acad. Sci. USA 2016, 113, E987–E996. [Google Scholar] [CrossRef]
- Jones, K.W. Chromosomal and Nuclear Location of Mouse Satellite DNA in Individual Cells. Nat. Cell Biol. 1970, 225, 912–915. [Google Scholar] [CrossRef] [PubMed]
- Pardue, M.L.; Gall, J.G. Chromosomal Localization of Mouse Satellite DNA. Science 1970, 168, 1356–1358. [Google Scholar] [CrossRef]
- Fransz, P.; De Jong, J.H.; Lysak, M.; Castiglione, M.R.; Schubert, I. Interphase chromosomes in Arabidopsis are organized as well defined chromocenters from which euchromatin loops emanate. Proc. Natl. Acad. Sci. USA 2002, 99, 14584–14589. [Google Scholar] [CrossRef] [PubMed]
- Jagannathan, M.; Cummings, R.; Yamashita, Y.M. A conserved function for pericentromeric satellite DNA. eLife 2018, 7, e34122. [Google Scholar] [CrossRef] [PubMed]
- Tagarro, I.; Fernández-Peralta, A.M.; González-Aguilera, J.J. Chromosomal localization of human satellites 2 and 3 by a FISH method using oligonucleotides as probes. Qual. Life Res. 1994, 93, 383–388. [Google Scholar] [CrossRef]
- Vissel, B.; Nagy, A.; Choo, K. A satellite III sequence shared by human chromosomes 13,14, and 21 that is contiguous with α satellite DNA. Cytogenet. Genome Res. 1992, 61, 81–86. [Google Scholar] [CrossRef]
- Meyne, J.; Goodwin, E.H.; Moyzis, R.K. Chromosome localization and orientation of the simple sequence repeat of human satellite I DNA. Chromosoma 1994, 103, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Moyzis, R.K.; Albright, K.L.; Bartholdi, M.F.; Cram, L.S.; Deaven, L.L.; Hildebrand, C.E.; Joste, N.E.; Longmire, J.L.; Meyne, J.; Schwarzacher-Robinson, T. Human chromosome-specific repetitive DNA sequences: Novel markers for genetic analysis. Chromosoma 1987, 95, 375–386. [Google Scholar] [CrossRef]
- Beauchamp, R.S.; Mitchell, A.R.; Buckland, R.A.; Bostock, C.J. Specific arrangements of human satellite III DNA sequences in human chromosomes. Chromosoma 1979, 71, 153–166. [Google Scholar] [CrossRef]
- Prosser, J.; Frommer, M.; Paul, C.; Vincent, P. Sequence relationships of three human satellite DNAs. J. Mol. Biol. 1986, 187, 145–155. [Google Scholar] [CrossRef]
- Tagarro, I.; Wiegant, J.; Raap, A.K. Assignment of human satellite 1 DNA as revealed by fluorescent In Situ hybridization with oligonucleotides. Qual. Life Res. 1994, 93, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Wevrick, R.; Fisher, R.B.; Ferguson-Smith, M.A.; Lin, C.C. Human centromeric DNAs. Qual. Life Res. 1997, 100, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Greig, G.M.; Willard, H.F. β satellite DNA: Characterization and localization of two subfamilies from the distal and proximal short arms of the human acrocentric chromosomes. Genomics 1992, 12, 573–580. [Google Scholar] [CrossRef]
- Schueler, M.G.; Dunn, J.M.; Bird, C.P.; Ross, M.T.; Viggiano, L.; Rocchi, M.; Willard, H.F.; Green, E.D. Progressive proximal expansion of the primate X chromosome centromere. Proc. Natl. Acad. Sci. USA 2005, 102, 10563–10568. [Google Scholar] [CrossRef]
- Welborn, J. Acquired Robertsonian translocations are not rare events in acute leukemia and lymphoma. Cancer Genet. Cytogenet. 2004, 151, 14–35. [Google Scholar] [CrossRef]
- Page, S.L.; Shin, J.-C.; Han, J.-Y.; Choo, K.H.A.; Shaffer, L.G. Breakpoint diversity illustrates distinct mechanisms for Robertsonian translocation formation. Hum. Mol. Genet. 1996, 5, 1279–1288. [Google Scholar] [CrossRef]
- Sullivan, B.A.; Jenkins, L.S.; Karson, E.M.; Leana-Cox, J.; Schwartz, S. Evidence for structural heterogeneity from molecular cytogenetic analysis of dicentric Robertsonian translocations. Am. J. Hum. Genet. 1996, 59, 167–175. [Google Scholar]
- Bandyopadhyay, R.; Heller, A.; Knox-DuBois, C.; McCaskill, C.; Berend, S.A.; Page, S.L.; Shaffer, L.G. Parental Origin and Timing of De Novo Robertsonian Translocation Formation. Am. J. Hum. Genet. 2002, 71, 1456–1462. [Google Scholar] [CrossRef]
- Capanna, E.; Gropp, A.; Winking, H.; Noack, G.; Civitelli, M.-V. Robertsonian metacentrics in the mouse. Chromosoma 1976, 58, 341–353. [Google Scholar] [CrossRef]
- Nachman, M.W.; Searle, J.B. Why is the house mouse karyotype so variable? Trends Ecol. Evol. 1995, 10, 397–402. [Google Scholar] [CrossRef]
- Han, J.Y.; Choo, K.H.; Shaffer, L.G. Molecular cytogenetic characterization of 17 rob(13q14q) Robertsonian translocations by FISH, narrowing the region containing the breakpoints. Am. J. Hum. Genet. 1994, 55, 960–967. [Google Scholar] [PubMed]
- Choo, K.H. Role of acrocentric cen-pter satellite DNA in Robertsonian translocation and chromosomal non-disjunction. Mol. Boil. Med. 1990, 7, 437–449. [Google Scholar]
- Garagna, S.; Page, J.; Fernandez-Donoso, R.; Zuccotti, M.; Searle, J.B. The Robertsonian phenomenon in the house mouse: Mutation, meiosis and speciation. Chromosoma 2014, 123, 529–544. [Google Scholar] [CrossRef]
- Guenatri, M.; Bailly, D.; Maison, C.; Almouzni, G. Mouse centric and pericentric satellite repeats form distinct functional heterochromatin. J. Cell Biol. 2004, 166, 493–505. [Google Scholar] [CrossRef]
- Thompson, H.L.; Schmidt, R.; Dean, C. Identification and distribution of seven classes of middle-repetitive DNA in the Arabidopsis thaliana genome. Nucleic Acids Res. 1996, 24, 3017–3022. [Google Scholar] [CrossRef]
- Brandes, A.; Thompson, H.; Dean, C.; Heslop-Harrison, J.S. Multiple repetitive DNA sequences in the paracentromeric regions of Arabidopsis thaliana L. Chromosom. Res. 1997, 5, 238–246. [Google Scholar] [CrossRef]
- Bauwens, S.; Van Oostveldt, P.; Engler, G.; Van Montagu, M. Distribution of the rDNA and three classes of highly repetitive DNA in the chromatin of interphase nuclei of Arabidopsis thaliana. Chromosoma 1991, 101, 41–48. [Google Scholar] [CrossRef]
- Stupar, R.M.; Song, J.; Tek, A.L.; Cheng, Z.; Dong, F.; Jiang, J. Highly condensed potato pericentromeric heterochromatin contains rDNA-related tandem repeats. Genetics 2002, 162, 1435–1444. [Google Scholar]
- Kuhn, G.C.S.; Küttler, H.; Moreira-Filho, O.; Heslop-Harrison, J.S.; Heslop-Harrison, J. The 1.688 Repetitive DNA of Drosophila: Concerted Evolution at Different Genomic Scales and Association with Genes. Mol. Biol. Evol. 2011, 29, 7–11. [Google Scholar] [CrossRef]
- Losada, A.; Villasante, A. Autosomal location of a new subtype of 1.688 satellite DNA of Drosophila melanogaster. Chromosom. Res. 1996, 4, 372–383. [Google Scholar] [CrossRef]
- Houtchens, K.; Lyttle, T.W. Responder (Rsp) Alleles in the Segregation Distorter (SD) System of Meiotic Drive in Drosophila may Represent a Complex Family of Satellite Repeat Sequences. Genetics 2003, 117, 291–302. [Google Scholar] [CrossRef]
- Plohl, M.; Mestrović, N.; Bruvo, B.; Ugarković, Đ. Similarity of Structural Features and Evolution of Satellite DNAs from Palorus subdepressus (Coleoptera) and Related Species. J. Mol. Evol. 1998, 46, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Lorite, P.; Palomeque, T.; Garner, I.; Petitpierre, E. Characterization and chromosome location of satellite DNA in the leaf beetle Chrysolina americana (Coleoptera, Chrysomelidae). Genetics 2000, 110, 143–150. [Google Scholar] [CrossRef]
- Palomeque, T.; Muñoz-López, M.; Carrillo, J.A.; Lorite, P. Characterization and evolutionary dynamics of a complex family of satellite DNA in the leaf beetle Chrysolina carnifex (Coleoptera, Chrysomelidae). Chromosom. Res. 2005, 13, 795–807. [Google Scholar] [CrossRef] [PubMed]
- Feliciello, I.; Chinali, G.; Ugarković, Đ. Structure and population dynamics of the major satellite DNA in the red flour beetle Tribolium castaneum. Genetics 2011, 139, 999–1008. [Google Scholar] [CrossRef] [PubMed]
- Feliciello, I.; Akrap, I.; Brajković, J.; Zlatar, I.; Ugarković, Đ. Satellite DNA as a Driver of Population Divergence in the Red Flour Beetle Tribolium castaneum. Genome Biol. Evol. 2015, 7, 228–239. [Google Scholar] [CrossRef]
- Shaw, D.D. Centromeres: Moving chromosomes through space and time. Trends Ecol. Evol. 1994, 9, 170–175. [Google Scholar] [CrossRef]
- Blattes, R.; Monod, C.; Susbielle, G.; Cuvier, O.; Wu, J.-H.; Hsieh, T.-S.; Laemmli, U.K.; Käs, E. Displacement of D1, HP1 and topoisomerase II from satellite heterochromatin by a specific polyamide. EMBO J. 2006, 25, 2397–2408. [Google Scholar] [CrossRef]
- Wu, M.-L. Association between a satellite DNA sequence and the responder of segregation distorter in D. melanogaster. Cell 1988, 54, 179–189. [Google Scholar] [CrossRef]
- Larracuente, A.M. The organization and evolution of the Responder satellite in species of the Drosophila melanogaster group: Dynamic evolution of a target of meiotic drive. BMC Evol. Biol. 2014, 14, 1–12. [Google Scholar] [CrossRef]
- Sandler, L.; Novitski, E. Meiotic Drive as an Evolutionary Force. Am. Nat. 1957, 91, 105–110. [Google Scholar] [CrossRef]
- Wu, C.-I.; True, J.R.; Johnson, N. Fitness reduction associated with the deletion of a satellite DNA array. Nat. Cell Biol. 1989, 341, 248–251. [Google Scholar] [CrossRef]
- Pavlek, M.; Gelfand, Y.; Plohl, M.; Meštrović, N. Genome-wide analysis of tandem repeats in Tribolium castaneum genome reveals abundant and highly dynamic tandem repeat families with satellite DNA features in euchromatic chromosomal arms. DNA Res. 2015, 22, 387–401. [Google Scholar] [CrossRef]
- Ugarkovic, D.; Podnar, M.; Plohl, M. Satellite DNA of the red flour beetle Tribolium castaneum—Comparative study of satellites from the genus Tribolium. Mol. Biol. Evol. 1996, 13, 1059–1066. [Google Scholar] [CrossRef]
- Blackburn, E.H. Telomeres and telomerase: Their mechanisms of action and the effects of altering their functions. FEBS Lett. 2004, 579, 859–862. [Google Scholar] [CrossRef]
- Bebikhov, D.V. Repeating sequences, organizing the telomeric region of chromosomes from the eukaryotic genome. Genetika 1993, 29, 373–387. [Google Scholar]
- Vítková, M.; Král, J.; Traut, W.; Zrzavý, J.; Marec, F. The evolutionary origin of insect telomeric repeats, (TTAGG) N. Chromosom. Res. 2005, 13, 145–156. [Google Scholar] [CrossRef]
- Riha, K.; Shippen, R.E. Telomere structure, function and maintenance in Arabidopsis. Chromosom. Res. 2003, 11, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Makarov, V.L.; Hirose, Y.; Langmore, J.P. Long G Tails at Both Ends of Human Chromosomes Suggest a C Strand Degradation Mechanism for Telomere Shortening. Cell 1997, 88, 657–666. [Google Scholar] [CrossRef]
- Casacuberta, E. Drosophila: Retrotransposons Making up Telomeres. Viruses 2017, 9, 192. [Google Scholar] [CrossRef] [PubMed]
- Mason, J.M.; Ransom, J.; Konev, A.Y. A Deficiency Screen for Dominant Suppressors of Telomeric Silencing in Drosophila. Genetics 2004, 168, 1353–1370. [Google Scholar] [CrossRef]
- Sheen, F.M.; Levis, R.W. Transposition of the LINE-like retrotransposon TART to Drosophila chromosome termini. Proc. Natl. Acad. Sci. USA 1994, 91, 12510–12514. [Google Scholar] [CrossRef]
- Mason, J.M.; Biessmann, H. The unusual telomeres of Drosophila. Trends Genet. 1995, 11, 58–62. [Google Scholar] [CrossRef]
- Levis, R.W.; Ganesan, R.; Houtchens, K.; Tolar, L.A.; Sheen, F.-M. Transposons in place of telomeric repeats at a Drosophila telomere. Cell 1993, 75, 1083–1093. [Google Scholar] [CrossRef]
- Roth, C.W.; Kobeski, F.; Walter, M.F.; Biessmann, H. Chromosome end elongation by recombination in the mosquito Anopheles gambiae. Mol. Cell. Biol. 1997, 17, 5176–5183. [Google Scholar] [CrossRef] [PubMed]
- Biessmann, H.; Kobeski, F.; Walter, M.F.; Kasravi, A.; Roth, C.W. DNA organization and length polymorphism at the 2L telomeric region of Anopheles gambiae. Insect Mol. Biol. 1998, 7, 83–93. [Google Scholar] [CrossRef]
- Compton, A.; Liang, J.; Chen, C.; Lukyanchikova, V.; Qi, Y.; Potters, M.; Settlage, R.; Miller, D.; Deschamps, S.; Mao, C.; et al. The Beginning of the End: A Chromosomal Assembly of the New World Malaria Mosquito Ends with a Novel Telomere. Genes Genomes Genet. 2020, 10, 3811–3819. [Google Scholar] [CrossRef]
- Madalena, C.R.G.; Amabis, J.M.; Gorab, E. Unusually short tandem repeats appear to reach chromosome ends of Rhynchosciara americana (Diptera: Sciaridae). Chromosoma 2010, 119, 613–623. [Google Scholar] [CrossRef]
- Flint, J.; Thomas, K.; Micklem, G.; Raynham, H.; Clark, K.; Doggett, N.A.; Andrew, A.; Higgs, D.R. The relationship between chromosome structure and function at a human telomeric region. Nat. Genet. 1997, 15, 252–257. [Google Scholar] [CrossRef]
- Flint, J.; Bates, G.P.; Clark, K.; Dorman, A.; Willingham, D.; Roe, B.A.; Micklem, G.; Higgs, D.R.; Louis, E.J. Sequence comparison of human and yeast telomeres identifies structurally distinct subtelomeric domains. Hum. Mol. Genet. 1997, 6, 1305–1313. [Google Scholar] [CrossRef]
- Chute, I.; Le, Y.; Ashley, T.; Dobson, M.J. The Telomere-Associated DNA from Human Chromosome 20p Contains a Pseudotelomere Structure and Shares Sequences with the Subtelomeric Regions of 4q and 18p. Genomics 1997, 46, 51–60. [Google Scholar] [CrossRef]
- Riethman, H.; Ambrosini, A.; Paul, S. Human subtelomere structure and variation. Chromosom. Res. 2005, 13, 505–515. [Google Scholar] [CrossRef]
- Mefford, H.C.; Trask, B.J. The complex structure and dynamic evolution of human subtelomeres. Nat. Rev. Genet. 2002, 3, 91–102. [Google Scholar] [CrossRef]
- Ambrosini, A.; Paul, S.; Hu, S.; Riethman, H. Human subtelomeric duplicon structure and organization. Genome Biol. 2007, 8, R151. [Google Scholar] [CrossRef]
- Riethman, H. Human subtelomeric copy number variations. Cytogenet. Genome Res. 2008, 123, 244–252. [Google Scholar] [CrossRef]
- Ohzeki, J.-I.; Nakano, M.; Okada, T.; Masumoto, H. CENP-B box is required for De Novo centromere chromatin assembly on human alphoid DNA. J. Cell Biol. 2002, 159, 765–775. [Google Scholar] [CrossRef]
- Fachinetti, D.; Han, J.S.; McMahon, M.A.; Ly, P.; Abdullah, A.; Wong, A.J.; Cleveland, D.W. DNA Sequence-Specific Binding of CENP-B Enhances the Fidelity of Human Centromere Function. Dev. Cell 2015, 33, 314–327. [Google Scholar] [CrossRef]
- Masumoto, H.; Masukata, H.; Muro, Y.; Nozaki, N.; Okazaki, T. A human centromere antigen (CENP-B) interacts with a short specific sequence in alphoid DNA, a human centromeric satellite. J. Cell Biol. 1989, 109, 1963–1973. [Google Scholar] [CrossRef]
- Mateo, L.; González, J. Pogo-like Transposases Have Been Repeatedly Domesticated into CENP-B-Related Proteins. Genome Biol. Evol. 2014, 6, 2008–2016. [Google Scholar] [CrossRef]
- Casola, C.; Hucks, D.; Feschotte, C. Convergent Domestication of pogo-like Transposases into Centromere-Binding Proteins in Fission Yeast and Mammals. Mol. Biol. Evol. 2007, 25, 29–41. [Google Scholar] [CrossRef]
- Rosandić, M.; Paar, V.; Basar, I.; Glunčić, M.; Pavin, N.; Pilaš, I. CENP-B box and pJα sequence distribution in human alpha satellite higher-order repeats (HOR). Chromosom. Res. 2006, 14, 735–753. [Google Scholar] [CrossRef]
- Paar, V.; Basar, I.; Rosandić, M.; Gluncić, M. Consensus Higher Order Repeats and Frequency of String Distributions in Human Genome. Curr. Genom. 2007, 8, 93–111. [Google Scholar] [CrossRef]
- Kasinathan, S.; Henikoff, S. Non-B-Form DNA Is Enriched at Centromeres. Mol. Biol. Evol. 2018, 35, 949–962. [Google Scholar] [CrossRef]
- Garavís, M.; Escaja, N.; Gabelica, V.; Villasante, A.; González, C. Centromeric Alpha-Satellite DNA Adopts Dimeric i-Motif Structures Capped by AT Hoogsteen Base Pairs. Chem. A Eur. J. 2015, 21, 9816–9824. [Google Scholar] [CrossRef]
- Garavís, M.; Méndez-Lago, M.; Gabelica, V.; Whitehead, S.L.; González, C.; Villasante, A. The structure of an endogenous Drosophila centromere reveals the prevalence of tandemly repeated sequences able to form i-motifs. Sci. Rep. 2015, 5, 13307. [Google Scholar] [CrossRef]
- Barceló, F.; Gutierrez, F.; Barjau, I.; Portugal, J. A Theoretical Perusal of the Satellite DNA Curvature in Tenebrionid Beetles. J. Biomol. Struct. Dyn. 1998, 16, 41–50. [Google Scholar] [CrossRef]
- Rojas-Rousse, D.; Bigot, Y.; Periquet, G. DNA insertions as a component of the evolution of unique satellite DNA families in two genera of parasitoid wasps: Diadromus and Eupelmus (Hymenoptera). Mol. Biol. Evol. 1993, 10, 383–396. [Google Scholar] [CrossRef]
- Landais, I.; Chavigny, P.; Castagnone, C.; Pizzol, J.; Abad, P.; Vanlerberghe-Masutti, F. Characterization of a highly conserved satellite DNA from the parasitoid wasp Trichogramma brassicae. Gene 2000, 255, 65–73. [Google Scholar] [CrossRef]
- Mravinac, B.; Ugarković, Ð.; Franjević, D.; Plohl, M. Long Inversely Oriented Subunits Form a Complex Monomer of Tribolium brevicornis Satellite DNA. J. Mol. Evol. 2005, 60, 513–525. [Google Scholar] [CrossRef]
- Rosén, M.; Castillejo-López, C.; Edström, J.-E. Telomere terminating with centromere-specific repeats is closely associated with a transposon derived gene in Chironomus pallidivittatus. Chromosoma 2002, 110, 532–541. [Google Scholar] [CrossRef] [PubMed]
- Pons, J.; Bruvo, B.; Petitpierre, E.; Plohl, M.; Ugarković, D.; Juan, C. Complex structural features of satellite DNA sequences in the genus Pimelia (Coleoptera: Tenebrionidae): Random differential amplification from a common ‘satellite DNA library’. Heredity 2004, 92, 418–427. [Google Scholar] [CrossRef] [PubMed]
- Barcelo, F.; Pons, J.; Petitpierre, E.; Barjau, I.; Portugal, J. Polymorphic Curvature of Satellite DNA in Three Subspecies of the Beetle Pimelia sparsa. J. Biol. Inorg. Chem. 1997, 244, 318–324. [Google Scholar] [CrossRef]
- Lobov, I.B.; Tsutsui, K.; Mitchell, A.R.; Podgornaya, O.I. Specificity of SAF-A and lamin B binding in vitro correlates with the satellite DNA bending state. J. Cell. Biochem. 2001, 83, 218–229. [Google Scholar] [CrossRef]
- de Lange, T. Shelterin: The protein complex that shapes and safeguards human telomeres. Genes Dev. 2005, 19, 2100–2110. [Google Scholar] [CrossRef] [PubMed]
- Denchi, E.L. Give me a break: How telomeres suppress the DNA damage response. DNA Repair 2009, 8, 1118–1126. [Google Scholar] [CrossRef]
- Griffith, J.D.; Comeau, L.; Rosenfield, S.; Stansel, R.M.; Bianchi, A.; Moss, H.; de Lange, T. Mammalian Telomeres End in a Large Duplex Loop. Cell 1999, 97, 503–514. [Google Scholar] [CrossRef]
- Ruis, P.; Boulton, S.J. The end protection problem—An unexpected twist in the tail. Genes Dev. 2021, 35, 1–21. [Google Scholar] [CrossRef]
- Muñoz-Jordán, J.L.; Cross, G.A.; De Lange, T.; Griffith, J.D. t-loops at trypanosome telomeres. EMBO J. 2001, 20, 579–588. [Google Scholar] [CrossRef]
- Tomáška, Ľ.; Cesare, A.J.; Alturki, T.M.; Griffith, J.D. Twenty years of t-loops: A case study for the importance of collaboration in molecular biology. DNA Repair 2020, 94, 102901. [Google Scholar] [CrossRef]
- Paeschke, K.; Simonsson, T.; Postberg, J.; Rhodes, D.; Lipps, H.J. Telomere end-binding proteins control the formation of G-quadruplex DNA structures In Vivo. Nat. Struct. Mol. Biol. 2005, 12, 847–854. [Google Scholar] [CrossRef]
- Schaffitzel, C.; Berger, I.; Postberg, J.; Hanes, J.; Lipps, H.J.; Plückthun, A. In Vitro generated antibodies specific for telomeric guanine-quadruplex DNA react with Stylonychia lemnae macronuclei. Proc. Natl. Acad. Sci. USA 2001, 98, 8572–8577. [Google Scholar] [CrossRef] [PubMed]
- Pagano, B.; Amato, J.; Iaccarino, N.; Cingolani, C.; Zizza, P.; Biroccio, A.; Novellino, E.; Randazzo, A. Looking for Efficient G-Quadruplex Ligands: Evidence for Selective Stabilizing Properties and Telomere Damage by Drug-Like Molecules. ChemMedChem 2015, 10, 640–649. [Google Scholar] [CrossRef]
- Granotier, C. Preferential binding of a G-quadruplex ligand to human chromosome ends. Nucleic Acids Res. 2005, 33, 4182–4190. [Google Scholar] [CrossRef] [PubMed]
- Tahara, H.; Shin-Ya, K.; Seimiya, H.; Yamada, H.; Tsuruo, T.; Ide, T. G-Quadruplex stabilization by telomestatin induces TRF2 protein dissociation from telomeres and anaphase bridge formation accompanied by loss of the 3′ telomeric overhang in cancer cells. Oncogene 2006, 25, 1955–1966. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, R.; Müller, S.; Yeoman, J.A.; Trentesaux, C.; Riou, J.-F.; Balasubramanian, S. A Novel Small Molecule That Alters Shelterin Integrity and Triggers a DNA-Damage Response at Telomeres. J. Am. Chem. Soc. 2008, 130, 15758–15759. [Google Scholar] [CrossRef]
- Canudas, S.; Smith, S. Differential regulation of telomere and centromere cohesion by the Scc3 homologues SA1 and SA2, respectively, in human cells. J. Cell Biol. 2009, 187, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Saint-André, C.D.L.R. Alternative ends: Telomeres and meiosis. Biochimie 2008, 90, 181–189. [Google Scholar] [CrossRef]
- Sen, D.; Gilbert, W. Formation of parallel four-stranded complexes by guanine-rich motifs in DNA and its implications for meiosis. Nat. Cell Biol. 1988, 334, 364–366. [Google Scholar] [CrossRef]
- Csink, A.K.; Henikoff, S. Something from nothing: The evolution and utility of satellite repeats. Trends Genet. 1998, 14, 200–204. [Google Scholar] [CrossRef]
- Blower, M.D.; Sullivan, B.A.; Karpen, G.H. Conserved Organization of Centromeric Chromatin in Flies and Humans. Dev. Cell 2002, 2, 319–330. [Google Scholar] [CrossRef]
- Sullivan, B.A.; Karpen, G.H. Centromeric chromatin exhibits a histone modification pattern that is distinct from both euchromatin and heterochromatin. Nat. Struct. Mol. Biol. 2004, 11, 1076–1083. [Google Scholar] [CrossRef]
- Palmer, D.K.; Day, M.H.W.; Wener, M.H.; Andrews, B.S.; Margolis, R.L. A 17-kD centromere protein (CENP-A) copurifies with nucleosome core particles and with histones. J. Cell Biol. 1987, 104, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Westhorpe, F.G.; Straight, A.F. The Centromere: Epigenetic Control of Chromosome Segregation during Mitosis. Cold Spring Harb. Perspect. Biol. 2014, 7, a015818. [Google Scholar] [CrossRef] [PubMed]
- Henikoff, S.; Thakur, J.; Kasinathan, S.; Talbert, P.B. Remarkable Evolutionary Plasticity of Centromeric Chromatin. Cold Spring Harb. Symp. Quant. Biol. 2017, 82, 71–82. [Google Scholar] [CrossRef]
- Westermann, S.; Cheeseman, I.M.; Anderson, S.; Yates, J.R.; Drubin, D.G.; Barnes, G. Architecture of the budding yeast kinetochore reveals a conserved molecular core. J. Cell Biol. 2003, 163, 215–222. [Google Scholar] [CrossRef]
- Welburn, J.P.; Cheeseman, I.M. Toward a Molecular Structure of the Eukaryotic Kinetochore. Dev. Cell 2008, 15, 645–655. [Google Scholar] [CrossRef]
- Hayden, K.E.; Strome, E.D.; Merrett, S.L.; Lee, H.-R.; Rudd, M.K.; Willard, H.F. Sequences Associated with Centromere Competency in the Human Genome. Mol. Cell. Biol. 2012, 33, 763–772. [Google Scholar] [CrossRef]
- Thakur, J.; Henikoff, S. Unexpected conformational variations of the human centromeric chromatin complex. Genes Dev. 2018, 32, 20–25. [Google Scholar] [CrossRef]
- Hasson, D.; Panchenko, T.; Salimian, K.J.; Salman, M.U.; Sekulic, N.; Alonso, A.; Warburton, P.E.; Black, B.E. The octamer is the major form of CENP-A nucleosomes at human centromeres. Nat. Struct. Mol. Biol. 2013, 20, 687–695. [Google Scholar] [CrossRef]
- Bodor, D.L.; Mata, J.F.; Sergeev, M.; David, A.F.; Salimian, K.J.; Panchenko, T.; Cleveland, D.W.; Black, B.E.; Shah, J.V.; Jansen, L.E. The quantitative architecture of centromeric chromatin. eLife 2014, 3, e02137. [Google Scholar] [CrossRef] [PubMed]
- Thakur, J.; Henikoff, S. CENPT bridges adjacent CENPA nucleosomes on young human α-satellite dimers. Genome Res. 2016, 26, 1178–1187. [Google Scholar] [CrossRef]
- Kato, H.; Jiang, J.; Zhou, B.-R.; Rozendaal, M.; Feng, H.; Ghirlando, R.; Xiao, T.S.; Straight, A.F.; Bai, Y. A Conserved Mechanism for Centromeric Nucleosome Recognition by Centromere Protein CENP-C. Science 2013, 340, 1110–1113. [Google Scholar] [CrossRef]
- Ando, S.; Yang, H.; Nozaki, N.; Okazaki, T.; Yoda, K. CENP-A, -B, and -C Chromatin Complex That Contains the I-Type α-Satellite Array Constitutes the Prekinetochore in HeLa Cells. Mol. Cell. Biol. 2002, 22, 2229–2241. [Google Scholar] [CrossRef] [PubMed]
- Bobkov, G.O.; Gilbert, N.; Heun, P. Centromere transcription allows CENP-A to transit from chromatin association to stable incorporation. J. Cell Biol. 2018, 217, 1957–1972. [Google Scholar] [CrossRef] [PubMed]
- Chan, F.L.; Marshall, O.J.; Saffery, R.; Kim, B.W.; Earle, E.; Choo, K.H.A.; Wong, L.H. Active transcription and essential role of RNA polymerase II at the centromere during mitosis. Proc. Natl. Acad. Sci. USA 2012, 109, 1979–1984. [Google Scholar] [CrossRef] [PubMed]
- Wong, L.H.; Brettingham-Moore, K.H.; Chan, L.; Quach, J.M.; Anderson, M.A.; Northrop, E.L.; Hannan, R.; Saffery, R.; Shaw, M.L.; Williams, E.; et al. Centromere RNA is a key component for the assembly of nucleoproteins at the nucleolus and centromere. Genome Res. 2007, 17, 1146–1160. [Google Scholar] [CrossRef]
- Du, Y.; Topp, C.N.; Dawe, R.K. DNA Binding of Centromere Protein C (CENPC) Is Stabilized by Single-Stranded RNA. PLoS Genet. 2010, 6, e1000835. [Google Scholar] [CrossRef]
- McNulty, S.M.; Sullivan, L.L.; Sullivan, B.A. Human Centromeres Produce Chromosome-Specific and Array-Specific Alpha Satellite Transcripts that Are Complexed with CENP-A and CENP-C. Dev. Cell 2017, 42, 226–240. [Google Scholar] [CrossRef]
- Yamagata, K.; Yamazaki, T.; Miki, H.; Ogonuki, N.; Inoue, K.; Ogura, A.; Baba, T. Centromeric DNA hypomethylation as an epigenetic signature discriminates between germ and somatic cell lineages. Dev. Biol. 2007, 312, 419–426. [Google Scholar] [CrossRef]
- Luo, S.; Preuss, D. Strand-biased DNA methylation associated with centromeric regions in Arabidopsis. Proc. Natl. Acad. Sci. USA 2003, 100, 11133–11138. [Google Scholar] [CrossRef]
- Li, Y.; Miyanari, Y.; Shirane, K.; Nitta, H.; Kubota, T.; Ohashi, H.; Okamoto, A.; Sasaki, H. Sequence-specific microscopic visualization of DNA methylation status at satellite repeats in individual cell nuclei and chromosomes. Nucleic Acids Res. 2013, 41, e186. [Google Scholar] [CrossRef]
- Koo, D.-H.; Han, F.; Birchler, J.A.; Jiang, J. Distinct DNA methylation patterns associated with active and inactive centromeres of the maize B chromosome. Genome Res. 2011, 21, 908–914. [Google Scholar] [CrossRef]
- Yan, H.; Kikuchi, S.; Neumann, P.; Zhang, W.; Wu, Y.; Chen, F.; Jiang, J. Genome-wide mapping of cytosine methylation revealed dynamic DNA methylation patterns associated with genes and centromeres in rice. Plant J. 2010, 63, 353–365. [Google Scholar] [CrossRef]
- Ichikawa, K.; Tomioka, S.; Suzuki, Y.; Nakamura, R.; Doi, K.; Yoshimura, J.; Kumagai, M.; Inoue, Y.; Uchida, Y.; Irie, N.; et al. Centromere evolution and CpG methylation during vertebrate speciation. Nat. Commun. 2017, 8, 1833. [Google Scholar] [CrossRef]
- Mitchell, A.R.; Jeppesen, P.; Nicol, L.; Morrison, H.; Kipling, D. Epigenetic control of mammalian centromere protein binding: Does DNA methylation have a role? J. Cell Sci. 1996, 109, 2199–2206. [Google Scholar] [PubMed]
- Gopalakrishnan, S.; Sullivan, B.A.; Trazzi, S.; Della Valle, G.; Robertson, K.D. DNMT3B interacts with constitutive centromere protein CENP-C to modulate DNA methylation and the histone code at centromeric regions. Hum. Mol. Genet. 2009, 18, 3178–3193. [Google Scholar] [CrossRef]
- Tartof, K.D.; Bishop, C.; Jones, M.; Hobbs, C.A.; Locke, J. Towards an understanding of position effect variegation. Dev. Genet. 1989, 10, 162–176. [Google Scholar] [CrossRef] [PubMed]
- Girton, J.R.; Johansen, K.M. Chapter 1 Chromatin Structure and the Regulation of Gene Expression: The Lessons of PEV in Drosophila. Adv. Genet. 2008, 61, 1–43. [Google Scholar] [CrossRef] [PubMed]
- Sass, G.L.; Henikoff, S. Comparative analysis of position-effect variegation mutations in Drosophila melanogaster delineates the targets of modifiers. Genetics 1998, 148, 733–741. [Google Scholar]
- Hall, L.L.; Byron, M.; Carone, D.M.; Whitfield, T.W.; Pouliot, G.P.; Fischer, A.; Jones, P.; Lawrence, J.B. Demethylated HSATII DNA and HSATII RNA Foci Sequester PRC1 and MeCP2 into Cancer-Specific Nuclear Bodies. Cell Rep. 2017, 18, 2943–2956. [Google Scholar] [CrossRef] [PubMed]
- Landers, C.C.; Rabeler, C.A.; Ferrari, E.K.; D’Alessandro, L.R.; Kang, D.D.; Malisa, J.; Bashir, S.M.; Carone, D.M. Ectopic expression of pericentric HSATII RNA results in nuclear RNA accumulation, MeCP2 recruitment, and cell division defects. Chromosoma 2021, 130, 75–90. [Google Scholar] [CrossRef] [PubMed]
- Bernard, P.; Maure, J.-F.; Partridge, J.F.; Genier, S.; Javerzat, J.-P.; Allshire, R.C. Requirement of Heterochromatin for Cohesion at Centromeres. Science 2001, 294, 2539–2542. [Google Scholar] [CrossRef] [PubMed]
- Schotta, G.; Sengupta, R.; Kubicek, S.; Malin, S.; Kauer, M.; Callén, E.; Celeste, A.; Pagani, M.; Opravil, S.; De La Rosa-Velazquez, I.A.; et al. A chromatin-wide transition to H4K20 monomethylation impairs genome integrity and programmed DNA rearrangements in the mouse. Genes Dev. 2008, 22, 2048–2061. [Google Scholar] [CrossRef] [PubMed]
- Schotta, G.; Lachner, M.; Sarma, K.; Ebert, A.; Sengupta, R.; Reuter, G.; Reinberg, D.; Jenuwein, T. A silencing pathway to induce H3-K9 and H4-K20 trimethylation at constitutive heterochromatin. Genes Dev. 2004, 18, 1251–1262. [Google Scholar] [CrossRef] [PubMed]
- Almouzni, G.; Probst, A.V. Heterochromatin maintenance and establishment: Lessons from the mouse pericentromere. Nucleus 2011, 2, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Hahn, M.; Dambacher, S.; Dulev, S.; Kuznetsova, A.Y.; Eck, S.; Wörz, S.; Sadic, D.; Schulte, M.; Mallm, J.-P.; Maiser, A.; et al. Suv4-20h2 mediates chromatin compaction and is important for cohesin recruitment to heterochromatin. Genes Dev. 2013, 27, 859–872. [Google Scholar] [CrossRef]
- Jørgensen, S.; Schotta, G.; Sørensen, C.S. Histone H4 Lysine 20 methylation: Key player in epigenetic regulation of genomic integrity. Nucleic Acids Res. 2013, 41, 2797–2806. [Google Scholar] [CrossRef]
- Lehnertz, B.; Ueda, Y.; Derijck, A.A.; Braunschweig, U.; Perez-Burgos, L.; Kubicek, S.; Chen, T.; Li, E.; Jenuwein, T.; Peters, A.H. Suv39h-Mediated Histone H3 Lysine 9 Methylation Directs DNA Methylation to Major Satellite Repeats at Pericentric Heterochromatin. Curr. Biol. 2003, 13, 1192–1200. [Google Scholar] [CrossRef]
- Probst, A.; Okamoto, I.; Casanova, M.; El Marjou, F.; Le Baccon, P.; Almouzni, G. A Strand-Specific Burst in Transcription of Pericentric Satellites Is Required for Chromocenter Formation and Early Mouse Development. Dev. Cell 2010, 19, 625–638. [Google Scholar] [CrossRef] [PubMed]
- Yadav, R.P.; Mäkelä, J.-A.; Hyssälä, H.; Cisneros-Montalvo, S.; Kotaja, N. DICER regulates the expression of major satellite repeat transcripts and meiotic chromosome segregation during spermatogenesis. Nucleic Acids Res. 2020, 48, 7135–7153. [Google Scholar] [CrossRef]
- Kishi, Y.; Kondo, S.; Gotoh, Y. Transcriptional Activation of Mouse Major Satellite Regions during Neuronal Differentiation. Cell Struct. Funct. 2012, 37, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Camacho, O.V.; Galan, C.; Swist-Rosowska, K.; Ching, R.; Gamalinda, M.; Karabiber, F.; De La Rosa-Velazquez, I.; Engist, B.; Koschorz, B.; Shukeir, N.; et al. Major satellite repeat RNA stabilize heterochromatin retention of Suv39h enzymes by RNA-nucleosome association and RNA:DNA hybrid formation. eLife 2017, 6, e25293. [Google Scholar] [CrossRef] [PubMed]
- Huo, X.; Ji, L.; Zhang, Y.; Lv, P.; Cao, X.; Wang, Q.; Yan, Z.; Dong, S.; Du, D.; Zhang, F.; et al. The Nuclear Matrix Protein SAFB Cooperates with Major Satellite RNAs to Stabilize Heterochromatin Architecture Partially through Phase Separation. Mol. Cell 2020, 77, 368–383. [Google Scholar] [CrossRef] [PubMed]
- Maison, C.; Bailly, D.; Peters, A.H.; Quivy, J.-P.; Roche, D.; Taddei, A.; Lachner, M.; Jenuwein, T.; Almouzni, G. Higher-order structure in pericentric heterochromatin involves a distinct pattern of histone modification and an RNA component. Nat. Genet. 2002, 30, 329–334. [Google Scholar] [CrossRef]
- Thakur, J.; Henikoff, S. Architectural RNA in chromatin organization. Biochem. Soc. Trans. 2020, 48, 1967–1978. [Google Scholar] [CrossRef] [PubMed]
- Thakur, J.; Fang, H.; Llagas, T.; Disteche, C.M.; Henikoff, S. Architectural RNA is required for heterochromatin organization. bioRxiv 2019, 784835. [Google Scholar] [CrossRef]
- Brown, K.E.; Guest, S.S.; Smale, S.T.; Hahm, K.; Merkenschlager, M.; Fisher, A.G. Association of Transcriptionally Silent Genes with Ikaros Complexes at Centromeric Heterochromatin. Cell 1997, 91, 845–854. [Google Scholar] [CrossRef]
- Hahm, K.; Cobb, B.S.; Mccarty, A.S.; Brown, K.E.; Klug, C.A.; Lee, R.; Akashi, K.; Weissman, I.L.; Fisher, A.G.; Smale, S.T. Helios, a T cell-restricted Ikaros family member that quantitatively associates with Ikaros at centromeric heterochromatin. Genes Dev. 1998, 12, 782–796. [Google Scholar] [CrossRef] [PubMed]
- Cobb, B.S.; Morales-Alcelay, S.; Kleiger, G.; Brown, K.E.; Fisher, A.G.; Smale, S.T. Targeting of Ikaros to pericentromeric heterochromatin by direct DNA binding. Genes Dev. 2000, 14, 2146–2160. [Google Scholar] [CrossRef]
- Gurel, Z.; Ronni, T.; Ho, S.; Kuchar, J.; Payne, K.J.; Turk, C.W.; Dovat, S. Recruitment of Ikaros to Pericentromeric Heterochromatin Is Regulated by Phosphorylation. J. Biol. Chem. 2008, 283, 8291–8300. [Google Scholar] [CrossRef]
- Kim, J.-H.; Ebersole, T.; Kouprina, N.; Noskov, V.N.; Ohzeki, J.-I.; Masumoto, H.; Mravinac, B.; Sullivan, B.A.; Pavlicek, A.; Dovat, S.; et al. Human gamma-satellite DNA maintains open chromatin structure and protects a transgene from epigenetic silencing. Genome Res. 2009, 19, 533–544. [Google Scholar] [CrossRef]
- Koering, C.E.; Pollice, A.; Zibella, M.P.; Bauwens, S.; Puisieux, A.; Brunori, M.; Brun, C.; Martins, L.; Sabatier, L.; Pulitzer, J.F.; et al. Human telomeric position effect is determined by chromosomal context and telomeric chromatin integrity. EMBO Rep. 2002, 3, 1055–1061. [Google Scholar] [CrossRef]
- Baur, J.A.; Zou, Y.; Shay, J.W.; Wright, W.E. Telomere Position Effect in Human Cells. Science 2001, 292, 2075–2077. [Google Scholar] [CrossRef] [PubMed]
- Pisano, S.; Pascucci, E.; Cacchione, S.; de Santis, P.; Savino, M. AFM imaging and theoretical modeling studies of sequence-dependent nucleosome positioning. Biophys. Chem. 2006, 124, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Makarov, V.L.; Lejnine, S.; Bedoyan, J.; Langmore, J.P. Nucleosomal organization of telomere-specific chromatin in rat. Cell 1993, 73, 775–787. [Google Scholar] [CrossRef]
- Mechelli, R.; Anselmi, C.; Cacchione, S.; de Santis, P.; Savino, M. Organization of telomeric nucleosomes: Atomic force microscopy imaging and theoretical modeling. FEBS Lett. 2004, 566, 131–135. [Google Scholar] [CrossRef]
- Filesi, I.; Cacchione, S.; de Santis, P.; Rossetti, L.; Savino, M. The main role of the sequence-dependent DNA elasticity in determining the free energy of nucleosome formation on telomeric DNAs. Biophys. Chem. 2000, 83, 223–237. [Google Scholar] [CrossRef]
- Ichikawa, Y.; Morohashi, N.; Nishimura, Y.; Kurumizaka, H.; Shimizu, M. Telomeric repeats act as nucleosome-disfavouring sequences In Vivo. Nucleic Acids Res. 2013, 42, 1541–1552. [Google Scholar] [CrossRef]
- Blasco, M.A. The epigenetic regulation of mammalian telomeres. Nat. Rev. Genet. 2007, 8, 299–309. [Google Scholar] [CrossRef]
- Jones, B.; Su, H.; Bhat, A.; Lei, H.; Bajko, J.; Hevi, S.; Baltus, G.A.; Kadam, S.; Zhai, H.; Valdez, R.; et al. The Histone H3K79 Methyltransferase Dot1L Is Essential for Mammalian Development and Heterochromatin Structure. PLoS Genet. 2008, 4, e1000190. [Google Scholar] [CrossRef]
- Bandaria, J.N.; Qin, P.; Berk, V.; Chu, S.; Yildiz, A. Shelterin Protects Chromosome Ends by Compacting Telomeric Chromatin. Cell 2016, 164, 735–746. [Google Scholar] [CrossRef] [PubMed]
- Raffa, G.D.; Ciapponi, L.; Cenci, G.; Gatti, M. Terminin: A protein complex that mediates epigenetic maintenance of Drosophila telomeres. Nucl. 2011, 2, 383–391. [Google Scholar] [CrossRef]
- Frydrychova, R.C.; Mason, J.M.; Archer, T.K. HP1 Is Distributed within Distinct Chromatin Domains at Drosophila Telomeres. Genetics 2008, 180, 121–131. [Google Scholar] [CrossRef]
- Raffa, G.D.; Siriaco, G.; Cugusi, S.; Ciapponi, L.; Cenci, G.; Wojcik, E.; Gatti, M. The Drosophila modigliani (moi) gene encodes a HOAP-interacting protein required for telomere protection. Proc. Natl. Acad. Sci. USA 2009, 106, 2271–2276. [Google Scholar] [CrossRef] [PubMed]
- Benetti, R.; Gonzalo, S.; Jaco, I.; Schotta, G.; Klatt, P.; Jenuwein, T.; Blasco, M.A. Suv4-20h deficiency results in telomere elongation and derepression of telomere recombination. J. Cell Biol. 2007, 178, 925–936. [Google Scholar] [CrossRef] [PubMed]
- Gonzalo, S.; García-Cao, M.; Fraga, M.F.; Schotta, G.; Peters, A.H.F.M.; Cotter, S.E.; Eguía, R.; Dean, D.C.; Esteller, M.; Jenuwein, T.; et al. Role of the RB1 family in stabilizing histone methylation at constitutive heterochromatin. Nat. Cell Biol. 2005, 7, 420–428. [Google Scholar] [CrossRef] [PubMed]
- Gonzalo, S.; Blasco, M.A. Role of Rb Family in the Epigenetic Definition of Chromatin. Cell Cycle 2005, 4, 752–755. [Google Scholar] [CrossRef] [PubMed]
- Pedram, M.; Sprung, C.N.; Gao, Q.; Lo, A.W.I.; Reynolds, G.E.; Murnane, J.P. Telomere Position Effect and Silencing of Transgenes near Telomeres in the Mouse. Mol. Cell. Biol. 2006, 26, 1865–1878. [Google Scholar] [CrossRef]
- Garrido-Ramos, M.A. Satellite DNA: An Evolving Topic. Genes 2017, 8, 230. [Google Scholar] [CrossRef]
- Gonzalo, S.; Jaco, I.; Fraga, M.F.; Chen, T.; Li, E.; Esteller, M.; Blasco, M.A. DNA methyltransferases control telomere length and telomere recombination in mammalian cells. Nat. Cell Biol. 2006, 8, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Schoeftner, S.; Blasco, M.A. Developmentally regulated transcription of mammalian telomeres by DNA-dependent RNA polymerase II. Nat. Cell Biol. 2007, 10, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Azzalin, C.M.; Reichenbach, P.; Khoriauli, L.; Giulotto, E.; Lingner, J. Telomeric Repeat Containing RNA and RNA Surveillance Factors at Mammalian Chromosome Ends. Science 2007, 318, 798–801. [Google Scholar] [CrossRef] [PubMed]
- Schoeftner, S.; Blasco, M.A. Chromatin regulation and non-coding RNAs at mammalian telomeres. Semin. Cell Dev. Biol. 2010, 21, 186–193. [Google Scholar] [CrossRef]
- Horard, B.; Gilson, E. Telomeric RNA enters the game. Nat. Cell Biol. 2008, 10, 113–115. [Google Scholar] [CrossRef]
- Savitsky, M.; Kwon, D.; Georgiev, P.; Kalmykova, A.; Gvozdev, V. Telomere elongation is under the control of the RNAi-based mechanism in the Drosophila germline. Genes Dev. 2006, 20, 345–354. [Google Scholar] [CrossRef]
- Xu, Y.; Kimura, T.; Komiyama, M. Human telomere RNA and DNA form an intermolecular G-quadruplex. Nucleic Acids Symp. Ser. 2008, 52, 169–170. [Google Scholar] [CrossRef] [PubMed]
- Zahler, A.M.; Williamson, J.R.; Cech, T.R.; Prescott, D.M. Inhibition of telomerase by G-quartet DMA structures. Nat. Cell Biol. 1991, 350, 718–720. [Google Scholar] [CrossRef]
- Xu, Y.; Kaminaga, K.; Komiyama, M. Human telomeric RNA in G-quadruplex structure. Nucleic Acids Symp. Ser. 2008, 52, 175–176. [Google Scholar] [CrossRef] [PubMed]
- Phan, A.T. Human telomeric G-quadruplex: Structures of DNA and RNA sequences. FEBS J. 2010, 277, 1107–1117. [Google Scholar] [CrossRef]
- Meštrovic, N.; Mravinac, B.; Juan, C.; Ugarkovic, Ð.; Plohl, M. Comparative study of satellite sequences and phylogeny of five species from the genus Palorus (Insecta, Coleoptera). Genome 2000, 43, 776–785. [Google Scholar] [CrossRef] [PubMed]
- Willard, H.F. Evolution of alpha satellite. Curr. Opin. Genet. Dev. 1991, 1, 509–514. [Google Scholar] [CrossRef]
- Biscotti, M.A.; Olmo, E.; Heslop-Harrison, J.S. (Pat) Repetitive DNA in eukaryotic genomes. Chromosom. Res. 2015, 23, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Palomeque, T.; Lorite, P. Satellite DNA in insects: A review. Heredity 2008, 100, 564–573. [Google Scholar] [CrossRef] [PubMed]
- Mestrovic, N.; Plohl, M.; Mravinac, B.; Ugarkovic, D. Evolution of satellite DNAs from the genus Palorus--experimental evidence for the “library” hypothesis. Mol. Biol. Evol. 1998, 15, 1062–1068. [Google Scholar] [CrossRef] [PubMed]
- Fry, K.; Salser, W. Nucleotide sequences of HS-α satellite DNA from kangaroo rat Dipodomys ordii and characterization of similar sequences in other rodents. Cell 1977, 12, 1069–1084. [Google Scholar] [CrossRef]
- Ruiz-Ruano, F.J.; López-León, M.D.; Cabrero, J.; Camacho, J.P.M. High-throughput analysis of the satellitome illuminates satellite DNA evolution. Sci. Rep. 2016, 6, 28333. [Google Scholar] [CrossRef]
- Cesari, M.; Luchetti, A.; Passamonti, M.; Scali, V.; Mantovani, B. Polymerase chain reaction amplification of the Bag320 satellite family reveals the ancestral library and past gene conversion events in Bacillus rossius (Insecta Phasmatodea). Gene 2003, 312, 289–295. [Google Scholar] [CrossRef]
- del Bosque, M.Q.; Navajas-Pérez, R.; Panero, J.; Fernández-González, A.J.; Garrido-Ramos, M. A satellite DNA evolutionary analysis in the North American endemic dioecious plant Rumex hastatulus (Polygonaceae). Genome 2011, 54, 253–260. [Google Scholar] [CrossRef]
- del Bosque, M.E.Q.; López-Flores, I.; Suárez-Santiago, V.N.; Garrido-Ramos, M.A. Satellite-DNA diversification and the evolution of major lineages in Cardueae (Carduoideae Asteraceae). J. Plant Res. 2014, 127, 575–583. [Google Scholar] [CrossRef]
- Feliciello, I.; Picariello, O.; Chinali, G. The first characterisation of the overall variability of repetitive units in a species reveals unexpected features of satellite DNA. Gene 2005, 349, 153–164. [Google Scholar] [CrossRef]
- Meštrović, N.; Mravinac, B.; Pavlek, M.; Vojvoda-Zeljko, T.; Šatović, E.; Plohl, M. Structural and functional liaisons between transposable elements and satellite DNAs. Chromosom. Res. 2015, 23, 583–596. [Google Scholar] [CrossRef]
- Belyayev, A.; Josefiová, J.; Jandová, M.; Mahelka, V.; Krak, K.; Mandák, B. Transposons and satellite DNA: On the origin of the major satellite DNA family in the Chenopodium genome. Mob. DNA 2020, 11, 1–10. [Google Scholar] [CrossRef]
- del Bosque, M.E.Q.; López-Flores, I.; Suárez-Santiago, V.N.; Garrido-Ramos, M.A. Differential spreading of HinfI satellite DNA variants during radiation in Centaureinae. Ann. Bot. 2013, 112, 1793–1802. [Google Scholar] [CrossRef] [PubMed]
- Wei, K.H.-C.; Grenier, J.K.; Barbash, D.A.; Clark, A.G. Correlated variation and population differentiation in satellite DNA abundance among lines ofDrosophila melanogaster. Proc. Natl. Acad. Sci. USA 2014, 111, 18793–18798. [Google Scholar] [CrossRef] [PubMed]
- Durfy, S.J.; Willard, H.F. Concerted evolution of primate alpha satellite DNA: Evidence for an ancestral sequence shared by gorilla and human X chromosome alpha satellite. J. Mol. Biol. 1990, 216, 555–566. [Google Scholar] [CrossRef]
- Waye, J.S.; Willard, H.F. Concerted evolution of alpha satellite DNA: Evidence for species specificity and a general lack of sequence conservation among alphoid sequences of higher primates. Chromosoma 1989, 98, 273–279. [Google Scholar] [CrossRef]
- Kimura, M.; Crow, J.F. The number of alleles that can be maintained in a finite population. Genetics 1964, 49, 725–738. [Google Scholar] [CrossRef]
- Smith, G. Evolution of repeated DNA sequences by unequal crossover. Science 1976, 191, 528–535. [Google Scholar] [CrossRef]
- Smith, G.P. Unequal Crossover and the Evolution of Multigene Families. Cold Spring Harb. Symp. Quant. Biol. 1974, 38, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Perelson, A.S.; Bell, G.I. Mathematical models for the evolution of multigene families by unequal crossing over. Nat. Cell Biol. 1977, 265, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Stephan, W.; Cho, S. Possible role of natural selection in the formation of tandem-repetitive noncoding DNA. Genet. 1994, 136, 333–341. [Google Scholar] [CrossRef]
- Stephan, W. Tandem-repetitive noncoding DNA: Forms and forces. Mol. Biol. Evol. 1989, 6, 198–212. [Google Scholar] [CrossRef] [PubMed]
- Dover, G.A. Molecular drive: A cohesive mode of species evolution. Nature 1982, 299, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Dover, G. Molecular drive. Trends Genet. 2002, 18, 587–589. [Google Scholar] [CrossRef]
- Liao, D.; Pavelitz, T.; Kidd, J.R.; Kidd, K.K.; Weiner, A.M. Concerted evolution of the tandemly repeated genes encoding human U2 snRNA (the RNU2 locus) involves rapid intrachromosomal homogenization and rare interchromosomal gene conversion. EMBO J. 1997, 16, 588–598. [Google Scholar] [CrossRef]
- Rice, W.R. Game of Thrones at Human Centromeres II. A new molecular/evolutionary model. bioRxiv 2019. [Google Scholar] [CrossRef]
- Kobayashi, T. Recombination Regulation by Transcription-Induced Cohesin Dissociation in rDNA Repeats. Science 2005, 309, 1581–1584. [Google Scholar] [CrossRef]
- Malik, H.S.; Henikoff, S. Adaptive evolution of Cid, a centromere-specific histone in Drosophila. Genetics 2001, 157, 1293–1298. [Google Scholar]
- Rhoades, M.M. Preferential Segregation in Maize. Genetics 1942, 27, 395–407. [Google Scholar] [CrossRef]
- Iwata-Otsubo, A.; Dawicki-McKenna, J.M.; Akera, T.; Falk, S.J.; Chmátal, L.; Yang, K.; Sullivan, B.A.; Schultz, R.M.; Lampson, M.A.; Black, B.E. Expanded Satellite Repeats Amplify a Discrete CENP-A Nucleosome Assembly Site on Chromosomes that Drive in Female Meiosis. Curr. Biol. 2017, 27, 2365–2373. [Google Scholar] [CrossRef] [PubMed]
- Chmátal, L.; Gabriel, S.I.; Mitsainas, G.P.; Martínez-Vargas, J.; Ventura, J.; Searle, J.B.; Schultz, R.M.; Lampson, M.A. Centromere Strength Provides the Cell Biological Basis for Meiotic Drive and Karyotype Evolution in Mice. Curr. Biol. 2014, 24, 2295–2300. [Google Scholar] [CrossRef]
- Fishman, L.; Saunders, A. Centromere-Associated Female Meiotic Drive Entails Male Fitness Costs in Monkeyflowers. Science 2008, 322, 1559–1562. [Google Scholar] [CrossRef]
- Saint-Leandre, B.; Christopher, C.; Levine, M.T. Adaptive evolution of an essential telomere protein restricts telomeric retrotransposons. eLife 2020, 9, e60987. [Google Scholar] [CrossRef]
- Saint-Leandre, B.; Levine, M.T. The Telomere Paradox: Stable Genome Preservation with Rapidly Evolving Proteins. Trends Genet. 2020, 36, 232–242. [Google Scholar] [CrossRef]
- Pontremoli, C.; Forni, D.; Cagliani, R.; Pozzoli, U.; Clerici, M.; Sironi, M. Evolutionary rates of mammalian telomere-stability genes correlate with karyotype features and female germline expression. Nucleic Acids Res. 2018, 46, 7153–7168. [Google Scholar] [CrossRef]
- Cutter, A.D. The polymorphic prelude to Bateson–Dobzhansky–Muller incompatibilities. Trends Ecol. Evol. 2012, 27, 209–218. [Google Scholar] [CrossRef]
- Brideau, N.J.; Flores, H.A.; Wang, J.; Maheshwari, S.; Wang, X.; Barbash, D.A. Two Dobzhansky-Muller Genes Interact to Cause Hybrid Lethality in Drosophila. Science 2006, 314, 1292–1295. [Google Scholar] [CrossRef] [PubMed]
- Satyaki, P.R.V.; Cuykendall, T.N.; Wei, K.H.-C.; Brideau, N.J.; Kwak, H.; Aruna, S.; Ferree, P.M.; Ji, S.; Barbash, D.A. The Hmr and Lhr Hybrid Incompatibility Genes Suppress a Broad Range of Heterochromatic Repeats. PLoS Genet. 2014, 10, e1004240. [Google Scholar] [CrossRef] [PubMed]
- Ferree, P.M.; Barbash, D.A. Species-Specific Heterochromatin Prevents Mitotic Chromosome Segregation to Cause Hybrid Lethality in Drosophila. PLoS Biol. 2009, 7, e1000234. [Google Scholar] [CrossRef] [PubMed]
- Ting, D.T.; Lipson, D.; Paul, S.; Brannigan, B.W.; Akhavanfard, S.; Coffman, E.J.; Contino, G.; Deshpande, V.; Iafrate, A.J.; Letovsky, S.; et al. Aberrant Overexpression of Satellite Repeats in Pancreatic and Other Epithelial Cancers. Science 2011, 331, 593–596. [Google Scholar] [CrossRef]
- Bersani, F.; Lee, E.; Kharchenko, P.V.; Xu, A.W.; Liu, M.; Xega, K.; MacKenzie, O.C.; Brannigan, B.W.; Wittner, B.S.; Jung, H.; et al. Pericentromeric satellite repeat expansions through RNA-derived DNA intermediates in cancer. Proc. Natl. Acad. Sci. USA 2015, 112, 15148–15153. [Google Scholar] [CrossRef]
- Ferreira, D.; Meles, S.; Escudeiro, A.; Mendes-da-Silva, A.; Adega, F.; Chaves, R. Satellite non-coding RNAs: The emerging players in cells, cellular pathways and cancer. Chromosom. Res. 2015, 23, 479–493. [Google Scholar] [CrossRef] [PubMed]
- Jackson, K.; Yu, M.C.; Arakawa, K.; Fiala, E.; Youn, B.; Fiegl, H.; Müller-Holzner, E.; Widschwendter, M.; Ehrlich, M. DNA hypomethylation is prevalent even in low-grade breast cancers. Cancer Biol. Ther. 2004, 3, 1225–1231. [Google Scholar] [CrossRef] [PubMed]
- Vukic, M.; Daxinger, L. DNA methylation in disease: Immunodeficiency, Centromeric instability, Facial anomalies syndrome. Essays Biochem. 2019, 63, 773–783. [Google Scholar] [CrossRef] [PubMed]
- Miga, K.H.; Koren, S.; Rhie, A.; Vollger, M.R.; Gershman, A.; Bzikadze, A.; Brooks, S.; Howe, E.; Porubsky, D.; Logsdon, G.A.; et al. Telomere-to-telomere assembly of a complete human X chromosome. Nature 2020, 585, 79–84. [Google Scholar] [CrossRef]
- Logsdon, G.A.; Vollger, M.R.; Hsieh, P.; Mao, Y.; Liskovykh, M.A.; Koren, S.; Nurk, S.; Mercuri, L.; Dishuck, P.C.; Rhie, A.; et al. The structure, function and evolution of a complete human chromosome 8. Nat. Cell Biol. 2021, 1–7. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Organism | Repeat Unit | Length | References |
|---|---|---|---|
| Human | α-satellite | 171-bp | [4] |
| M. musculus | Minor satellite | 120-bp | [34,35] |
| MS3 | 150-bp | ||
| MS4 | 300-bp | ||
| Z. mays | CentC | 156-bp | [36,37,38] |
| CRM | ~7000-bp | ||
| O. sativa | RCS2 or CentO | 155-bp | [39] |
| CRR | 4400- and 9200-bp | ||
| L. nivea | LCS1 | 178-bp | [40] |
| A. thaliana | pAL1 | 178-bp | [41,42] |
| R. sativus | pRA5/pRB | ~177-bp | [43] |
| D. melanogaster | Prodsat 10-mer AATAACATAG | 10-bp | [44,45] |
| AATAT | 5-bp | ||
| AATAG | 5-bp | ||
| AAGAT | 5-bp | ||
| G2/Jockey-3 | Up to a few kb | ||
| D. simulans | 10-mer AATAGAATTG | 10-bp | [44,45] |
| Simcent1 | 154-bp | ||
| Simcent2 | 88-bp | ||
| G2/Jockey-3 | Up to a few kb |
| Organism | Repeat Unit | Monomer Length | Reference |
|---|---|---|---|
| Human | Satellite I | 17-bp, 25-bp | [23,62,102,105] |
| Satellite II | 10–80-bp | ||
| Satellite III | 5-bp, 10-bp | ||
| α Satellites | 68-bp | ||
| α Satellites | 220-bp | ||
| M. musculus | Major satellites | 234-bp | [24,34] |
| TRPC-21A | 21-bp | ||
| TLC | 146-bp | ||
| A.thaliana | pAL1/pAtMr | 180-bp | [42,117,118,119] |
| Athila retrotransposons | Up to 14,100-bp | ||
| 500 bp repeat | 500-bp | ||
| 160 bp repeat | 160-bp | ||
| Dispersed repeats-163A, 164A, 278A and 106B | 1000–2000-bp | ||
| S. bulbocastanum | 2D8 | 5900-bp | [120] |
| D. melanogaster | 1.688 gm/cm3 buoyant density family | 353-bp, 356-bp, 260-bp and 359-bp | [121,122,123] |
| Rsp | 120-bp + 120-bp | ||
| Prod | 10-bp | ||
| AAGAT | 5-bp | ||
| AATAG | 5-bp | ||
| AAGAG | 5-bp | ||
| P. subdepressus | T2A5T octanucleotide containing 72-bp-long repeat | 8-bp, 22-bp | [124] |
| C. americana | 189 bp repeat | 189-bp | [125] |
| C. carnifex | Complex satellite repeat | 211-bp, 477-bp, 633-bp | [126] |
| T. castaneum | TCAST1 | 377-bp and 362-bp | [127,128] |
| Tcast2a | 359-bp | ||
| Tcast2b | 179-bp | ||
| Tcast2c | 300-bp |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thakur, J.; Packiaraj, J.; Henikoff, S. Sequence, Chromatin and Evolution of Satellite DNA. Int. J. Mol. Sci. 2021, 22, 4309. https://doi.org/10.3390/ijms22094309
Thakur J, Packiaraj J, Henikoff S. Sequence, Chromatin and Evolution of Satellite DNA. International Journal of Molecular Sciences. 2021; 22(9):4309. https://doi.org/10.3390/ijms22094309
Chicago/Turabian StyleThakur, Jitendra, Jenika Packiaraj, and Steven Henikoff. 2021. "Sequence, Chromatin and Evolution of Satellite DNA" International Journal of Molecular Sciences 22, no. 9: 4309. https://doi.org/10.3390/ijms22094309
APA StyleThakur, J., Packiaraj, J., & Henikoff, S. (2021). Sequence, Chromatin and Evolution of Satellite DNA. International Journal of Molecular Sciences, 22(9), 4309. https://doi.org/10.3390/ijms22094309