
Neuromedin U, a Key Molecule in Metabolic Disorders

{kind=link}
{kind=link}
Abstract
1. Introduction
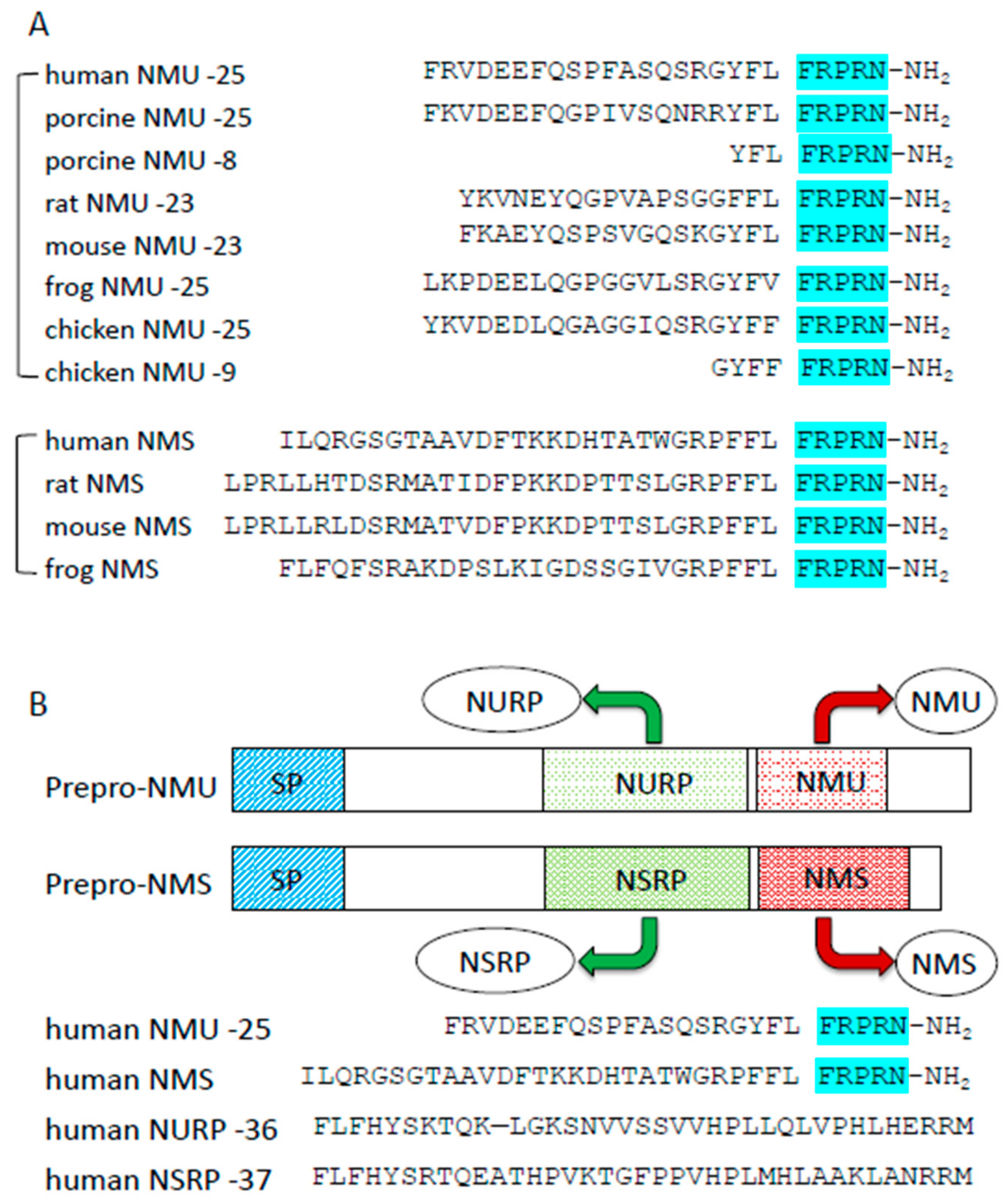
2. Biology of NMU and NMU-Related Peptides
2.1. Structure of NMU and Its Receptors
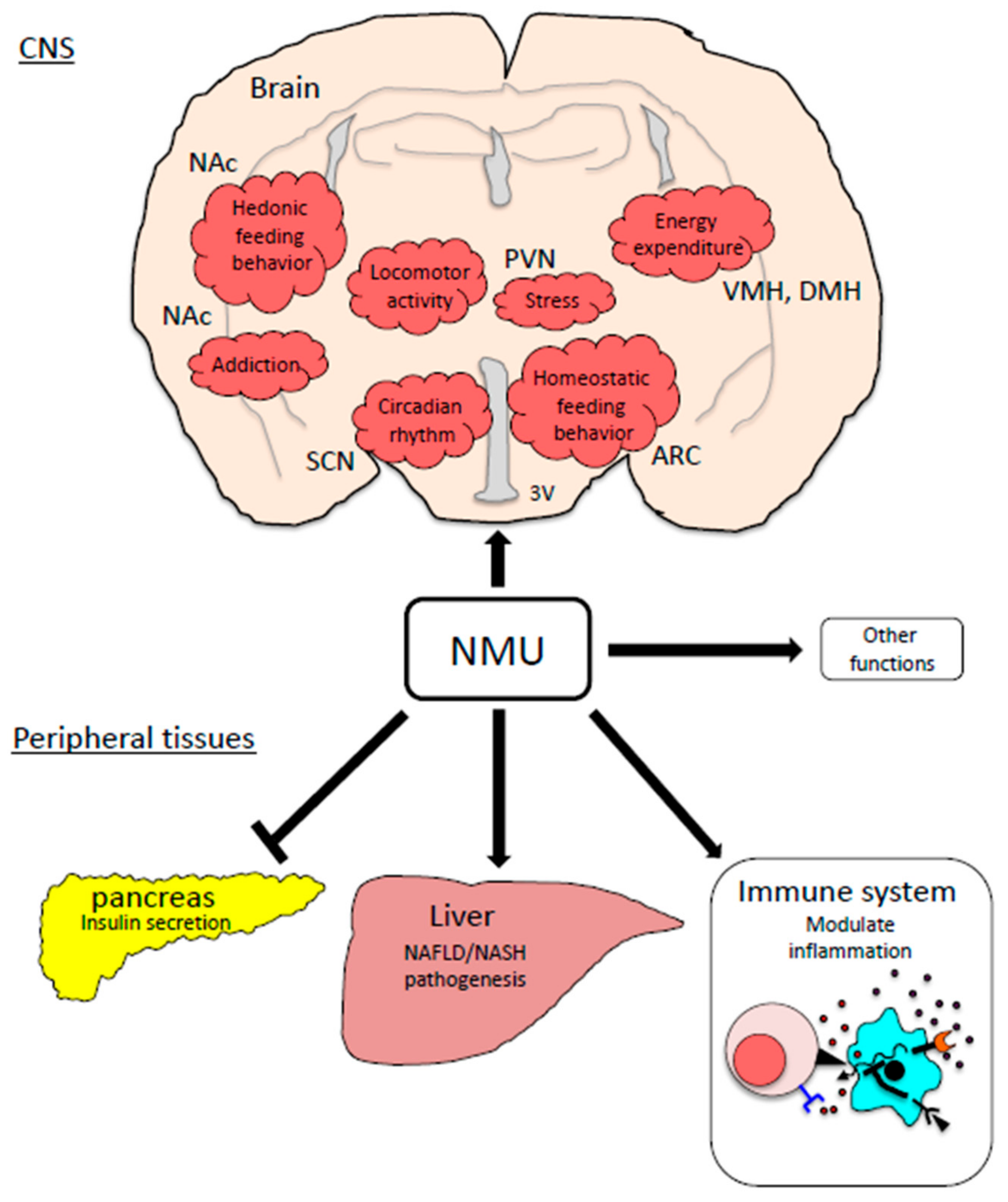
2.2. Distribution of NMU
2.3. NMU Receptor Distributions
2.4. Neuromedin S, Another Ligand of NMU Receptors
3. Homeostatic Regulation of Feeding Behavior
4. The Brain Reward System and Stress, and Their Relationships with the NMU System
5. NMU as a Modulator of Other Metabolic Disorders
6. Insulin Secretion
7. Inflammation
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cummings, D.E.; Schwartz, M.W. Genetics and Pathophysiology of Human Obesity. Annu. Rev. Med. 2003, 54, 453–471. [Google Scholar] [CrossRef] [PubMed]
- Kalra, S.P.; Dube, M.G.; Pu, S.; Xu, B.; Horvath, T.L.; Kalra, P.S. Interacting appetite-regulating pathways in the hy-pothalamic regulation of body weight. Endocr. Rev. 1999, 20, 68–100. [Google Scholar] [PubMed]
- Waterson, M.J.; Horvath, T.L. Neuronal Regulation of Energy Homeostasis: Beyond the Hypothalamus and Feeding. Cell Metab. 2015, 22, 962–970. [Google Scholar] [CrossRef] [PubMed]
- Minamino, N.; Kangawa, K.; Matsuo, H. Neuromedin U-8 and U-25: Novel uterus stimulating and hypertensive peptides identified in porcine spinal cord. Biochem. Biophys. Res. Commun. 1985, 130, 1078–1085. [Google Scholar] [CrossRef]
- Kojima, M.; Haruno, R.; Nakazato, M.; Date, Y.; Murakami, N.; Hanada, R.; Matsuo, H.; Kangawa, K. Purification and Identification of Neuromedin U as an Endogenous Ligand for an Orphan Receptor GPR66 (FM3). Biochem. Biophys. Res. Commun. 2000, 276, 435–438. [Google Scholar] [CrossRef]
- Howard, A.D.; Wang, R.; Pong, S.-S.; Mellin, T.N.; Strack, A.; Guan, X.-M.; Zeng, Z.; Jr, D.L.W.; Feighner, S.D.; Nunes, C.N.; et al. Identification of receptors for neuromedin U and its role in feeding. Nat. Cell Biol. 2000, 406, 70–74. [Google Scholar] [CrossRef]
- Nakazato, M.; Hanada, R.; Murakami, N.; Date, Y.; Mondal, M.S.; Kojima, M.; Yoshimatsu, H.; Kangawa, K.; Matsukura, S. Central Effects of Neuromedin U in the Regulation of Energy Homeostasis. Biochem. Biophys. Res. Commun. 2000, 277, 191–194. [Google Scholar] [CrossRef]
- Hanada, T.; Date, Y.; Shimbara, T.; Sakihara, S.; Murakami, N.; Hayashi, Y.; Kanai, Y.; Suda, T.; Kangawa, K.; Nakazato, M. Central actions of neuromedin U via corticotropin-releasing hormone. Biochem. Biophys. Res. Commun. 2003, 311, 954–958. [Google Scholar] [CrossRef]
- Hanada, R.; Nakazato, M.; Murakami, N.; Sakihara, S.; Yoshimatsu, H.; Toshinai, K.; Hanada, T.; Suda, T.; Kangawa, K.; Matsukura, S.; et al. A role for neuromedin U in stress response. Biochem. Biophys. Res. Commun. 2001, 289, 225–228. [Google Scholar] [CrossRef]
- Hanada, R.; Teranishi, H.; Pearson, J.T.; Kurokawa, M.; Hosoda, H.; Fukushima, N.; Fukue, Y.; Serino, R.; Fujihara, H.; Ueta, Y.; et al. Neuromedin U has a novel anorexigenic effect independent of the leptin signaling pathway. Nat. Med. 2004, 10, 1067–1073. [Google Scholar] [CrossRef]
- Nakahara, K.; Hanada, R.; Murakami, N.; Teranishi, H.; Ohgusu, H.; Fukushima, N.; Moriyama, M.; Ida, T.; Kangawa, K.; Kojima, M. The gut–brain peptide neuromedin U is involved in the mammalian circadian oscillator system. Biochem. Biophys. Res. Commun. 2004, 318, 156–161. [Google Scholar] [CrossRef]
- Moriyama, M.; Sato, T.; Inoue, H.; Fukuyama, S.; Teranishi, H.; Kangawa, K.; Kano, T.; Yoshimura, A.; Kojima, M. The neuropeptide neuromedin U promotes inflammation by direct activation of mast cells. J. Exp. Med. 2005, 202, 217–224. [Google Scholar] [CrossRef]
- Kowalski, T.J.; Spar, B.D.; Markowitz, L.; Maguire, M.; Golovko, A.; Yang, S.; Farley, C.; Cook, J.A.; Tetzloff, G.; Hoos, L.; et al. Transgenic overexpression of neuromedin U promotes leanness and hypophagia in mice. J. Endocrinol. 2005, 185, 151–164. [Google Scholar] [CrossRef]
- Minamino, N.; Kangawa, K.; Honzawa, M.; Matsuo, H. Isolation and structural determination of rat neuromedin U. Biochem. Biophys. Res. Commun. 1988, 156, 355–360. [Google Scholar] [CrossRef]
- Murphy, R.; Turner, C.; Furness, J.; Parker, L.; Giraud, A. Isolation and microsequence analysis of a novel form of neuromedin U from guinea pig small intestine. Peptides 1990, 11, 613–617. [Google Scholar] [CrossRef]
- Kage, R.; O’Harte, F.; Thim, L.; Conlon, J.M. Rabbit neuromedin U-25: Lack of conservation of a posttranslational processing site. Regul. Pept. 1991, 33, 191–198. [Google Scholar] [CrossRef]
- O’Harte, F.; Bockman, C.S.; Abel, P.W.; Conlon, J.M. Isolation, structural characterization and pharmacological ac-tivity of dog neuromedin U. Peptides 1991, 12, 11–15. [Google Scholar] [CrossRef]
- Austin, C.; Lo, G.; Nandha, K.A.; Meleagros, L.; Bloom, S.R. Cloning and characterization of the cDNA encoding the human neuromedin U (NmU) precursor: NmU expression in the human gastrointestinal tract. J. Mol. Endocrinol. 1995, 14, 157–169. [Google Scholar] [CrossRef]
- O’Harte, F.; Bockman, C.S.; Zeng, W.; Abel, P.W.; Harvey, S.; Conlon, J. Primary structure and pharmacological activity of a nonapeptide related to neuromedin U isolated from chicken intestine. Peptides 1991, 12, 809–812. [Google Scholar] [CrossRef]
- Domin, J.; Benito-Orfila, M.A.; Nandha, K.A.; Aitken, A.; Bloom, S.R. The purification and sequence analysis of an avian neuromedin U. Regul. Pept. 1992, 41, 1–8. [Google Scholar] [CrossRef]
- Domin, J.; Yiangou, Y.G.; Spokes, R.A.; Aitken, A.; Parmar, K.B.; Chrysanthou, B.J.; Bloom, S.R. The distribution, purification, and pharmacological action of an amphibian neuromedin U. J. Biol. Chem. 1989, 264, 20881–20885. [Google Scholar] [CrossRef]
- Salmon, A.L.; Johnsen, A.H.; Bienert, M.; McMurray, G.; Nandha, K.A.; Bloom, S.R.; Shaw, C. Isolation, Structural Characterization, and Bioactivity of a Novel Neuromedin U Analog from the Defensive Skin Secretion of the Australasian Tree Frog, Litoria caerulea. J. Biol. Chem. 2000, 275, 4549–4554. [Google Scholar] [CrossRef]
- Maruyama, K.; Konno, N.; Ishiguro, K.; Wakasugi, T.; Uchiyama, M.; Shioda, S.; Matsuda, K. Isolation and character-isation of four cDNAs encoding neuromedin U (NMU) from the brain and gut of goldfish, and the inhibitory effect of a deduced NMU on food intake and locomotor activity. J. Neuroendocrinol. 2008, 20, 71–78. [Google Scholar]
- Hedrick, J.A.; Morse, K.; Shan, L.; Qiao, X.; Pang, L.; Wang, S.; Laz, T.; Gustafson, E.L.; Bayne, M.; Monsma, F.J. Identification of a Human Gastrointestinal Tract and Immune System Receptor for the Peptide Neuromedin U. Mol. Pharmacol. 2000, 58, 870–875. [Google Scholar] [CrossRef] [PubMed]
- Funes, S.; Hedrick, J.A.; Yang, S.; Shan, L.; Bayne, M.; Monsma, F.J., Jr.; Gustafson, E.L. Cloning and characteriza-tion of murine neuromedin U receptors. Peptides 2002, 23, 1607–1615. [Google Scholar] [CrossRef]
- Sakura, N.; Ohta, S.; Uchida, Y.; Kurosawa, K.; Okimura, K.; Hashimoto, T. Structure-activity relationships of rat neu-romedin U for smooth muscle contraction. Chem. Pharm. Bull. 1991, 39, 2016–2020. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Fujii, R.; Hosoya, M.; Fukusumi, S.; Kawamata, Y.; Habata, Y.; Hinuma, S.; Onda, H.; Nishimura, O.; Fujino, M. Iden-tification of neuromedin U as the cognate ligand of the orphan G protein-coupled receptor FM-3. J. Biol. Chem. 2000, 275, 21068–21074. [Google Scholar] [CrossRef] [PubMed]
- Raddatz, R.; Wilson, A.E.; Artymyshyn, R.; Bonini, J.A.; Borowsky, B.; Boteju, L.W.; Zhou, S.; Kouranova, E.V.; Nagorny, R.; Guevarra, M.S.; et al. Identification and Characterization of Two Neuromedin U Receptors Differentially Expressed in Peripheral Tissues and the Central Nervous System. J. Biol. Chem. 2000, 275, 32452–32459. [Google Scholar] [CrossRef]
- Szekeres, P.G.; Muir, A.I.; Spinage, L.D.; Miller, J.E.; Butler, S.I.; Smith, A.; Rennie, G.I.; Murdock, P.R.; Fitzgerald, L.R.; Wu, H.-L.; et al. Neuromedin U Is a Potent Agonist at the Orphan G Protein-coupled Receptor FM3. J. Biol. Chem. 2000, 275, 20247–20250. [Google Scholar] [CrossRef]
- Hosoya, M.; Moriya, T.; Kawamata, Y.; Ohkubo, S.; Fujii, R.; Matsui, H.; Shintani, Y.; Fukusumi, S.; Habata, Y.; Hinuma, S.; et al. Identification and Functional Characterization of a Novel Subtype of Neuromedin U Receptor. J. Biol. Chem. 2000, 275, 29528–29532. [Google Scholar] [CrossRef]
- Shan, L.; Qiao, X.; Crona, J.H.; Behan, J.; Wang, S.; Laz, T.; Bayne, M.; Gustafson, E.L.; Monsma, F.J.; Hedrick, J.A. Identification of a Novel Neuromedin U Receptor Subtype Expressed in the Central Nervous System. J. Biol. Chem. 2000, 275, 39482–39486. [Google Scholar] [CrossRef]
- Holmgren, S.; Jensen, J. Evolution of vertebrate neuropeptides. Brain Res. Bull. 2001, 55, 723–735. [Google Scholar] [CrossRef]
- Hsu, S.H.; Luo, C.W. Molecular dissection of G protein preference using Gsalpha chimeras reveals novel ligand signal-ing of GPCRs. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E1021–E1029. [Google Scholar] [CrossRef]
- Torres, R.; Croll, S.D.; Vercollone, J.; Reinhardt, J.; Griffiths, J.; Zabski, S.; Anderson, K.D.; Adams, N.C.; Gowen, L.; Sleeman, M.W.; et al. Mice genetically deficient in neuromedin U receptor 2, but not neuromedin U receptor 1, have impaired nociceptive responses. Pain 2007, 130, 267–278. [Google Scholar] [CrossRef]
- Abbondanzo, S.J.; Manfra, D.J.; Chen, S.C.; Pinzon-Ortiz, M.; Sun, Y.; Phillips, J.E.; Laverty, M.; Vassileva, G.; Hu, W.; Yang, S.; et al. Nmur1−/− mice are not protected from cutaneous inflammation. Biochem. Biophys. Res. Commun. 2009, 378, 777–782. [Google Scholar] [CrossRef]
- Rao, S.M.; Auger, J.L.; Gaillard, P.; Weissleder, R.; Wada, E.; Torres, R.; Kojima, M.; Benoist, C.; Mathis, D.; Binstadt, B.A. The neuropeptide neuromedin U promotes autoantibody-mediated arthritis. Arthritis Res. Ther. 2012, 14, R29. [Google Scholar] [CrossRef]
- Takahashi, K.; Furukawa, C.; Takano, A.; Ishikawa, N.; Kato, T.; Hayama, S.; Suzuki, C.; Yasui, W.; Inai, K.; Sone, S.; et al. The Neuromedin U-Growth Hormone Secretagogue Receptor 1b/Neurotensin Receptor 1 Oncogenic Signaling Pathway as a Therapeutic Target for Lung Cancer. Cancer Res. 2006, 66, 9408–9419. [Google Scholar] [CrossRef]
- Matsuo, Y.; Yanase, Y.; Irifuku, R.; Takahagi, S.; Mihara, S.; Ishii, K.; Kawaguchi, T.; Tanaka, A.; Iwamoto, K.; Watanuki, H.; et al. Neuromedin U directly induces degranulation of skin mast cells, presumably via MRGPRX2. Allergy 2018, 73, 2256–2260. [Google Scholar] [CrossRef]
- Domin, J.; Ghatei, M.A.; Chohan, P.; Bloom, S.R. Neuromedin U—A study of its distribution in the rat. Peptides 1987, 8, 779–784. [Google Scholar] [CrossRef]
- Nakashima, Y.; Ida, T.; Sato, T.; Nakamura, Y.; Takahashi, T.; Mori, K.; Miyazato, M.; Kangawa, K.; Kusukawa, J.; Kojima, M. Neuromedin U is necessary for normal gastrointestinal motility and is regulated by serotonin. Ann. N. Y. Acad. Sci. 2010, 1200, 104–111. [Google Scholar] [CrossRef]
- Augood, S.; Keast, J.; Emson, P. Distribution and characterisation of neuromedin U-like immunoreactivity in rat brain and intestine and in guinea pig intestine. Regul. Pept. 1988, 20, 281–292. [Google Scholar] [CrossRef]
- Honzawa, M.; Sudoh, T.; Minamino, N.; Kangawa, K.; Matsuo, H. Neuromedin U-like immunoreactivity in rat intes-tine: Regional distribution and immunohistochemical study. Neuropeptides 1990, 15, 1–9. [Google Scholar] [CrossRef]
- Ballesta, J.; Carlei, F.; Bishop, A.E.; Steel, J.H.; Gibson, S.J.; Fahey, M.; Hennessey, R.; Domin, J.; Bloom, S.R.; Polak, J.M. Occurrence and developmental pattern of neuromedin U-immunoreactive nerves in the gastrointestinal tract and brain of the rat. Neuroscience 1988, 25, 797–816. [Google Scholar] [CrossRef]
- Furness, J.B.; Pompolo, S.; Murphy, R.; Giraud, A. Projections of neurons with neuromedin U-like immunoreactivity in the small intestine of the guinea-pig. Cell Tissue Res. 1989, 257, 415–422. [Google Scholar] [CrossRef]
- Domin, J.; Al-Madani, A.M.; Desperbasques, M.; Bishop, A.E.; Polak, J.M.; Bloom, S.R. Neuromedin U-like immuno-reactivity in the thyroid gland of the rat. Cell Tissue Res. 1990, 260, 131–135. [Google Scholar] [CrossRef]
- Graham, E.S.; Turnbull, Y.; Fotheringham, P.; Nilaweera, K.; Mercer, J.G.; Morgan, P.J.; Barrett, P. Neuromedin U and Neuromedin U receptor-2 expression in the mouse and rat hypothalamus: Effects of nutritional status. J. Neurochem. 2003, 87, 1165–1173. [Google Scholar] [CrossRef]
- Westfall, T.D.; McCafferty, G.P.; Pullen, M.; Gruver, S.; Sulpizio, A.C.; Aiyar, V.N.; Disa, J.; Contino, L.C.; Mannan, I.J.; Hieble, J.P. Characterization of neuromedin U effects in canine smooth muscle. J. Pharmacol. Exp. Ther. 2002, 301, 987–992. [Google Scholar] [CrossRef]
- Mori, K.; Miyazato, M.; Ida, T.; Murakami, N.; Serino, R.; Ueta, Y.; Kojima, M.; Kangawa, K. Identification of neu-romedin S and its possible role in the mammalian circadian oscillator system. EMBO J. 2005, 24, 325–335. [Google Scholar] [CrossRef]
- Mori, K.; Ida, T.; Fudetani, M.; Mori, M.; Kaiya, H.; Hino, J.; Nakahara, K.; Murakami, N.; Miyazato, M.; Kangawa, K. Identification of neuromedin U precursor-related peptide and its possible role in the regulation of prolactin release. Sci. Rep. 2017, 7, 10468. [Google Scholar] [CrossRef]
- Nakahara, K.; Maruyama, K.; Ensho, T.; Mori, K.; Miyazato, M.; Kangawa, K.; Uemura, R.; Sakoda, H.; Nakazato, M.; Murakami, N. Neuromedin U suppresses prolactin secretion via dopamine neurons of the arcuate nucleus. Biochem. Biophys. Res. Commun. 2020, 521, 521–526. [Google Scholar] [CrossRef]
- Ensho, T.; Maruyama, K.; Mori, K.; Miyazato, M.; Kangawa, K.; Nakahara, K.; Murakami, N. Neuromedin U precur-sor-related peptide (NURP) exerts neuromedin U-like sympathetic nerve action in the rat. Biochem. Biophys. Res. Commun. 2017, 492, 412–418. [Google Scholar] [CrossRef]
- Niimi, M.; Murao, K.; Taminato, T. Central administration of neuromedin U activates neurons in ventrobasal hypo-thalamus and brainstem. Endocrine 2001, 16, 201–206. [Google Scholar] [CrossRef]
- Ivanov, T.R.; Lawrence, C.B.; Stanley, P.J.; Luckman, S.M. Evaluation of neuromedin U actions in energy homeo-stasis and pituitary function. Endocrinology 2002, 143, 3813–3821. [Google Scholar] [CrossRef]
- Ozaki, Y.; Onaka, T.; Nakazato, M.; Saito, J.; Kanemoto, K.; Matsumoto, T.; Ueta, Y. Centrally administered neu-romedin U activates neurosecretion and induction of c-fos messenger ribonucleic acid in the paraventricular and su-praoptic nuclei of rat. Endocrinology 2002, 143, 4320–4329. [Google Scholar] [CrossRef][Green Version]
- Yokota, M.; Ozaki, Y.; Sakamoto, F.; Yamada, S.; Saito, J.; Fujihara, H.; Ueta, Y. Fos Expression in CRF-containing Neurons in the Rat Paraventricular Nucleus after Central Administration of Neuromedin U. Stress 2004, 7, 109–112. [Google Scholar] [CrossRef]
- Fang, L.; Zhang, M.; Li, C.; Dong, S.; Hu, Y. Chemical genetic analysis reveals the effects of NMU2R on the expression of peptide hormones. Neurosci. Lett. 2006, 404, 148–153. [Google Scholar] [CrossRef]
- Zeng, H.; Gragerov, A.; Hohmann, J.G.; Pavlova, M.N.; Schimpf, B.A.; Xu, H.; Wu, L.-J.; Toyoda, H.; Zhao, M.-G.; Rohde, A.D.; et al. Neuromedin U Receptor 2-Deficient Mice Display Differential Responses in Sensory Perception, Stress, and Feeding. Mol. Cell. Biol. 2006, 26, 9352–9363. [Google Scholar] [CrossRef]
- Takayama, K.; Mori, K.; Tanaka, A.; Sasaki, Y.; Sohma, Y.; Taguchi, A.; Taniguchi, A.; Sakane, T.; Yamamoto, A.; Miyazato, M.; et al. A chemically stable peptide agonist to neuromedin U receptor type 2. Bioorganic Med. Chem. 2020, 28, 115454. [Google Scholar] [CrossRef]
- Tanaka, A.; Takayama, K.; Furubayashi, T.; Mori, K.; Takemura, Y.; Amano, M.; Maeda, C.; Inoue, D.; Kimura, S.; Kiriyama, A.; et al. Transnasal Delivery of the Peptide Agonist Specific to Neuromedin-U Receptor 2 to the Brain for the Treatment of Obesity. Mol. Pharm. 2019, 17, 32–39. [Google Scholar] [CrossRef]
- Schwartz, M.W.; Seeley, R.J.; Campfield, L.A.; Burn, P.; Baskin, D.G. Identification of targets of leptin action in rat hypothalamus. J. Clin. Investig. 1996, 98, 1101–1106. [Google Scholar] [CrossRef]
- Schwartz, M.W.; Seeley, R.J.; Woods, S.C.; Weigle, D.S.; Campfield, L.A.; Burn, P.; Baskin, D.G. Leptin increases hypothalamic pro-opiomelanocortin mRNA expression in the rostral arcuate nucleus. Diabetes 1997, 46, 2119–2123. [Google Scholar] [CrossRef] [PubMed]
- Wren, A.M.; Small, C.J.; Abbott, C.R.; Jethwa, P.; Kennedy, A.R.; Murphy, K.G.; Stanley, S.A.; Zollner, A.N.; Ghatei, M.A.; Bloom, S.R. Hypothalamic Actions of Neuromedin U. Endocrinology 2002, 143, 4227–4234. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.W.; Dinulescu, D.M.; Cone, R.D. Genetic Models of Obesity and Energy Balance in the Mouse. Annu. Rev. Genet. 2000, 34, 687–745. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M.W.; Peskind, E.; Raskind, M.; Boyko, E.J.; Porte, D. Cerebrospinal fluid leptin levels: Relationship to plasma levels and to adiposity in humans. Nat. Med. 1996, 2, 589–593. [Google Scholar] [CrossRef]
- Caro, J.F.; Kolaczynski, J.W.; Nyce, M.R.; Ohannesian, J.P.; Opentanova, I.; Goldman, W.H.; Lynn, R.B.; Zhang, P.-L.; Sinha, M.K.; Considine, R.V. Decreased cerebrospinal-fluid/serum leptin ratio in obesity: A possible mechanism for leptin resistance. Lancet 1996, 348, 159–161. [Google Scholar] [CrossRef]
- Friedman, J.M.; Halaas, J.L. Leptin and the regulation of body weight in mammals. Nat. Cell Biol. 1998, 395, 763–770. [Google Scholar] [CrossRef]
- Jethwa, P.H.; Smith, K.L.; Small, C.J.; Abbott, C.R.; Darch, S.J.; Murphy, K.G.; Seth, A.; Semjonous, N.M.; Patel, S.R.; Todd, J.F.; et al. Neuromedin U partially mediates leptin-induced hypothalamo-pituitary ad-renal (HPA) stimulation and has a physiological role in the regulation of the HPA axis in the rat. Endocrinology 2006, 147, 2886–2892. [Google Scholar] [CrossRef][Green Version]
- Peier, A.M.; Desai, K.; Hubert, J.; Du, X.; Yang, L.; Qian, Y.; Kosinski, J.R.; Metzger, J.M.; Pocai, A.; Nawrocki, A.R.; et al. Effects of Peripherally Administered Neuromedin U on Energy and Glucose Homeostasis. Endocrinology 2011, 152, 2644–2654. [Google Scholar] [CrossRef]
- Hainerová, I.; Torekov, S.S.; Ek, J.; Finková, M.; Borch-Johnsen, K.; Jørgensen, T.; Madsen, O.D.; Lebl, J.; Hansen, T.; Pedersen, O. Association between Neuromedin U Gene Variants and Overweight and Obesity. J. Clin. Endocrinol. Metab. 2006, 91, 5057–5063. [Google Scholar] [CrossRef]
- Kenny, P.J. Reward Mechanisms in Obesity: New Insights and Future Directions. Neuron 2011, 69, 664–679. [Google Scholar] [CrossRef]
- Lutter, M.; Nestler, E.J. Homeostatic and Hedonic Signals Interact in the Regulation of Food Intake. J. Nutr. 2009, 139, 629–632. [Google Scholar] [CrossRef]
- Egecioglu, E.; Ploj, K.; Xu, X.; Bjursell, M.; Salomé, N.; Andersson, N.; Ohlsson, C.; Taube, M.; Hansson, C.; Bohlooly, M.Y.; et al. Central NMU signaling in body weight and energy balance regulation: Evidence from NMUR2 deletion and chronic central NMU treatment in mice. Am. J. Physiol. Metab. 2009, 297, E708–E716. [Google Scholar] [CrossRef]
- Peier, A.; Kosinski, J.; Cox-York, K.; Qian, Y.; Desai, K.; Feng, Y.; Trivedi, P.; Hastings, N.; Marsh, D.J. The Antiobesity Effects of Centrally Administered Neuromedin U and Neuromedin S Are Mediated Predominantly by the Neuromedin U Receptor 2 (NMUR2). Endocrinology 2009, 150, 3101–3109. [Google Scholar] [CrossRef]
- Benzon, C.; Johnson, S.; McCue, D.; Li, D.; Green, T.; Hommel, J. Neuromedin U receptor 2 knockdown in the paraventricular nucleus modifies behavioral responses to obesogenic high-fat food and leads to increased body weight. Neuroscience 2014, 258, 270–279. [Google Scholar] [CrossRef]
- Ignar, D.M.; Goetz, A.S.; Noble, K.N.; Carballo, L.H.; Stroup, A.E.; Fisher, J.C.; Boucheron, J.A.; Brainard, T.A.; Larkin, A.L.; Epperly, A.H.; et al. Regulation of ingestive be-haviors in the rat by GSK1521498, a novel micro-opioid receptor-selective inverse agonist. J. Pharmacol. Exp. Ther. 2011, 339, 24–34. [Google Scholar] [CrossRef]
- Corwin, R.L.; Avena, N.M.; Boggiano, M.M. Feeding and reward: Perspectives from three rat models of binge eating. Physiol. Behav. 2011, 104, 87–97. [Google Scholar] [CrossRef]
- Jerlhag, E. Gut-brain axis and addictive disorders: A review with focus on alcohol and drugs of abuse. Pharmacol. Ther. 2019, 196, 1–14. [Google Scholar] [CrossRef]
- Berthoud, H.R.; Munzberg, H.; Morrison, C.D. Blaming the Brain for Obesity: Integration of Hedonic and Homeostat-ic Mechanisms. Gastroenterology 2017, 152, 1728–1738. [Google Scholar] [CrossRef]
- Gumbs, M.C.R.; Heuvel, J.K.V.D.; La Fleur, S.E. The effect of obesogenic diets on brain Neuropeptide Y. Physiol. Behav. 2016, 162, 161–173. [Google Scholar] [CrossRef]
- Kasper, J.M.; McCue, D.L.; Milton, A.J.; Szwed, A.; Sampson, C.M.; Huang, M.; Carlton, S.; Meltzer, H.Y.; Cunning-ham, K.A.; Hommel, J.D. Gamma-Aminobutyric Acidergic Projections from the Dorsal Raphe to the Nucleus Accum-bens Are Regulated by Neuromedin U. Biol. Psychiatry 2016, 80, 878–887. [Google Scholar] [CrossRef]
- Vallof, D.; Ulenius, L.; Egecioglu, E.; Engel, J.A.; Jerlhag, E. Central administration of the anorexigenic peptide neu-romedin U decreases alcohol intake and attenuates alcohol-induced reward in rodents. Addict. Biol. 2017, 22, 640–651. [Google Scholar] [CrossRef] [PubMed]
- Vallof, D.; Vestlund, J.; Engel, J.A.; Jerlhag, E. The Anorexigenic Peptide Neuromedin U (NMU) Attenuates Ampheta-mine-Induced Locomotor Stimulation, Accumbal Dopamine Release and Expression of Conditioned Place Preference in Mice. PLoS ONE 2016, 11, e0154477. [Google Scholar] [CrossRef] [PubMed]
- Lydall, G.J.; Bass, N.J.; McQuillin, A.; Lawrence, J.; Anjorin, A.; Kandaswamy, R.; Pereira, A.; Guerrini, I.; Curtis, D.; Vine, A.E.; et al. Confirmation of prior evidence of genetic susceptibility to alcoholism in a genome-wide association study of comorbid alcoholism and bipolar disorder. Psychiatr. Genet. 2011, 21, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Vallof, D.; Kalafateli, A.L.; Jerlhag, E. Brain region-specific neuromedin U signalling regulates alcohol-related behav-iours and food intake in rodents. Addict. Biol. 2020, 25, e12764. [Google Scholar] [CrossRef]
- Anan, M.; Higa, R.; Shikano, K.; Shide, M.; Soda, A.; Apolinario, M.E.C.; Mori, K.; Shin, T.; Miyazato, M.; Mimata, H.; et al. Cocaine has some effect on neuromedin U expressing neurons related to the brain reward system. Heliyon 2020, 6, e03947. [Google Scholar] [CrossRef]
- Bowers, E.; Singer, K. Obesity-induced inflammation: The impact of the hematopoietic stem cell niche. JCI Insight 2021, 6, 3. [Google Scholar] [CrossRef]
- Sarma, S.; MacNamara, J.; Livingston, S.; Samels, M.; Haykowsky, M.J.; Berry, J.; Levine, B.D. Impact of severe obe-sity on exercise performance in heart failure with preserved ejection fraction. Physiol. Rep. 2020, 8, e14634. [Google Scholar] [CrossRef]
- Cohen, J.C.; Horton, J.D.; Hobbs, H.H. Human Fatty Liver Disease: Old Questions and New Insights. Science 2011, 332, 1519–1523. [Google Scholar] [CrossRef]
- Hebbard, L.; George, J. Animal models of nonalcoholic fatty liver disease. Nat. Rev. Gastroenterol. Hepatol. 2010, 8, 35–44. [Google Scholar] [CrossRef]
- Ludwig, J.; Viggiano, T.R.; McGill, D.B.; Oh, B.J. Nonalcoholic steatohepatitis: Mayo Clinic experiences with a hitherto unnamed disease. Mayo Clin. Proc. 1980, 55, 434–438. [Google Scholar]
- Tilg, H.; Adolph, T.E.; Moschen, A.R. Multiple Parallel Hits Hypothesis in Nonalcoholic Fatty Liver Disease: Revisited after a Decade. Hepatology 2021, 73, 833–842. [Google Scholar] [CrossRef]
- Teranishi, H.; Hayashi, M.; Higa, R.; Mori, K.; Miyazawa, T.; Hino, J.; Amano, Y.; Tozawa, R.; Ida, T.; Hanada, T.; et al. Role of neuromedin U in accelerating of non-alcoholic steatohepa-titis in mice. Peptides 2018, 99, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Sakoda, H.; Miura, A.; Shimizu, K.; Mori, K.; Miyazato, M.; Takayama, K.; Hayashi, Y.; Nakazato, M. Neuromedin U suppresses glucose-stimulated insulin secretion in pancreatic beta cells. Biochem. Biophys. Res. Commun. 2017, 493, 677–683. [Google Scholar] [CrossRef]
- Alfa, R.W.; Park, S.; Skelly, K.R.; Poffenberger, G.; Jain, N.; Gu, X.; Kockel, L.; Wang, J.; Liu, Y.; Powers, A.C.; et al. Suppression of insulin production and secretion by a decretin hormone. Cell Metab. 2015, 21, 323–334. [Google Scholar] [CrossRef]
- Takayama, K.; Mori, K.; Tanaka, A.; Nomura, E.; Sohma, Y.; Mori, M.; Taguchi, A.; Taniguchi, A.; Sakane, T.; Yamamoto, A.; et al. Discovery of a Human Neuromedin U Recep-tor 1-Selective Hexapeptide Agonist with Enhanced Serum Stability. J. Med. Chem. 2017, 60, 5228–5234. [Google Scholar] [CrossRef]
- Baggio, L.L.; Drucker, D.J. Biology of Incretins: GLP-1 and GIP. Gastroenterology 2007, 132, 2131–2157. [Google Scholar] [CrossRef]
- Ensho, T.; Maruyama, K.; Qattali, A.W.; Yasuda, M.; Uemura, R.; Murakami, N.; Nakahara, K. Comparison of glu-cose tolerance between wild-type mice and mice with double knockout of neuromedin U and neuromedin S. J. Vet. Med. Sci. 2019, 81, 1305–1312. [Google Scholar] [CrossRef]
- Wouters, K.; Gaens, K.; Bijnen, M.; Verboven, K.; Jocken, J.; Wetzels, S.; Wijnands, E.; Hansen, D.; Van Greevenbroek, M.; Duijvestijn, A.; et al. Circulating classical monocytes are associated with CD11c+ macrophages in human visceral adipose tissue. Sci. Rep. 2017, 7, 42665. [Google Scholar] [CrossRef]
- Asterholm, I.W.; Tao, C.; Morley, T.S.; Wang, Q.A.; Delgado-Lopez, F.; Wang, Z.V.; Scherer, P.E. Adipocyte Inflammation Is Essential for Healthy Adipose Tissue Expansion and Remodeling. Cell Metab. 2014, 20, 103–118. [Google Scholar] [CrossRef]
- Moriyama, M.; Fukuyama, S.; Inoue, H.; Matsumoto, T.; Satoru, F.; Tanaka, K.; Kinjyo, I.; Kano, T.; Yoshimura, A.; Kojima, M. The neuropeptide neuromedin U activates eosinophils and is involved in allergen-induced eosinophilia. Am. J. Physiol. Cell. Mol. Physiol. 2006, 290, L971–L977. [Google Scholar] [CrossRef]
- Moriyama, M.; Matsukawa, A.; Kudoh, S.; Takahashi, T.; Sato, T.; Kano, T.; Yoshimura, A.; Kojima, M. The neuro-peptide neuromedin U promotes IL-6 production from macrophages and endotoxin shock. Biochem. Biophys. Res. Commun. 2006, 341, 1149–1154. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.N.; Appelbaum, E.R.; Carpenter, D.C.; Cox, R.F.; Disa, J.; Foley, J.J.; Ghosh, S.K.; Naselsky, D.P.; Pullen, M.A.; Sarau, H.M.; et al. Neuromedin U elicits cytokine re-lease in murine Th2-type T cell clone D10.G4.1. J. Immunol. 2004, 173, 7230–7238. [Google Scholar] [CrossRef] [PubMed]
- Wallrapp, A.; Riesenfeld, S.J.; Burkett, P.R.; Abdulnour, R.E.; Nyman, J.; Dionne, D.; Hofree, M.; Cuoco, M.S.; Rod-man, C.; Farouq, D.; et al. The neuropeptide NMU am-plifies ILC2-driven allergic lung inflammation. Nature 2017, 549, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, V.; Chesné, J.; Ribeiro, H.; García-Cassani, B.; Carvalho, T.; Bouchery, T.; Shah, K.; Barbosa-Morais, N.L.; Harris, N.; Veiga-Fernandes, H. Neuronal regulation of type 2 innate lymphoid cells via neuromedin U. Nat. Cell Biol. 2017, 549, 277–281. [Google Scholar] [CrossRef]
- Klose, C.S.N.; Mahlakõiv, T.; Moeller, J.B.; Rankin, L.C.; Flamar, A.-L.; Kabata, H.; Monticelli, L.A.; Moriyama, S.; Putzel, G.G.; Rakhilin, N.; et al. The neuropeptide neuromedin U stimulates innate lymphoid cells and type 2 inflammation. Nat. Cell Biol. 2017, 549, 282–286. [Google Scholar] [CrossRef]
- Sugita, K.; Steer, C.A.; Martinez-Gonzalez, I.; Altunbulakli, C.; Morita, H.; Castro-Giner, F.; Kubo, T.; Wawrzyniak, P.; Ruckert, B.; Sudo, K.; et al. Type 2 innate lym-phoid cells disrupt bronchial epithelial barrier integrity by targeting tight junctions through IL-13 in asthmatic patients. J. Allergy Clin. Immunol. 2018, 141, 300–310.e11. [Google Scholar] [CrossRef]
- Serafini, N.; Vosshenrich, C.A.J.; Di Santo, J.P. Transcriptional regulation of innate lymphoid cell fate. Nat. Rev. Immunol. 2015, 15, 415–428. [Google Scholar] [CrossRef]
- Cording, S.; Medvedovic, J.; Aychek, T.; Eberl, G. Innate lymphoid cells in defense, immunopathology and immuno-therapy. Nat. Immunol. 2016, 17, 755–757. [Google Scholar] [CrossRef]
- Miethe, S.; Karsonova, A.; Karaulov, A.; Renz, H. Obesity and asthma. J. Allergy Clin. Immunol. 2020, 146, 685–693. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teranishi, H.; Hanada, R. Neuromedin U, a Key Molecule in Metabolic Disorders. Int. J. Mol. Sci. 2021, 22, 4238. https://doi.org/10.3390/ijms22084238
Teranishi H, Hanada R. Neuromedin U, a Key Molecule in Metabolic Disorders. International Journal of Molecular Sciences. 2021; 22(8):4238. https://doi.org/10.3390/ijms22084238
Chicago/Turabian StyleTeranishi, Hitoshi, and Reiko Hanada. 2021. "Neuromedin U, a Key Molecule in Metabolic Disorders" International Journal of Molecular Sciences 22, no. 8: 4238. https://doi.org/10.3390/ijms22084238
APA StyleTeranishi, H., & Hanada, R. (2021). Neuromedin U, a Key Molecule in Metabolic Disorders. International Journal of Molecular Sciences, 22(8), 4238. https://doi.org/10.3390/ijms22084238