
Alterations of the Skin and Gut Microbiome in Psoriasis and Psoriatic Arthritis

 ,
, 
Abstract
1. Introduction
2. Results
2.1. Microbiome Analysis Techniques
2.2. Alterations in Microbiome of the Skin in Psoriasis
2.3. The Role of Skin Microbiome in Psoriasis Pathogenesis
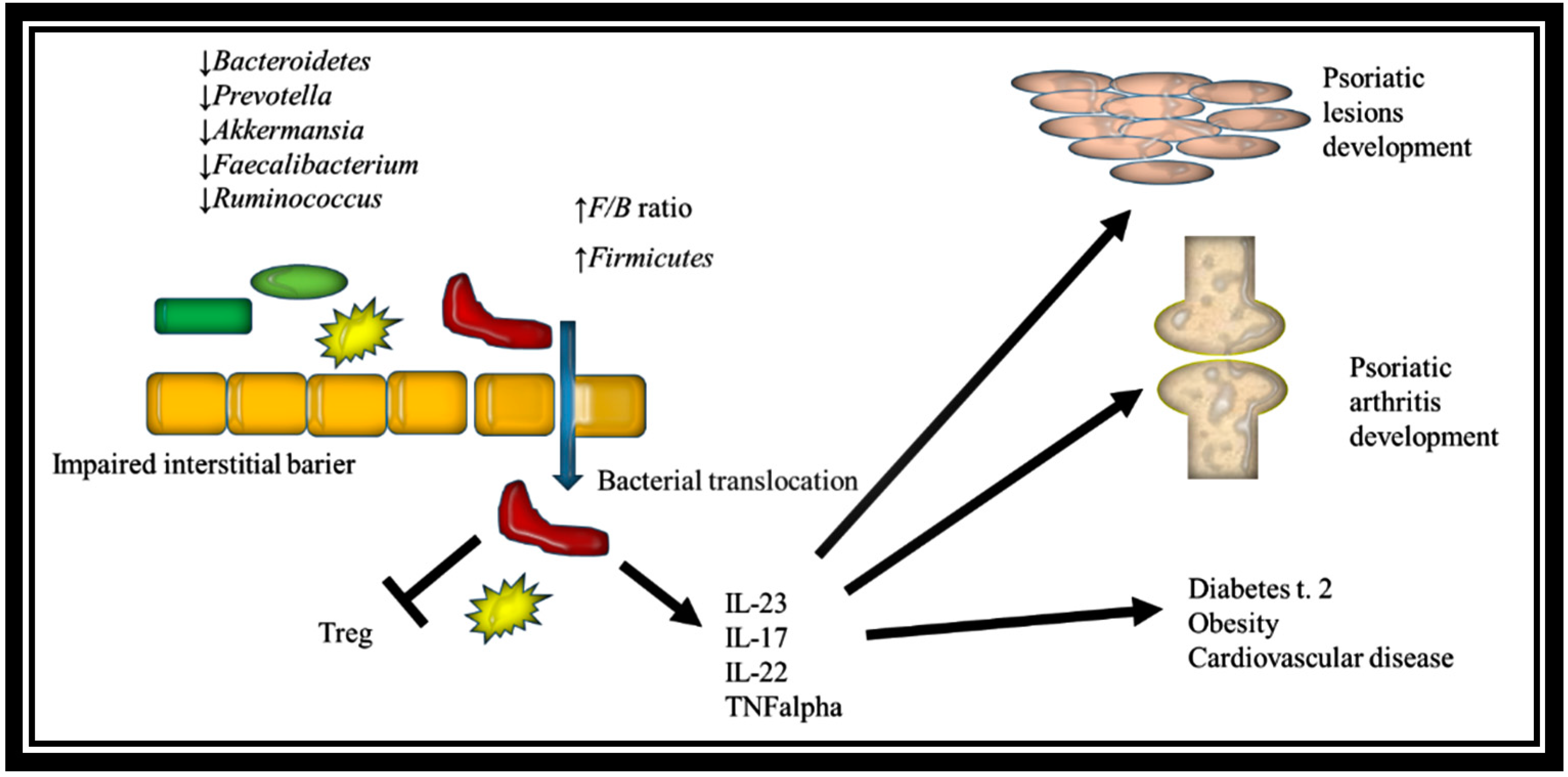
2.4. Gut Dysbiosis in Psoriasis
2.5. Role of Gut Microbiome in Psoriasis Pathogenesis
2.6. The Role of Intestinal Dysbiosis in Psoriatic Arthritis
2.7. Microbiome and Psoriasis Treatment
2.8. Probiotics and Dietary Approaches in Psoriasis
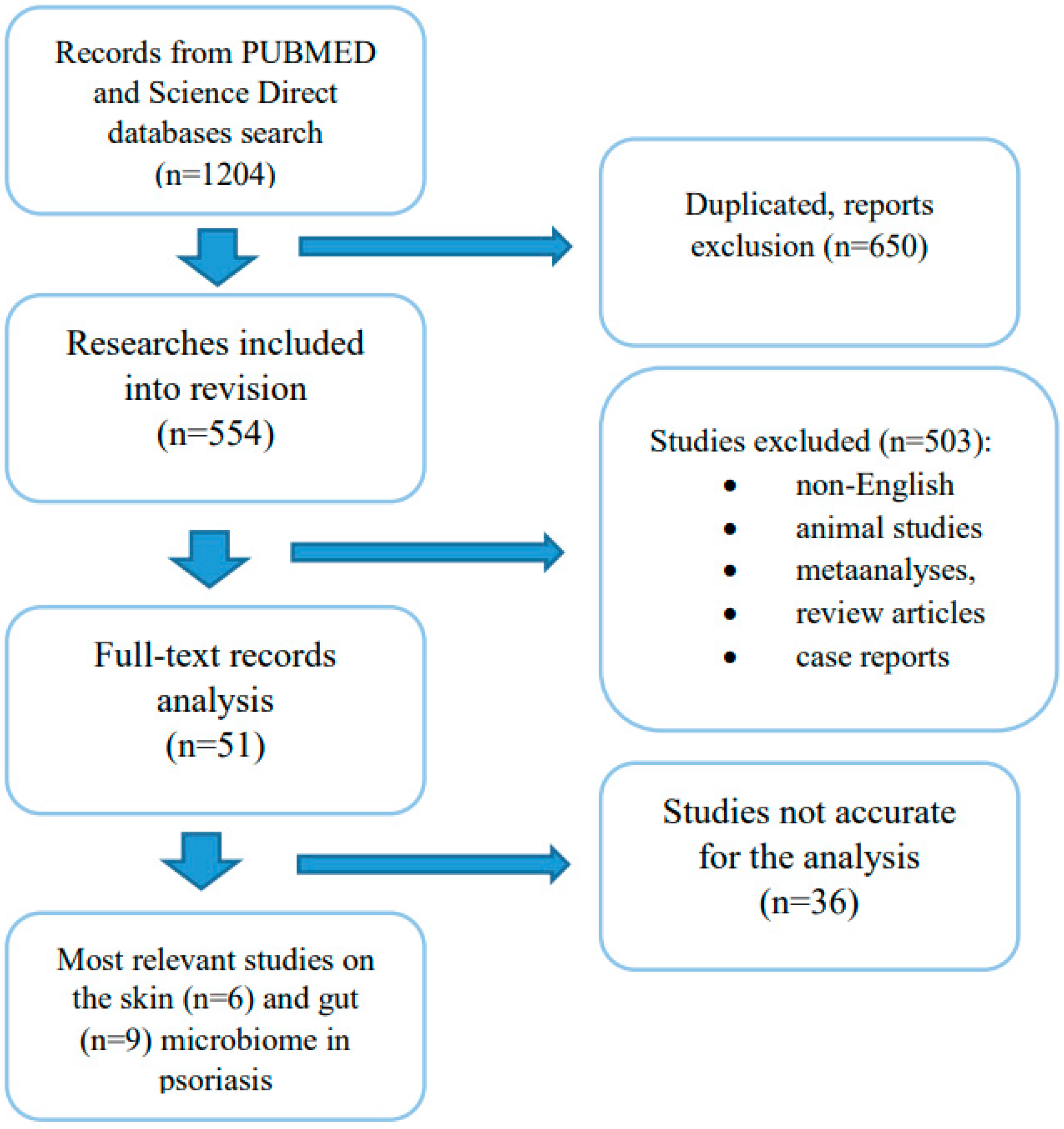
3. Materials and Methods
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| IBD | inflammatory bowel disease |
| PCR | polymerase chain reaction |
| rRNA | ribosomal ribonucleic acid |
| DNA | deoxyribonucleic acid |
| PASI | Psoriasis Activity and Severity Index |
| TNFα | tumour necrosis factor alpha |
| IL-1 | interleukin 1 |
| IL-6 | interleukin 6 |
| IL-8 | interleukin 8 |
| MAMPs | microbial-associated molecular patterns |
| PRRs | pattern recognition receptors |
| pDC | plasmacytoid dendritic cells |
| iNOS | inducible nitric oxide synthase |
| mDC | myeloid dendritic cells |
| Th17 | T helper 17 |
| IL-17 | interleukin 17 |
| IL-22 | interleukin 22 |
| IL-36α | interleukin 36 alpha |
| F/B ratio | Firmicutes-to-Bacteroidetes ratio |
| NF-κB | nuclear factor kappa light chain enhancer of activated B cells |
| MCFAs | medium-chain fatty acids |
| CRP | C-reactive protein |
| Kcasp1Tg-mice | keratinocyte-specific caspase-1 transgenic mice |
| I-FABP | intestinal fatty acid binding protein |
| IFN-γ | gamma interferon |
| Treg | T regulatory cells |
| SCFAs | short-chain fatty acids |
| PPAR | peroxisome proliferator-activated receptor |
| IAld | Indole-3-aldehyde |
| IgA | Immunoglobulin A |
| RANKL | receptor activator of nuclear factor kappa-B ligand |
| TMAO | Trimethylamine-N-oxide |
| TMA | trimethylamine |
| IL-23 | interleukin 23 |
| SpA | spondyloarthropathies |
| TCR | T-Cell Antigen Receptors |
| TNF∆ARE/+ mice | TNF overexpressing mice |
| MAIT | mucosal-associated invariant T |
| iNKT | invariant natural killer T cells |
| NB-UVB | narrow band ultraviolet radiation |
| MDA | malondialdehyde |
| ROS | reactive oxygen species |
| AGA | anti-gliadin antibodies |
| IBD | inflammatory bowel disease |
| PCR | polymerase chain reaction |
| rRNA | ribosomal ribonucleic acid |
| DNA | deoxyribonucleic acid |
| MAMPs | microbial-associated molecular patterns |
| pDC | plasmacytoid dendritic cells |
| TNFα | tumour necrosis factor alpha |
| IL-1 | interleukin 1 |
| IL-6 | interleukin 6 |
| IL-8 | interleukin 8 |
| iNOS | inducible nitric oxide synthase |
| mDC | myeloid dendritic cells |
| Kcasp1Tg-mice | keratinocyte-specific caspase-1 transgenic mice |
| Treg | T regulatory cells |
| SCFA | short-chain fatty acid |
| MCFAs | medium-chain fatty acids |
| PPAR | peroxisome proliferator-activated receptor |
| IAld | Indole-3-aldehyde |
| RANKL | receptor activator of nuclear factor kappa-B ligand |
| F/B ratio | Firmicutes to Bacteroidetes ratio |
| TMAO | Trimethylamine-N-oxide |
| SpA | spondyloarthropathies |
| TNF∆ARE/+ mice | TNF overexpressing mice |
| MAIT | mucosal-associated invariant T |
| iNKT | invariant natural killer T cells |
| NB-UVB | narrow band ultraviolet radiation |
| MDA | malondialdehyde |
| ROS | reactive oxygen species |
| CRP | C-reactive protein |
| AGA | anti-gliadin antibodies |
| PASI | Psoriasis Activity and Severity Index |
References
- Iannone, L.F.; Bennardo, L.; Palleria, C.; Roberti, R.; De Sarro, C.; Naturale, M.D.; Dastoli, S.; Donato, L.; Manti, A.; Valenti, G.; et al. Safety profile of biologic drugs for psoriasis in clinical practice: An Italian prospective pharmacovigilance study. PLoS ONE 2020, 15, e0241575. [Google Scholar] [CrossRef]
- Dattola, A.; Silvestri, M.; Bennardo, L.; Passante, M.; Rizzuto, F.; Dastoli, S.; Patruno, C.; Bianchi, L.; Nisticò, S.P. A novel vehicle for the treatment of psoriasis. Dermatol. Ther. 2020, 33, e13185. [Google Scholar] [CrossRef]
- Dattola, A.; Silvestri, M.; Tamburi, F.; Amoruso, G.F.; Bennardo, L.; Nisticò, S.P. Emerging role of anti-IL23 in the treatment of psoriasis: When humanized is very promising. Dermatol. Ther. 2020, 33, e14504. [Google Scholar] [CrossRef] [PubMed]
- Hsu, D.K.; Funga, M.A.; Chen, H.L. Role of skin and gut microbiota in the pathogenesis of psoriasis, an inflammatory skin disease. Med. Microecol. 2020, 100016, 1–8. [Google Scholar] [CrossRef]
- Oliveira, M.F.; Rocha, B.O.; Duarte, G.V. Psoriasis: Classical and emerging comorbidities. An. Bras. Dermatol. 2015, 90, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Langan, E.A.; Griffiths, C.E.M.; Solbach, W.; Knobloch, J.K.; Zillikens, D.; Thaçi, D. The role of the microbiome in psoriasis: Moving from disease description to treatment selection? Br. J. Dermatol. 2018, 178, 1020–1027. [Google Scholar] [CrossRef] [PubMed]
- Human Microbiome Project Consortium. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Grice, E.A.; Kong, H.H.; Conlan, S.; Deming, C.B.; Davis, J.; Young, A.C.; Bouffard, G.G.; Blakesley, R.W.; Murray, P.R.; Green, E.D.; et al. Topographical and temporal diversity of the human skin microbiome. Science 2009, 324, 1190–1192. [Google Scholar] [CrossRef]
- Grice, E.A.; Segre, J.A. The skin microbiome. Nat. Rev. Microbiol. 2011, 9, 244–253. [Google Scholar] [CrossRef]
- Yan, D.; Issa, N.; Afifi, L.; Jeon, C.; Chang, H.W.; Liao, W. The role of the skin and gut microbiome in psoriatic disease. Curr. Dermatol. Rep. 2017, 6, 94–103. [Google Scholar] [CrossRef]
- Alekseyenko, A.V.; Perez-Perez, G.I.; De Souza, A.; Strober, B.; Gao, Z.; Bihan, M.; Li, K.; Methe, B.A.; Blaser, M.J. Community differentiation of the cutaneous microbiota in psoriasis. Microbiome 2013, 1, 31. [Google Scholar] [CrossRef] [PubMed]
- Fyhrquist, N.; Muirhead, G.; Prast-Nielsen, S.; Jeanmougin, M.; Olah, P.; Skoog, T.; Jules-Clement, G.; Feld, M.; Barrientos-Somarribas, M.; Sinkko, H.; et al. Microbe-host interplay in atopic dermatitis and psoriasis. Nat. Commun. 2019, 10, 4703. [Google Scholar] [CrossRef] [PubMed]
- Quan, C.; Chen, X.Y.; Li, X.; Xue, F.; Chen, L.H.; Liu, N.; Wang, B.; Wang, L.Q.; Wang, X.P.; Yang, H.; et al. Psoriatic lesions are characterized by higher bacterial load and imbalance between Cutibacterium and Corynebacterium. J. Am. Acad. Dermatol. 2020, 82, 955–961. [Google Scholar] [CrossRef]
- Visser, M.J.E.; Kell, D.B.; Pretorius, E. Bacterial dysbiosis and translocation in psoriasis vulgaris. Front. Cell Infect. Microbiol. 2019, 9, 7. [Google Scholar] [CrossRef]
- Thio, H.B. The microbiome in psoriasis and psoriatic arthritis: The skin perspective. J. Rheumatol. Suppl. 2018, 94, 30–31. [Google Scholar] [CrossRef]
- Gao, Z.; Tseng, C.H.; Strober, B.E.; Pei, Z.; Blaser, M.J. Substantial alterations of the cutaneous bacterial biota in psoriatic lesions. PLoS ONE 2008, 3, e2719. [Google Scholar] [CrossRef]
- Fahlen, A.; Engstrand, L.; Baker, B.S.; Powles, A.; Fry, L. Comparison of bacterial microbiota in skin biopsies from normal and psoriatic skin. Arch. Dermatol. Res. 2012, 304, 15–22. [Google Scholar] [CrossRef]
- Lober, C.W.; Belew, P.W.; Rosenberg, E.W.; Bale, G. Patch tests with killed sonicated microflora in patients with psoriasis. Arch. Dermatol. 1982, 118, 322–325. [Google Scholar] [CrossRef] [PubMed]
- Engstrand, L.; Fry, L. Investigation of the skin microbiome: Swabs vs. biopsies. Br. J. Dermatol. 2019, 181, 572–579. [Google Scholar] [CrossRef]
- Dei-Cas, I.; Giliberto, F.; Luce, L. Metagenomic analysis of gut microbiota in non-treated plaque psoriasis patients stratified by disease severity: Development of a new Psoriasis-Microbiome Index. Sci. Rep. 2020, 10, 12754. [Google Scholar] [CrossRef] [PubMed]
- Luger, T.A.; Loser, K. Novel insights into the pathogenesis of psoriasis. Clin. Immunol. 2018, 186, 43–45. [Google Scholar] [CrossRef]
- Elfatoiki, F.Z.; El Azhari, M.; El Kettani, A.; Serhier, Z.; Othmani, M.B.; Timinouni, M.; Benchikhi, H.; Chiheb, S.; Fellah, H. Psoriasis and Staphylococcus aureus skin colonization in Moroccan patients. Pan Afr. Med. J. 2016, 23, 33. [Google Scholar] [CrossRef] [PubMed]
- Tett, A.; Pasolli, E.; Farina, S.; Truong, D.T.; Asnicar, F.; Zolfo, M. Unexplored diversity and strain-level structure of the skin microbiome associated with psoriasis. NPJ Biofilms Microbiomes 2017, 3, 14. [Google Scholar] [CrossRef] [PubMed]
- Stehlikova, Z.; Kostovcik, M.; Kostovcikova, K.; Kverka, M.; Juzlova, K.; Rob, F.; Hercogova, J. Dysbiosis of Skin Microbiota in Psoriatic Patients: Co-occurrence of Fungal and Bacterial Communities. Front. Microbiol. 2019, 10, 438. [Google Scholar] [CrossRef] [PubMed]
- Findley, K.; Oh, J.; Yang, J.; Conlan, S.; Deming, C.; Meyer, J.A. Topographic diversity of fungal and bacterial communities in human skin. Nature 2013, 498, 367–370. [Google Scholar] [CrossRef]
- Watanabe, S.; Kano, R.; Sato, H.; Nakamura, Y.; Hasegawa, A. The effects of Malassezia yeasts on cytokine production by human keratinocytes. J. Invest. Dermatol. 2001, 116, 769–773. [Google Scholar] [CrossRef] [PubMed]
- Fauler, J.; Neumann, C.; Tsikas, D.; Frolich, J. Enhanced synthesis of cysteinyl leukotrienes in psoriasis. J. Invest. Dermatol. 1992, 99, 8–11. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Buentke, E.; Heffler, L.C.; Wilson, J.L.; Wallin, R.P.; Lofman, C.; Chambers, B.J. Natural killer and dendritic cell contact in lesional atopic dermatitis skin–Malassezia-influenced cell interaction. J. Invest. Dermatol. 2002, 119, 850–857. [Google Scholar] [CrossRef]
- Selander, C.; Engblom, C.; Nilsson, G.; Scheynius, A.; Andersson, C.L. TLR2/MyD88-dependent and -independent activation of mast cell IgE responses by the skin commensal yeast Malassezia sympodialis. J. Immunol. 2009, 182, 4208–4216. [Google Scholar] [CrossRef]
- Chang, H.W.; Yan, D.; Singh, R.; Liu, J.; Lu, X.; Ucmak, D. Alteration of the cutaneous microbiome in psoriasis and potential role in Th17 polarization. Microbiome 2018, 6, 154. [Google Scholar] [CrossRef]
- Assarsson, M.; Soderman, J.; Dienus, O.; Seifert, O. Significant differences in the bacterial microbiome of the pharynx and skin in patients with psoriasis compared with healthy controls. Acta Derm. Venereol. 2020, 100, 00273. [Google Scholar] [CrossRef]
- Chu, H.; Mazmanian, S.K. Innate immune recognition of the microbiota promotes host-microbial symbiosis. Nat. Immunol. 2013, 14, 668–675. [Google Scholar] [CrossRef] [PubMed]
- Belkaid, Y.; Hand, T.W. Role of the microbiota in immunity and inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [PubMed]
- Fry, L.; Baker, B.S.; Powles, A.V. Is chronic plaque psoriasis triggered by microbiota in the skin? Br. J. Dermatol. 2013, 169, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Fry, L.; Baker, B.S.; Powles, A.V.; Engstrand, L. Psoriasis is not an autoimmune disease? Exp. Dermatol. 2015, 24, 241–244. [Google Scholar] [CrossRef] [PubMed]
- Hashiguchi, Y.; Yabe, R.; Chung, S.H.; Murayama, M.A.; Yoshida, K. IL-36alpha from skin-resident cells plays an important role in the pathogenesis of imiquimod-induced psoriasiform dermatitis by forming a local autoamplification loop. J. Immunol. 2018, 201, 167–182. [Google Scholar] [CrossRef]
- Zakostelska, Z.; Malkova, J.; Klimesova, K.; Rossmann, P.; Hornova, M.; Novosadova, I.; Stehlíkova, Z. Intestinal microbiota promotes psoriasis-like skin inflammation by enhancing Th17 response. PLoS ONE 2015, 11, e0159539. [Google Scholar] [CrossRef]
- Valdimarsson, H.; Baker, B.S.; Jonsdottir, I.; Powles, A.; Fry, L. Psoriasis: A T-cell-mediated autoimmune disease induced by streptococcal superantigens? Immunol. Today 1995, 16, 145–149. [Google Scholar] [CrossRef]
- Leung, D.Y.; Travers, J.B.; Giorno, R.; Norris, D.A.; Skinner, R.; Aelion, J.; Kazemi, L.V.; Kim, M.H.; Trumble, A.E.; Kotb, M.; et al. Evidence for a streptococcal superantigendriven process in acute guttate psoriasis. J. Clin. Invest. 1995, 96, 2106–2112. [Google Scholar] [CrossRef]
- Kolbinger, F.; Loesche., C.; Valentin, M.A. b-Defensin 2 is a responsive biomarker of IL-17A-driven skin pathology in patients with psoriasis. J. Allergy. Clin. Immunol. 2017, 139, 923–932. [Google Scholar] [CrossRef]
- Komine, M. Recent Advances in Psoriasis Research; the Clue to Mysterious Relation to Gut Microbiome. Int. J. Mol. Sci. 2020, 21, 2582. [Google Scholar] [CrossRef] [PubMed]
- Scher, J.U.; Ubeda, C.; Artacho, A.; Attur, M.; Isaac, S.; Reddy, S.M. Decreased bacterial diversity characterizes the altered gut microbiota in patients with psoriatic arthritis, resembling dysbiosis in inflammatory bowel disease. Arthritis. Rheumatol. 2015, 67, 128–139. [Google Scholar] [CrossRef]
- Alesa, D.I.; Alshamrani, H.M.; Alzahrani, Y.A.; Alamssi, D.N.; Alzahrani, N.S.; Almohammadi, M.E. The role of gut microbiome in the pathogenesis of psoriasis and the therapeutic effects of probiotics. J. Family Med. Prim. Care 2019, 8, 3496–3503. [Google Scholar] [CrossRef]
- Fry, L.; Baker, B.S. Triggering psoriasis: The role of infections and medications. Clin. Dermatol. 2007, 25, 606–615. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Ho, H.J.; Tseng, C.H.; Lai, Z.L.; Shieh, J.J.; Wu, C.Y. Intestinal microbiota profiling and predicted metabolic dysregulation in psoriasis patients. Exp. Dermatol. 2018, 27, 1336–1343. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, C.; Tan, J.; Macia, L.; Mackay, C.R. The nutrition-gut microbiome-physiology axis and allergic diseases. Immunol. Rev. 2017, 278, 277–295. [Google Scholar] [CrossRef]
- Mazidi, M.; Rezaie, P.; Kengne, A.P.; Mobarhan, M.G.; Ferns, G.A. Gut microbiome and metabolic syndrome. Diabetes Metab. Syndr. 2016, 10, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Santoru, M.L.; Piras, C.; Murgia, A.; Palmas, V.; Camboni, T.; Liggi, S.; Ibba, I.; Lai, M.A.; Orru, S.; Blois, S.; et al. Cross sectional evaluation of the gut-microbiome metabolome axis in an Italian cohort of IBD patients. Sci. Rep. 2017, 7, 9523. [Google Scholar] [CrossRef]
- Pascal, V.; Pozuelo, M.; Borruel, N.; Casellas, F.; Campos, D.; Santiago, A.; Martinez, X.; Varela, E.; Sarrabayrouse, G.; Machiels, K.; et al. A microbial signature for Crohn’s disease. Gut 2017, 66, 813–822. [Google Scholar] [CrossRef]
- Shapiro, J.; Cohen, N.A.; Shalev, V.; Uzan, A.; Koren, O.; Maharshak, N. Psoriatic patients have a distinct structural and functional fecal microbiota compared with controls. J. Dermatol. 2019, 46, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.; Zhao, S.; Zhu, W.; Wu, L.; Li, J.; Shen, M.; Lei, L.; Chen, X.; Peng, C. The Akkermansia muciniphila is a gut microbiota signature in psoriasis. Exp. Dermatol. 2018, 27, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Scher, J.U. The microbiome in psoriasis and psoriatic arthritis: Joints. J. Rheumatol. Suppl. 2018, 94, 32–35. [Google Scholar] [CrossRef] [PubMed]
- Sikora, M.; Stec, A.; Chrabaszcz, M.; Knot, A.; Waskiel-Burnat, A.; Rakowska, A.; Olszewska, M.; Rudnicka, L. Gut Microbiome in Psoriasis: An Updated Review. Pathogens 2020, 9, 463. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Zhang, M.; Wang, Y.; Dorfman, R.G.; Liu, H.; Yu, T.; Chen, X.; Tang, D.; Xu, L.; Yin, Y.; et al. Faecalibacterium prausnitzii produces butyrate to maintain Th17/treg balance and to ameliorate colorectal colitis by inhibiting histone deacetylase 1. Inflamm. Bowel. Dis. 2018, 24, 1926–1940. [Google Scholar] [CrossRef]
- Huang, X.L.; Zhang, X.; Fei, X.Y.; Chen, Z.G.; Hao, Y.P.; Zhang, S.; Zhang, M.M.; Yu, Y.Q.; Yu, C.G. Faecalibacterium prausnitzii supernatant ameliorates dextran sulfate sodium induced colitis by regulating Th17 cell differentiation. World J. Gastroenterol. 2016, 22, 5201–5210. [Google Scholar] [CrossRef]
- Stefia, L.V.; Lee, J.; Patel, J.; de Sousa, S.R.; Legrand, J.; Rodero, M.; Burman, S.; Linedale, R.; Morrison, M.; Khosrotehrani, K. Secretome components from Faecalibacterium prausnitzii strains A2-165 and AHMP21 Modulate Cutaneous Wound Inflammation. J. Investig. Dermatol. 2020, 140, 2312–2315.e6. [Google Scholar] [CrossRef] [PubMed]
- Anbazhagan, A.N.; Priyamvada, S.; Gujral, T.; Bhattacharyya, S.; Alrefai, W.A.; Dudeja, P.K. A novel anti-inflammatory role of GPR120 in intestinal epithelial cells. Am. J. Physiol. Cell Physiol. 2016, 310, C612–C621. [Google Scholar] [CrossRef]
- Masallat, D.; Moemen, D. State A. Gut bacterial microbiota in psoriasis: A case control study. Afr. J. Microbiol. Res. 2016, 7, 1337–1343. [Google Scholar] [CrossRef]
- Eppinga, H.; Sperna Weiland, C.J.; Thio, H.B.; van der Woude, C.J.; Nijsten, T.E.C.; Peppelenbosch, M.P.; Konstantinov, S.R. Similar depletion of protective Faecalibacterium prausnitzii in psoriasis and inflammatory bowel disease, but not in hidradenitis suppurativa. J. Crohns Colitis 2016, 10, 1067–1075. [Google Scholar] [CrossRef]
- Hidalgo-Cantabrana, C.; Gomez, J.; Delgado, S. Gut microbiota dysbiosis in a cohort of patients with psoriasis. Br. J. Dermatol. 2019, 181, 1287–1295. [Google Scholar] [CrossRef]
- Zhang, X.; Shi, L.; Sun, T.; Guo, K.; Geng, S. Dysbiosis of gut microbiota and its correlation with dysregulation of cytokines in psoriasis patients. BMC Microbiol. 2021, 21, 78. [Google Scholar] [CrossRef] [PubMed]
- Valentini, V.; Silvestri, V.; Marraffa, F.; Greco, G.; Bucalo, A.; Grassi, S.; Gagliardi, A.; Mazzotta, A.; Ottini, L.; Richetta, A.G. Gut microbiome profile in psoriatic patients treated and untreated with biologic therapy. J. Dermatol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Okada, K.; Matsushima, Y.; Mizutani, K.; Yamanaka, K. The Role of Gut Microbiome in Psoriasis: Oral Administration of Staphylococcus aureus and Streptococcus danieliae Exacerbates Skin Inflammation of Imiquimod-Induced Psoriasis-Like Dermatitis. Int. J. Mol. Sci. 2020, 21, 3303. [Google Scholar] [CrossRef]
- Sikora, M.; Chrabaszcz, M.; Maciejewski, C.; Zaremba, M.; Waskiel, A.; Olszewska, M. Intestinal barrier integrity in patients with plaque psoriasis. J. Dermatol. 2018, 45, 1468–1470. [Google Scholar] [CrossRef] [PubMed]
- Sikora, M.; Stec, A.; Chrabaszcz, M.; Waskiel-Burnat, A.; Zaremba, M.; Olszewska, M. Intestinal fatty acid binding protein, a biomarker of intestinal barrier, is associated with severity of psoriasis. J. Clin. Med. 2019, 8, 1021. [Google Scholar] [CrossRef]
- Ramírez-Boscá, A.; Navarro-López, V.; Martínez-Andrés, A.; Such, J.; Francés, R.; Horga de la Parte, J.; Asín-Llorca, M. Identification of Bacterial DNA in the Peripheral Blood of Patients with Active Psoriasis. JAMA Derm. 2015, 151, 670–671. [Google Scholar] [CrossRef]
- Fu, Y.; Lee, C.H.; Chi, C.C. Association of psoriasis with inflammatory bowel disease: A systematic review and meta-analysis. JAMA Dermatol. 2018, 154, 1417–1423. [Google Scholar] [CrossRef]
- Ely, P.H. Is psoriasis a bowel disease? Successful treatment with bile acids and bioflavonoids suggests it is. Clin. Dermatol. 2018, 36, 376–389. [Google Scholar] [CrossRef]
- Sitkin, S.; Pokrotnieks, J. Clinical potential of anti-inflammatory effects of Faecalibacterium prausnitzii and butyrate in inflammatory bowel disease. Inflamm. Bowel. Dis. 2019, 25, 40–41. [Google Scholar] [CrossRef]
- Bian, X.; Wu, W.; Yang, L.; Lv, L.; Wang, Q.; Li, Y.; Ye, J.; Fang, D.; Wu, J.; Jiang, X.; et al. Administration of Akkermansia muciniphila ameliorates dextran sulfate sodiuminduced ulcerative colitis in mice. Front. Microbiol. 2019, 10, 2259. [Google Scholar] [CrossRef]
- Smith, P.M.; Howitt, M.R.; Panikov, N.; Michaud, M.; Gallini, C.A.; Bohlooly, Y.M.; Glickman, J.N.; Garrett, W.S. The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef]
- Vinolo, M.A.R.; Rodrigues, H.G.; Nachbar, R.T.; Curi, R. Regulation of inflammation by short chain fatty acids. Nutrients 2011, 3, 858–876. [Google Scholar] [CrossRef] [PubMed]
- Plöger, S.; Stumpff, F.; Penner, G.B.; Schulzke, J.D.; Gäbel, G.; Martens, H.; Shen, Z.; Günzel, D.; Aschenbach, J.R. Microbial butyrate, and its role for barrier function in the gastrointestinal tract. Ann. N. Y. Acad. Sci. 2012, 1258, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Meijer, K.; de Vos, P.; Priebe, M.G. Butyrate and other short-chain fatty acids as modulators of immunity: What relevance for health? Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 715–721. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, A.; Bruhs, A.; Schwarz, T. The short-chain fatty acid sodium butyrate functions as a regulator of the skin immune system. J. Invest. Dermatol. 2017, 137, 855–864. [Google Scholar] [CrossRef]
- Codoner, F.M.; Ramírez-Bosca, A.; Climent, E. Gut microbial composition in patients with psoriasis. Sci. Rep. 2018, 8, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, S.C.; Barbara, G.; Buurman, W. Intestinal permeability e a new target for disease prevention and therapy. BMC Gastroenterol. 2014, 18, 189. [Google Scholar] [CrossRef]
- De Preter, V.; Machiels, K.; Joossens, M.; Arijs, I.; Matthys, C.; Vermeire, S.; Rutgeerts, P.; Verbeke, K. Faecal metabolite profiling identifies medium-chain fatty acids as discriminating compounds in IBD. Gut 2015, 64, 447–458. [Google Scholar] [CrossRef]
- Liberato, M.V.; Nascimento, A.S.; Ayers, S.D.; Lin, J.Z.; Cvoro, A.; Silveira, R.L.; Martínez, L.; Souza, P.C.; Saidemberg, D.; Deng, T.; et al. Medium chain fatty acids are selective peroxisome proliferator activated receptor (PPAR) γ activators and pan-PPAR partial agonists. PLoS ONE 2012, 7, e36297. [Google Scholar] [CrossRef]
- Bassaganya-Riera, J.; Hontecillas, R.; Horne, W.T.; Sandridge, M.; Herfarth, H.H.; Bloomfeld, R.; Isaacs, K.L. Conjugated linoleic acid modulates immune responses in patients with mild to moderately active Crohn’s disease. Clin. Nutr. 2012, 31, 721–727. [Google Scholar] [CrossRef]
- Haghikia, A.; Jörg, S.; Duscha, A.; Berg, J.; Manzel, A.; Waschbisch, A.; Hammer, A.; Lee, D.H.; May, C.; Wilck, N.; et al. Dietary Fatty Acids Directly Impact Central Nervous System Autoimmunity via the Small Intestine. Immunity 2015, 43, 817–829. [Google Scholar] [CrossRef]
- Bhutia, Y.D.; Ganapathy, V. Short, but smart: SCFAs train T cells in the gut to fight autoimmunity in the brain. Immunity 2015, 43, 629–631. [Google Scholar] [CrossRef]
- Zelante, T.; Iannitti, R.G.; Cunha, C.; De Luca, A.; Giovannini, G.; Pieraccini, G. Tryptophan catabolites from microbiota engage aryl hydrocarbon receptor and balance mucosal reactivity via interleukin-22. Immunity 2013, 39, 372–385. [Google Scholar] [CrossRef]
- Rothhammer, V.; Mascanfroni, I.D.; Bunse, L.; Takenaka, M.C.; Kenison, J.E.; Mayo, L. Type I interferons and microbial metabolites of tryptophan modulate astrocyte activity and central nervous system inflammation via the aryl hydrocarbon receptor. Nat. Med. 2016, 22, 586–597. [Google Scholar] [CrossRef]
- Zhai, R.; Xue, X.; Zhang, L.; Yang, X.; Zhao, L.; Zhang, C. Strain-Specific Anti-inflammatory Properties of Two Akkermansia muciniphila Strains on Chronic Colitis in Mice. Front. Cell. Infect. Microbiol. 2019, 9, 239. [Google Scholar] [CrossRef] [PubMed]
- Giacinto, C.D.; Marinaro, M.; Sanchez, M. Probiotics ameliorate recurrent Th1-mediated murine colitis by inducing IL-10 and IL-10-dependent TGF-b-bearing regulatory cells. J. Immunol. 2005, 174, 3237–3246. [Google Scholar] [CrossRef] [PubMed]
- Groeger, D.; O’Mahony, L.; Murphy, E.F. Bifidobacterium infantis 35624 modulates host inflammatory processes beyond the gut. Gut Microb. 2013, 4, 325–339. [Google Scholar] [CrossRef] [PubMed]
- Wedrychowicz, A.; Tomasik, P.; Zajac, A.; Fyderek, K. Prognostic value of assessment of stool and serum IL-1beta, IL-1ra and IL 6 concentrations in children with active and inactive ulcerative colitis. Arch. Med. Sci. 2018, 14, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Enocksson, A.; Lundberg, J.; Weitzberg, E.; Norrby-Teglund, A.; Svenungsson, B. Rectal nitric oxide gas and stool cytokine levels during the course of infectious gastroenteritis. Clin. Diagnos. Lab. Immunol. 2004, 11, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Natsuaki, Y.; Egawa, G.; Nakamizo, S.; Ono, S.; Hanakawa, S.; Okada, T. Perivascular leukocyte clusters are essential for efficient activation of effector T cells in the skin. Nat. Immunol. 2014, 15, 1064–1069. [Google Scholar] [CrossRef]
- Kryczek, I.; Bruce, A.T.; Gudjonsson, J.E.; Johnston, A.; Aphale, A.; Vatan, L. Induction of IL-17+ T cell trafficking and development by IFN-gamma: Mechanism and pathological relevance in psoriasis. J. Immunol. 2008, 181, 4733–4741. [Google Scholar] [CrossRef] [PubMed]
- Baliwag, J.; Barnes, D.H.; Johnston, A. Cytokines in psoriasis. Cytokine 2015, 73, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Di Paolo, N.C.; Shayakhmetov, D.M. Interleukin 1alpha and the inflammatory process. Nat. Immunol. 2016, 17, 906–913. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, C.A.; Monteleone, G.; McLaughlin, J.T.; Paus, R. The gut-skin axis in health and disease: A paradigm with therapeutic implications. Bioessays 2016, 38, 1167–1176. [Google Scholar] [CrossRef]
- Cho, C.E.; Taesuwan, S.; Malysheva, O.V. Trimethylamine-N-oxide (TMAO) response to animal source foods varies among healthy young men and is influenced by their gut microbiota composition: A randomized controlled trial. Mol. Nutr. Food Res. 2017, 61, 1600324. [Google Scholar] [CrossRef]
- Qin, J.; Li, Y.; Cai, Z. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012, 490, 55–60. [Google Scholar] [CrossRef]
- Benhadou, F.; Mintoff, D.; Schnebert, B.; Thio, H.B. Psoriasis and Microbiota: A Systematic Review. Diseases 2018, 6, 47. [Google Scholar] [CrossRef]
- Wang, Z.; Klipfell, E.; Bennett, B.J. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef]
- Koeth, R.A.; Wang, Z.; Levison, B.S. Intestinal microbiota metabolism of L-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat. Med. 2013, 19, 576–585. [Google Scholar] [CrossRef]
- Komaroff, A.L. The microbiome and risk for obesity and diabetes. J. Am. Med. Assoc. 2017, 317, 355–356. [Google Scholar] [CrossRef]
- Perry, R.J.; Peng, L.; Barry, N.A. Acetate mediates a microbiome-brain-b cell axis promoting metabolic syndrome. Nature 2016, 534, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Yin, J.; Zhang, J. Butyrate improves insulin sensitivity and increases energy expenditure in mice. Diabetes 2009, 58, 1509–1517. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Backhed, F.; Turnbaugh, P. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef]
- Rosenbaum, M.; Knight, R.; Leibel, R.L. The gut microbiota in human energy homeostasis and obesity. Trends Endocrinol. Metab. TEM 2015, 26, 93–501. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Cerdeira, C.; Cordeiro-Rodríguez, M.; Carnero-Gregorio, M.; López-Barcenas, A.; Martínez-Herrera, E.; Fabbrocini, G.; Sinani, A.; Arenas-Guzmán, R.; González-Cespón, J.L. Biomarkers of Inflammation in Obesity-Psoriatic Patients. Mediators Inflamm. 2019, 7353420. [Google Scholar] [CrossRef]
- Abuabara, K.; Azfar, R.S.; Shin, D.B. Cause-specific mortality in patients with severe psoriasis: A population-based cohort study in the United Kingdom. Br. J. Dermatol. 2010, 163, 586–592. [Google Scholar] [CrossRef]
- Gilis, E.; Mortier, C.; Venken, K.; Debusschere, K.; Vereecke, L.; Elewaut, D. The Role of the Microbiome in Gut and Joint Inflammation in Psoriatic Arthritis and Spondyloarthritis. J. Rheumatol. Suppl. 2018, 94, 36–39. [Google Scholar] [CrossRef]
- Van Praet, L.; Jacques, P.; Van den Bosch, F.; Elewaut, D. The transition of acute to chronic bowel inflammation in spondyloarthritis. Nat. Rev. Rheumatol. 2012, 85, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Benham, H.; Rehaume, L.M.; Hasnain, S.Z.; Velasco, J.; Baillet, A.C.; Ruutu, M. Interleukin-23 mediates the intestinal response to microbial β-1,3-glucan, and the development of spondyloarthritis pathology in SKG mice. Arthritis. Rheumatol. 2014, 66, 1755–1767. [Google Scholar] [CrossRef]
- Rehaume, L.M.; Mondot, S.; Aguirre de Carcer, D.; Velasco, J.; Benham, H.; Hasnain, S.Z. ZAP-70 genotype disrupts the relationship between microbiota and host, leading to spondyloarthritis and ileitis in SKG mice. Arthritis. Rheumatol. 2014, 66, 2780–2792. [Google Scholar] [CrossRef]
- Ruutu, M.; Thomas, G.; Steck, R.; Degli-Esposti, M.A.; Zinkernagel, M.S.; Alexander, K. β-glucan triggers spondylarthritis and Crohn’s disease-like ileitis in SKG mice. Arthritis. Rheum. 2012, 64, 2211–2222. [Google Scholar] [CrossRef]
- Jacques, P.; Venken, K.; Van Beneden, K.; Hammad, H.; Seeuws, S.; Drennan, M.B. Invariant natural killer T cells are natural regulators of murine spondylarthritis. Arthritis. Rheum. 2010, 62, 988–999. [Google Scholar] [CrossRef]
- Schaubeck, M.; Clavel, T.; Calasan, J.; Lagkouvardos, I.; Haange, S.B.; Jehmlich, N. Dysbiotic gut microbiota causes transmissible Crohn’s disease-like ileitis independent of failure in antimicrobial defence. Gut 2016, 65, 225–237. [Google Scholar] [CrossRef]
- Honda, K.; Littman, D.R. The microbiome in infectious disease and inflammation. Annu. Rev. Immunol. 2012, 30, 759–795. [Google Scholar] [CrossRef] [PubMed]
- Salio, M.; Silk, J.D.; Jones, E.Y.; Cerundolo, V. Biology of CD1- and MR1-restricted T cells. Annu. Rev. Immunol. 2014, 32, 323–366. [Google Scholar] [CrossRef] [PubMed]
- Eken, A.; Singh, A.K.; Oukka, M. Interleukin 23 in Crohn’s disease. Inflamm. Bowel. Dis. 2014, 20, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Van de Wiele, T.; Van Praet, J.T.; Marzorati, M.; Drennan, M.B.; Elewaut, D. How the microbiota shapes rheumatic diseases. Nat. Rev. Rheumatol. 2016, 12, 398–411. [Google Scholar] [CrossRef]
- Van Praet, J.T.; Donovan, E.; Vanassche, I.; Drennan, M.B.; Windels, F.; Dendooven, A. Commensal microbiota influence systemic autoimmune responses. EMBO J. 2015, 34, 466–474. [Google Scholar] [CrossRef] [PubMed]
- Zanvit, P.; Konkel, J.E.; Jiao, X.; Kasagi, S.; Zhang, D.; Wu, R. Antibiotics in neonatal life increase murine susceptibility to experimental psoriasis. Nat. Commun. 2015, 6, 8424. [Google Scholar] [CrossRef]
- Montoya, J.; Matta, N.B.; Suchon, P.; Guzian, M.C.; Lambert, N.C.; Mattei, J.P. Patients with ankylosing spondylitis have been breast fed less often than healthy controls: A case-control retrospective study. Ann. Rheum. Dis. 2016, 75, 879–882. [Google Scholar] [CrossRef]
- Asquith, M.; Davin, S.; Stauffer, P.; Michell, C.; Janowitz, C.; Lin, P. Intestinal metabolites are profoundly altered in the context of HLA-B27 expression and functionally modulate disease in a rat model of spondyloarthritis. Arthritis Rheumatol. 2017, 69, 1984–1995. [Google Scholar] [CrossRef]
- Peric, M.; Koglin, S.; Dombrowski, Y.; Groß, K.; Bradac, E.; Büchau, A.; Steinmeyer, A.; Zügel, U.; Ruzicka, T.; Schauber, J. Vitamin D analogs differentially control antimicrobial peptide/ “Alarmin” expression in psoriasis. PLoS ONE 2009, 4, e6340. [Google Scholar] [CrossRef] [PubMed]
- López-García, B.; Lee, P.H.; Gallo, R.L. Expression, and potential function of cathelicidin antimicrobial peptides in dermatophytosis and tinea versicolor. J. Antimicrob. Chemother. 2006, 57, 877–882. [Google Scholar] [CrossRef] [PubMed]
- Assarsson, M.; Duvetorp, A.; Dienus, O.; Söderman, J.; Seifert, O. Significant Changes in the Skin Microbiome in Patients with Chronic Plaque Psoriasis after Treatment with Narrowband Ultraviolet B. Acta Dermato. Venereol. 2018, 98, 428–436. [Google Scholar] [CrossRef] [PubMed]
- Darlenski, R.; Hristakieva, E.; Aydin, U.; Gancheva, D.; Gancheva, T.; Zheleva, A.; Gadjeva, V.; Fluhr, J.W. Epidermal barrier and oxidative stress parameters improve during in 311 nm narrow band UVB phototherapy of plaque type psoriasis. J. Dermatol. Sci. 2018, 91, 28–34. [Google Scholar] [CrossRef]
- Bosman, E.S.; Albert, A.Y.; Lui, H.; Dutz, J.P.; Vallance, B.A. Skin Exposure to Narrow, Band Ultraviolet (UVB) Light Modulates the Human Intestinal Microbiome. Front. Microbiol. 2019, 10, 2410. [Google Scholar] [CrossRef] [PubMed]
- Langan, E.A.; Künstner, A.; Miodovnik, M.; Zillikens, D.; Thaçi, D.; Baines, J.F.; Ibrahim, S.M.; Solbach, W.; Knobloch, J.K. Combined culture and metagenomic analyses reveal significant shifts in the composition of the cutaneous microbiome in psoriasis. Br. J. Dermatol. 2019, 181, 1254–1264. [Google Scholar] [CrossRef]
- Yeh, N.L.; Hsu, C.Y.; Tsai, T.F.; Chiu, H.Y. Gut Microbiome in Psoriasis is Perturbed Differently During Secukinumab and Ustekinumab Therapy and Associated with Response to Treatment. Clin. Drug Investig. 2019, 39, 1195–1203. [Google Scholar] [CrossRef]
- Nermes, M.; Kantele, J.M.; Atosuo, T.J.; Salminen, S.; Isolauri, E. Interaction of orally administered Lactobacillus rhamnosus GG with skin and gut microbiota and humoral immunity in infants with atopic dermatitis. Clin. Exp. Allergy 2011, 41, 370–377. [Google Scholar] [CrossRef]
- Vijayashankar, M.; Raghunath, N. Pustular psoriasis responding to probiotics–A new insight. Our Dermatol. Online 2012, 3, 326. [Google Scholar] [CrossRef]
- Chen, Y.H.; Wu, C.S.; Chao, Y.H.; Lin, C.C.; Tsai, H.Y.; Li, Y.R.; Chen, Y.Z.; Tsai, W.H.; Chen, Y.K. Lactobacillus pentosus GMNL-77 inhibits skin lesions in imiquimod-induced psoriasis-like mice. J. Food Drug Anal. 2017, 25, 559–566. [Google Scholar] [CrossRef]
- Suarez-Farinas, M.; Li, K.; Fuentes-Duculan, J.; Hayden, K.; Brodmerkel, C.; Krueger, J.G. Expanding the psoriasis disease profile: Interrogation of the skin and serum of patients with moderate-to-severe psoriasis. J. Invest. Dermatol. 2012, 132, 2552–2564. [Google Scholar] [CrossRef]
- Aroniadis, O.C.; Brandt, L.J. Fecal microbiota transplantation: Past, present, and future. Curr. Opin. Gastroenterol. 2013, 29, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Glassner, K.L.; Abraham, B.P.; Quigley, E.M.M. The microbiome and inflammatory bowel disease. J. Allergy Clin. Immunol. 2020, 145, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Kelly, C.R.; Ihunnah, C.; Fischer, M.; Khoruts, A.; Surawicz, C.; Afzali, A.; Aroniadis, O.; Barto, A.; Borody, T.; Giovanelli, A.; et al. Fecal microbiota transplant for treatment of Clostridium difficile infection in immunocompromised patients. Am. J. Gastroenterol. 2014, 109, 1065–1071. [Google Scholar] [CrossRef]
- Michaelsson, G.; Gerden, B.; Ottosson, M.; Parra, A.; Sjöberg, O.; Hjelmquist, G.; Lööf, L. Patients with psoriasis often have increased serum levels of IgA antibodies to gliadin. Br. J. Dermatol. 1993, 129, 667–673. [Google Scholar] [CrossRef]
- Michaëlsson, G.; Gerden, B.; Hagforsen, E.; Nilsson, B.; Pihl-Lundin, I.; Kraaz, W.; Hjelmquist, G.; Lööf, L. Psoriasis patients with antibodies to gliadin can be improved by a gluten-free diet. Br. J. Dermatol. 2000, 142, 44–51. [Google Scholar] [CrossRef]
- Michaëlsson, G.; Åhs, S.; Hammarström, I.; Lundin, I.P.; Hagforsen, E. Gluten-free diet in psoriasis patients with antibodies to gliadin results in decreased expression of tissue transglutaminase and fewer Ki67+ cells in the dermis. Acta Dermato. Venereol. 2003, 83, 425–429. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Author | Study Group | Analysed Sample | Method of Analysis | Results |
|---|---|---|---|---|
| Fahlen et al., 2012 [17] | Psoriasis patients (n = 10) Healthy controls (n = 12) | Skin biopsy | 16S rRNA sequesting (V3–V4 hypervariable region) | ↑ Proteobacteria phylum ↓ Firmicutes and Actinobacteria phyla ↓ Streptococci and Cutibacterium genera ↑ Staphylococci genera |
| Alekseyenko et al., 2013 [11] | Psoriasis patients (n = 54) Healthy controls (n = 37) | Skin swab | 16S rRNA sequesting (V1–V3 hypervariable region) | ↓ Proteobacteria phylum ↑ Actinobacteria and Firmicutes phyla |
| Chang et al., 2018 [30] | Psoriasis patients (n = 28) Healthy controls (n = 26) | Skin swab | 16S rRNA sequesting (V1–V3 hypervariable region) | ↓ Actinobacteria phylum ↑ Proteobacteria phylum ↓ Cutibacterium, Ethanoligenens and Macrococcus genera ↑ Pseudomonas genera ↓ Cutibacterium acnes, Cutibacterium granulosum, Staphylococcus epidermidis ↑ Staphylococcus aureus and Staphylococcus pettenkoferi |
| Fyhrquist et al., 2019 [12] | Atopic dermatitis patients (n = 82) Psoriasis patients (n = 119) Healthy controls (n = 115) | Skin swab | 16S rRNA sequesting (V1-V4 hypervariable region) | ↑ Finegoldia, Neisseriaceae, Corynebacterium kroppenstedtii, Corynebacterium simulans ↓ Lactobacilli, Burkholderia spp., Cutibacterium acnes |
| Assarsson et al., 2020 [31] | Psoriasis patients (n = 50) Healthy controls (n = 77) | Skin swab | 16S rRNA sequesting (V3-V4 hypervariable region) | ↑ Firmicutes and Protebacteria phyla ↓ Fusobacteria and Cyanobacteria phyla |
| Quan et al., 2020 [13] | Psoriasis patients (n = 27) Healthy controls (n = 19) | Skin swab | 16S rRNA sequesting (V3–V4 hypervariable region) | ↓ Deinococcus and Thermus phyla ↑ Corynebacterium genera ↓ Cutibacterium genera |
| Author | Study Group | Analysed Sample | Method of Analysis | Results |
|---|---|---|---|---|
| Scher et al., 2015 [42] | Psoriasis patients (n = 15) Psoriatic arthritis patients (n = 16) Healthy controls (n = 17) | Fecal sample | 16s rRNA sequencing (V1 -V2 hypervariable region) | ↓ diversity in psoriasis and psoriatic arthritis ↓ Bacteroidetes phylum in psoriasis versus psoriatic arthritis ↓ Akkermansia, Alistipes, Parabacteroides, Pseudobutyrivibrio, Ruminococcus and Coprococcus genera in psoriatic arthritis and psoriasis ↓ Coprobacillus genera in psoriasis when compared to psoriatic arthritis |
| Masallat et al., 2016 [58] | Psoriasis patients (n = 45) Healthy controls (n = 45) | Fecal sample | Real-time PCR | ↓ Actinobacteria phylum ↑ Bifidobacterium Collinsella, Dorea Ruminococcus, Slackia and Subdoligranulum genera ↑ F/B ratio |
| Eppinga et al., 2016 [59] | Psoriasis patients (n = 29) IBD patients (n = 31) HS patients (n = 17)-Psoriasis and IBD patients (n = 13) HS and IBD patients (n = 17) Healthy controls (n = 33) | Fecal sample | Quantitative PCR | ↓ F. prausnitzii ↑ E. coli in patients with psoriasis and concomitant psoriasis and IBD |
| Tan et al., 2018 [51] | Psoriasis patients (n = 14) Healthy controls (n = 14) | Fecal sample | 16s rDNA sequencing (V4 hypervariable region) | ↓ Akkermansia muciniphila, Verrucomicrobia and Tenericutes phyla Mollicutes and Verrucomicrobiae ↑ Bacteroides genera, Clostridium citroniae spp. and Enterococcus genera |
| Chen et al., 2018 [45] | Psoriasis patients (n = 32) Non-psoriasis controls (n = 64) | Fecal sample | 16s rRNA sequencing (V3 - V4 hypervariable region) | ↑ Firmicutes ↓ Bacteroidetes ↑ F/B ratio ↓ Akkermansia genus |
| Hidalgo -Cantabrana et al., 2019 [60] | Psoriasis patients (n = 19) Healthy controls (n = 20) | Fecal sample | 16s rRNA sequencing (V2 -V3 hypervariable region) | ↓ diversity ↑ Firmicutes ↓ Bacteroidetes ↑ F/B ratio ↑ Actinobacteria ↓ Proteobacteria phylum, Alistipes, Bacteroides, Barnesiella, Faecalibacterium, Parabacteroides and Paraprevotella genera |
| Shapiro et al., 2019 [50] | Psoriasis patients (n = 24) Non-psoriasis controls (n = 24) | Fecal sample | 16s rRNA sequencing (V4 hypervariable region) | ↑ Firmicutes ↓ Bacteroidetes ↑ F/B ratio ↑ Actinobacteria phylum, Blautia and Faecalibacterium genera ↓ Proteobacteria phylum, Prevotella genarum, Ruminococcus gnavus, Dorea formicigenerans and Collinsella aerofaciens spp. |
| Zhang et al., 2021 [61] | Psoriasis patients (n = 30) Healthy controls (n = 30) | Fecal sample | 16s rRNA sequencing | ↑ Faecalibacterium and Megamonas taxa |
| Valentini et al., 2021 [62] | Psoriasis patients treated with biologic therapy (n = 10) Psoriasis patients not treated with biologic therapy (n = 20) | Fecal sample | 16s rRNA sequencing | ↓ diversity of biologically treated patients vs. untreated patients |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olejniczak-Staruch, I.; Ciążyńska, M.; Sobolewska-Sztychny, D.; Narbutt, J.; Skibińska, M.; Lesiak, A. Alterations of the Skin and Gut Microbiome in Psoriasis and Psoriatic Arthritis. Int. J. Mol. Sci. 2021, 22, 3998. https://doi.org/10.3390/ijms22083998
Olejniczak-Staruch I, Ciążyńska M, Sobolewska-Sztychny D, Narbutt J, Skibińska M, Lesiak A. Alterations of the Skin and Gut Microbiome in Psoriasis and Psoriatic Arthritis. International Journal of Molecular Sciences. 2021; 22(8):3998. https://doi.org/10.3390/ijms22083998
Chicago/Turabian StyleOlejniczak-Staruch, Irmina, Magdalena Ciążyńska, Dorota Sobolewska-Sztychny, Joanna Narbutt, Małgorzata Skibińska, and Aleksandra Lesiak. 2021. "Alterations of the Skin and Gut Microbiome in Psoriasis and Psoriatic Arthritis" International Journal of Molecular Sciences 22, no. 8: 3998. https://doi.org/10.3390/ijms22083998
APA StyleOlejniczak-Staruch, I., Ciążyńska, M., Sobolewska-Sztychny, D., Narbutt, J., Skibińska, M., & Lesiak, A. (2021). Alterations of the Skin and Gut Microbiome in Psoriasis and Psoriatic Arthritis. International Journal of Molecular Sciences, 22(8), 3998. https://doi.org/10.3390/ijms22083998