
Periostin Is Required for the Maintenance of Muscle Fibers during Muscle Regeneration



Abstract
1. Introduction
2. Results
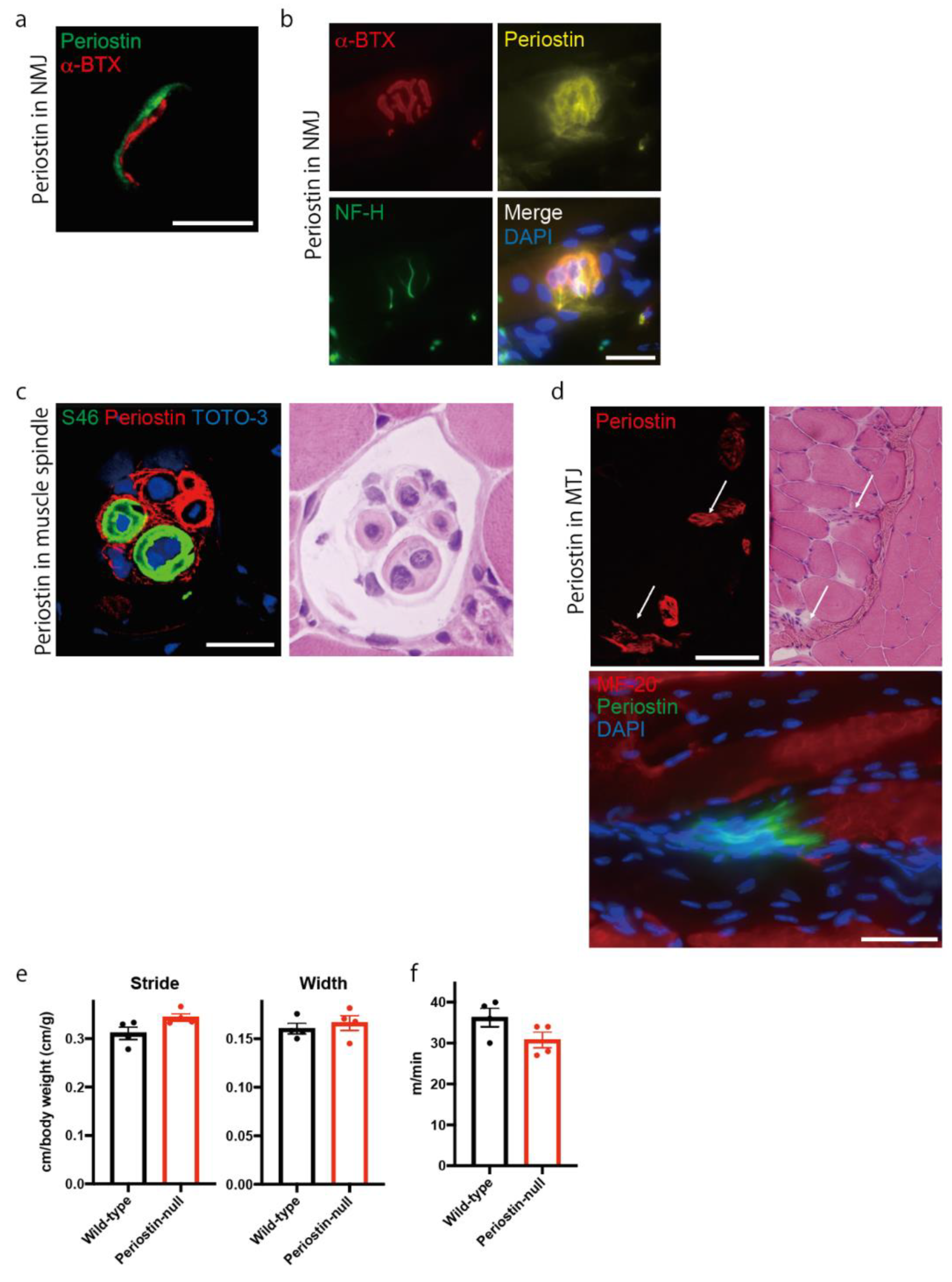
2.1. Periostin Is Localized at the Neuromuscular Junction, Muscle Spindle, and Myotendinous Junction in Skeletal Muscle
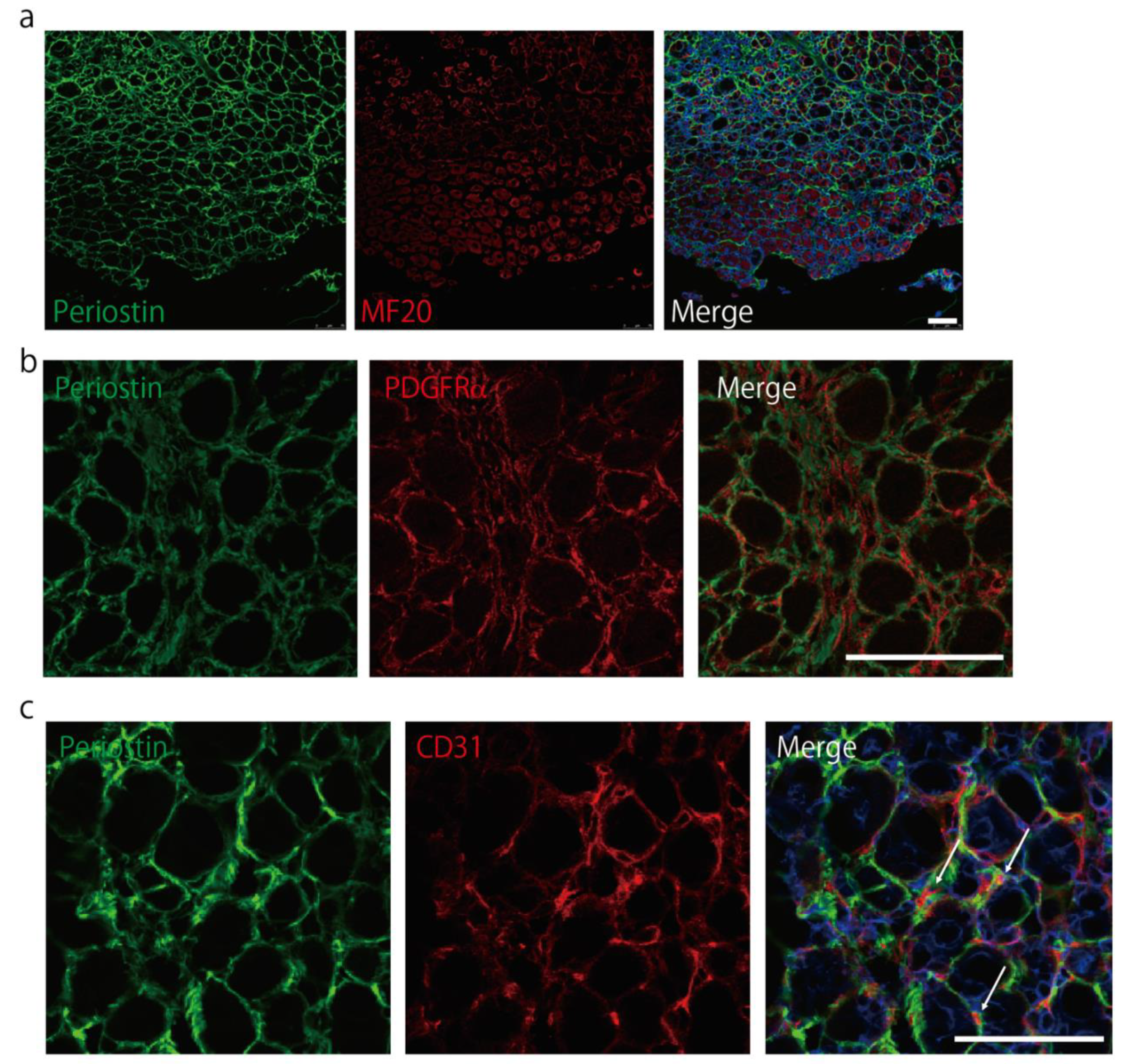
2.2. Periostin Is Expressed in the Interstitial Space during Muscle Regeneration
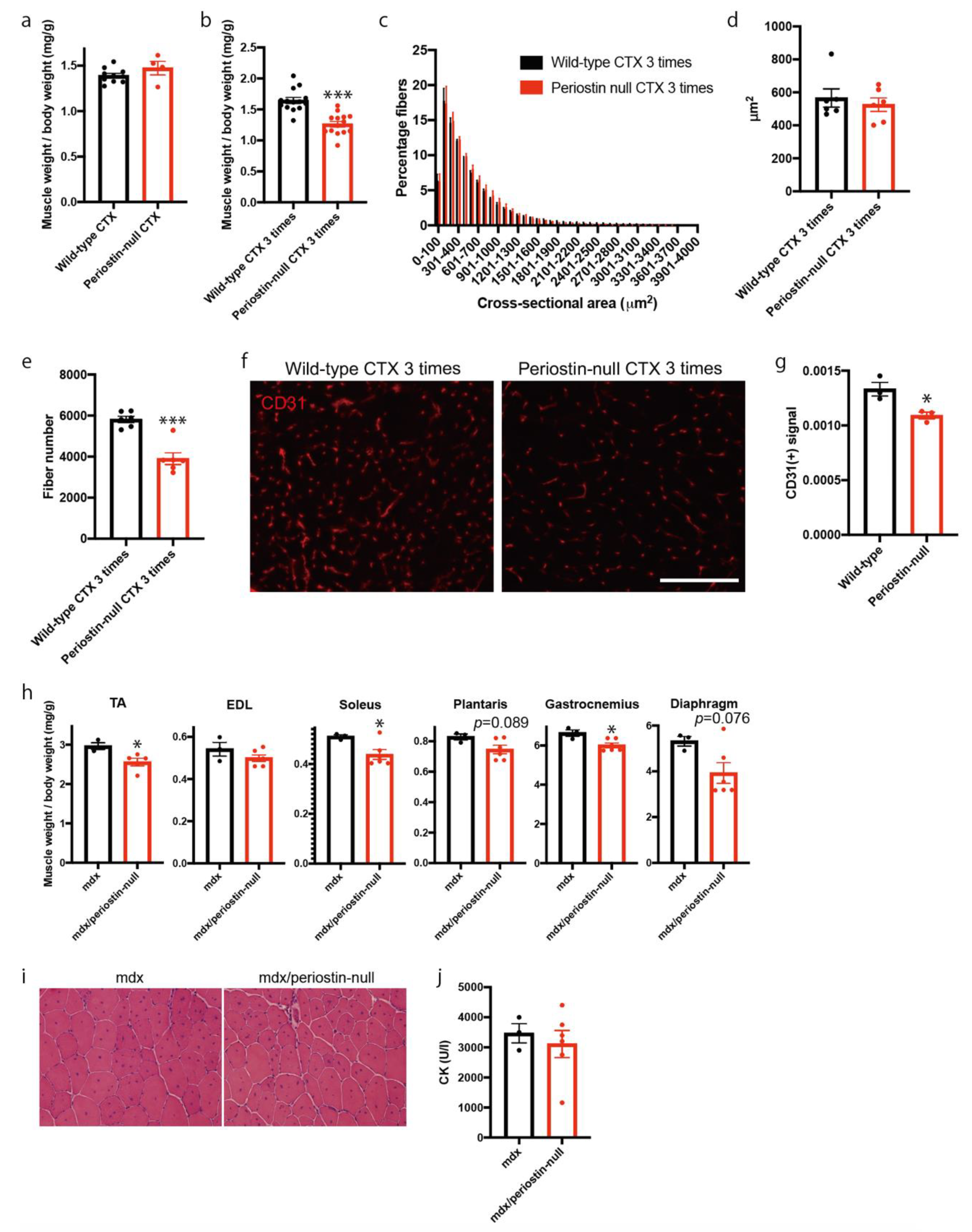
2.3. Loss of Periostin Caused a Decrease in Muscle Fibers during Repeated Muscle Regeneration
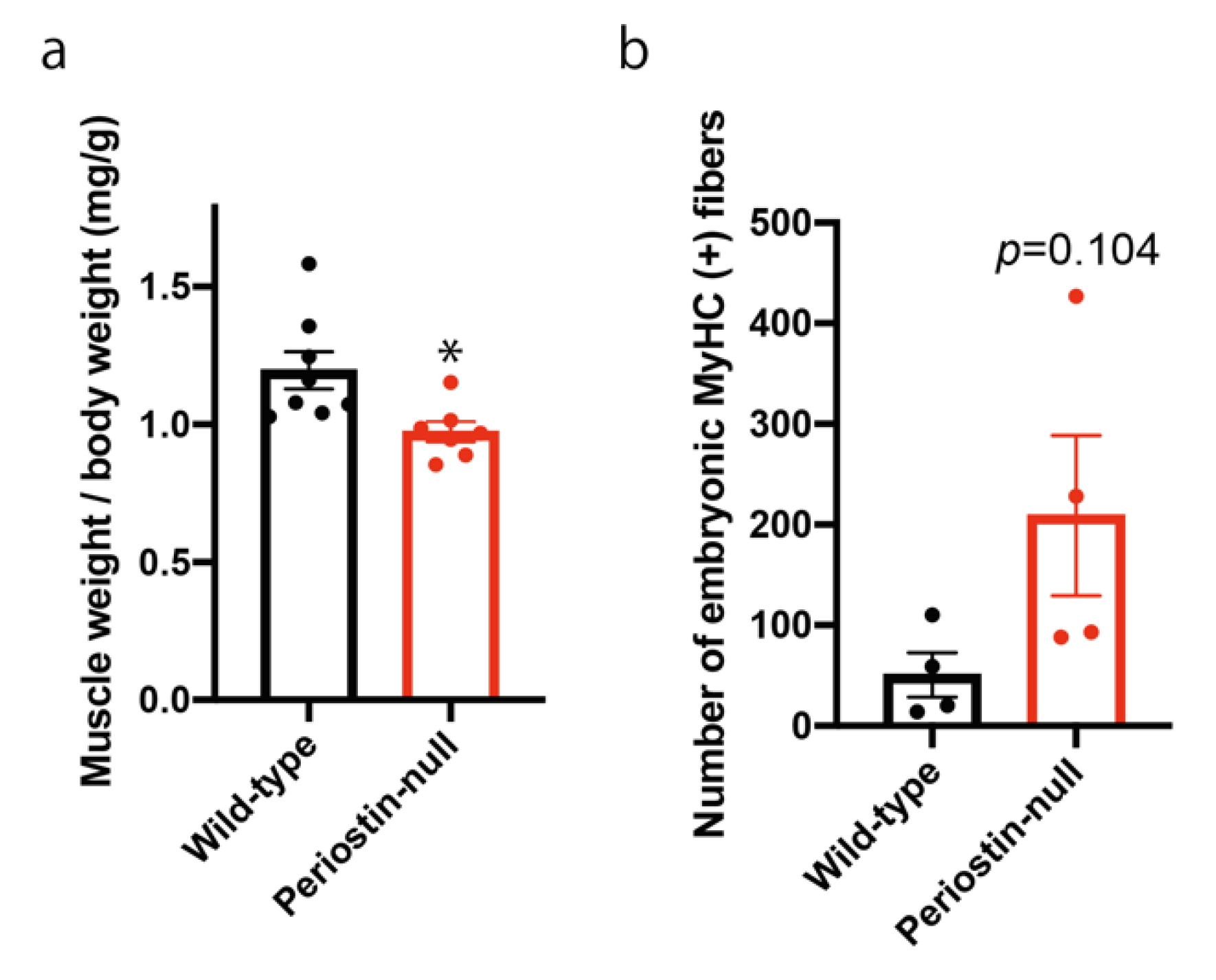
2.4. Loss of Periostin in Mice with a DBA/2 Genetic Background Delayed Muscle Regeneration
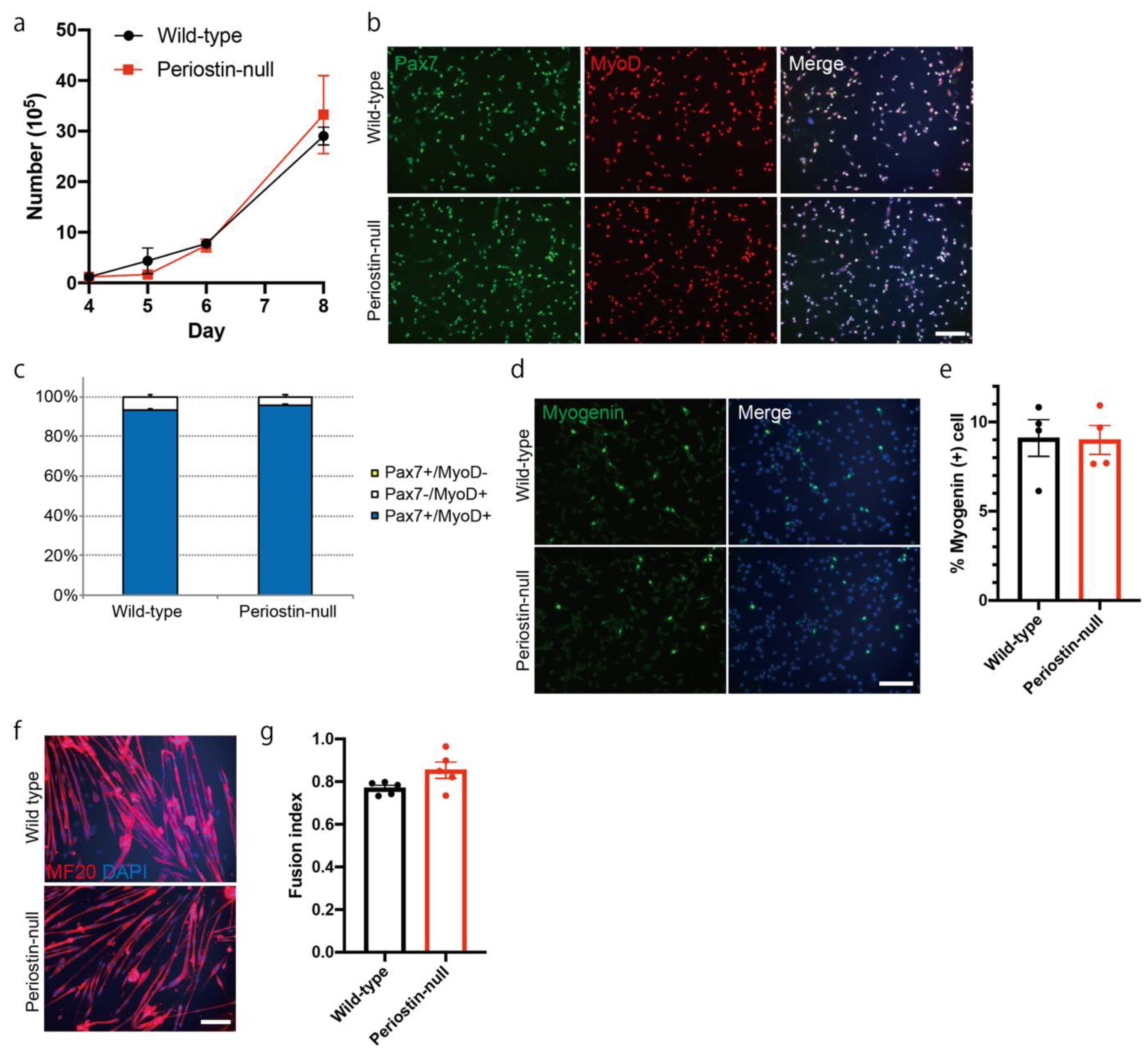
2.5. The Function of Satellite Cells was Not Impaired in Periostin-Null Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Exercise Model
4.3. Materials
4.4. RNA Isolation and Reverse Transcription–Polymerase Chain Reaction Analysis (RT-PCR)
4.5. Histological and Immunohistochemical Analysis, and Immunocytochemistry
4.6. Single Muscle Fiber Preparation and Cell Culture
4.7. Induction of Muscle Regeneration
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Segawa, M.; Fukada, S.-I.; Yamamoto, H.; Yahagi, H.; Kanematsu, M.; Sato, M.; Ito, T.; Uezumi, A.; Hayashi, S.; Miyagoe-Suzuki, Y. Suppression of macrophage functions impairs skeletal muscle regeneration with severe fibrosis. Exp. Cell Res. 2008, 314, 3232–3244. [Google Scholar] [CrossRef]
- Feige, P.; Brun, C.E.; Ritso, M.; Rudnicki, M.A. Orienting Muscle Stem Cells for Regeneration in Homeostasis, Aging, and Disease. Cell Stem Cell 2018, 23, 653–664. [Google Scholar] [CrossRef]
- Swinehart, I.T.; Badylak, S.F. Extracellular matrix bioscaffolds in tissue remodeling and morphogenesis. Dev. Dyn. 2016, 245, 351–360. [Google Scholar] [CrossRef]
- Takeshita, S.; Kikuno, R.; Tezuka, K.; Amann, E. Osteoblast-specific factor 2: Cloning of a putative bone adhesion protein with homology with the insect protein fasciclin I. Biochem. J. 1993, 294, 271–278. [Google Scholar] [CrossRef]
- Kudo, A. Introductory review: Periostin—Gene and protein structure. Cell. Mol. Life Sci. 2017, 74, 4259–4268. [Google Scholar] [CrossRef]
- Horiuchi, K.; Amizuka, N.; Takeshita, S.; Takamatsu, H.; Katsuura, M.; Ozawa, H.; Toyama, Y.; Bonewald, L.F.; Kudo, A. Identification and Characterization of a Novel Protein, Periostin, with Restricted Expression to Periosteum and Periodontal Ligament and Increased Expression by Transforming Growth Factor β. J. Bone Miner. Res. 1999, 14, 1239–1249. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Kashima, T.G.; Nishiyama, T.; Shimazu, K.; Morishita, Y.; Shimazaki, M.; Kii, I.; Horie, H.; Nagai, H.; Kudo, A.; et al. Periostin Is Expressed in Pericryptal Fibroblasts and Cancer-associated Fibroblasts in the Colon. J. Histochem. Cytochem. 2008, 56, 753–764. [Google Scholar] [CrossRef]
- Shimazaki, M.; Nakamura, K.; Kii, I.; Kashima, T.; Amizuka, N.; Li, M.; Saito, M.; Fukuda, K.; Nishiyama, T.; Kitajima, S.; et al. Periostin is essential for cardiac healingafter acute myocardial infarction. J. Exp. Med. 2008, 205, 295–303. [Google Scholar] [CrossRef]
- Goetsch, S.C.; Hawke, T.J.; Gallardo, T.D.; Richardson, J.A.; Garry, D.J. Transcriptional profiling and regulation of the extracellular matrix during muscle regeneration. Physiol. Genom. 2003, 14, 261–271. [Google Scholar] [CrossRef]
- Lorts, A.; Schwanekamp, J.A.; Baudino, T.A.; McNally, E.M.; Molkentin, J.D. Deletion of periostin reduces muscular dystrophy and fibrosis in mice by modulating the transforming growth factor- pathway. Proc. Natl. Acad. Sci. USA 2012, 109, 10978–10983. [Google Scholar] [CrossRef] [PubMed]
- Kudo, A. Periostin in fibrillogenesis for tissue regeneration: Periostin actions inside and outside the cell. Cell. Mol. Life Sci. 2011, 68, 3201–3207. [Google Scholar] [CrossRef] [PubMed]
- Kii, I.; Amizuka, N.; Minqi, L.; Kitajima, S.; Saga, Y.; Kudo, A. Periostin is an extracellular matrix protein required for eruption of incisors in mice. Biochem. Biophys. Res. Commun. 2006, 342, 766–772. [Google Scholar] [CrossRef] [PubMed]
- Norris, R.A.; Damon, B.; Mironov, V.; Kasyanov, V.; Ramamurthi, A.; Moreno-Rodriguez, R.; Trusk, T.; Potts, J.D.; Goodwin, R.L.; Davis, J.; et al. Periostin regulates collagen fibrillogenesis and the biomechanical properties of connective tissues. J. Cell. Biochem. 2007, 101, 695–711. [Google Scholar] [CrossRef]
- Takayama, G.; Arima, K.; Kanaji, T.; Toda, S.; Tanaka, H.; Shoji, S.; McKenzie, A.N.; Nagai, H.; Hotokebuchi, T.; Izuhara, K. Periostin: A novel component of subepithelial fibrosis of bronchial asthma downstream of IL-4 and IL-13 signals. J. Allergy Clin. Immunol. 2006, 118, 98–104. [Google Scholar] [CrossRef]
- Kii, I.; Nishiyama, T.; Li, M.; Matsumoto, K.-I.; Saito, M.; Amizuka, N.; Kudo, A. Incorporation of Tenascin-C into the Extracellular Matrix by Periostin Underlies an Extracellular Meshwork Architecture. J. Biol. Chem. 2010, 285, 2028–2039. [Google Scholar] [CrossRef]
- Baril, P.; Gangeswaran, R.; Mahon, P.C.; Caulee, K.; Kocher, H.M.; Harada, T.; Zhu, M.; Kalthoff, H.; Crnogorac-Jurcevic, T.; Lemoine, N.R. Periostin promotes invasiveness and resistance of pancreatic cancer cells to hypoxia-induced cell death: Role of the β4 integrin and the PI3k pathway. Oncogene 2006, 26, 2082–2094. [Google Scholar] [CrossRef] [PubMed]
- Gillan, L.; Matei, D.; Fishman, D.A.; Gerbin, C.S.; Karlan, B.Y.; Chang, D.D. Periostin secreted by epithelial ovarian carcinoma is a ligand for αVβ3 and αVβ5 integrins and promotes cell motility. Cancer Res. 2002, 62, 5358–5364. [Google Scholar]
- Siriwardena, B.S.M.S.; Kudo, Y.; Ogawa, I.; Kitagawa, M.; Kitajima, S.; Hatano, H.; Tilakaratne, W.M.; Miyauchi, M.; Takata, T. Periostin is frequently overexpressed and enhances invasion and angiogenesis in oral cancer. Br. J. Cancer 2006, 95, 1396–1403. [Google Scholar] [CrossRef]
- Hara, M.; Yokota, K.; Saito, T.; Kobayakawa, K.; Kijima, K.; Yoshizaki, S.; Okazaki, K.; Yoshida, S.; Matsumoto, Y.; Harimaya, K.; et al. Periostin Promotes Fibroblast Migration and Inhibits Muscle Repair After Skeletal Muscle Injury. J. Bone Jt. Surg. Am. Vol. 2018, 100, e108. [Google Scholar] [CrossRef]
- Kudo, H.; Amizuka, N.; Araki, K.; Inohaya, K.; Kudo, A. Zebrafish periostin is required for the adhesion of muscle fiber bundles to the myoseptum and for the differentiation of muscle fibers. Dev. Biol. 2004, 267, 473–487. [Google Scholar] [CrossRef]
- Motohashi, N.; Uezumi, A.; Yada, E.; Fukada, S.-I.; Fukushima, K.; Imaizumi, K.; Miyagoe-Suzuki, Y.; Takeda, S. Muscle CD31(−) CD45(−) Side Population Cells Promote Muscle Regeneration by Stimulating Proliferation and Migration of Myoblasts. Am. J. Pathol. 2008, 173, 781–791. [Google Scholar] [CrossRef] [PubMed]
- De Lageneste, O.D.; Julien, A.; Abou-Khalil, R.; Frangi, G.; Carvalho, C.; Cagnard, N.; Cordier, C.; Conway, S.J.; Colnot, C. Periosteum contains skeletal stem cells with high bone regenerative potential controlled by Periostin. Nat. Commun. 2018, 9, 1–15. [Google Scholar] [CrossRef]
- Hardy, D.; Besnard, A.; Latil, M.; Jouvion, G.; Briand, D.; Thépenier, C.; Pascal, Q.; Guguin, A.; Gayraud-Morel, B.; Cavaillon, J.-M.; et al. Comparative Study of Injury Models for Studying Muscle Regeneration in Mice. PLoS ONE 2016, 11, e0147198. [Google Scholar] [CrossRef]
- Bulfield, G.; Siller, W.G.; Wight, P.A.; Moore, K.J. X chromosome-linked muscular dystrophy (mdx) in the mouse. Proc. Natl. Acad. Sci. USA 1984, 81, 1189–1192. [Google Scholar] [CrossRef] [PubMed]
- Fukada, S.-I.; Morikawa, D.; Yamamoto, Y.; Yoshida, T.; Sumie, N.; Yamaguchi, M.; Ito, T.; Miyagoe-Suzuki, Y.; Takeda, S.; Tsujikawa, K.; et al. Genetic Background Affects Properties of Satellite Cells and mdx Phenotypes. Am. J. Pathol. 2010, 176, 2414–2424. [Google Scholar] [CrossRef]
- Grady, R.; Zhou, H.; Cunningham, J.M.; Henry, M.D.; Campbell, K.P.; Sanes, J.R. Maturation and Maintenance of the Neuromuscular Synapse. Neuron 2000, 25, 279–293. [Google Scholar] [CrossRef]
- Maier, A.; Mayne, R. Distribution of connective tissue proteins in chick muscle spindles as revealed by monoclonal antibodies: A unique distribution of brachionectin/tenascin. Am. J. Anat. 1987, 180, 226–236. [Google Scholar] [CrossRef]
- Welser, J.V.; Rooney, J.E.; Cohen, N.C.; Gurpur, P.B.; Singer, C.A.; Evans, R.A.; Haines, B.A.; Burkin, D.J. Myotendinous Junction Defects and Reduced Force Transmission in Mice that Lack α7 Integrin and Utrophin. Am. J. Pathol. 2009, 175, 1545–1554. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Deng, R.; Chai, Y.; Chen, H.; Hu, B.; Wang, X.; Zhu, S.; Cao, Y.; Ni, S.; Wan, M.; et al. Macrophage-lineage TRAP+ cells recruit periosteum-derived cells for periosteal osteogenesis and regeneration. J. Clin. Investig. 2019, 129, 2578–2594. [Google Scholar] [CrossRef]
- Shao, R.; Bao, S.; Bai, X.; Blanchette, C.; Anderson, R.M.; Dang, T.; Gishizky, M.L.; Marks, J.R.; Wang, X.-F. Acquired Expression of Periostin by Human Breast Cancers Promotes Tumor Angiogenesis through Up-Regulation of Vascular Endothelial Growth Factor Receptor 2 Expression. Mol. Cell. Biol. 2004, 24, 3992–4003. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; LiZhou, L.; Gao, F.; Xingxin, L.; Qin, P.; Liang, X.; Zhang, J.; Qiao, X.; Lin, L.; Zhao, Q.; et al. Periostin promotes tumor angiogenesis in pancreatic cancer via Erk/VEGF signaling. Oncotarget 2016, 7, 40148–40159. [Google Scholar] [CrossRef]
- Yamada, M.; Sankoda, Y.; Tatsumi, R.; Mizunoya, W.; Ikeuchi, Y.; Sunagawa, K.; Allen, R.E. Matrix metalloproteinase-2 mediates stretch-induced activation of skeletal muscle satellite cells in a nitric oxide-dependent manner. Int. J. Biochem. Cell Biol. 2008, 40, 2183–2191. [Google Scholar] [CrossRef]
- Kanematsu, A.; Marui, A.; Yamamoto, S.; Ozeki, M.; Hirano, Y.; Yamamoto, M.; Ogawa, O.; Komeda, M.; Tabata, Y. Type I collagen can function as a reservoir of basic fibroblast growth factor. J. Control. Release 2004, 99, 281–292. [Google Scholar] [CrossRef]
- Chen, X.; Li, Y. Role of matrix metalloproteinases in skeletal muscle. Cell Adhes. Migr. 2009, 3, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Blain, A.M.; Straub, V.W. δ-Sarcoglycan-deficient muscular dystrophy: From discovery to therapeutic approaches. Skelet. Muscle 2011, 1, 13. [Google Scholar] [CrossRef]
- Turk, R.; Hsiao, J.J.; Smits, M.M.; Ng, B.H.; Pospisil, T.C.; Jones, K.S.; Campbell, K.P.; Wright, M.E. Molecular Signatures of Membrane Protein Complexes Underlying Muscular Dystrophy. Mol. Cell. Proteom. 2016, 15, 2169–2185. [Google Scholar] [CrossRef]
- Ito, N.; Ruegg, U.T.; Kudo, A.; Miyagoe-Suzuki, Y.; Takeda, S. Activation of calcium signaling through Trpv1 by nNOS and peroxynitrite as a key trigger of skeletal muscle hypertrophy. Nat. Med. 2013, 19, 101–106. [Google Scholar] [CrossRef]
- Ito, N.; Ruegg, U.T.; Takeda, S. ATP-Induced Increase in Intracellular Calcium Levels and Subsequent Activation of mTOR as Regulators of Skeletal Muscle Hypertrophy. Int. J. Mol. Sci. 2018, 19, 2804. [Google Scholar] [CrossRef]
- Ito, N.; Kii, I.; Shimizu, N.; Tanaka, H.; Takeda, S. Direct reprogramming of fibroblasts into skeletal muscle progenitor cells by transcription factors enriched in undifferentiated subpopulation of satellite cells. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Clone | Dilution | Source, Catalog Number |
|---|---|---|---|
| rabbit anti-periostin | polyclonal | 300 | Reference [12] |
| mouse anti-Neurofilament H (NF-H), Nonphosphorylated | SMI32 | 1500 | Bioletend (San Diego, CA, USA), 801701 |
| mouse anti-S46 | S46 | 800 | Developmental Studies Hybridoma Bank (Iowa City, IA, USA), S46 |
| mouse anti-BF-45 | BF-45 | 40 | Developmental Studies Hybridoma Bank (Iowa City, IA, USA), BF-45 |
| rat-anti-CD31 | 390 | 100 | Immunotech (Ocala, FL, USA) |
| rat-anti-PDGFRa | APA5 | 2000 | BD Pharmingen (Franklin Lakes, NJ, USA), 558774 |
| mouse- anti-MF20 | MF20 | 500 | R&D Systems (Minneapolis, MN, USA), MAB4470 |
| rat anti-laminin-a2 | 4H8-2 | 200 | Alexis Biochemicals (San Diego, CA, USA), ALX-804-190 |
| mouse anti-Pax7 | PAX7 | 80 | Santa Cruz Biotech (Santa Cruz, CA, USA), sc-81648 |
| mouse anti-MyoD | 5.8A | 400 | Santa Cruz Biotech (Santa Cruz, CA, USA), sc-32758 |
| mouse anti-myogenin | F5D | 1000 | Santa Cruz Biotech (Santa Cruz, CA, USA), sc-12732 |
| Primer Sequence | |
|---|---|
| periostin for all isoforms | 5′-atgtacaacaatctggggcttt-3′ |
| 5′-cgacaccatttgtggcaatc-3′ | |
| PDGFRa | 5′-gcatcttcgacaacctctacac-3′ |
| 5′-accatcatgccaggataggg-3′ | |
| TGFb1 | 5′-cagagaagaactgctgtgtgcg-3′ |
| 5′-cgggttgtgttggttgtagagg-3′ | |
| periostin pair 1 | 5′-gataaatacatccaaatcaagtttgttcg-3′ |
| 5′-cgtggatcacttctgtcaccgtttcgc-3′ | |
| periostin pair 2 | 5′-ctgaaaaacagactcgggaagaacg-3’ |
| 5′-aaactctgtggtctggcctctggg-3′ | |
| periostin pair 3 | 5′-gataaaatacatccaaatcaagtttgttcg-3′ |
| 5′-ctgaaaaacagactcgggaagaacg-3′ | |
| BMP2 | 5′-cagtgggagagcttcgacgtca-3′ |
| 5′-ggagacacctgggttctcctct-3′ | |
| BMP4 | 5′-ggccaaacgtagtcccaagca-3′ |
| 5′-ttccagcccacgtcactgaag-3′ | |
| Acvr1 | 5′-aggagagtcaatgctgtcct-3′ |
| 5′-gagacatctgcttccgtcaa-3′ | |
| 18s rRNA | 5′-taccctggcggtgggattaac-3′ |
| 5′-cgagagaagaccacgccaac-3′ | |
| TATA-binding protein (TBP) | 5′-cagcctcagtacagcaatcaac-3′ |
| 5′-taggggtcataggagtcattgg-3′ | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ito, N.; Miyagoe-Suzuki, Y.; Takeda, S.; Kudo, A. Periostin Is Required for the Maintenance of Muscle Fibers during Muscle Regeneration. Int. J. Mol. Sci. 2021, 22, 3627. https://doi.org/10.3390/ijms22073627
Ito N, Miyagoe-Suzuki Y, Takeda S, Kudo A. Periostin Is Required for the Maintenance of Muscle Fibers during Muscle Regeneration. International Journal of Molecular Sciences. 2021; 22(7):3627. https://doi.org/10.3390/ijms22073627
Chicago/Turabian StyleIto, Naoki, Yuko Miyagoe-Suzuki, Shin’ichi Takeda, and Akira Kudo. 2021. "Periostin Is Required for the Maintenance of Muscle Fibers during Muscle Regeneration" International Journal of Molecular Sciences 22, no. 7: 3627. https://doi.org/10.3390/ijms22073627
APA StyleIto, N., Miyagoe-Suzuki, Y., Takeda, S., & Kudo, A. (2021). Periostin Is Required for the Maintenance of Muscle Fibers during Muscle Regeneration. International Journal of Molecular Sciences, 22(7), 3627. https://doi.org/10.3390/ijms22073627