
Comparative Study of Structural Changes of Polylactide and Poly(ethylene terephthalate) in the Presence of Trichoderma viride

 , ,
, ,
Abstract
1. Introduction
2. Results
2.1. Identification and Characterization of T. viride GZ1 Metabolic Activity
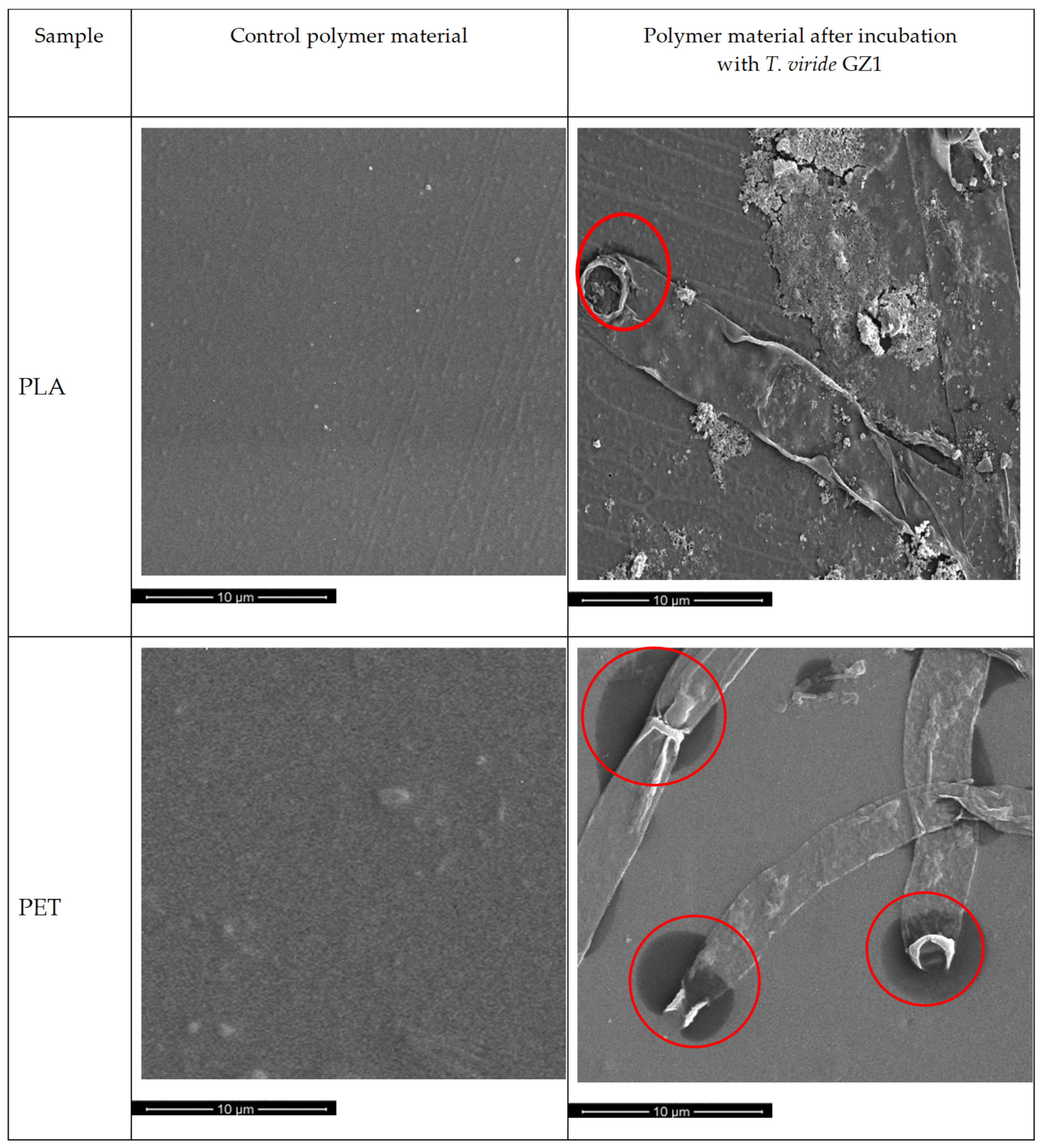
2.2. Adhesion and Growth of T. viride GZ1 on Polymer Material
2.3. Analysis of PET and PLA Biodegradation by T. viride GZ1
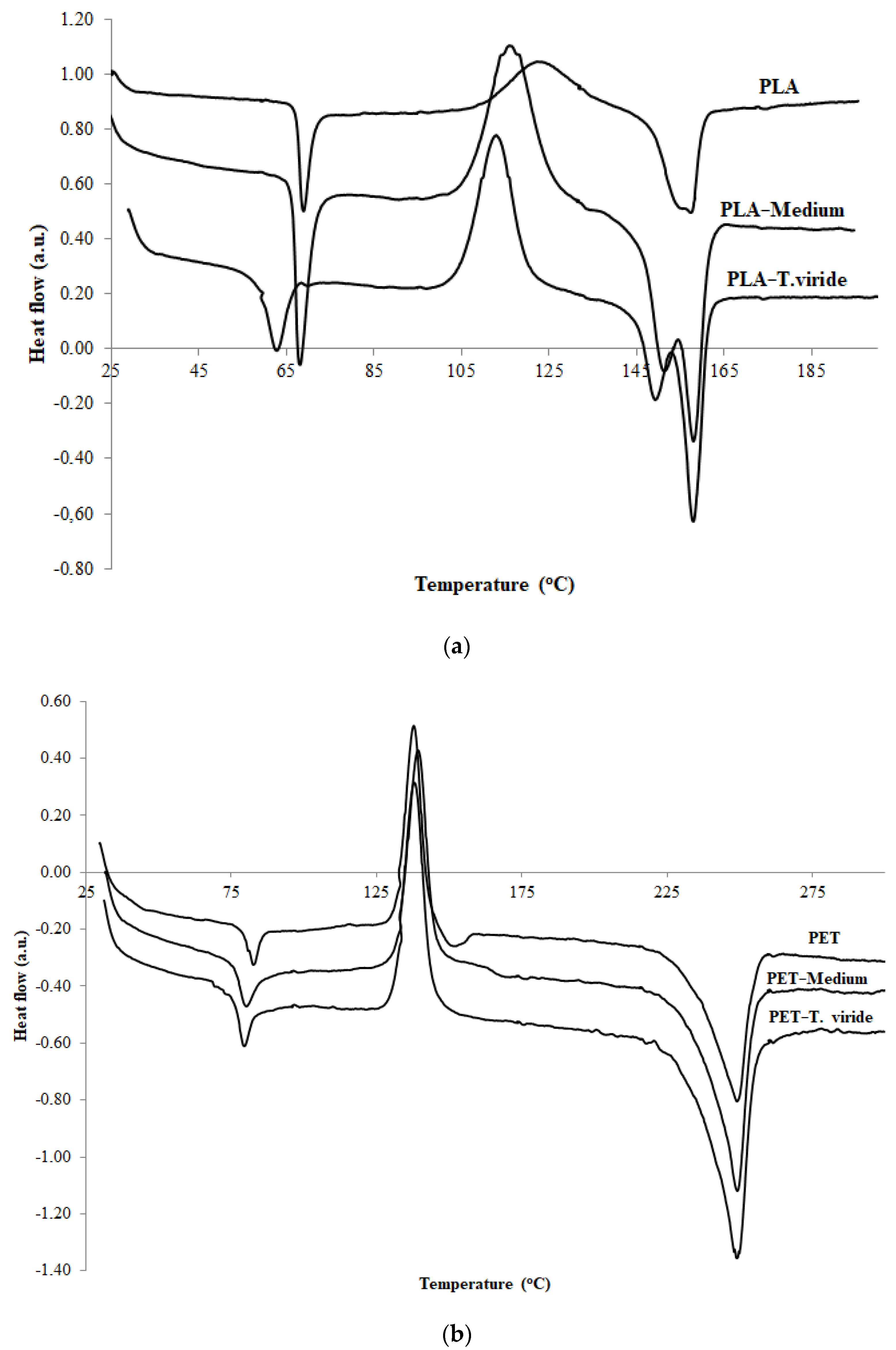
2.3.1. Thermal Properties
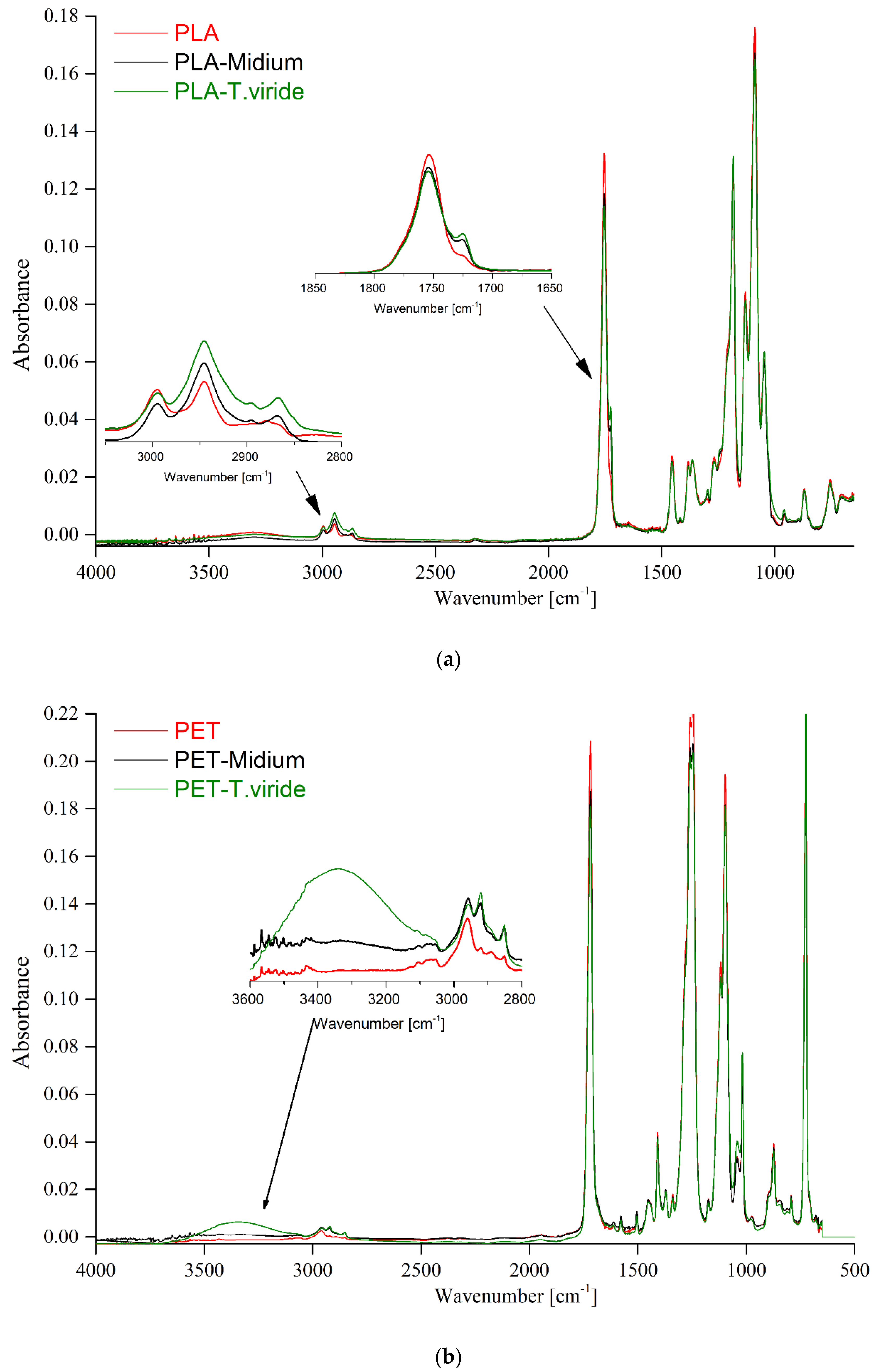
2.3.2. Fourier-Transform Infrared Spectroscopy (FTIR)
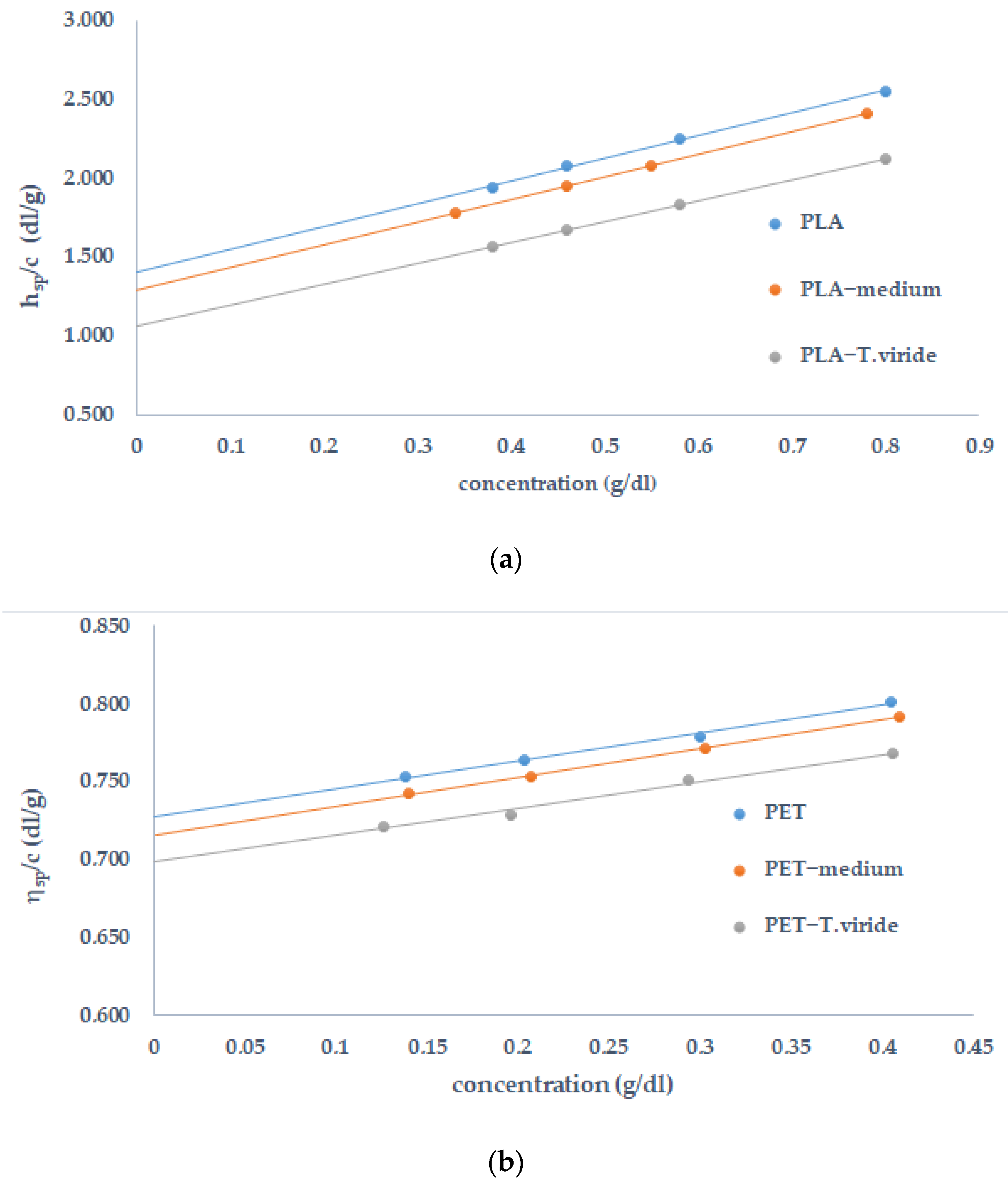
2.4. Determination of Molecular Weight of Polymers Incubacted with the Fungus
2.5. Hydrophobins
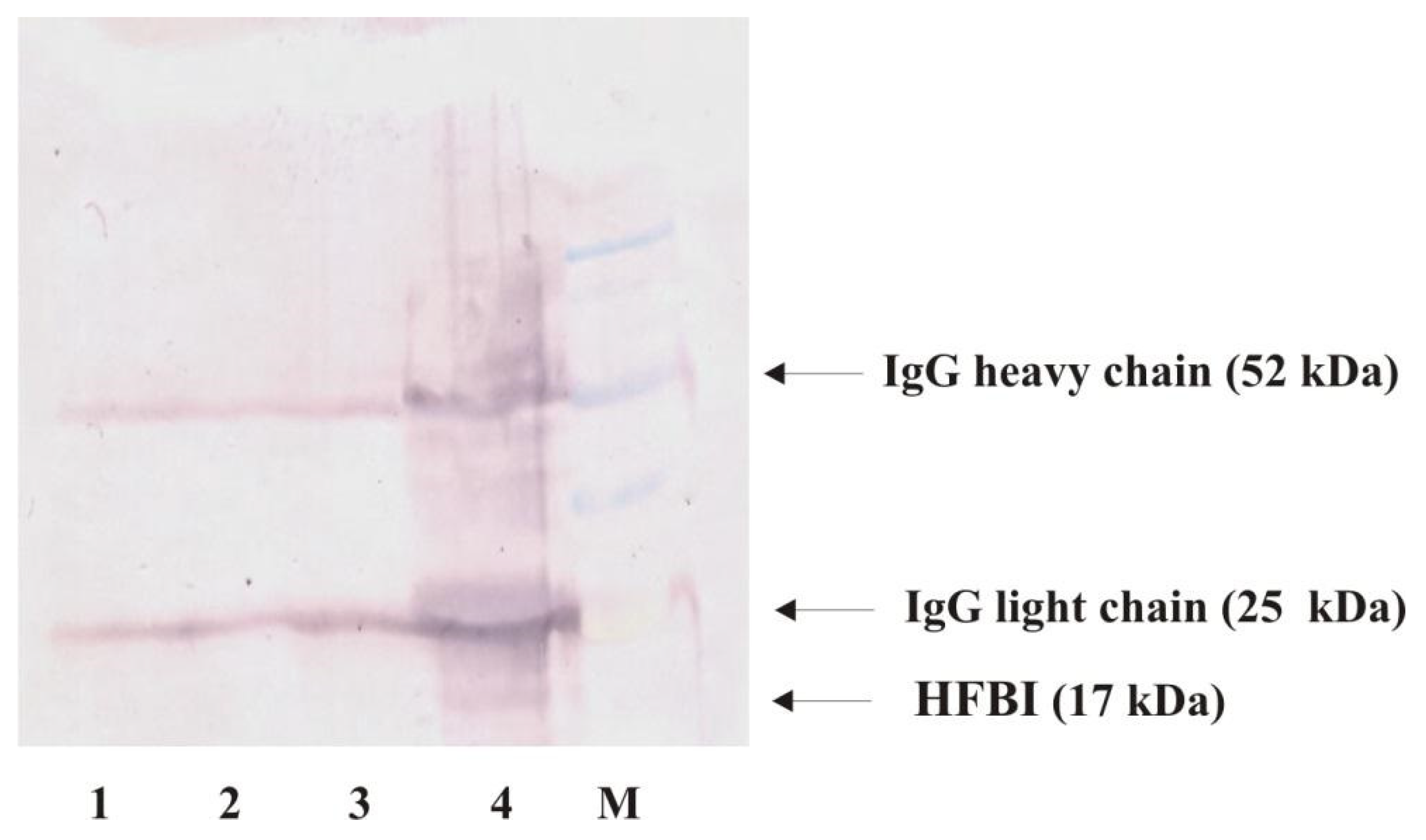
2.5.1. Detection of Hydrophobins in Fungal Culture
2.5.2. Detection of Hydrophobin Film
3. Discussion
3.1. Identification and Metabolic Activity of T. viride GZ1
3.2. Changes in the Structure and Thermal Properties of PET and PLA after Incubation with the Fungus
3.3. T. viride GZ1 Hydrophobin Proteins and Their Potential in Biodegradation
4. Materials and Methods
4.1. Polymer Materials
4.2. Molecular Identification of T. viride GZ1 and Fungal Culture Conditions
4.3. Characteristics of the T. viride GZ1 Strain
4.3.1. Analysis of Fungal Metabolic Activity
4.3.2. Growth of T. viride GZ1 on Polymer Materials
4.4. Assessment of the Properties of Polymeric Materials after Incubation with the Fungus
4.4.1. Scanning Electron Microscopy (SEM)
4.4.2. Differential Scanning Calorimetry (DSC)
4.4.3. Fourier-Transform Infrared Spectroscopy (FTIR)
4.4.4. Viscosity Measurements
4.5. Hydrophobins
4.5.1. Detection of T. viride GZ1 Hydrophobins
4.5.2. Detection of Hydrophobin Film—Atomic Force Microscopy (AFM)
4.6. Statistical Analysis
5. Conclusions
- The fungi adhere to the surface of the material and the expression of hydrophobin-coding genes increases.
- Increase in the amount of hydrophobins in the environment of the fungus.
- A hydrophobic film forms on the polymer surface.
- Hydrolytic enzymes produced by T. viride are immobilized on the surface of the film and increase in effectiveness.
- Biodegradable changes of PLA and PET.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ragaert, K.; Delva, L.; Van Geem, K. Mechanical and chemical recycling of solid plastic waste. Waste Manag. 2017, 69, 24–58. [Google Scholar] [CrossRef]
- Biundo, A.; Ribitsch, D.; Guebitz, G.M. Surface engineering of polyester-degrading enzymes to improve efficiency and tune specificity. Appl. Microbiol. Biotechnol. 2018, 102, 3551–3559. [Google Scholar] [CrossRef]
- Janczak, K.; Dąbrowska, G.; Hrynkiewicz, K.; Raszkowska-Kaczor, A. Searching for the fungi capable to grow on the polymeric materials. Przem. Chem. 2014, 93, 1206–1209. [Google Scholar] [CrossRef]
- Janczak, K.; Znajewska, Z.; Narbutt, O.; Raszkowska-Kaczor, A.; Dąbrowska, G. Serratia sp. in the PLA and PCL-supporting biodegradation preparations. Przem. Chem. 2016, 95, 943–947. [Google Scholar] [CrossRef]
- Szumigaj, J.; Zakowska, Z.; Klimek, L.; Rosicka-Kaczmarek, J.; Bartkowiak, A. Assessment of polylactide foil degradation as a result of filamentous fungi activity. Pol. J. Environ. Stud. 2008, 17, 335–341. [Google Scholar]
- Tokiwa, Y.; Calabia, B.P. Biodegradability and biodegradation of poly(lactide). Appl. Microbiol. Biotechnol. 2006, 72, 244–251. [Google Scholar] [CrossRef]
- Ohkita, T.; Lee, S.-H. Thermal degradation and biodegradability of poly (lactic acid)/corn starch biocomposites. J. Appl. Polym. Sci. 2006, 100, 3009–3017. [Google Scholar] [CrossRef]
- Qi, X.; Ren, Y.; Wang, X. New advances in the biodegradation of poly(lactic) acid. Int. Biodeterior. Biodegrad. 2017, 117, 215–223. [Google Scholar] [CrossRef]
- Kawai, F.; Kawabata, T.; Oda, M. Current knowledge on enzymatic PET degradation and its possible application to waste stream management and other fields. Appl. Microbiol. Biotechnol. 2019, 103, 4253–4268. [Google Scholar] [CrossRef] [PubMed]
- Müller, R.-J.; Kleeberg, I.; Deckwer, W.-D. Biodegradation of polyesters containing aromatic constituents. J. Biotechnol. 2001, 86, 87–95. [Google Scholar] [CrossRef]
- Stasiek, A.; Raszkowska-Kaczor, A.; Janczak, K. Foaming of modified polylactide. Przem. Chem. 2014, 93, 217–219. [Google Scholar] [CrossRef]
- Sánchez, C. Fungal potential for the degradation of petroleum-based polymers: An overview of macro- and microplastics biodegradation. Biotechnol. Adv. 2020, 40, 107501. [Google Scholar] [CrossRef]
- Casalini, T.; Rossi, F.; Castrovinci, A.; Perale, G. A perspective on polylactic acid-based polymers use for nanoparticles synthesis and applications. Front. Bioeng. Biotechnol. 2019, 7, 259. [Google Scholar] [CrossRef]
- Shaxson, L. Structuring policy problems for plastics, the environment and human health: Reflections from the UK. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 2141–2151. [Google Scholar] [CrossRef] [PubMed]
- Head, I.M.; Swannell, R.P.J. Bioremediation of petroleum hydrocarbon contaminants in marine habitats. Curr. Opin. Biotechnol. 1999, 10, 234–239. [Google Scholar] [CrossRef]
- Znajewska, Z.; Dąbrowska, G.B.; Hrynkiewicz, K.; Janczak, K. Biodegradation of polycaprolactone by Trichoderma viride fungi. Przem. Chem. 2018, 97, 1676–1679. [Google Scholar] [CrossRef]
- Kannahi, M.; Thamizhmarai, T. Biodegradation of plastic by Aspergillus sp. Int. J. Trend Sci. Res. Dev. 2018, 2, 683–690. [Google Scholar] [CrossRef]
- Romero, M.C.; Salvioli, M.L.; Cazau, M.C.; Arambarri, A.M. Pyrene degradation by yeasts and filamentous fungi. Environ. Pollut. 2002, 117, 159–163. [Google Scholar] [CrossRef]
- Abdel-Motaal, F.F.; El-Sayed, M.A.; El-Zayat, S.A.; Ito, S.I. Biodegradation of poly (ε-caprolactone) (PCL) film and foam plastic by Pseudozyma japonica sp. nov., a novel cutinolytic ustilaginomycetous yeast species. 3 Biotech 2014, 4, 507–512. [Google Scholar] [CrossRef]
- Janczak, K.; Hrynkiewicz, K.; Znajewska, Z.; Dąbrowska, G. Use of rhizosphere microorganisms in the biodegradation of PLA and PET polymers in compost soil. Int. Biodeterior. Biodegrad. 2018, 130, 65–75. [Google Scholar] [CrossRef]
- Loredo-Treviño, A.; García, G.; Velasco-Téllez, A.; Rodríguez-Herrera, R.; Aguilar, C.N. Polyurethane foam as substrate for fungal strains. Adv. Biosci. Biotechnol. 2011, 2, 52–58. [Google Scholar] [CrossRef]
- Vertommen, M.A.M.E.; Nierstrasz, V.A.; van der Veer, M.; Warmoeskerken, M.M.C.G. Enzymatic surface modification of poly(ethylene terephthalate). J. Biotechnol. 2005, 120, 376–386. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Hiraga, K.; Takehana, T.; Taniguchi, I.; Yamaji, H.; Maeda, Y.; Toyohara, K.; Miyamoto, K.; Kimura, Y.; Oda, K. A bacterium that degrades and assimilates poly(ethylene terephthalate). Science 2016, 351, 1196–1199. [Google Scholar] [CrossRef]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef]
- Kashyap, P.L.; Rai, P.; Srivastava, A.K.; Kumar, S. Trichoderma for climate resilient agriculture. World J. Microbiol. Biotechnol. 2017, 33, 155. [Google Scholar] [CrossRef] [PubMed]
- Scholtmeijer, K.; Wessels, J.G.H.; Wösten, H.A.B. Fungal hydrophobins in medical and technical applications. Appl. Microbiol. Biotechnol. 2001, 56, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Przylucka, A.; Akcapinar, G.B.; Chenthamara, K.; Cai, F.; Grujic, M.; Karpenko, J.; Livoi, M.; Shen, Q.; Kubicek, C.P.; Druzhinina, I.S. HFB7—A novel orphan hydrophobin of the Harzianum and Virens clades of Trichoderma, is involved in response to biotic and abiotic stresses. Fungal Genet. Biol. 2017, 102, 63–76. [Google Scholar] [CrossRef]
- Tymiński, Ł.P.; Znajewska, Z.; Dąbrowska, G.B. Characteristics and functions of hydrophobins and their use in manifold industries. Postępy Mikrobiol. Adv. Microbiol. 2019, 57, 374–384. [Google Scholar] [CrossRef]
- Raffaini, G.; Milani, R.; Ganazzoli, F.; Resnati, G.; Metrangolo, P. Atomistic simulation of hydrophobin HFBII conformation in aqueous and fluorous media and at the water/ vacuum interface. J. Mol. Graph. Model. 2016, 63, 8–14. [Google Scholar] [CrossRef]
- Cicatiello, P.; Dardano, P.; Pirozzi, M.; Gravagnuolo, A.M.; De Stefano, L.; Giardina, P. Self-assembly of two hydrophobins from marine fungi affected by interaction with surfaces. Biotechnol. Bioeng. 2017, 114, 2173–2186. [Google Scholar] [CrossRef]
- Pham, C.L.L.; Rodríguez de Francisco, B.; Valsecchi, I.; Dazzoni, R.; Pillé, A.; Lo, V.; Ball, S.R.; Cappai, R.; Wien, F.; Kwan, A.H.; et al. Probing structural changes during self-assembly of surface-active hydrophobin proteins that form functional amyloids in fungi. J. Mol. Biol. 2018, 430, 3784–3801. [Google Scholar] [CrossRef]
- Wessels, J.G.H. Developmental regulation of fungal cell wall formation. Annu. Rev. Phytopathol. 1994, 32, 413–437. [Google Scholar] [CrossRef]
- Guzmán-Guzmán, P.; Alemán-Duarte, M.I.; Delaye, L.; Herrera-Estrella, A.; Olmedo-Monfil, V. Identification of effector-like proteins in Trichoderma spp. and role of a hydrophobin in the plant-fungus interaction and mycoparasitism. BMC Genet. 2017, 18, 16. [Google Scholar] [CrossRef] [PubMed]
- Espino-Rammer, L.; Ribitsch, D.; Przylucka, A.; Marold, A.; Greimel, K.J.; Acero, E.H.; Guebitz, G.M.; Kubicek, C.P.; Druzhinina, I.S. Two novel class ii hydrophobins from Trichoderma spp. stimulate enzymatic hydrolysis of poly(ethylene terephthalate) when expressed as fusion proteins. Appl. Environ. Microbiol. 2013, 79, 4230–4238. [Google Scholar] [CrossRef]
- Khalesi, M.; Gebruers, K.; Derdelinckx, G. Recent advances in fungal hydrophobin towards using in industry. Protein J. 2015, 34, 243–255. [Google Scholar] [CrossRef]
- Kulkarni, S.; Nene, S.; Joshi, K. Production of hydrophobins from fungi. Process Biochem. 2017, 61, 1–11. [Google Scholar] [CrossRef]
- Houmadi, S.; Ciuchi, F.; De Santo, M.P.; De Stefano, L.; Rea, I.; Giardina, P.; Almenante, A.; Lacaze, E.; Giocondo, M. Langmuir-blodgett film of hydrophobin protein from Pleurotus ostreatus at the air-water interface. Langmuir 2008, 24, 12953–12957. [Google Scholar] [CrossRef] [PubMed]
- Jensen, B.G.; Andersen, M.R.; Pedersen, M.H.; Frisvad, J.C.; Søndergaard, I. Hydrophobins from Aspergillus species cannot be clearly divided into two classes. BMC Res. Notes 2010, 3, 344. [Google Scholar] [CrossRef]
- Seidl-Seiboth, V.; Gruber, S.; Sezerman, U.; Schwecke, T.; Albayrak, A.; Neuhof, T.; von Döhren, H.; Baker, S.E.; Kubicek, C.P. Novel hydrophobins from Trichoderma define a new hydrophobin subclass: Protein properties, evolution, regulation and processing. J. Mol. Evol. 2011, 72, 339–351. [Google Scholar] [CrossRef]
- Pitocchi, R.; Cicatiello, P.; Birolo, L.; Piscitelli, A.; Bovio, E.; Cristina Varese, G.; Giardina, P. Cerato-platanins from marine fungi as effective protein biosurfactants and bioemulsifiers. Int. J. Mol. Sci. 2020, 21, 2913. [Google Scholar] [CrossRef]
- Ribitsch, D.; Herrero Acero, E.; Przylucka, A.; Zitzenbacher, S.; Marold, A.; Gamerith, C.; Tscheließnig, R.; Jungbauer, A.; Rennhofer, H.; Lichtenegger, H.; et al. Enhanced cutinase-catalyzed hydrolysis of polyethylene terephthalate by covalent fusion to hydrophobins. Appl. Environ. Microbiol. 2015, 81, 3586–3592. [Google Scholar] [CrossRef]
- Puspitasari, N.; Tsai, S.-L.; Lee, C.-K. Fungal hydrophobin RolA enhanced PETase hydrolysis of polyethylene terephthalate. Appl. Biochem. Biotechnol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Olewnik-Kruszkowska, E.; Koter, I.; Skopińska-Wiśniewska, J.; Richert, J. Degradation of polylactide composites under UV irradiation at 254 nm. J. Photochem. Photobiol. A Chem. 2015, 311, 144–153. [Google Scholar] [CrossRef]
- Mecozzi, M.; Nisini, L. The differentiation of biodegradable and non-biodegradable polyethylene terephthalate (PET) samples by FTIR spectroscopy: A potential support for the structural differentiation of PET in environmental analysis. Infrared Phys. Technol. 2019, 101, 119–126. [Google Scholar] [CrossRef]
- Lai, W.C.; Liau, W. Bin Thermo-oxidative degradation of poly(ethylene glycol)/poly(L-lactic acid) blends. Polymer (Guildf) 2003, 44, 8103–8109. [Google Scholar] [CrossRef]
- Kaur, P. Determination of Mark-Houwink Parameters for Polylactide. J. Emerg. Technol. Innov. Res. 2018, 5, 52–57. [Google Scholar]
- Li, K.; Song, X.; Zhang, D. Molecular weight evaluation of depolymerized poly(ethylene terephthalate) using intrinsic viscosity. J. Appl. Polym. Sci. 2008, 109, 1294–1297. [Google Scholar] [CrossRef]
- Cicatiello, P.; Gravagnuolo, A.M.; Gnavi, G.; Varese, G.C.; Giardina, P. Marine fungi as source of new hydrophobins. Int. J. Biol. Macromol. 2016, 92, 1229–1233. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.Y.; Rhee, Y.H. Biodegradation of microbial and synthetic polyesters by fungi. Appl. Microbiol. Biotechnol. 2003, 61, 300–308. [Google Scholar] [CrossRef]
- Suyama, T.; Tokiwa, Y.; Ouichanpagdee, P.; Kanagawa, T.; Kamagata, Y. Phylogenetic affiliation of soil bacteria that degrade aliphatic polyesters available commercially as biodegradable plastics. Appl. Environ. Microbiol. 1998, 64, 5008–5011. [Google Scholar] [CrossRef]
- Guigón-López, C.; Carvajal-Millán, E.; De León-Renova, N.P.; Vargas-Albores, F.; Bravo-Luna, L.; Guerrero-Prieto, V.M. Microcalorimetric measurement of Trichoderma spp. growth at different temperatures. Thermochim. Acta 2010, 509, 40–45. [Google Scholar] [CrossRef]
- Krueger, M.C.; Harms, H.; Schlosser, D. Prospects for microbiological solutions to environmental pollution with plastics. Appl. Microbiol. Biotechnol. 2015, 99, 8857–8874. [Google Scholar] [CrossRef] [PubMed]
- Wei, R.; Oeser, T.; Zimmermann, W. Synthetic polyester-hydrolyzing enzymes from thermophilic Actinomycetes. Adv. Appl. Microbiol. 2014, 89, 267–305. [Google Scholar] [CrossRef] [PubMed]
- Santo, M.; Weitsman, R.; Sivan, A. The role of the copper-binding enzyme—laccase—in the biodegradation of polyethylene by the actinomycete Rhodococcus ruber. Int. Biodeterior. Biodegrad. 2013, 84, 204–210. [Google Scholar] [CrossRef]
- Narancic, T.; O’Connor, K.E. Plastic waste as a global challenge: Are biodegradable plastics the answer to the plastic waste problem? Microbiology 2019, 165, 129–137. [Google Scholar] [CrossRef]
- Carniel, A.; Valoni, É.; Nicomedes, J.; Gomes, A.d.C.; de Castro, A.M. Lipase from Candida antarctica (CALB) and cutinase from Humicola insolens act synergistically for PET hydrolysis to terephthalic acid. Process Biochem. 2017, 59, 84–90. [Google Scholar] [CrossRef]
- Kim, J.M.; Jeon, C.O. Isolation and characterization of a new benzene, toluene, and ethylbenzene degrading bacterium, Acinetobacter sp. B113. Curr. Microbiol. 2009, 58, 70–75. [Google Scholar] [CrossRef]
- Akutsu, Y.; Nakajima-Kambe, T.; Nomura, N.; Nakahara, T. Purification and properties of a polyester polyurethane-degrading enzyme from Comamonas acidovorans TB-35. Appl. Environ. Microbiol. 1998, 64, 62–67. [Google Scholar] [CrossRef]
- Nimchua, T.; Eveleigh, D.E.; Sangwatanaroj, U.; Punnapayak, H. Screening of tropical fungi producing polyethylene terephthalate-hydrolyzing enzyme for fabric modification. J. Ind. Microbiol. Biotechnol. 2008, 35, 843. [Google Scholar] [CrossRef] [PubMed]
- Nimchua, T.; Punnapayak, H.; Zimmermann, W. Comparison of the hydrolysis of polyethylene terephthalate fibers by a hydrolase from Fusarium oxysporum LCH I and Fusarium solani f. sp. pisi. Biotechnol. J. 2007, 2, 361–364. [Google Scholar] [CrossRef]
- Nair, J.; Okamitsu, K. Microbial inoculants for small scale composting of putrescible kitchen wastes. Waste Manag. 2010, 30, 977–982. [Google Scholar] [CrossRef]
- Mutschlechner, M.; Illmer, P.; Wagner, A.O. Biological pre-treatment: Enhancing biogas production using the highly cellulolytic fungus Trichoderma viride. Waste Manag. 2015, 43, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.A.; Hasan, F.; Hameed, A.; Ahmed, S. Biological degradation of plastics: A comprehensive review. Biotechnol. Adv. 2008, 26, 246–265. [Google Scholar] [CrossRef] [PubMed]
- Sivan, A. New perspectives in plastic biodegradation. Curr. Opin. Biotechnol. 2011, 22, 422–426. [Google Scholar] [CrossRef] [PubMed]
- Wei, R.; Zimmermann, W. Microbial enzymes for the recycling of recalcitrant petroleum-based plastics: How far are we? Microb. Biotechnol. 2017, 10, 1308–1322. [Google Scholar] [CrossRef] [PubMed]
- Webb, H.; Arnott, J.; Crawford, R.; Ivanova, E. Plastic degradation and its environmental implications with special reference to poly(ethylene terephthalate). Polymers 2012, 5, 1–18. [Google Scholar] [CrossRef]
- Urbanek, A.K.; Rymowicz, W.; Strzelecki, M.C.; Kociuba, W.; Franczak, Ł.; Mirończuk, A.M. Isolation and characterization of Arctic microorganisms decomposing bioplastics. AMB Express 2017, 7, 148. [Google Scholar] [CrossRef]
- Munir, E.; Harefa, R.S.M.; Priyani, N.; Suryanto, D. Plastic degrading fungi Trichoderma viride and Aspergillus nomius isolated from local landfill soil in Medan. IOP Conf. Ser. Earth Environ. Sci. 2018, 126, 012145. [Google Scholar] [CrossRef]
- Lipsa, R.; Tudorachi, N.; Darie-Nita, R.N.; Oprică, L.; Vasile, C.; Chiriac, A. Biodegradation of poly(lactic acid) and some of its based systems with Trichoderma viride. Int. J. Biol. Macromol. 2016, 88, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Olewnik-Kruszkowska, E.; Nowaczyk, J.; Kadac, K. Effect of ozone exposure on thermal and structural properties of polylactide based composites. Polym. Test. 2016, 56, 299–307. [Google Scholar] [CrossRef]
- Olewnik-Kruszkowska, E.; Burkowska-But, A.; Tarach, I.; Walczak, M.; Jakubowska, E. Biodegradation of polylactide-based composites with an addition of a compatibilizing agent in different environments. Int. Biodeterior. Biodegrad. 2020, 147, 104840. [Google Scholar] [CrossRef]
- Tabi, T.; Sajo, I.E.; Szabo, F.; Luyt, A.S.; Kovacs, J.G. Crystalline structure of annealed polylactic acid and its relation to processing. Express Polym. Lett. 2010, 4, 659–668. [Google Scholar] [CrossRef]
- Stepczyńska, M.; Rytlewski, P. Enzymatic degradation of flax-fibers reinforced polylactide. Int. Biodeterior. Biodegrad. 2018, 126, 160–166. [Google Scholar] [CrossRef]
- Itävaara, M.; Karjomaa, S.; Selin, J.F. Biodegradation of polylactide in aerobic and anaerobic thermophilic conditions. Chemosphere 2002, 46, 879–885. [Google Scholar] [CrossRef]
- Stloukal, P.; Kucharczyk, P. Acceleration of polylactide degradation under biotic and abiotic conditions through utilization of a new, experimental, highly compatible additive. Polym. Degrad. Stab. 2017, 142, 217–225. [Google Scholar] [CrossRef]
- Richert, A.; Olewnik-Kruszkowska, E.; Adamska, E.; Tarach, I. Enzymatic degradation of bacteriostatic polylactide composites. Int. Biodeterior. Biodegrad. 2019, 142, 103–108. [Google Scholar] [CrossRef]
- Zaaba, N.F.; Jaafar, M. A review on degradation mechanisms of polylactic acid: Hydrolytic, photodegradative, microbial, and enzymatic degradation. Polym. Eng. Sci. 2020, 60, 2061–2075. [Google Scholar] [CrossRef]
- Nakkabi, A.; Elmoualij, N.; Saad, I.K.; Fahim, M.F. Biodegradation of Poly (Ethylene Terephthalate) by Bacillus Subtilis. Int. J. Recent Adv. Multidiscip. Res. 2015, 2, 1060–1062. [Google Scholar]
- Farzi, A.; Dehnad, A.; Fotouhi, A.F. Biodegradation of polyethylene terephthalate waste using Streptomyces species and kinetic modeling of the process. Biocatal. Agric. Biotechnol. 2019, 17, 25–31. [Google Scholar] [CrossRef]
- Benitez, T.; Rincón, A.M.; Limón, M.C.; Codón, A.C. Biocontrol mechanisms of Trichoderma strains. Int. Microbiol. 2004, 7, 249–260. [Google Scholar]
- Kershaw, M.J.; Talbot, N.J. Hydrophobins and repellents: Proteins with fundamental roles in fungal morphogenesis. Fungal Genet. Biol. 1998, 23, 18–33. [Google Scholar] [CrossRef] [PubMed]
- Linder, M.B.; Szilvay, G.R.; Nakari-Setälä, T.; Penttilä, M.E. Hydrophobins: The protein-amphiphiles of filamentous fungi. FEMS Microbiol. Rev. 2005, 29, 877–896. [Google Scholar] [CrossRef]
- Hou, S.; Yang, K.; Qin, M.; Feng, X.Z.; Guan, L.; Yang, Y.; Wang, C. Patterning of cells on functionalized poly(dimethylsiloxane) surface prepared by hydrophobin and collagen modification. Biosens. Bioelectron. 2008, 24, 912–916. [Google Scholar] [CrossRef]
- Cox, P.W.; Hooley, P. Hydrophobins: New prospects for biotechnology. Fungal Biol. Rev. 2009, 23, 40–47. [Google Scholar] [CrossRef]
- Cox, A.R.; Aldred, D.L.; Russell, A.B. Exceptional stability of food foams using class II hydrophobin HFBII. Food Hydrocoll. 2009, 23, 366–376. [Google Scholar] [CrossRef]
- Yamasaki, R.; Takatsuji, Y.; Asakawa, H.; Fukuma, T.; Haruyama, T. Flattened-top domical water drops formed through self-organization of hydrophobin membranes: A structural and mechanistic study using atomic force microscopy. ACS Nano 2016, 10, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Li, J.; Yang, H.; Shin, H.-J. Fungal and mushroom hydrophobins: A review. J. Mushroom 2017, 15, 1–7. [Google Scholar] [CrossRef]
- Wösten, H.A.B.; De Vocht, M.L. Hydrophobins, the fungal coat unravelled. Biochim. Biophys. Acta Rev. Biomembr. 2000, 1469, 79–86. [Google Scholar] [CrossRef]
- Whiteford, J.R.; Spanu, P.D. Hydrophobins and the interactions between fungi and plants. Mol. Plant Pathol. 2002, 3, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Gandier, J.-A.; Master, E. Pichia pastoris is a suitable host for the heterologous expression of predicted class I and class II hydrophobins for discovery, study, and application in biotechnology. Microorganisms 2018, 6, 3. [Google Scholar] [CrossRef]
- Pawłowska, B.K.; Sobieszczańska, B.M. Amyloids, common proteins among microorganisms. Postępy Mikrobiol. Adv. Microbiol. 2017, 56, 77–87. [Google Scholar]
- Ley, K.; Christofferson, A.; Penna, M.; Winkler, D.; Maclaughlin, S.; Yarovsky, I. Surface-water interface induces conformational changes critical for protein adsorption: Implications for monolayer formation of EAS hydrophobin. Front. Mol. Biosci. 2015, 2, 64. [Google Scholar] [CrossRef]
- Lo, V.; Ren, Q.; Pham, C.; Morris, V.; Kwan, A.; Sunde, M. Fungal hydrophobin proteins produce self-assembling protein films with diverse structure and chemical stability. Nanomaterials 2014, 4, 827–843. [Google Scholar] [CrossRef]
- Yu, L.; Zhang, B.; Szilvay, G.R.; Sun, R.; Jänis, J.; Wang, Z.; Feng, S.; Xu, H.; Linder, M.B.; Qiao, M. Protein HGFI from the edible mushroom Grifola frondosa is a novel 8 kDa class I hydrophobin that forms rodlets in compressed monolayers. Microbiology 2008, 154, 1677–1685. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Maeda, H.; Yoneda, S.; Ohtaki, S.; Yamagata, Y.; Hasegawa, F.; Gomi, K.; Nakajima, T.; Abe, K. The fungal hydrophobin RolA recruits polyesterase and laterally moves on hydrophobic surfaces. Mol. Microbiol. 2005, 57, 1780–1796. [Google Scholar] [CrossRef]
- Raja, H.A.; Miller, A.N.; Pearce, C.J.; Oberlies, N.H. Fungal identification using molecular tools: A primer for the natural products research community. J. Nat. Prod. 2017, 80, 756–770. [Google Scholar] [CrossRef] [PubMed]
- Vancov, T.; Keen, B. Amplification of soil fungal community DNA using the ITS86F and ITS4 primers. FEMS Microbiol. Lett. 2009, 296, 91–96. [Google Scholar] [CrossRef]
- Hrynkiewicz, K.; Baum, C.; Leinweber, P. Density, metabolic activity, and identity of cultivable rhizosphere bacteria on Salix viminalis in disturbed arable and landfill soils. J. Plant Nutr. Soil Sci. 2010, 173, 747–756. [Google Scholar] [CrossRef]
- Ogita, Z.; Markert, C.L. A miniaturized system for electrophoresis on polyacrylamide gels. Anal. Biochem. 1979, 99, 233–241. [Google Scholar] [CrossRef]
- Hammer, O.; Harper, D.A.T.; Ryan, P. PAST: Paleontological statistics software package for education and data analsis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Tg [°C] | Tc [°C] | ∆Hc [J/g] | Tm [°C] | ∆Hm [J/g] |
|---|---|---|---|---|---|
| PLA | 67.9 | 122.2 | −18.4 | 157.4 | 18.1 |
| PLA-medium | 67.0 | 115.9 | −28.8 | 157.8/151.2 | 30.5 |
| PLA-T. viride | 60.5 | 112.8 | −32.0 | 157.8/149.2 | 36.1 |
| PET | 80.8 | 137.8 | −26.8 | 248.9 | 37.5 |
| PET-medium | 77.9 | 138.0 | −30.8 | 249.1 | 46.9 |
| PET-T. viride | 77.8 | 139.4 | −31.6 | 249.1 | 47.8 |
| Sample | [η] | Mn (g/mol) |
|---|---|---|
| PLA | 1.1645 | 52,919 |
| PLA-medium | 1.0948 | 47,044 |
| PLA-T. viride | 0.8678 | 36,765 |
| PET | 0.7270 | 32,226 |
| PET-medium | 0.7153 | 31,518 |
| PET-T. viride | 0.6983 | 30,496 |
| Sample | Ra (nm) | Rq (nm) | Rmax (nm) |
|---|---|---|---|
| PLA | 2.89 ± 0.09 a | 3.48 ± 0.08 a | 37.0 ± 2.7 a |
| PLA-medium | 3.78 ± 0.11 a | 4.76 ± 0.07 a | 47.0 ± 4.2 a |
| PLA-T. viride | 8.19 ± 0.08 b | 13.60 ± 0.12 b | 157.0 ± 9.9 b |
| PET | 2.12 ± 0.11 a | 3.30 ± 0.09 a | 53.6 ± 5.5 a |
| PET-medium | 3.27 ± 0.20 a | 4.19 ± 0.15 a | 54.2 ± 8.1 a |
| PET-T. viride | 7.67 ± 0.18 b | 9.46 ± 0.11 b | 73.2 ± 4.6 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dąbrowska, G.B.; Garstecka, Z.; Olewnik-Kruszkowska, E.; Szczepańska, G.; Ostrowski, M.; Mierek-Adamska, A. Comparative Study of Structural Changes of Polylactide and Poly(ethylene terephthalate) in the Presence of Trichoderma viride. Int. J. Mol. Sci. 2021, 22, 3491. https://doi.org/10.3390/ijms22073491
Dąbrowska GB, Garstecka Z, Olewnik-Kruszkowska E, Szczepańska G, Ostrowski M, Mierek-Adamska A. Comparative Study of Structural Changes of Polylactide and Poly(ethylene terephthalate) in the Presence of Trichoderma viride. International Journal of Molecular Sciences. 2021; 22(7):3491. https://doi.org/10.3390/ijms22073491
Chicago/Turabian StyleDąbrowska, Grażyna B., Zuzanna Garstecka, Ewa Olewnik-Kruszkowska, Grażyna Szczepańska, Maciej Ostrowski, and Agnieszka Mierek-Adamska. 2021. "Comparative Study of Structural Changes of Polylactide and Poly(ethylene terephthalate) in the Presence of Trichoderma viride" International Journal of Molecular Sciences 22, no. 7: 3491. https://doi.org/10.3390/ijms22073491
APA StyleDąbrowska, G. B., Garstecka, Z., Olewnik-Kruszkowska, E., Szczepańska, G., Ostrowski, M., & Mierek-Adamska, A. (2021). Comparative Study of Structural Changes of Polylactide and Poly(ethylene terephthalate) in the Presence of Trichoderma viride. International Journal of Molecular Sciences, 22(7), 3491. https://doi.org/10.3390/ijms22073491