
Specialized Pro-Resolving Mediators and the Lymphatic System

Abstract
1. Introduction
2. The Lymphatic System
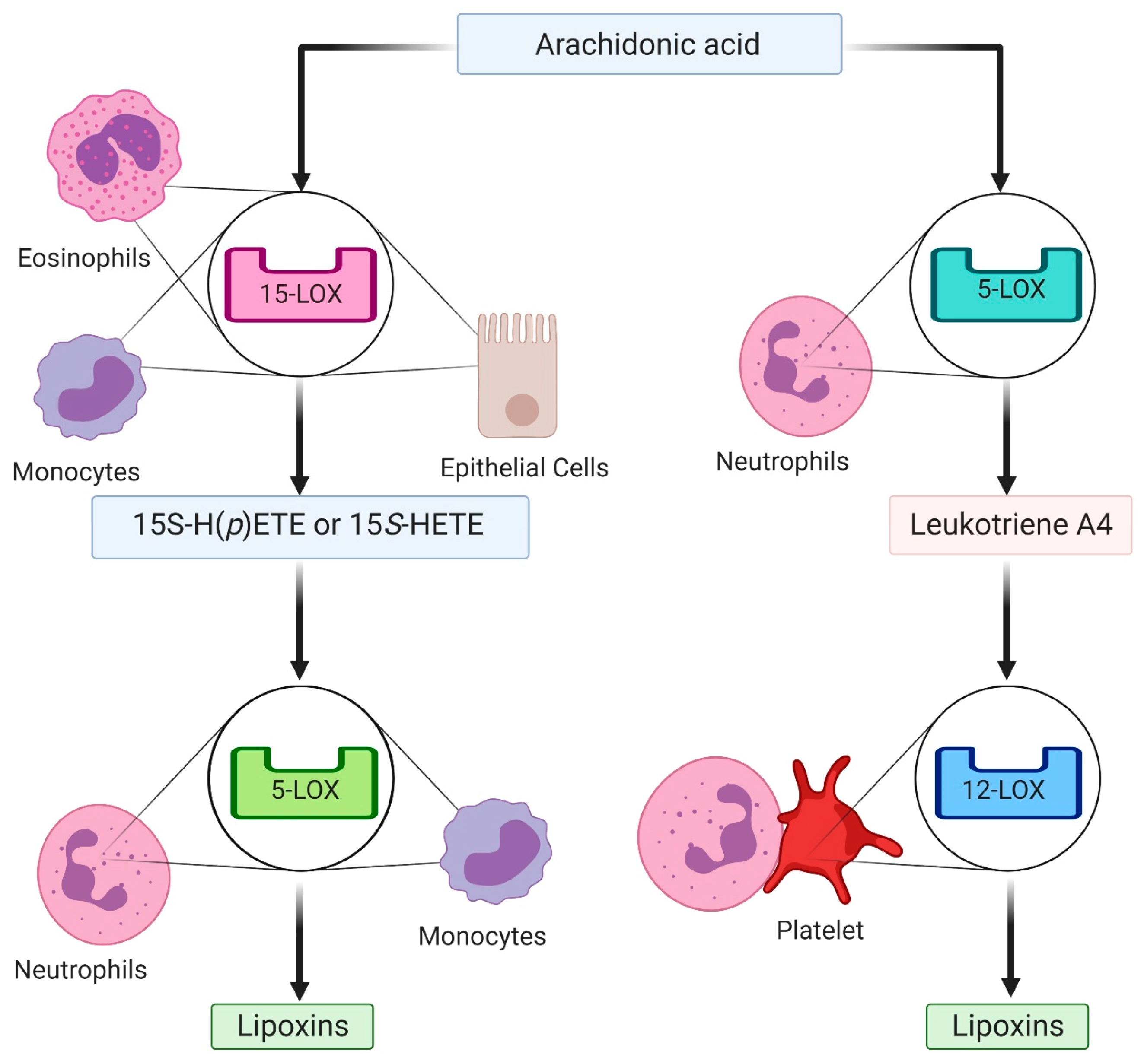
3. Specialized Pro-Resolving Mediators in Resolution of Inflammation
4. Lymphatics in Inflammatory Diseases
4.1. Cardiovascular Disease
4.2. Inflammatory Bowel Disease
4.3. Dry Eye Disease
5. Specialized Pro-Resolving Mediators and the Lymphatic System
5.1. Specialized Pro-Resolving Mediators in the Lymph Nodes and Spleen

{kind=link}
| Species | SPMs | Location/Disease | Sample Collection Time | Refs |
|---|---|---|---|---|
| Homo Sapiens | RvD1, RvD5, RvD6 and RvE3 | Axillary lymph nodes (from patients with Stage IV breast cancer, cardiorespiratory failure or craniocerebral injuries). | Post mortem | [107] |
| PGE2, LXA4 and LXB4 | Lymph nodes (from patients with Crohn’s disease, liver cirrhosis or heart failure). | Post mortem | ||
| RvD5, RvE1, RvE2, RvE3, PD1, 10S,17S-diHDHA, PGD2, PGE2, PGF2α, MaR2, 7S,14S-diHDHA and LXA4 | Spleens (from patients with sepsis along with leukaemia, heart failure or pneumonia). | Post mortem | ||
| African Green Monkey | LXA4 and LXB4, DHA-derived 7S,14S-diHDHA | Terminal ileum in Crohn’s disease model. Surgically induced lymphatic obstruction. | 7 days post-injury | [110] |
| D-series resolvins, protectins, and maresins | 61 days post-injury (significantly increased than day 7) | |||
| MaR1, 7S,14S-diHDHA, RvD5, RvD6, PDX, AT LXA4, LXB4, LTB4, and RvE2 | Colon in Crohn’s disease model. Surgically induced lymphatic obstruction. | 21 days post-injury |
5.2. Specialized Pro-Resolving Mediators in the Gastrointestinal Tract
5.3. Specialized Pro-Resolving Mediators in the Lungs
5.4. Specialized Pro-Resolving Mediators and the Eyes
5.5. Specialized Pro-Resolving Mediators and the Peritoneum
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schwager, S.; Detmar, M. Inflammation and lymphatic function. Front. Immunol. 2019, 10, 308. [Google Scholar] [CrossRef]
- Choi, I.; Lee, S.; Hong, Y.-K. The new era of the lymphatic system: No longer secondary to the blood vascular system. Cold Spring Harb. Perspect. Med. 2012, 2, a006445. [Google Scholar] [CrossRef]
- Aspelund, A.; Robciuc, M.R.; Karaman, S.; Makinen, T.; Alitalo, K. Lymphatic system in cardiovascular medicine. Circ. Res. 2016, 118, 515–530. [Google Scholar] [CrossRef] [PubMed]
- Kaipainen, A.; Korhonen, J.; Mustonen, T.; van Hinsbergh, V.W.; Fang, G.H.; Dumont, D.; Breitman, M.; Alitalo, K. Expression of the fms-like tyrosine kinase 4 gene becomes restricted to lymphatic endothelium during development. Proc. Natl. Acad. Sci. USA 1995, 92, 3566–3570. [Google Scholar] [CrossRef]
- Hirakawa, S.; Detmar, M.; Karaman, S. Lymphatics in nanophysiology. Adv. Drug Deliv. Rev. 2014, 74, 12–18. [Google Scholar] [CrossRef]
- Liao, S.; von der Weid, P.Y. Lymphatic system: An active pathway for immune protection. Semin. Cell Dev. Biol. 2015, 38, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Goyal, S.; Chauhan, S.K.; El Annan, J.; Nallasamy, N.; Zhang, Q.; Dana, R. Evidence of corneal lymphangiogenesis in dry eye disease: A potential link to adaptive immunity? Arch. Ophthalmol. 2010, 128, 819–824. [Google Scholar] [CrossRef] [PubMed]
- Mandal, R.V.; Mark, E.J.; Kradin, R.L. Organizing pneumonia and pulmonary lymphatic architecture in diffuse alveolar damage. Hum. Pathol. 2008, 39, 1234–1238. [Google Scholar] [CrossRef] [PubMed]
- Fullerton, J.N.; Gilroy, D.W. Resolution of inflammation: A new therapeutic frontier. Nat. Rev. Drug Discov. 2016, 15, 551–567. [Google Scholar] [CrossRef]
- Rosales, C. Neutrophil: A cell with many roles in inflammation or several cell types? Front. Physiol. 2018, 9, 113. [Google Scholar] [CrossRef]
- Li, Y.; Wang, W.; Yang, F.; Xu, Y.; Feng, C.; Zhao, Y. The regulatory roles of neutrophils in adaptive immunity. Cell Commun. Signal. 2019, 17, 147. [Google Scholar] [CrossRef] [PubMed]
- Headland, S.E.; Norling, L.V. The resolution of inflammation: Principles and challenges. Semin. Immunol. 2015, 27, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Godson, C.; Mitchell, S.; Harvey, K.; Petasis, N.A.; Hogg, N.; Brady, H.R. Cutting edge: Lipoxins rapidly stimulate nonphlogistic phagocytosis of apoptotic neutrophils by monocyte-derived macrophages. J. Immunol. 2000, 164, 1663–1667. [Google Scholar] [CrossRef]
- Schett, G.; Neurath, M.F. Resolution of chronic inflammatory disease: Universal and tissue-specific concepts. Nat. Commun. 2018, 9, 3261. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, T.; Gilroy, D.W. Chronic inflammation: A failure of resolution? Int. J. Exp. Pathol. 2007, 88, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Ungaro, F.; Rubbino, F.; Danese, S.; D’Alessio, S. Actors and factors in the resolution of intestinal inflammation: Lipid mediators as a new approach to therapy in inflammatory bowel diseases. Front. Immunol. 2017, 8, 1331. [Google Scholar] [CrossRef]
- Jiang, X.; Tian, W.; Nicolls, M.R.; Rockson, S.G. The lymphatic system in obesity, insulin resistance, and cardiovascular diseases. Front. Physiol. 2019, 10, 1402. [Google Scholar] [CrossRef]
- Stump, B.; Cui, Y.; Kidambi, P.; Lamattina, A.M.; El-Chemaly, S. Lymphatic changes in respiratory diseases: More than just remodeling of the lung? Am. J. Respir. Cell Mol. Biol. 2017, 57, 272–279. [Google Scholar] [CrossRef]
- Alexander, J.S.; Ganta, V.C.; Jordan, P.A.; Witte, M.H. Gastrointestinal lymphatics in health and disease. Pathophysiology 2010, 17, 315–335. [Google Scholar] [CrossRef]
- Al-Kofahi, M.; Yun, J.W.; Minagar, A.; Alexander, J.S. Anatomy and roles of lymphatics in inflammatory diseases. Clin. Exp. Neuroimmunol. 2017, 8, 199–214. [Google Scholar] [CrossRef]
- Huggenberger, R.; Siddiqui, S.S.; Brander, D.; Ullmann, S.; Zimmermann, K.; Antsiferova, M.; Werner, S.; Alitalo, K.; Detmar, M. An important role of lymphatic vessel activation in limiting acute inflammation. Blood 2011, 117, 4667–4678. [Google Scholar] [CrossRef]
- Cifarelli, V.; Eichmann, A. The intestinal lymphatic system: Functions and metabolic implications. Cell Mol. Gastroenterol. Hepatol. 2019, 7, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Dixon, J.B.; Raghunathan, S.; Swartz, M.A. A tissue-engineered model of the intestinal lacteal for evaluating lipid transport by lymphatics. Biotechnol. Bioeng. 2009, 103, 1224–1235. [Google Scholar] [CrossRef]
- Russo, E.; Nitschké, M.; Halin, C. Dendritic cell interactions with lymphatic endothelium. Lymphat Res. Biol. 2013, 11, 172–182. [Google Scholar] [CrossRef]
- Banerji, S.; Ni, J.; Wang, S.X.; Clasper, S.; Su, J.; Tammi, R.; Jones, M.; Jackson, D.G. LYVE-1, a new homologue of the CD44 glycoprotein, is a lymph-specific receptor for hyaluronan. J. Cell Biol. 1999, 144, 789–801. [Google Scholar] [CrossRef] [PubMed]
- Breiteneder-Geleff, S.; Soleiman, A.; Kowalski, H.; Horvat, R.; Amann, G.; Kriehuber, E.; Diem, K.; Weninger, W.; Tschachler, E.; Alitalo, K.; et al. Angiosarcomas express mixed endothelial phenotypes of blood and lymphatic capillaries: Podoplanin as a specific marker for lymphatic endothelium. Am. J. Pathol. 1999, 154, 385–394. [Google Scholar] [CrossRef]
- Wigle, J.T.; Oliver, G. Prox1 function is required for the development of the murine lymphatic system. Cell 1999, 98, 769–778. [Google Scholar] [CrossRef]
- Kukk, E.; Lymboussaki, A.; Taira, S.; Kaipainen, A.; Jeltsch, M.; Joukov, V.; Alitalo, K. VEGF-C receptor binding and pattern of expression with VEGFR-3 suggests a role in lymphatic vascular development. Dev. Camb. Engl. 1996, 122, 3829–3837. [Google Scholar]
- Jha, S.K.; Rauniyar, K.; Jeltsch, M. Key molecules in lymphatic development, function, and identification. Ann. Anat. 2018, 219, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Dumont, D.J.; Jussila, L.; Taipale, J.; Lymboussaki, A.; Mustonen, T.; Pajusola, K.; Breitman, M.; Alitalo, K. Cardiovascular failure in mouse embryos deficient in VEGF receptor-3. Science 1998, 282, 946–949. [Google Scholar] [CrossRef] [PubMed]
- Partanen, T.A.; Arola, J.; Saaristo, A.; Jussila, L.; Ora, A.; Miettinen, M.; Stacker, S.A.; Achen, M.G.; Alitalo, K. VEGF-C and VEGF-D expression in neuroendocrine cells and their receptor, VEGFR-3, in fenestrated blood vessels in human tissues. FASEB 2000, 14, 2087–2096. [Google Scholar] [CrossRef]
- Bouta, E.M.; Bell, R.D.; Rahimi, H.; Xing, L.; Wood, R.W.; Bingham, C.O., III; Ritchlin, C.T.; Schwarz, E.M. Targeting lymphatic function as a novel therapeutic intervention for rheumatoid arthritis. Nat. Rev. Rheumatol. 2018, 14, 94–106. [Google Scholar] [CrossRef]
- Alitalo, K. The lymphatic vasculature in disease. Nat. Med. 2011, 17, 1371–1380. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kataru, R.P.; Koh, G.Y. Regulation and implications of inflammatory lymphangiogenesis. Trends Immunol. 2012, 33, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kataru, R.P.; Koh, G.Y. Inflammation-associated lymphangiogenesis: A double-edged sword? J. Clin. Investig. 2014, 124, 936–942. [Google Scholar] [CrossRef] [PubMed]
- Karkkainen, M.J.; Haiko, P.; Sainio, K.; Partanen, J.; Taipale, J.; Petrova, T.V.; Jeltsch, M.; Jackson, D.G.; Talikka, M.; Rauvala, H.; et al. Vascular endothelial growth factor C is required for sprouting of the first lymphatic vessels from embryonic veins. Nat. Immunol. 2004, 5, 74–80. [Google Scholar] [CrossRef]
- Guo, R.; Zhou, Q.; Proulx, S.T.; Wood, R.; Ji, R.C.; Ritchlin, C.T.; Pytowski, B.; Zhu, Z.; Wang, Y.J.; Schwarz, E.M.; et al. Inhibition of lymphangiogenesis and lymphatic drainage via vascular endothelial growth factor receptor 3 blockade increases the severity of inflammation in a mouse model of chronic inflammatory arthritis. Arthritis Rheum. 2009, 60, 2666–2676. [Google Scholar] [CrossRef]
- Levick, J.R.; Michel, C.C. Microvascular fluid exchange and the revised Starling principle. Cardiovasc. Res. 2010, 87, 198–210. [Google Scholar] [CrossRef]
- Srinivasan, S.; Vannberg, F.O.; Dixon, J.B. Lymphatic transport of exosomes as a rapid route of information dissemination to the lymph node. Sci. Rep. 2016, 6, 24436. [Google Scholar] [CrossRef]
- Clement, C.C.; Rotzschke, O.; Santambrogio, L. The lymph as a pool of self-antigens. Trends Immunol. 2011, 32, 6–11. [Google Scholar] [CrossRef]
- Jackson, D.G. Leucocyte trafficking via the lymphatic vasculature—Mechanisms and consequences. Front. Immunol. 2019, 10, 471. [Google Scholar] [CrossRef]
- Voisin, M.B.; Nourshargh, S. Neutrophil trafficking to lymphoid tissues: Physiological and pathological implications. J. Pathol. 2019, 247, 662–671. [Google Scholar] [CrossRef] [PubMed]
- Hampton, H.R.; Chtanova, T. Lymphatic migration of immune cells. Front. Immunol. 2019, 10, 1168. [Google Scholar] [CrossRef]
- Card, C.M.; Yu, S.S.; Swartz, M.A. Emerging roles of lymphatic endothelium in regulating adaptive immunity. J. Clin. Investig. 2014, 124, 943–952. [Google Scholar] [CrossRef]
- Clement, C.C.; Cannizzo, E.S.; Nastke, M.-D.; Sahu, R.; Olszewski, W.; Miller, N.E.; Stern, L.J.; Santambrogio, L. An expanded self-antigen peptidome is carried by the human lymph as compared to the plasma. PLoS ONE 2010, 5, e9863. [Google Scholar] [CrossRef] [PubMed]
- Förster, R.; Davalos-Misslitz, A.C.; Rot, A. CCR7 and its ligands: Balancing immunity and tolerance. Nat. Rev. Immunol. 2008, 8, 362–371. [Google Scholar] [CrossRef]
- Piao, W.; Xiong, Y.; Famulski, K.; Brinkman, C.C.; Li, L.; Toney, N.; Wagner, C.; Saxena, V.; Simon, T.; Bromberg, J.S. Regulation of T cell afferent lymphatic migration by targeting LTβR-mediated non-classical NFκB signaling. Nat. Commun. 2018, 9, 3020. [Google Scholar] [CrossRef] [PubMed]
- Piao, W.; Xiong, Y.; Li, L.; Saxena, V.; Smith, K.D.; Hippen, K.L.; Paluskievicz, C.; Willsonshirkey, M.; Blazar, B.R.; Abdi, R.; et al. Regulatory T cells condition lymphatic endothelia for enhanced transendothelial migration. Cell Rep. 2020, 30, 1052–1062.e1055. [Google Scholar] [CrossRef]
- Schwab, J.M.; Chiang, N.; Arita, M.; Serhan, C.N. Resolvin E1 and protectin D1 activate inflammation-resolution programmes. Nature 2007, 447, 869–874. [Google Scholar] [CrossRef]
- Aspelund, A.; Antila, S.; Proulx, S.T.; Karlsen, T.V.; Karaman, S.; Detmar, M.; Wiig, H.; Alitalo, K. A dural lymphatic vascular system that drains brain interstitial fluid and macromolecules. J. Exp. Med. 2015, 212, 991–999. [Google Scholar] [CrossRef]
- Ahn, J.H.; Cho, H.; Kim, J.-H.; Kim, S.H.; Ham, J.-S.; Park, I.; Suh, S.H.; Hong, S.P.; Song, J.-H.; Hong, Y.-K.; et al. Meningeal lymphatic vessels at the skull base drain cerebrospinal fluid. Nature 2019, 572, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Louveau, A.; Smirnov, I.; Keyes, T.J.; Eccles, J.D.; Rouhani, S.J.; Peske, J.D.; Derecki, N.C.; Castle, D.; Mandell, J.W.; Lee, K.S.; et al. Structural and functional features of central nervous system lymphatic vessels. Nature 2015, 523, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Hsu, M.; Rayasam, A.; Kijak, J.A.; Choi, Y.H.; Harding, J.S.; Marcus, S.A.; Karpus, W.J.; Sandor, M.; Fabry, Z. Neuroinflammation-induced lymphangiogenesis near the cribriform plate contributes to drainage of CNS-derived antigens and immune cells. Nat. Commun. 2019, 10, 229. [Google Scholar] [CrossRef] [PubMed]
- Esposito, E.; Ahn, B.J.; Shi, J.; Nakamura, Y.; Park, J.H.; Mandeville, E.T.; Yu, Z.; Chan, S.J.; Desai, R.; Hayakawa, A.; et al. Brain-to-cervical lymph node signaling after stroke. Nat. Commun. 2019, 10, 5306. [Google Scholar] [CrossRef]
- Miao, Z.; Schultzberg, M.; Wang, X.; Zhao, Y. Role of polyunsaturated fatty acids in ischemic stroke—A perspective of specialized pro-resolving mediators. Clin. Nutr. 2021. [Google Scholar] [CrossRef]
- Serhan, C.N. Discovery of specialized pro-resolving mediators marks the dawn of resolution physiology and pharmacology. Mol. Asp. Med. 2017, 58, 1–11. [Google Scholar] [CrossRef]
- Serhan, C.N.; Hamberg, M.; Samuelsson, B. Trihydroxytetraenes: A novel series of compounds formed from arachidonic acid in human leukocytes. Biochem. Biophys. Res. Commun. 1984, 118, 943–949. [Google Scholar] [CrossRef]
- Serhan, C.N.; Savill, J. Resolution of inflammation: The beginning programs the end. Nat. Immunol. 2005, 6, 1191–1197. [Google Scholar] [CrossRef]
- Basil, M.C.; Levy, B.D. Specialized pro-resolving mediators: Endogenous regulators of infection and inflammation. Nat. Rev. Immunol. 2016, 16, 51–67. [Google Scholar] [CrossRef]
- Serhan, C.N.; Hamberg, M.; Samuelsson, B. Lipoxins: Novel series of biologically active compounds formed from arachidonic acid in human leukocytes. Proc. Natl. Acad. Sci. USA 1984, 81, 5335–5339. [Google Scholar] [CrossRef]
- Serhan, C.N.; Sheppard, K.A. Lipoxin formation during human neutrophil-platelet interactions. Evidence for the transformation of leukotriene A4 by platelet 12-lipoxygenase in vitro. J. Clin. Investig. 1990, 85, 772–780. [Google Scholar] [CrossRef]
- Serhan, C.N. Lipoxins and aspirin-triggered 15-epi-lipoxins are the first lipid mediators of endogenous anti-inflammation and resolution. ProstaglandinsLeukot. Essent. Fat. Acids 2005, 73, 141–162. [Google Scholar] [CrossRef]
- Serhan, C.N. On the relationship between leukotriene and lipoxin production by human neutrophils: Evidence for differential metabolism of 15-HETE and 5-HETE. Biochim. Et Biophys. Acta Mol. Cell Res. 1989, 1004, 158–168. [Google Scholar] [CrossRef]
- Brezinski, M.E.; Serhan, C.N. Selective incorporation of (15S)-hydroxyeicosatetraenoic acid in phosphatidylinositol of human neutrophils: Agonist-induced deacylation and transformation of stored hydroxyeicosanoids. Proc. Natl. Acad. Sci. USA 1990, 87, 6248–6252. [Google Scholar] [CrossRef] [PubMed]
- Takano, T.; Fiore, S.; Maddox, J.F.; Brady, H.R.; Petasis, N.A.; Serhan, C.N. Aspirin-triggered 15-epi-lipoxin A4 (LXA4) and LXA4 stable analogues are potent inhibitors of acute inflammation: Evidence for anti-inflammatory receptors. J. Exp. Med. 1997, 185, 1693–1704. [Google Scholar] [CrossRef]
- McMahon, B.; Godson, C. Lipoxins: Endogenous regulators of inflammation. Am. J. Physiol. Ren. Physiol. 2004, 286, F189–F201. [Google Scholar] [CrossRef]
- Shinohara, M.; Serhan, C.N. Novel endogenous proresolving molecules: Essential fatty acid-derived and gaseous mediators in the resolution of inflammation. J. Atheroscler. Thromb. 2016, 23, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Borgeson, E.; Johnson, A.M.; Lee, Y.S.; Till, A.; Syed, G.H.; Ali-Shah, S.T.; Guiry, P.J.; Dalli, J.; Colas, R.A.; Serhan, C.N.; et al. Lipoxin A4 attenuates obesity-induced adipose inflammation and associated liver and kidney disease. Cell Metab. 2015, 22, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Clish, C.B.; Brannon, J.; Colgan, S.P.; Chiang, N.; Gronert, K. Novel functional sets of lipid-derived mediators with antiinflammatory actions generated from omega-3 fatty acids via cyclooxygenase 2-nonsteroidal antiinflammatory drugs and transcellular processing. J. Exp. Med. 2000, 192, 1197–1204. [Google Scholar] [CrossRef] [PubMed]
- Arita, M.; Bianchini, F.; Aliberti, J.; Sher, A.; Chiang, N.; Hong, S.; Yang, R.; Petasis, N.A.; Serhan, C.N. Stereochemical assignment, antiinflammatory properties, and receptor for the omega-3 lipid mediator resolvin E1. J. Exp. Med. 2005, 201, 713–722. [Google Scholar] [CrossRef]
- Campbell, E.L.; Louis, N.A.; Tomassetti, S.E.; Canny, G.O.; Arita, M.; Serhan, C.N.; Colgan, S.P. Resolvin E1 promotes mucosal surface clearance of neutrophils: A new paradigm for inflammatory resolution. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2007, 21, 3162–3170. [Google Scholar] [CrossRef]
- Tani, Y.; Isobe, Y.; Imoto, Y.; Segi-Nishida, E.; Sugimoto, Y.; Arai, H.; Arita, M. Eosinophils control the resolution of inflammation and draining lymph node hypertrophy through the proresolving mediators and CXCL13 pathway in mice. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2014, 28, 4036–4043. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Porter, T.F.; Lu, Y.; Oh, S.F.; Pillai, P.S.; Serhan, C.N. Resolvin E1 metabolome in local inactivation during inflammation-resolution. J. Immunol. 2008, 180, 3512. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.F.; Pillai, P.S.; Recchiuti, A.; Yang, R.; Serhan, C.N. Pro-resolving actions and stereoselective biosynthesis of 18S E-series resolvins in human leukocytes and murine inflammation. J. Clin. Investig. 2011, 121, 569–581. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Chiang, N.; Dalli, J.; Levy, B.D. Lipid mediators in the resolution of inflammation. Cold Spring Harb. Perspect. Biol. 2014, 7, a016311. [Google Scholar] [CrossRef]
- Dalli, J.; Winkler, J.W.; Colas, R.A.; Arnardottir, H.; Cheng, C.Y.; Chiang, N.; Petasis, N.A.; Serhan, C.N. Resolvin D3 and aspirin-triggered resolvin D3 are potent immunoresolvents. Chem. Biol. 2013, 20, 188–201. [Google Scholar] [CrossRef]
- Ariel, A.; Li, P.L.; Wang, W.; Tang, W.X.; Fredman, G.; Hong, S.; Gotlinger, K.H.; Serhan, C.N. The docosatriene protectin D1 is produced by TH2 skewing and promotes human T cell apoptosis via lipid raft clustering. J. Biol. Chem. 2005, 280, 43079–43086. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Gronert, K. The role of pro-resolving lipid mediators in ocular diseases. Mol. Asp. Med. 2017, 58, 37–43. [Google Scholar] [CrossRef]
- Sugimoto, M.A.; Sousa, L.P.; Pinho, V.; Perretti, M.; Teixeira, M.M. Resolution of inflammation: What controls its onset? Front. Immunol. 2016, 7, 160. [Google Scholar] [CrossRef]
- Linton, M.F.; Yancey, P.G.; Davies, S.S.; Jerome, W.G.; Linton, E.F.; Song, W.L.; Doran, A.C.; Vickers, K.C. Endotext—The Role of Lipids and Lipoproteins in Atherosclerosis; MDText.com, Inc.: South Dartmouth, MA, USA, 2019. [Google Scholar]
- Martel, C.; Li, W.; Fulp, B.; Platt, A.M.; Gautier, E.L.; Westerterp, M.; Bittman, R.; Tall, A.R.; Chen, S.H.; Thomas, M.J.; et al. Lymphatic vasculature mediates macrophage reverse cholesterol transport in mice. J. Clin. Investig. 2013, 123, 1571–1579. [Google Scholar] [CrossRef]
- Bergström, I.G. Pro-and Anti-Inflammatory Actions in Coronary Artery Dsease with Focus on CD 56 + T Cells and Annexin A 1; Linköping University Electronic Press: Linköping, Sweden, 2015. [Google Scholar]
- Luk, E.; Gotlieb, A.I. Atherosclerosis. In Pathobiology of Human Disease; McManus, L.M., Mitchell, R.N., Eds.; Academic Press: San Diego, CA, USA, 2014; pp. 2970–2985. [Google Scholar] [CrossRef]
- Khera, A.V.; Cuchel, M.; de la Llera-Moya, M.; Rodrigues, A.; Burke, M.F.; Jafri, K.; French, B.C.; Phillips, J.A.; Mucksavage, M.L.; Wilensky, R.L.; et al. Cholesterol efflux capacity, high-density lipoprotein function, and atherosclerosis. N. Engl. J. Med. 2011, 364, 127–135. [Google Scholar] [CrossRef]
- Lim, H.Y.; Thiam, C.H.; Yeo, K.P.; Bisoendial, R.; Hii, C.S.; McGrath, K.C.; Tan, K.W.; Heather, A.; Alexander, J.S.; Angeli, V. Lymphatic vessels are essential for the removal of cholesterol from peripheral tissues by SR-BI-mediated transport of HDL. Cell Metab. 2013, 17, 671–684. [Google Scholar] [CrossRef] [PubMed]
- Milasan, A.; Smaani, A.; Martel, C. Early rescue of lymphatic function limits atherosclerosis progression in Ldlr(−/−) mice. Atherosclerosis 2019, 283, 106–119. [Google Scholar] [CrossRef] [PubMed]
- Vieira, J.M.; Norman, S.; Villa Del Campo, C.; Cahill, T.J.; Barnette, D.N.; Gunadasa-Rohling, M.; Johnson, L.A.; Greaves, D.R.; Carr, C.A.; Jackson, D.G.; et al. The cardiac lymphatic system stimulates resolution of inflammation following myocardial infarction. J. Clin. Investig. 2018, 128, 3402–3412. [Google Scholar] [CrossRef]
- Henri, O.; Pouehe, C.; Houssari, M.; Galas, L.; Nicol, L.; Edwards-Lévy, F.; Henry, J.P.; Dumesnil, A.; Boukhalfa, I.; Banquet, S.; et al. Selective stimulation of cardiac lymphangiogenesis reduces myocardial edema and fibrosis leading to improved cardiac function following myocardial infarction. Circulation 2016, 133, 1484–1497. [Google Scholar] [CrossRef] [PubMed]
- Vuorio, T.; Ylä-Herttuala, E.; Laakkonen, J.P.; Laidinen, S.; Liimatainen, T.; Ylä-Herttuala, S. Downregulation of VEGFR3 signaling alters cardiac lymphatic vessel organization and leads to a higher mortality after acute myocardial infarction. Sci. Rep. 2018, 8, 16709. [Google Scholar] [CrossRef]
- Brakenhielm, E.; Alitalo, K. Cardiac lymphatics in health and disease. Nat. Rev. Cardiol. 2019, 16, 56–68. [Google Scholar] [CrossRef]
- Geleff, S.; Schoppmann, S.F.; Oberhuber, G. Increase in podoplanin-expressing intestinal lymphatic vessels in inflammatory bowel disease. Virchows Arch. Int. J. Pathol. 2003, 442, 231–237. [Google Scholar] [CrossRef]
- Pedica, F.; Ligorio, C.; Tonelli, P.; Bartolini, S.; Baccarini, P. Lymphangiogenesis in Crohn’s disease: An immunohistochemical study using monoclonal antibody D2-40. Virchows Arch. 2008, 452, 57–63. [Google Scholar] [CrossRef]
- Tonelli, F.; Giudici, F.; Liscia, G. Is lymphatic status related to regression of inflammation in Crohn’s disease? World J Gastrointest Surg 2012, 4, 228–233. [Google Scholar] [CrossRef][Green Version]
- Rahier, J.F.; Dubuquoy, L.; Colombel, J.F.; Jouret-Mourin, A.; Delos, M.; Ferrante, M.; Sokol, H.; Hertogh, G.D.; Salleron, J.; Geboes, K.; et al. Decreased lymphatic vessel density is associated with postoperative endoscopic recurrence in Crohn’s disease. Inflamm. Bowel Dis. 2013, 19, 2084–2090. [Google Scholar] [CrossRef] [PubMed]
- Kühn, R.; Löhler, J.; Rennick, D.; Rajewsky, K.; Müller, W. Interleukin-10-deficient mice develop chronic enterocolitis. Cell 1993, 75, 263–274. [Google Scholar] [CrossRef]
- Spencer, D.M.; Veldman, G.M.; Banerjee, S.; Willis, J.; Levine, A.D. Distinct inflammatory mechanisms mediate early versus late colitis in mice. Gastroenterology 2002, 122, 94–105. [Google Scholar] [CrossRef] [PubMed]
- D’Alessio, S.; Correale, C.; Tacconi, C.; Gandelli, A.; Pietrogrande, G.; Vetrano, S.; Genua, M.; Arena, V.; Spinelli, A.; Peyrin-Biroulet, L.; et al. VEGF-C-dependent stimulation of lymphatic function ameliorates experimental inflammatory bowel disease. J. Clin. Investig. 2014, 124, 3863–3878. [Google Scholar] [CrossRef]
- Jurisic, G.; Sundberg, J.P.; Detmar, M. Blockade of VEGF receptor-3 aggravates inflammatory bowel disease and lymphatic vessel enlargement. Inflamm. Bowel Dis. 2013, 19, 1983–1989. [Google Scholar] [CrossRef] [PubMed]
- Pflugfelder, S.C.; de Paiva, C.S. The Pathophysiology of Dry Eye Disease: What We Know and Future Directions for Research. Ophthalmology 2017, 124, S4–S13. [Google Scholar] [CrossRef]
- Plsková, J.; Holán, V.; Filipec, M.; Forrester, J.V. Lymph node removal enhances corneal graft survival in mice at high risk of rejection. Bmc Ophthalmol. 2004, 4, 3. [Google Scholar] [CrossRef]
- Serhan, C.N. Treating inflammation and infection in the 21st century: New hints from decoding resolution mediators and mechanisms. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2017, 31, 1273–1288. [Google Scholar] [CrossRef]
- Colby, J.K.; Abdulnour, R.E.; Sham, H.P.; Dalli, J.; Colas, R.A.; Winkler, J.W.; Hellmann, J.; Wong, B.; Cui, Y.; El-Chemaly, S.; et al. Resolvin D3 and aspirin-triggered resolvin D3 are protective for injured epithelia. Am. J. Pathol. 2016, 186, 1801–1813. [Google Scholar] [CrossRef]
- Jin, Y.; Arita, M.; Zhang, Q.; Saban, D.R.; Chauhan, S.K.; Chiang, N.; Serhan, C.N.; Dana, R. Anti-angiogenesis effect of the novel anti-inflammatory and pro-resolving lipid mediators. Investig. Ophthalmol. Vis. Sci. 2009, 50, 4743–4752. [Google Scholar] [CrossRef]
- Kain, V.; Ingle, K.A.; Colas, R.A.; Dalli, J.; Prabhu, S.D.; Serhan, C.N.; Joshi, M.; Halade, G.V. Resolvin D1 activates the inflammation resolving response at splenic and ventricular site following myocardial infarction leading to improved ventricular function. J. Mol. Cell Cardiol. 2015, 84, 24–35. [Google Scholar] [CrossRef]
- Gao, Y.; Su, J.; Zhang, Y.; Chan, A.; Sin, J.H.; Wu, D.; Min, K.; Gronert, K. Dietary DHA amplifies LXA(4) circuits in tissues and lymph node PMN and is protective in immune-driven dry eye disease. Mucosal. Immunol. 2018, 11, 1674–1683. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Tani, Y.; Nakanishi, H.; Taguchi, R.; Arita, M.; Arai, H. Eosinophils promote resolution of acute peritonitis by producing proresolving mediators in mice. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2011, 25, 561–568. [Google Scholar] [CrossRef]
- Colas, R.A.; Shinohara, M.; Dalli, J.; Chiang, N.; Serhan, C.N. Identification and signature profiles for pro-resolving and inflammatory lipid mediators in human tissue. Am. J. Physiol. Cell Physiol. 2014, 307, C39–C54. [Google Scholar] [CrossRef]
- Frolov, A.; Yang, L.; Dong, H.; Hammock, B.D.; Crofford, L.J. Anti-inflammatory properties of prostaglandin E2: Deletion of microsomal prostaglandin E synthase-1 exacerbates non-immune inflammatory arthritis in mice. ProstaglandinsLeukot. Essent. Fat. Acids 2013, 89, 351–358. [Google Scholar] [CrossRef]
- Halade, G.V.; Norris, P.C.; Kain, V.; Serhan, C.N.; Ingle, K.A. Splenic leukocytes define the resolution of inflammation in heart failure. Sci. Signal 2018, 11, eaao1818. [Google Scholar] [CrossRef] [PubMed]
- Becker, F.; Romero, E.; Goetzmann, J.; Hasselschwert, D.L.; Dray, B.; Vanchiere, J.; Fontenot, J.; Yun, J.W.; Norris, P.C.; White, L.; et al. Endogenous specialized proresolving mediator profiles in a novel experimental model of lymphatic obstruction and intestinal inflammation in african green monkeys. Am. J. Pathol. 2019, 189, 1953–1972. [Google Scholar] [CrossRef]
- Onali, S.; Favale, A.; Fantini, M.C. The resolution of intestinal inflammation: The peace-keeper’s perspective. Cells 2019, 8, 344. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.X.; Colgan, S.P. Special pro-resolving mediator (SPM) actions in regulating gastro-intestinal inflammation and gut mucosal immune responses. Mol. Asp. Med. 2017, 58, 93–101. [Google Scholar] [CrossRef]
- Newson, J.; Stables, M.; Karra, E.; Arce-Vargas, F.; Quezada, S.; Motwani, M.; Mack, M.; Yona, S.; Audzevich, T.; Gilroy, D.W. Resolution of acute inflammation bridges the gap between innate and adaptive immunity. Blood 2014, 124, 1748–1764. [Google Scholar] [CrossRef] [PubMed]
- El-Chemaly, S.; Levine, S.J.; Moss, J. Lymphatics in lung disease. Ann. N. Y. Acad. Sci. 2008, 1131, 195–202. [Google Scholar] [CrossRef]
- Baluk, P.; Tammela, T.; Ator, E.; Lyubynska, N.; Achen, M.G.; Hicklin, D.J.; Jeltsch, M.; Petrova, T.V.; Pytowski, B.; Stacker, S.A.; et al. Pathogenesis of persistent lymphatic vessel hyperplasia in chronic airway inflammation. J. Clin. Investig. 2005, 115, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Mori, M.; Andersson, C.K.; Graham, G.J.; Löfdahl, C.G.; Erjefält, J.S. Increased number and altered phenotype of lymphatic vessels in peripheral lung compartments of patients with COPD. Respir. Res. 2013, 14, 65. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Min, K.; Zhang, Y.; Su, J.; Greenwood, M.; Gronert, K. Female-specific downregulation of tissue polymorphonuclear neutrophils drives impaired regulatory T cell and amplified effector T cell responses in autoimmune dry eye disease. J. Immunol. 2015, 195, 3086–3099. [Google Scholar] [CrossRef]
- Chiang, N.; Dalli, J.; Colas, R.A.; Serhan, C.N. Identification of resolvin D2 receptor mediating resolution of infections and organ protection. J. Exp. Med. 2015, 212, 1203–1217. [Google Scholar] [CrossRef]
- Gilroy, D.W.; Colville-Nash, P.R.; Willis, D.; Chivers, J.; Paul-Clark, M.J.; Willoughby, D.A. Inducible cyclooxygenase may have anti-inflammatory properties. Nat. Med. 1999, 5, 698–701. [Google Scholar] [CrossRef] [PubMed]

| SPMs | Disease Model | Species | Molecular Effect | Refs |
|---|---|---|---|---|
| AT-RvD3 and RvD3 | Acute respiratory distress syndrome | Mouse—injection of hydrochloric acid into the mainstem bronchus | Decreased oedema, alveolar wall thickening, haemorrhage and leukocyte influx | [102] |
| Significantly increased LYVE-1+ vessels and increase VEGFR3+ vessels post injury | ||||
| AT-LXA4 | Corneal neovascularization | Mouse—suture or micropellet (containing IL-1β & VEGF-A) | Inhibited angiogenesis but not lymphangiogenesis. No effect on lymphangiogenic VEGF-D or VEGFR-3, reduction of neutrophils post injury | [103] |
| Peritonitis | Mouse—zymosan induced | Reduced PMN infiltration. Doubled the amount of zymosan-carrying leukocytes in the draining lymph nodes and a significant increase in the spleen | [49] | |
| RvD1 | Corneal neovascularization | Mouse—suture or micropellet (containing IL-1β & VEGF-A) | No effect on lymphangiogenesis or expression of lymphangiogenic VEGF-Dor VEGFR-3. Decreased neutrophil count post injury | [103] |
| Post-myocardial infarction | Mouse—surgical ligation of the left anterior descending coronary artery | Conserved transient changes in splenic weight following MI. Reduced pulmonary oedema. Significant increase neutrophils present in spleen and rapid clearance. Altered production of SPMs in spleen. Induced rapid clearance of macrophages | [104] | |
| RvE1 | Corneal neovascularization | Mouse—suture or micropellet (containing IL-1β & VEGF-A) | Prevented angiogenesis. Did not alter lymphangiogenesis or VEGF-D or VEGFR-3 expression. Reduced neutrophils post-injury. | [103] |
| Peritonitis | Mouse—Zymosan induced | Increased mononuclear cell infiltration, Increased zymosan-carrying leukocytes in the spleen | [49] | |
| DHA supplement (diet enriched for 40% DHA) | Dry eye disease | Mouse—desiccating stress (combination of continuous airflow, low humidity and inhibiting lacrimal gland function) | Increased LXA4 levels in the draining lymph nodes, increased LXA4 production from lymph node-derived regulatory PMN | [105] |
| PD1 | Peritonitis | Mouse—zymosan induced | Rescued impaired lymphatic drainage in eosinophil-depleted mice. Reduced PMN infiltration. Increased zymosan-carrying leukocyte to the spleen 4-fold and 6-fold in lymph nodes | [106] |
| LXA4 | N/A | Mouse—eosinophil-depleted mice | Rescue impaired lymph node hypertrophy | [72] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kraft, J.D.; Blomgran, R.; Lundgaard, I.; Quiding-Järbrink, M.; Bromberg, J.S.; Börgeson, E. Specialized Pro-Resolving Mediators and the Lymphatic System. Int. J. Mol. Sci. 2021, 22, 2750. https://doi.org/10.3390/ijms22052750
Kraft JD, Blomgran R, Lundgaard I, Quiding-Järbrink M, Bromberg JS, Börgeson E. Specialized Pro-Resolving Mediators and the Lymphatic System. International Journal of Molecular Sciences. 2021; 22(5):2750. https://doi.org/10.3390/ijms22052750
Chicago/Turabian StyleKraft, Jamie D., Robert Blomgran, Iben Lundgaard, Marianne Quiding-Järbrink, Jonathan S. Bromberg, and Emma Börgeson. 2021. "Specialized Pro-Resolving Mediators and the Lymphatic System" International Journal of Molecular Sciences 22, no. 5: 2750. https://doi.org/10.3390/ijms22052750
APA StyleKraft, J. D., Blomgran, R., Lundgaard, I., Quiding-Järbrink, M., Bromberg, J. S., & Börgeson, E. (2021). Specialized Pro-Resolving Mediators and the Lymphatic System. International Journal of Molecular Sciences, 22(5), 2750. https://doi.org/10.3390/ijms22052750