
Different miRNA Profiles in Plasma Derived Small and Large Extracellular Vesicles from Patients with Neurodegenerative Diseases

 , , , , , ,
, , , , , ,  and add
Show full author list
and add
Show full author list
Abstract
1. Introduction
2. Results
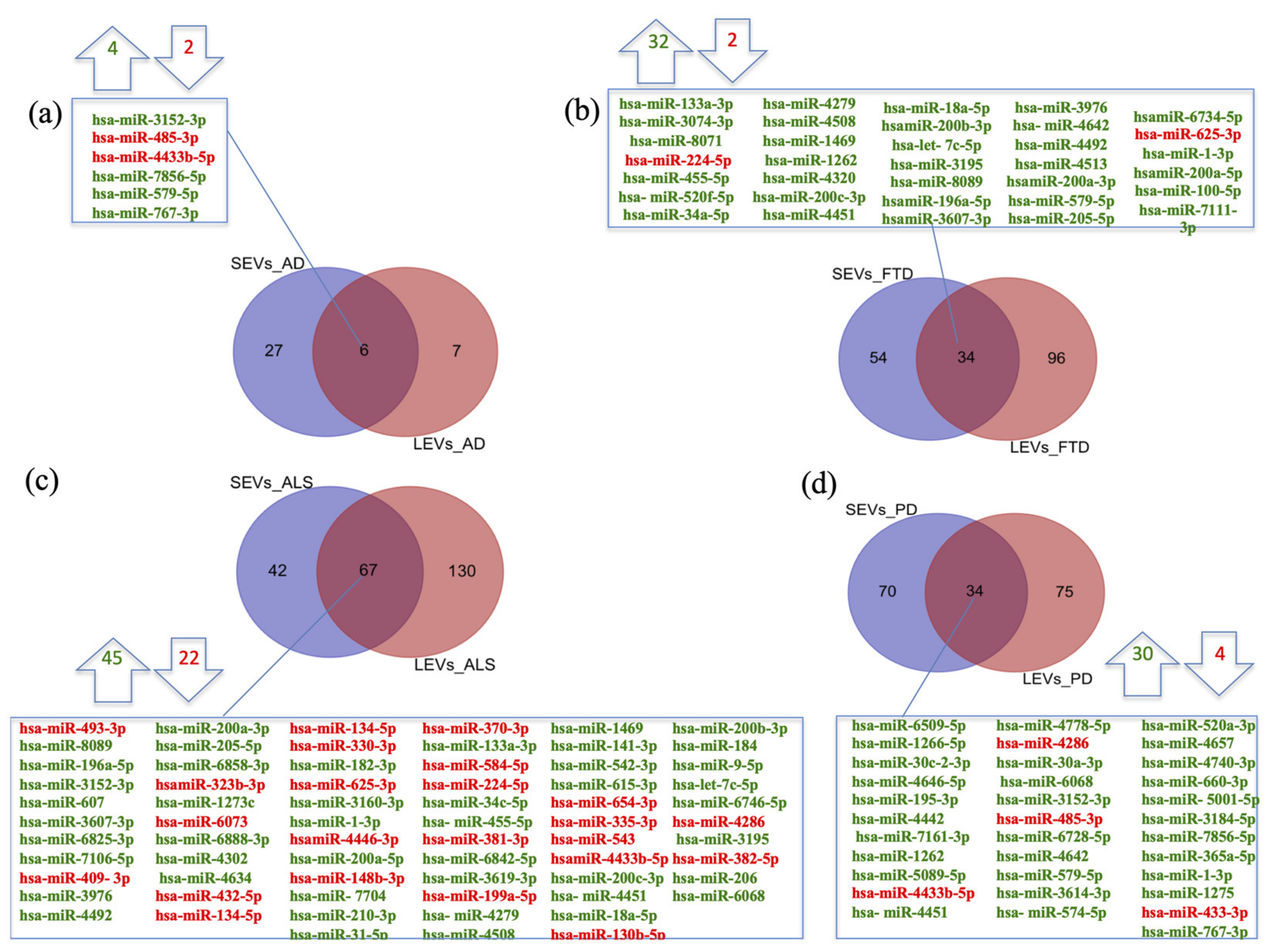
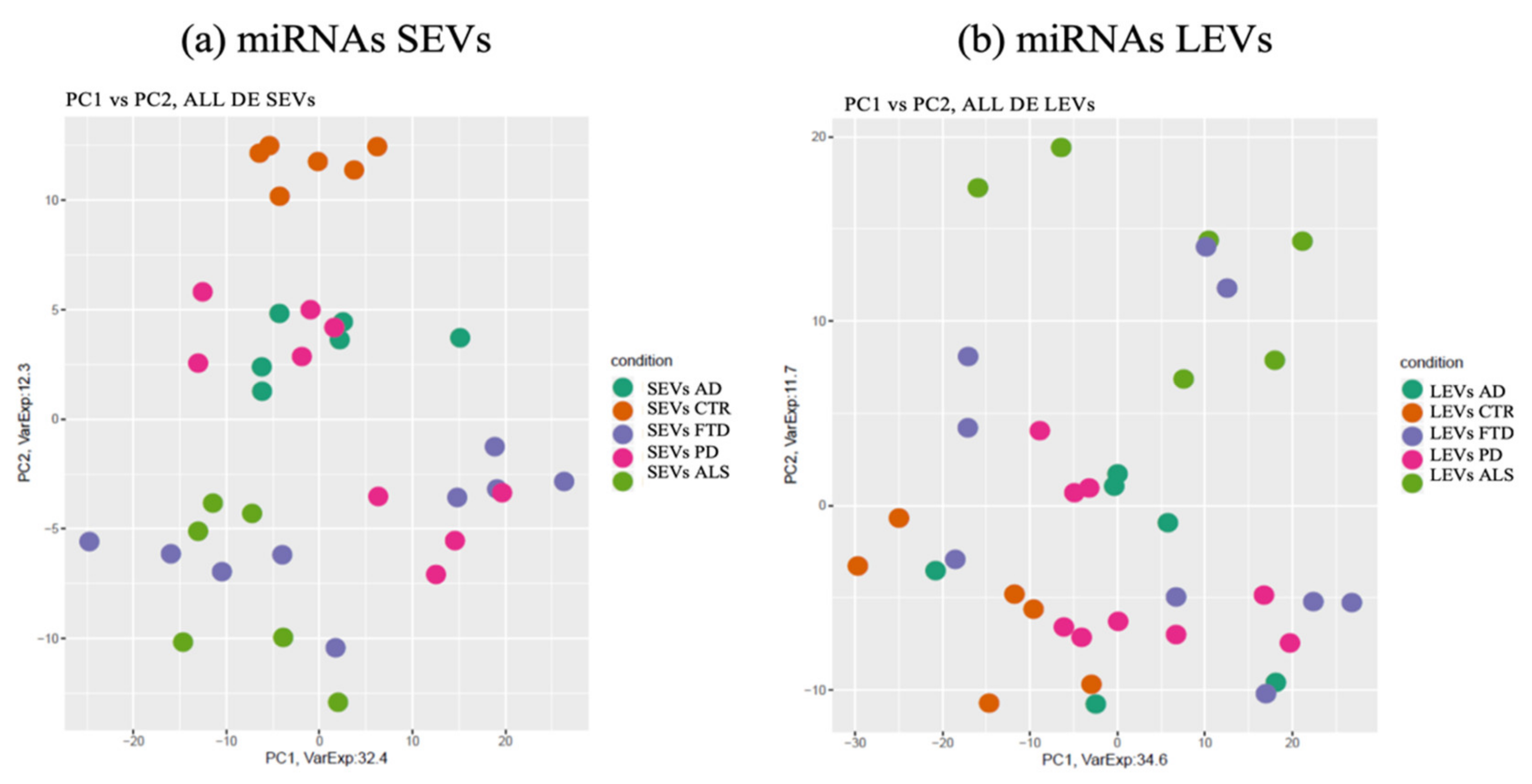
2.1. miRNAs Selectively Traffic into SEVs and LEVs
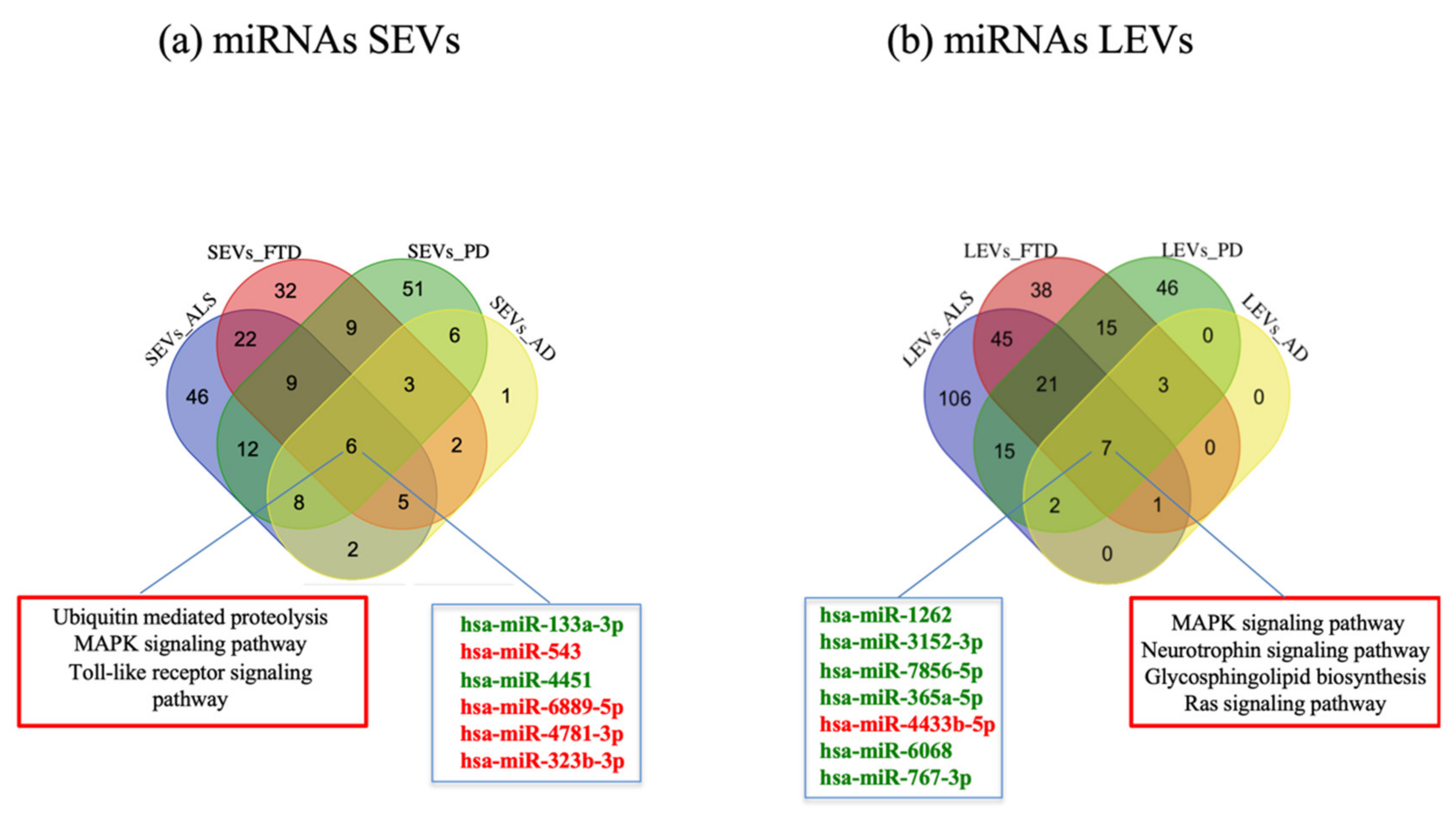
2.2. miRNAs Expression Profiles and Common Pathways in SEVs and LEVs of NDs
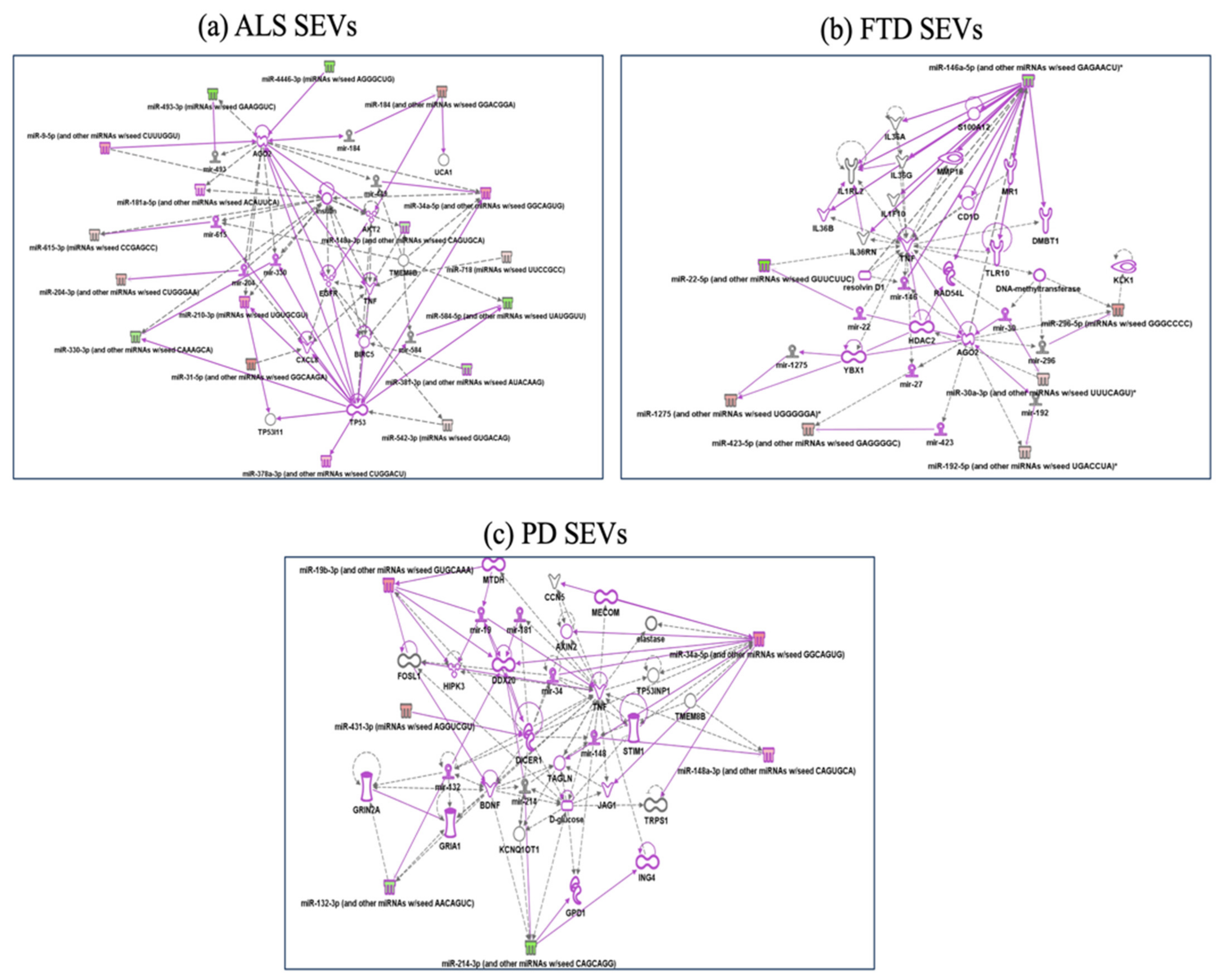
2.3. Specific miRNAs Pathway Analysis in SEVs of NDs
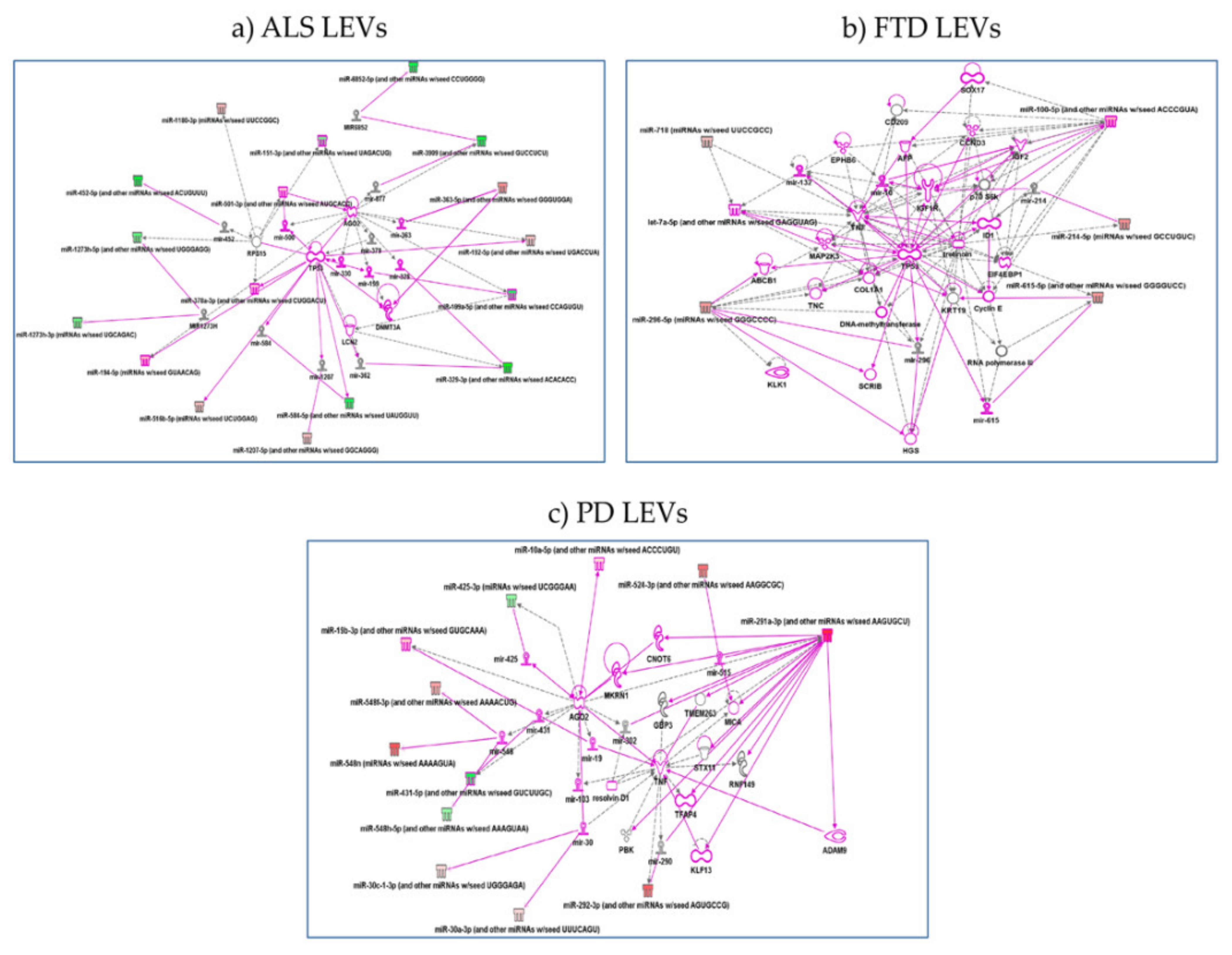
2.4. Specific miRNAs Pathway Analysis in LEVs of NDs
3. Discussion
4. Materials and Methods
4.1. Study Subjects
4.2. LEVs and SEVs Isolation
4.3. RNA Extraction
4.4. RNA Libraries Preparations
4.5. Bioinformatic Data Analysis
4.6. Pathways Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s disease |
| ALS | Amyotrophic Lateral Sclerosis |
| CTRs | healthy controls |
| DE miRNAs | differentially expressed miRNAs |
| ESCRT | Endosomal Sorting Complexes Required for Transport |
| EVs | extracellular vesicles |
| EXOs | exosomes |
| FTD | frontotemporal dementia |
| GWAS | Genome-wide association studies |
| ILVs | intraluminal vesicles |
| IPA | Ingenuity pathway analysis |
| LBs | Lewy bodies |
| LEVs | large extracellular vesicles |
| lncRNA | long non coding RNA |
| miRNA | microRNA |
| MVs | microvesicles |
| NDs | neurodegenerative diseases |
| PCA | principal component analysis |
| PD | Parkinson’s disease |
| RBPs | RNA-binding proteins |
| SEVs | small extracellular vesicles |
| SN | substantia nigra |
| SNPs | single nucleotide polymorphisms |
| TLRs | Toll like receptors |
References
- Gitler, A.D.; Dhillon, P.; Shorter, J. Neurodegenerative disease: Models, mechanisms, and a new hope. Dis. Model. Mech. 2017, 10, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Habib, R.; Noureen, N.; Nadeem, N. Decoding Common Features of Neurodegenerative Disorders: From Differentially Expressed Genes to Pathways. Curr. Genom. 2018, 19, 300–312. [Google Scholar] [CrossRef] [PubMed]
- Montie, H.L.; Durcan, T.M. The Cell and Molecular Biology of Neurodegenerative Diseases: An Overview. Front. Neurol. 2013, 4, 194. [Google Scholar] [CrossRef] [PubMed]
- García, J.C.; Bustos, R.H. The Genetic Diagnosis of Neurodegenerative Diseases and Therapeutic Perspectives. Brain Sci. 2018, 8, 222. [Google Scholar] [CrossRef] [PubMed]
- Obeso, J.A.; Stamelou, M.; Goetz, C.G.; Poewe, W.; Lang, A.E.; Weintraub, D.; Burn, D.; Halliday, G.M.; Bezard, E. Past, present, and future of Parkinson’s disease: A special essay on the 200th Anniversary of the Shaking Palsy. Mov. Disord. 2017, 32, 1264–1310. [Google Scholar] [CrossRef]
- Cereda, C.; Leoni, E.; Milani, P.; Pansarasa, O.; Mazzini, G.; Guareschi, S.; Alvisi, E.; Ghiroldi, A.; Diamanti, L.; Bernuzzi, S.; et al. Altered intracellular localization of SOD1 in leukocytes from patients with sporadic amyotrophic lateral sclerosis. PLoS ONE 2013, 8, e75916. [Google Scholar] [CrossRef]
- Guareschi, S.; Cova, E.; Cereda, C.; Ceroni, M.; Donetti, E.; Bosco, D.A.; Trotti, D.; Pasinelli, P. An over-oxidized form of superoxide dismutase found in sporadic amyotrophic lateral sclerosis with bulbar onset shares a toxic mechanism with mutant SOD1. Proc. Natl. Acad. Sci. USA 2012, 109, 5074–5079. [Google Scholar] [CrossRef] [PubMed]
- Pansarasa, O.; Bordoni, M.; Diamanti, L.; Sproviero, D.; Gagliardi, S.; Cereda, C. SOD1 in Amyotrophic Lateral Sclerosis: “Ambivalent” Behavior Connected to the Disease. Int. J. Mol. Sci. 2018, 19, 1345. [Google Scholar] [CrossRef]
- Gagliardi, S.; Cova, E.; Davin, A.; Guareschi, S.; Abel, K.; Alvisi, E.; Laforenza, U.; Ghidoni, R.; Cashman, J.R.; Ceroni, M.; et al. SOD1 mRNA expression in sporadic amyotrophic lateral sclerosis. Neurobiol. Dis. 2010, 39, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Fogh, I.; Lin, K.; Tiloca, C.; Rooney, J.; Gellera, C.; Diekstra, F.P.; Ratti, A.; Shatunov, A.; Van Es, M.A.; Proitsi, P.; et al. Association of a Locus in theCAMTA1Gene With Survival in Patients with Sporadic Amyotrophic Lateral Sclerosis. JAMA Neurol. 2016, 73, 812–820. [Google Scholar] [CrossRef]
- Arai, T.; Hasegawa, M.; Akiyama, H.; Ikeda, K.; Nonaka, T.; Mori, H.; Mann, D.; Tsuchiya, K.; Yoshida, M.; Hashizume, Y.; et al. TDP-43 is a component of ubiquitin-positive tau-negative inclusions in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Biochem. Biophys. Res. Commun. 2006, 351, 602–611. [Google Scholar] [CrossRef]
- Bott, N.T.; Radke, A.; Stephens, M.L.; Kramer, J.H. Frontotemporal dementia: Diagnosis, deficits and management. Neurodegener. Dis. Manag. 2014, 4, 439–454. [Google Scholar] [CrossRef] [PubMed]
- Götzl, J.K.; Lang, C.M.; Haass, C.; Capell, A. Impaired protein degradation in FTLD and related disorders. Ageing Res. Rev. 2016, 32, 122–139. [Google Scholar] [CrossRef] [PubMed]
- Belzil, V.V.; Gendron, T.F.; Petrucelli, L. RNA-mediated toxicity in neurodegenerative disease. Mol. Cell. Neurosci. 2013, 56, 406–419. [Google Scholar] [CrossRef]
- Ling, S.C.; Polymenidou, M.; Cleveland, D.W. Converging mechanisms in ALS and FTD: Disrupted RNA and protein homestasis. Neuron 2013, 79, 416–438. [Google Scholar] [CrossRef] [PubMed]
- Liu, E.Y.; Cali, C.P.; Lee, E.B. RNA metabolism in neurodegenerative disease. Dis. Model. Mech. 2017, 10, 509–518. [Google Scholar] [CrossRef]
- Iraci, N.; Leonardi, T.; Gessler, F.; Vega, B.; Pluchino, S. Focus on Extracellular Vesicles: Physiological Role and Signalling Properties of Extracellular Membrane Vesicles. Int. J. Mol. Sci. 2016, 17, 171. [Google Scholar] [CrossRef] [PubMed]
- Fiandaca, M.S.; Kapogiannis, D.; Mapstone, M.; Boxer, A.; Eitan, E.; Schwartz, J.B.; Abner, E.L.; Petersen, R.C.; Federoff, H.J.; Miller, B.L.; et al. Identification of preclinical Alzheimer’s disease by a profile of pathogenic proteins in neurally de-rived blood exosomes: A case-control study. Alzheimers Dement. 2015, 11, 600.e1–601.e1. [Google Scholar] [CrossRef]
- Goetzl, E.J.; Boxer, A.L.; Schwartz, J.B.; Abner, E.L.; Petersen, R.C.; Miller, B.L.; Kapogiannis, D. Altered lysosomal proteins in neural-derived plasma exosomes in preclinical Alzheimer disease. Neurology 2015, 85, 40–47. [Google Scholar] [CrossRef]
- Fatima, F.; Nawaz, M. Vesiculated Long Non-Coding RNAs: Offshore Packages Deciphering Trans-Regulation between Cells, Cancer Progression and Resistance to Therapies. Non-Coding RNA 2017, 3, 10. [Google Scholar] [CrossRef]
- Matsumoto, J.; Stewart, T.; Banks, W.A.; Zhang, J. The Transport Mechanism of Extracellular Vesicles at the Blood-Brain Barrier. Curr. Pharm. Des. 2017, 23, 6206–6214. [Google Scholar] [CrossRef]
- Cocucci, E.; Meldolesi, J. Ectosomes and exosomes: Shedding the confusion between extracellular vesicles. Trends Cell Biol. 2015, 25, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Yao, Y.F.; Qu, M.W.; Li, G.C.; Zhang, F.B.; Rui, H.C. Circulating exosomal miRNAs as diagnostic biomarkers in Parkinson’s disease. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 5278–5283. [Google Scholar] [CrossRef]
- Gámez-Valero, A.; Campdelacreu, J.; Vilas, D.; Ispierto, L.; Reñé, R.; Álvarez, R.; Armengol, M.P.; Borràs, F.E.; Beyer, K. Ex-ploratory study on microRNA profiles from plasma-derived extracellular vesicles in Alzheimer’s disease and dementia with Lewy bodies. Transl. Neurodegener. 2019, 8, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Katsu, M.; Hama, Y.; Utsumi, J.; Takashina, K.; Yasumatsu, H.; Mori, F.; Wakabayashi, K.; Shoji, M.; Sasaki, H. MicroRNA expression profiles of neuron-derived extracellular vesicles in plasma from patients with amyotrophic lateral sclerosis. Neurosci. Lett. 2019, 708, 134176. [Google Scholar] [CrossRef] [PubMed]
- Sproviero, D.; La Salvia, S.; Giannini, M.; Crippa, V.; Gagliardi, S.; Bernuzzi, S.; Diamanti, L.; Ceroni, M.; Pansarasa, O.; Poletti, A.; et al. Pathological Proteins Are Transported by Extracellular Vesicles of Sporadic Amyotrophic Lateral Sclerosis Patients. Front. Neurosci. 2018, 12, 487. [Google Scholar] [CrossRef]
- Morasso, C.F.; Sproviero, D.; Mimmi, M.C.; Giannini, M.; Gagliardi, S.; Vanna, R.; Diamanti, L.; Bernuzzi, S.; Piccotti, F.; Truffi, M.; et al. Raman spectroscopy reveals biochemical differences in plasma derived extra-cellular vesicles from sporadic Amyotrophic Lateral Sclerosis patients. Nanomedicine 2020, 29, 102249. [Google Scholar] [CrossRef] [PubMed]
- Sproviero, D.; La Salvia, S.; Colombo, F.; Zucca, S.; Pansarasa, O.; Diamanti, L.; Costa, A.; Lova, L.; Giannini, M.; Gagliardi, S.; et al. Leukocyte derived microvesicles as disease progres-sion biomarkers in slow progressing amyotrophic lateral sclerosis patients. Front. Neurosci. 2019, 13, 344. [Google Scholar] [CrossRef]
- Bowie, A.G.; Unterholzner, L. Viral evasion and subversion of pattern-recognition receptor signalling. Nat. Rev. Immunol. 2008, 8, 911–922. [Google Scholar] [CrossRef]
- Mazin, A.V.; Mazina, O.M.; Bugreev, D.V.; Rossi, M.J. Rad54, the motor of homologous recombination. DNA Repair 2010, 9, 286–302. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Sato, S.; Ishii, K.J.; Coban, C.; Hemmi, H.; Yamamoto, M.; Terai, K.; Matsuda, M.; Inoue, J.-I.; Uematsu, S.; et al. Interferon-α induction through Toll-like receptors involves a direct interaction of IRF7 with MyD88 and TRAF6. Nat. Immunol. 2004, 5, 1061–1068. [Google Scholar] [CrossRef]
- Bi, F.; Huang, C.; Tong, J.; Qiu, G.; Huang, B.; Wu, Q.; Li, F.; Xu, Z.; Bowser, R.; Xia, X.-G.; et al. Reactive astrocytes secrete lcn2 to promote neuron death. Proc. Natl. Acad. Sci. USA 2013, 110, 4069–4074. [Google Scholar] [CrossRef]
- Liu, C.; Yao, M.-D.; Li, C.-P.; Shan, K.; Yang, H.; Wang, J.-J.; Liu, B.; Li, X.-M.; Yao, J.; Jiang, Q.; et al. Silencing of Circular RNA-ZNF609 Ameliorates Vascular Endothelial Dysfunction. Theranostics 2017, 7, 2863–2877. [Google Scholar] [CrossRef]
- Wang, Y.; Baskerville, S.; Shenoy, A.; Babiarz, J.E.; Baehner, L.; Blelloch, R. Embryonic stem cell-specific microRNAs regulate the G1-S transition and promote rapid proliferation. Nat. Genet. 2008, 40, 1478–1483. [Google Scholar] [CrossRef]
- Recchiuti, A.; Krishnamoorthy, S.; Fredman, G.; Chiang, N.; Serhan, C.N. MicroRNAs in resolution of acute inflammation: Identification of novel resolvin D1-miRNA circuits. FASEB J. 2011, 25, 544–560. [Google Scholar] [CrossRef]
- Conley, A.; Minciacchi, V.R.; Lee, D.H.; Knudsen, B.S.; Karlan, B.Y.; Citrigno, L.; Viglietto, G.; Tewari, M.; Freeman, M.R.; Demichelis, F.; et al. High-throughput sequencing of two populations of extracellular vesicles provides an mRNA signature that can be detected in the circulation of breast cancer patients. RNA Biol. 2017, 14, 305–316. [Google Scholar] [CrossRef]
- Crescitelli, R.; Lässer, C.; Szabó, T.G.; AKittel, M.E.; Dianzani, I.; Buzás, E.I.; Lötvall, J. Distinct RNA profiles in subpopulations of extracellular vesicles: Apoptotic bodies, microvesicles and exosomes. JEVs 2013, 2, 20677. [Google Scholar] [CrossRef]
- Waller, R.; Goodall, E.F.; Milo, M.; Cooper-Knock, J.; Da Costa, M.; Hobson, E.; Kazoka, M.; Wollff, H.; Heath, P.R.; Shaw, P.J.; et al. Serum miRNAs miR-206, 143–3p and 374b-5p as potential biomarkers for amyotrophic lateral sclerosis (ALS). Neurobiol. Aging 2017, 55, 123–131. [Google Scholar] [CrossRef] [PubMed]
- De Andrade, H.M.T.; de Albuquerque, M.; Avansini, S.H.; Rocha, C.d.S.; Dogini, D.B.; Nucci, A.; Carvalho, B.; Lopes-Cendes, I.; França, M.C., Jr. MicroRNAs-424 and 206 are potential prognostic markers in spinal onset amyotrophic lateral sclerosis. J. Neurol. Sci. 2016, 368, 19–24. [Google Scholar] [CrossRef]
- Raheja, R.; Regev, K.; Healy, B.C.; Mazzola, M.A.; Beynon, V.; Von Glehn, F.; Paul, A.; Diaz-Cruz, C.; Gholipour, T.; Glanz, B.I.; et al. Correlating serum micrornas and clinical parameters in amyotrophic lateral sclerosis. Muscle Nerve 2018, 58, 261–269. [Google Scholar] [CrossRef]
- Apolloni, S.; Parisi, C.; Volonté, C. MicroRNAs: Newcomers into the ALS Picture. CNS Neurol. Disord. Drug Targets 2015, 14, 194–207. [Google Scholar] [CrossRef]
- Modigliani, S.D.; Morlando, M.; Errichelli, L.; Sabatelli, M.; Bozzoni, I. An ALS-associated Mutation in the FUS 3′-UTR Disrupts a microRNA-FUS Regulatory Circuitry. Nat. Commun. 2014, 5, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Watts, M.E.; Williams, S.M.; Nithianantharajah, J.; Claudianos, C. Hypoxia-Induced MicroRNA-210 Targets Neurodegenerative Pathways. Non-Coding RNA 2018, 4, 10. [Google Scholar] [CrossRef]
- Ferrari, R.; Kapogiannis, D.; Huey, E.D.; Momeni, P. FTD and ALS: A Tale of Two Diseases. Curr. Alzheimer Res. 2011, 8, 273–294. [Google Scholar] [CrossRef]
- Achi, E.Y.; Rudnicki, S.A. ALS and Frontotemporal Dysfunction: A Review. Neurol. Res. Int. 2012, 2012, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Liscic, R.M. Als and Ftd: Insights into the disease mechanisms and therapeutic targets. Eur. J. Pharmacol. 2017, 817, 2–6. [Google Scholar] [CrossRef] [PubMed]
- Park, H.K.; Chung, S.J. New Perspective on Parkinsonism in Frontotemporal Lobar Degeneration. J. Mov. Disord. 2013, 6, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Armentero, M.T.; Sinforiani, E.; Ghezzi, C.; Bazzini, E.; Levandis, G.; Ambrosi, G.; Zangaglia, R.; Pacchetti, C.; Cereda, C.; Cova, E.; et al. Peripheral expression of key regulatory kinases in Alzheimer’s disease and Parkinson’s disease. Neurobiol. Aging 2011, 32, 2142–2151. [Google Scholar] [CrossRef] [PubMed]
- Yáñez-Mó, M.; Siljander, P.R.M.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological Properties of Extracellular Vesicles and their Physiological Functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed]
- Fiebich, B.L.; Batista, C.R.A.; Saliba, S.W.; Yousif, N.M.; De Oliveira, A.C.P. Role of Microglia TLRs in Neurodegeneration. Front. Cell. Neurosci. 2018, 12, 329. [Google Scholar] [CrossRef]
- Ciechanover, A.; Schwartz, A.L. The ubiquitin-mediated proteolytic pathway: Mechanisms of recognition of the proteolytic substrate and involvement in the degradation of native cellular proteins. FASEB J. 1994, 8, 182–191. [Google Scholar] [CrossRef]
- Layfield, R.; Cavey, J.R.; Lowe, J. Role of ubiquitin-mediated proteolysis in the pathogenesis of neurodegenerative disorders. Ageing Res. Rev. 2003, 2, 343–356. [Google Scholar] [CrossRef]
- Raiborg, C.; Stenmark, H. The ESCRT machinery in endosomal sorting of ubiquitylated membrane proteins. Nat. Cell Biol. 2009, 458, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Numakawa, T.; Suzuki, S.; Kumamaru, E.; Adachi, N.; Richards, M.; Kunugi, H. BDNF function and intracellular signaling in neurons. Histol. Histopathol. 2010, 25, 2. [Google Scholar] [CrossRef]
- Czubowicz, K.; Jęśko, H.; Wencel, P.; Lukiw, W.J.; Strosznajder, R.P. The Role of Ceramide and Sphingosine-1-Phosphate in Alzheimer’s Disease and Other Neurodegenerative Disorders. Mol. Neurobiol. 2019, 56, 5436–5455. [Google Scholar] [CrossRef]
- Kim, J.; Inoue, K.; Ishii, J.; Vanti, W.B.; Voronov, S.V.; Murchison, E.; Hannon, G.; Abeliovich, A.A. MicroRNA feedback circuit in midbrain dopamine neurons. Science 2007, 317, 1220–1224. [Google Scholar] [CrossRef] [PubMed]
- Satoh, J.; Kino, Y.; Niida, S. MicroRNA-Seq data analysis pipeline to identify blood biomarkers for Alzheimer’s disease from public data. Biomark. Insights 2015, 10, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Li, L.; Huang, C.; Li, X.; Peng, Y.; Li, J. MicroRNA-323–3p with clinical potential in rheumatoid arthritis. Alzheimer’s disease and ectopic pregnancy. Expert. Opin. Ther. Targets 2014, 18, 153–158. [Google Scholar] [CrossRef] [PubMed]
- McKhann, G.M.; Knopman, D.S.; Chertkow, H.; Hyman, B.T.; Jack, C.R., Jr.; Kawas, C.H.; Klunk, W.E.; Koroshetz, W.J.; Manly, J.J.; Mayeux, R.; et al. The diagnosis of dementia due to Alzheimer’s disease: Recommendations from the national institute on aging—Alzheimer’s association work groups on diagnostic guidelines for Alzheimer’s disease. Alzheimers Dement. 2011, 7, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Postuma, R.B.; Berg, D.; Stern, M.; Poewe, W.; Olanow, C.W.; Oertel, W.; Obeso, J.; Marek, K.; Litvan, I.; Lang, A.E.; et al. MDS clinical diagnostic criteria for Parkinson’s disease. Mov. Disord. 2015, 30, 1591–1601. [Google Scholar] [CrossRef] [PubMed]
- Chare, L.; Hodges, J.R.; Leyton, C.E.; McGinley, C.; Tan, R.H.; Kril, J.K.; Halliday, G.M. New criteria for frontotemporal dementia syndromes: Clinical and pathological diagnostic implications. J. Neurol. Neurosurg. Psychiatry 2014, 85, 865–870. [Google Scholar] [CrossRef]
- Brooks, B.R.; Miller, R.G.; Swash, M.; Munsat, T.L. El Escorial revisited: Revised criteria for the diagnosis of amyotrophic lateral sclerosis. Amyotroph. Lateral Scler. 2000, 1, 293–299. [Google Scholar] [CrossRef]
- Cordero, F.; Beccuti, M.; Arigoni, M.; Donatelli, S.; Calogero, R.A. Optimizing a Massive Parallel Sequencing Workflow for Quantitative miRNA Expression Analysis. PLoS ONE 2012, 7, e31630. [Google Scholar] [CrossRef]
- Kulkarni, N.; Alessandrì, L.; Panero, R.; Arigoni, M.; Olivero, M.; Ferrero, G.; Cordero, F.; Beccuti, M.; Calogero, R.A. Reproducible bioinformatics project: A community for reproducible bioinformatics analysis pipelines. BMC Bioinform. 2018, 19, 5–13. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNAs | AD | FTD | ALS | PD | ||||
|---|---|---|---|---|---|---|---|---|
| SEVs | LEVs | SEVs | LEVs | SEVs | LEVs | SEVs | LEVs | |
| up-regulated | 17 | 10 | 49 | 113 | 80 | 128 | 85 | 87 |
| down-regulated | 16 | 3 | 39 | 17 | 29 | 69 | 19 | 22 |
| Total | 33 | 13 | 88 | 130 | 109 | 197 | 104 | 109 |
| miRNA | ||
|---|---|---|
| NDs | Common miRNAs in SEVs and LEVs (%) | |
| SEVs | LEVs | |
| AD | 18.2 | 46.2 |
| FTD | 38.6 | 25.2 |
| ALS | 61.5 | 34 |
| PD | 32.7 | 31.2 |
| Patients | Sex | Age | Education (years) | Disease Duration (months) | Familiarity | cMMSE † | Tau †† | pTau ††† | Aβ †††† 1-42 |
|---|---|---|---|---|---|---|---|---|---|
| FTD1 | M | 54 | 8 | 24 | No | 8,97 | <10 | 10 | 683 |
| FTD2 | F | 57 | 8 | 36 | No | 15.74 | 419 | 43 | 616 |
| FTD3 | M | 70 | 8 | 36 | No | 18 | NA | NA | NA |
| FTD4 | M | 62 | 8 | 34 | No | 17 | 164 | 42 | 1312 |
| FTD5 | M | 52 | 8 | 34 | No | 17.97 | 195 | 42 | 1378 |
| FTD6 | M | 69 | 11 | 36 | No | 21 | NA | NA | NA |
| FTD7 | M | 56 | 13 | 60 | No | 17.99 | <10 | 7 | 360 |
| FTD8 | M | 49 | 5 | 12 | No | 18.31 | 799 | 68 | 1780 |
| FTD9 | F | 75 | 8 | 44 | No | 17 | NA | NA | NA |
| Column Title | CTRs | AD | FTD | ALS | PD |
|---|---|---|---|---|---|
| Recruited subjects | 6 | 6 | 9 | 6 | 9 |
| Age (mean ± SD) | 55 ± 5.2 | 77 ± 3.7 | 60 ± 6.7 | 72 ± 6.3 | 69 ± 3.6 |
| Males % | 43% | 50% | 78% | 50% | 60% |
| Females % | 67% | 50% | 22% | 50% | 40% |
| Patients | Sex | Age | Education (years) | Disease Duration (months) | Familiarity | Onset | Imaging † | MMSE †† | CDR ††† |
|---|---|---|---|---|---|---|---|---|---|
| AD1 | M | 78 | 5 | 50 | No | Amnesic | 1.3 | 15 | 2 |
| AD2 | M | 70 | 8 | 48 | Yes | Amnesic | 1 | 19 | 1.5 |
| AD3 | F | 79 | 5 | 48 | No | Amnesic | 2 | 20 | 1 |
| AD4 | M | 77 | 5 | 40 | No | Amnesic | 1 | 13 | 2 |
| AD5 | F | 82 | 5 | 38 | Yes | Amnesic | 1 | 18 | 1 |
| AD6 | F | 76 | 5 | 42 | Yes | Amnesic | 2.3 | 17 | 2 |
| Patients | Sex | Age | Education (years) | Disease Duration (months) | Familiarity | Site of Onset | ALSFRS † | Therapy (Riluzole) |
|---|---|---|---|---|---|---|---|---|
| ALS1 | F | 69 | 13 | 19 | No | Spinal | 9 | |
| ALS2 | F | 70 | NA | 52 | No | Spinal | 29 | 19 |
| ALS3 | F | 69 | NA | 11 | No | Spinal | 34 | 20 |
| ALS4 | M | 71 | 5 | 24 | No | Spinal | 46 | 13 |
| ALS5 | M | 72 | 13 | 67 | No | Spinal | 28 | 18 |
| ALS6 | M | 67 | 5 | 28 | No | Spinal | 40 | 17 |
| Patients | Sex | Age | Education (years) | Disease Duration (years) | Familiarity | Affected Side | UPDRS III † | Therapy LEDD †† | Disorders of Behaviour |
|---|---|---|---|---|---|---|---|---|---|
| PD1 | F | 70 | NA | 8 | No | Right | 12 | 350 | No |
| PD2 | M | 73 | NA | 7 | Yes | Right | 10 | 320 | ICD |
| PD3 | M | 68 | 8 | 19 | No | Left | 44 | 800 | ICD |
| PD4 | M | 69 | NA | 13 | Yes | Right | 18 | 500 | No |
| PD5 | F | 73 | 8 | 10 | Yes | Right | 17 | 1000 | No |
| PD6 | M | 76 | NA | 7 | No | Right | 18 | 650 | No |
| PD7 | F | 68 | NA | 6 | No | Right | 20 | 600 | No |
| PD8 | M | 76 | 8 | 13 | Yes | Left | 35 | 690 | No |
| PD9 | F | 73 | NA | 13 | Yes | Left | 15 | 350 | ICD |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sproviero, D.; Gagliardi, S.; Zucca, S.; Arigoni, M.; Giannini, M.; Garofalo, M.; Olivero, M.; Dell’Orco, M.; Pansarasa, O.; Bernuzzi, S.; et al. Different miRNA Profiles in Plasma Derived Small and Large Extracellular Vesicles from Patients with Neurodegenerative Diseases. Int. J. Mol. Sci. 2021, 22, 2737. https://doi.org/10.3390/ijms22052737
Sproviero D, Gagliardi S, Zucca S, Arigoni M, Giannini M, Garofalo M, Olivero M, Dell’Orco M, Pansarasa O, Bernuzzi S, et al. Different miRNA Profiles in Plasma Derived Small and Large Extracellular Vesicles from Patients with Neurodegenerative Diseases. International Journal of Molecular Sciences. 2021; 22(5):2737. https://doi.org/10.3390/ijms22052737
Chicago/Turabian StyleSproviero, Daisy, Stella Gagliardi, Susanna Zucca, Maddalena Arigoni, Marta Giannini, Maria Garofalo, Martina Olivero, Michela Dell’Orco, Orietta Pansarasa, Stefano Bernuzzi, and et al. 2021. "Different miRNA Profiles in Plasma Derived Small and Large Extracellular Vesicles from Patients with Neurodegenerative Diseases" International Journal of Molecular Sciences 22, no. 5: 2737. https://doi.org/10.3390/ijms22052737
APA StyleSproviero, D., Gagliardi, S., Zucca, S., Arigoni, M., Giannini, M., Garofalo, M., Olivero, M., Dell’Orco, M., Pansarasa, O., Bernuzzi, S., Avenali, M., Cotta Ramusino, M., Diamanti, L., Minafra, B., Perini, G., Zangaglia, R., Costa, A., Ceroni, M., Perrone-Bizzozero, N. I., ... Cereda, C. (2021). Different miRNA Profiles in Plasma Derived Small and Large Extracellular Vesicles from Patients with Neurodegenerative Diseases. International Journal of Molecular Sciences, 22(5), 2737. https://doi.org/10.3390/ijms22052737