
Novel Molecular Insights into Human Lipid-Mediated T Cell Immunity


Abstract
1. Introduction
2. CD1d Recognition by Conventional Natural Killer T Cells
3. ‘Atypical’ Molecular Recognition of CD1d-Lipid by Human NKT TCRs
3.1. CD1d-α-GalCer Recognition by Human ‘Atypical’ NKT αβTCRs
3.2. Beyond the Molecular Recognition of CD1d-Lipids by αβTCRs
4. Molecular Presentation of Novel Self-Lipids by Group 1 CD1
4.1. Presentation of Self-Lipids by CD1a in Human Skin
4.2. CD1b Presenting Self- and Microbial-Lipids
4.3. The Molecular Diversity of Lipids Presentation by CD1c
5. Molecular Mechanism Underpinning the Recognition of Group 1 CD1-Restricted Self-Lipids
5.1. Beyond the Left-Right Mismatch Recognition of CD1a
5.2. Left/Right-Centric Mechanism of CD1b-Lipid Recognition in Diseases
5.3. Beyond the Buried Ligand Model of CD1c Recognition
6. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| APC | Antigen presenting cell |
| CD1 | Cluster of differentiation 1 |
| CDR | Complementarity determining region |
| iNKT | Invariant Natural Killer T cells |
| MHC | Major histocompatibility complex |
| PBMC | Peripheral blood mononuclear cell |
| TCR | T cell receptor |
| TRAV | Human T cell receptor alpha variable gene |
| TRBV | Human T cell receptor beta variable gene |
| TRDV | Human T cell receptor delta variable gene |
| TRGV | Human T cell receptor gamma variable gene |
References
- Rossjohn, J.; Gras, S.; Miles, J.J.; Turner, S.J.; Godfrey, D.I.; McCluskey, J. T cell antigen receptor recognition of antigen-presenting molecules. Annu. Rev. Immunol. 2015, 33, 169–200. [Google Scholar] [CrossRef] [PubMed]
- Brigl, M.; Brenner, M.B. CD1: Antigen presentation and T cell function. Annu. Rev. Immunol. 2004, 22, 817–890. [Google Scholar] [CrossRef] [PubMed]
- Moody, D.B.; Zajonc, D.M.; Wilson, I.A. Anatomy of CD1-lipid antigen complexes. Nat. Rev. Immunol. 2005, 5, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Rossjohn, J.; Pellicci, D.G.; Patel, O.; Gapin, L.; Godfrey, D.I. Recognition of CD1d-restricted antigens by natural killer T cells. Nat. Rev. Immunol. 2012, 12, 845–857. [Google Scholar] [CrossRef] [PubMed]
- Godfrey, D.I.; MacDonald, H.R.; Kronenberg, M.; Smyth, M.J.; Van Kaer, L. NKT cells: What’s in a name? Nat. Rev. Immunol. 2004, 4, 231–237. [Google Scholar] [CrossRef]
- Lantz, O.; Bendelac, A. An invariant T cell receptor alpha chain is used by a unique subset of major histocompatibility complex class I-specific CD4+ and CD4-8- T cells in mice and humans. J. Exp. Med. 1994, 180, 1097–1106. [Google Scholar] [CrossRef] [PubMed]
- Godfrey, D.I.; Uldrich, A.P.; McCluskey, J.; Rossjohn, J.; Moody, D.B. The burgeoning family of unconventional T cells. Nat. Immunol. 2015, 16, 1114–1123. [Google Scholar] [CrossRef]
- Borg, N.A.; Kjer-Nielsen, L.; McCluskey, J.; Rossjohn, J. Structural insight into natural killer T cell receptor recognition of CD1d. Adv. Exp. Med. Biol. 2007, 598, 20–34. [Google Scholar] [CrossRef]
- Patel, O.; Pellicci, D.G.; Gras, S.; Sandoval-Romero, M.L.; Uldrich, A.P.; Mallevaey, T.; Clarke, A.J.; Le Nours, J.; Theodossis, A.; Cardell, S.L.; et al. Recognition of CD1d-sulfatide mediated by a type II natural killer T cell antigen receptor. Nat. Immunol. 2012, 13, 857–863. [Google Scholar] [CrossRef] [PubMed]
- Girardi, E.; Maricic, I.; Wang, J.; Mac, T.T.; Iyer, P.; Kumar, V.; Zajonc, D.M. Type II natural killer T cells use features of both innate-like and conventional T cells to recognize sulfatide self antigens. Nat. Immunol. 2012, 13, 851–856. [Google Scholar] [CrossRef] [PubMed]
- Brigl, M.; van den Elzen, P.; Chen, X.; Meyers, J.H.; Wu, D.; Wong, C.H.; Reddington, F.; Illarianov, P.A.; Besra, G.S.; Brenner, M.B.; et al. Conserved and heterogeneous lipid antigen specificities of CD1d-restricted NKT cell receptors. J. Immunol. 2006, 176, 3625–3634. [Google Scholar] [CrossRef] [PubMed]
- Gadola, S.D.; Dulphy, N.; Salio, M.; Cerundolo, V. Valpha24-JalphaQ-independent, CD1d-restricted recognition of alpha-galactosylceramide by human CD4(+) and CD8alphabeta(+) T lymphocytes. J. Immunol. 2002, 168, 5514–5520. [Google Scholar] [CrossRef]
- Constantinides, M.G.; Picard, D.; Savage, A.K.; Bendelac, A. A naive-like population of human CD1d-restricted T cells expressing intermediate levels of promyelocytic leukemia zinc finger. J. Immunol. 2011, 187, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Sagaseta, J.; Kung, J.E.; Savage, P.B.; Gumperz, J.; Adams, E.J. The molecular basis for recognition of CD1d/alpha-galactosylceramide by a human non-Valpha24 T cell receptor. PLoS Biol. 2012, 10, e1001412. [Google Scholar] [CrossRef] [PubMed]
- Le Nours, J.; Praveena, T.; Pellicci, D.G.; Gherardin, N.A.; Ross, F.J.; Lim, R.T.; Besra, G.S.; Keshipeddy, S.; Richardson, S.K.; Howell, A.R.; et al. Atypical natural killer T-cell receptor recognition of CD1d-lipid antigens. Nat. Commun. 2016, 7, 10570. [Google Scholar] [CrossRef]
- Uldrich, A.P.; Le Nours, J.; Pellicci, D.G.; Gherardin, N.A.; McPherson, K.G.; Lim, R.T.; Patel, O.; Beddoe, T.; Gras, S.; Rossjohn, J.; et al. CD1d-lipid antigen recognition by the gammadelta TCR. Nat. Immunol. 2013, 14, 1137–1145. [Google Scholar] [CrossRef]
- Luoma, A.M.; Castro, C.D.; Mayassi, T.; Bembinster, L.A.; Bai, L.; Picard, D.; Anderson, B.; Scharf, L.; Kung, J.E.; Sibener, L.V.; et al. Crystal structure of Vdelta1 T cell receptor in complex with CD1d-sulfatide shows MHC-like recognition of a self-lipid by human gammadelta T cells. Immunity 2013, 39, 1032–1042. [Google Scholar] [CrossRef]
- Pellicci, D.G.; Uldrich, A.P.; Le Nours, J.; Ross, F.; Chabrol, E.; Eckle, S.B.; de Boer, R.; Lim, R.T.; McPherson, K.; Besra, G.; et al. The molecular bases of delta/alphabeta T cell-mediated antigen recognition. J. Exp. Med. 2014, 211, 2599–2615. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Picard, D.; Anderson, B.; Chaudhary, V.; Luoma, A.; Jabri, B.; Adams, E.J.; Savage, P.B.; Bendelac, A. The majority of CD1d-sulfatide-specific T cells in human blood use a semiinvariant Vdelta1 TCR. Eur. J. Immunol. 2012, 42, 2505–2510. [Google Scholar] [CrossRef]
- Hardman, C.S.; Chen, Y.L.; Salimi, M.; Jarrett, R.; Johnson, D.; Jarvinen, V.J.; Owens, R.J.; Repapi, E.; Cousins, D.J.; Barlow, J.L.; et al. CD1a presentation of endogenous antigens by group 2 innate lymphoid cells. Sci. Immunol. 2017, 2. [Google Scholar] [CrossRef]
- Cernadas, M.; Cavallari, M.; Watts, G.; Mori, L.; De Libero, G.; Brenner, M.B. Early recycling compartment trafficking of CD1a is essential for its intersection and presentation of lipid antigens. J. Immunol. 2010, 184, 1235–1241. [Google Scholar] [CrossRef]
- Betts, R.J.; Perkovic, A.; Mahapatra, S.; Del Bufalo, A.; Camara, K.; Howell, A.R.; Martinozzi Teissier, S.; De Libero, G.; Mori, L. Contact sensitizers trigger human CD1-autoreactive T-cell responses. Eur. J. Immunol. 2017, 47, 1171–1180. [Google Scholar] [CrossRef] [PubMed]
- De Jong, A.; Cheng, T.Y.; Huang, S.; Gras, S.; Birkinshaw, R.W.; Kasmar, A.G.; Van Rhijn, I.; Pena-Cruz, V.; Ruan, D.T.; Altman, J.D.; et al. CD1a-autoreactive T cells recognize natural skin oils that function as headless antigens. Nat. Immunol. 2014, 15, 177–185. [Google Scholar] [CrossRef]
- Cotton, R.N.; Cheng, T.Y.; Wegrecki, M.; Le Nours, J.; Orgill, D.P.; Pomahac, B.; Talbot, S.G.; Willis, R.A.; Altman, J.D.; de Jong, A.; et al. Human skin is colonized by T cells that recognize CD1a independently of lipid. J. Clin. Investig. 2021, 131. [Google Scholar] [CrossRef]
- Birkinshaw, R.W.; Pellicci, D.G.; Cheng, T.-Y.; Keller, A.N.; Sandoval-Romero, M.; Gras, S.; de Jong, A.; Uldrich, A.P.; Moody, D.B.; Godfrey, D.I.; et al. αβ T cell antigen receptor recognition of CD1a presenting self lipid ligands. Nat. Immunol. 2015, 16, 258–266. [Google Scholar] [CrossRef]
- Zajonc, D.M.; Crispin, M.D.; Bowden, T.A.; Young, D.C.; Cheng, T.Y.; Hu, J.; Costello, C.E.; Rudd, P.M.; Dwek, R.A.; Miller, M.J.; et al. Molecular mechanism of lipopeptide presentation by CD1a. Immunity 2005, 22, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Zajonc, D.M.; Elsliger, M.A.; Teyton, L.; Wilson, I.A. Crystal structure of CD1a in complex with a sulfatide self antigen at a resolution of 2.15 A. Nat. Immunol. 2003, 4, 808–815. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Hu, Y.; Yongqing, T.; Kim, J.; Hughes, V.A.; Le Nours, J.; Marquez, E.A.; Purcell, A.W.; Wan, Q.; Sugita, M.; et al. CD1a on Langerhans cells controls inflammatory skin disease. Nat. Immunol. 2016, 17, 1159–1166. [Google Scholar] [CrossRef] [PubMed]
- Nicolai, S.; Wegrecki, M.; Cheng, T.Y.; Bourgeois, E.A.; Cotton, R.N.; Mayfield, J.A.; Monnot, G.C.; Le Nours, J.; Van Rhijn, I.; Rossjohn, J.; et al. Human T cell response to CD1a and contact dermatitis allergens in botanical extracts and commercial skin care products. Sci. Immunol. 2020, 5. [Google Scholar] [CrossRef]
- Shahine, A. The intricacies of self-lipid antigen presentation by CD1b. Mol. Immunol. 2018, 104, 27–36. [Google Scholar] [CrossRef]
- Mansour, S.; Tocheva, A.S.; Cave-Ayland, C.; Machelett, M.M.; Sander, B.; Lissin, N.M.; Molloy, P.E.; Baird, M.S.; Stubs, G.; Schroder, N.W.; et al. Cholesteryl esters stabilize human CD1c conformations for recognition by self-reactive T cells. Proc. Natl. Acad. Sci. USA 2016, 113, E1266–E1275. [Google Scholar] [CrossRef]
- Gras, S.; Van Rhijn, I.; Shahine, A.; Cheng, T.Y.; Bhati, M.; Tan, L.L.; Halim, H.; Tuttle, K.D.; Gapin, L.; Le Nours, J.; et al. T cell receptor recognition of CD1b presenting a mycobacterial glycolipid. Nat. Commun. 2016, 7, 13257. [Google Scholar] [CrossRef] [PubMed]
- Shahine, A.; Van Rhijn, I.; Cheng, T.Y.; Iwany, S.; Gras, S.; Moody, D.B.; Rossjohn, J. A molecular basis of human T cell receptor autoreactivity toward self-phospholipids. Sci. Immunol. 2017, 2. [Google Scholar] [CrossRef]
- Shahine, A.; Reinink, P.; Reijneveld, J.F.; Gras, S.; Holzheimer, M.; Cheng, T.Y.; Minnaard, A.J.; Altman, J.D.; Lenz, S.; Prandi, J.; et al. A T-cell receptor escape channel allows broad T-cell response to CD1b and membrane phospholipids. Nat. Commun. 2019, 10, 56. [Google Scholar] [CrossRef] [PubMed]
- Wun, K.S.; Reijneveld, J.F.; Cheng, T.Y.; Ladell, K.; Uldrich, A.P.; Le Nours, J.; Miners, K.L.; McLaren, J.E.; Grant, E.J.; Haigh, O.L.; et al. T cell autoreactivity directed toward CD1c itself rather than toward carried self lipids. Nat. Immunol. 2018, 19, 397–406. [Google Scholar] [CrossRef]
- Cotton, R.N.; Shahine, A.; Rossjohn, J.; Moody, D.B. Lipids hide or step aside for CD1-autoreactive T cell receptors. Curr. Opin. Immunol. 2018, 52, 93–99. [Google Scholar] [CrossRef]
- Batuwangala, T.; Shepherd, D.; Gadola, S.D.; Gibson, K.J.; Zaccai, N.R.; Fersht, A.R.; Besra, G.S.; Cerundolo, V.; Jones, E.Y. The crystal structure of human CD1b with a bound bacterial glycolipid. J. Immunol. 2004, 172, 2382–2388. [Google Scholar] [CrossRef]
- Van Rhijn, I.; Kasmar, A.; de Jong, A.; Gras, S.; Bhati, M.; Doorenspleet, M.E.; de Vries, N.; Godfrey, D.I.; Altman, J.D.; de Jager, W.; et al. A conserved human T cell population targets mycobacterial antigens presented by CD1b. Nat. Immunol. 2013, 14, 706–713. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Alles, L.F.; Versluis, K.; Maveyraud, L.; Vallina, A.T.; Sansano, S.; Bello, N.F.; Gober, H.J.; Guillet, V.; de la Salle, H.; Puzo, G.; et al. Endogenous phosphatidylcholine and a long spacer ligand stabilize the lipid-binding groove of CD1b. EMBO J. 2006, 25, 3684–3692. [Google Scholar] [CrossRef] [PubMed]
- Van Rhijn, I.; Gherardin, N.A.; Kasmar, A.; de Jager, W.; Pellicci, D.G.; Kostenko, L.; Tan, L.L.; Bhati, M.; Gras, S.; Godfrey, D.I.; et al. TCR bias and affinity define two compartments of the CD1b-glycolipid-specific T Cell repertoire. J. Immunol. 2014, 192, 4054–4060. [Google Scholar] [CrossRef] [PubMed]
- Scharf, L.; Li, N.S.; Hawk, A.J.; Garzon, D.; Zhang, T.; Fox, L.M.; Kazen, A.R.; Shah, S.; Haddadian, E.J.; Gumperz, J.E.; et al. The 2.5 A structure of CD1c in complex with a mycobacterial lipid reveals an open groove ideally suited for diverse antigen presentation. Immunity 2010, 33, 853–862. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Ly, D.; Li, N.S.; Altman, J.D.; Piccirilli, J.A.; Moody, D.B.; Adams, E.J. Molecular basis of mycobacterial lipid antigen presentation by CD1c and its recognition by alphabeta T cells. Proc. Natl. Acad. Sci. USA 2014, 111, E4648–E4657. [Google Scholar] [CrossRef] [PubMed]
- De Jong, A.; Pena-Cruz, V.; Cheng, T.Y.; Clark, R.A.; Van Rhijn, I.; Moody, D.B. CD1a-autoreactive T cells are a normal component of the human alphabeta T cell repertoire. Nat. Immunol. 2010, 11, 1102–1109. [Google Scholar] [CrossRef] [PubMed]
- De Lalla, C.; Lepore, M.; Piccolo, F.M.; Rinaldi, A.; Scelfo, A.; Garavaglia, C.; Mori, L.; De Libero, G.; Dellabona, P.; Casorati, G. High-frequency and adaptive-like dynamics of human CD1 self-reactive T cells. Eur. J. Immunol. 2011, 41, 602–610. [Google Scholar] [CrossRef]
- Bourgeois, E.A.; Subramaniam, S.; Cheng, T.Y.; De Jong, A.; Layre, E.; Ly, D.; Salimi, M.; Legaspi, A.; Modlin, R.L.; Salio, M.; et al. Bee venom processes human skin lipids for presentation by CD1a. J. Exp. Med. 2015, 212, 149–163. [Google Scholar] [CrossRef]
- Jarrett, R.; Salio, M.; Lloyd-Lavery, A.; Subramaniam, S.; Bourgeois, E.; Archer, C.; Cheung, K.L.; Hardman, C.; Chandler, D.; Salimi, M.; et al. Filaggrin inhibits generation of CD1a neolipid antigens by house dust mite-derived phospholipase. Sci. Transl. Med. 2016, 8, 325ra18. [Google Scholar] [CrossRef]
- Subramaniam, S.; Aslam, A.; Misbah, S.A.; Salio, M.; Cerundolo, V.; Moody, D.B.; Ogg, G. Elevated and cross-responsive CD1a-reactive T cells in bee and wasp venom allergic individuals. Eur. J. Immunol. 2016, 46, 242–252. [Google Scholar] [CrossRef]
- Cheung, K.L.; Jarrett, R.; Subramaniam, S.; Salimi, M.; Gutowska-Owsiak, D.; Chen, Y.L.; Hardman, C.; Xue, L.; Cerundolo, V.; Ogg, G. Psoriatic T cells recognize neolipid antigens generated by mast cell phospholipase delivered by exosomes and presented by CD1a. J. Exp. Med. 2016, 213, 2399–2412. [Google Scholar] [CrossRef]
- Moody, D.B.; Reinhold, B.B.; Guy, M.R.; Beckman, E.M.; Frederique, D.E.; Furlong, S.T.; Ye, S.; Reinhold, V.N.; Sieling, P.A.; Modlin, R.L.; et al. Structural requirements for glycolipid antigen recognition by CD1b-restricted T cells. Science 1997, 278, 283–286. [Google Scholar] [CrossRef] [PubMed]
- Van Rhijn, I.; van Berlo, T.; Hilmenyuk, T.; Cheng, T.Y.; Wolf, B.J.; Tatituri, R.V.; Uldrich, A.P.; Napolitani, G.; Cerundolo, V.; Altman, J.D.; et al. Human autoreactive T cells recognize CD1b and phospholipids. Proc. Natl. Acad. Sci. USA 2016, 113, 380–385. [Google Scholar] [CrossRef]
- Horvath, S.E.; Daum, G. Lipids of mitochondria. Prog. Lipid Res. 2013, 52, 590–614. [Google Scholar] [CrossRef]
- Morita, S.Y.; Terada, T. Enzymatic measurement of phosphatidylglycerol and cardiolipin in cultured cells and mitochondria. Sci. Rep. 2015, 5, 11737. [Google Scholar] [CrossRef]
- Bagchi, S.; Li, S.; Wang, C.R. CD1b-autoreactive T cells recognize phospholipid antigens and contribute to antitumor immunity against a CD1b(+) T cell lymphoma. Oncoimmunology 2016, 5, e1213932. [Google Scholar] [CrossRef] [PubMed]
- Bagchi, S.; He, Y.; Zhang, H.; Cao, L.; Van Rhijn, I.; Moody, D.B.; Gudjonsson, J.E.; Wang, C.R. CD1b-autoreactive T cells contribute to hyperlipidemia-induced skin inflammation in mice. J. Clin. Investig. 2017, 127, 2339–2352. [Google Scholar] [CrossRef] [PubMed]
- Reinink, P.; Shahine, A.; Gras, S.; Cheng, T.Y.; Farquhar, R.; Lopez, K.; Suliman, S.A.; Reijneveld, J.F.; Le Nours, J.; Tan, L.L.; et al. A TCR beta-Chain Motif Biases toward Recognition of Human CD1 Proteins. J. Immunol. 2019, 203, 3395–3406. [Google Scholar] [CrossRef]
- Reijneveld, J.F.; Ocampo, T.A.; Shahine, A.; Gully, B.S.; Vantourout, P.; Hayday, A.C.; Rossjohn, J.; Moody, D.B.; Van Rhijn, I. Human gammadelta T cells recognize CD1b by two distinct mechanisms. Proc. Natl. Acad. Sci. USA 2020, 117, 22944–22952. [Google Scholar] [CrossRef] [PubMed]
- Le Nours, J.; Gherardin, N.A.; Ramarathinam, S.H.; Awad, W.; Wiede, F.; Gully, B.S.; Khandokar, Y.; Praveena, T.; Wubben, J.M.; Sandow, J.J.; et al. A class of gammadelta T cell receptors recognize the underside of the antigen-presenting molecule MR1. Science 2019, 366, 1522–1527. [Google Scholar] [CrossRef]
- Ly, D.; Kasmar, A.G.; Cheng, T.Y.; de Jong, A.; Huang, S.; Roy, S.; Bhatt, A.; van Summeren, R.P.; Altman, J.D.; Jacobs, W.R., Jr.; et al. CD1c tetramers detect ex vivo T cell responses to processed phosphomycoketide antigens. J. Exp. Med. 2013, 210, 729–741. [Google Scholar] [CrossRef]
- Moody, D.B.; Ulrichs, T.; Muhlecker, W.; Young, D.C.; Gurcha, S.S.; Grant, E.; Rosat, J.P.; Brenner, M.B.; Costello, C.E.; Besra, G.S.; et al. CD1c-mediated T-cell recognition of isoprenoid glycolipids in Mycobacterium tuberculosis infection. Nature 2000, 404, 884–888. [Google Scholar] [CrossRef]
- Guo, T.; Koo, M.Y.; Kagoya, Y.; Anczurowski, M.; Wang, C.H.; Saso, K.; Butler, M.O.; Hirano, N. A Subset of Human Autoreactive CD1c-Restricted T Cells Preferentially Expresses TRBV4-1(+) TCRs. J. Immunol. 2018, 200, 500–511. [Google Scholar] [CrossRef]
- Roy, S.; Ly, D.; Castro, C.D.; Li, N.S.; Hawk, A.J.; Altman, J.D.; Meredith, S.C.; Piccirilli, J.A.; Moody, D.B.; Adams, E.J. Molecular Analysis of Lipid-Reactive Vdelta1 gammadelta T Cells Identified by CD1c Tetramers. J. Immunol. 2016, 196, 1933–1942. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
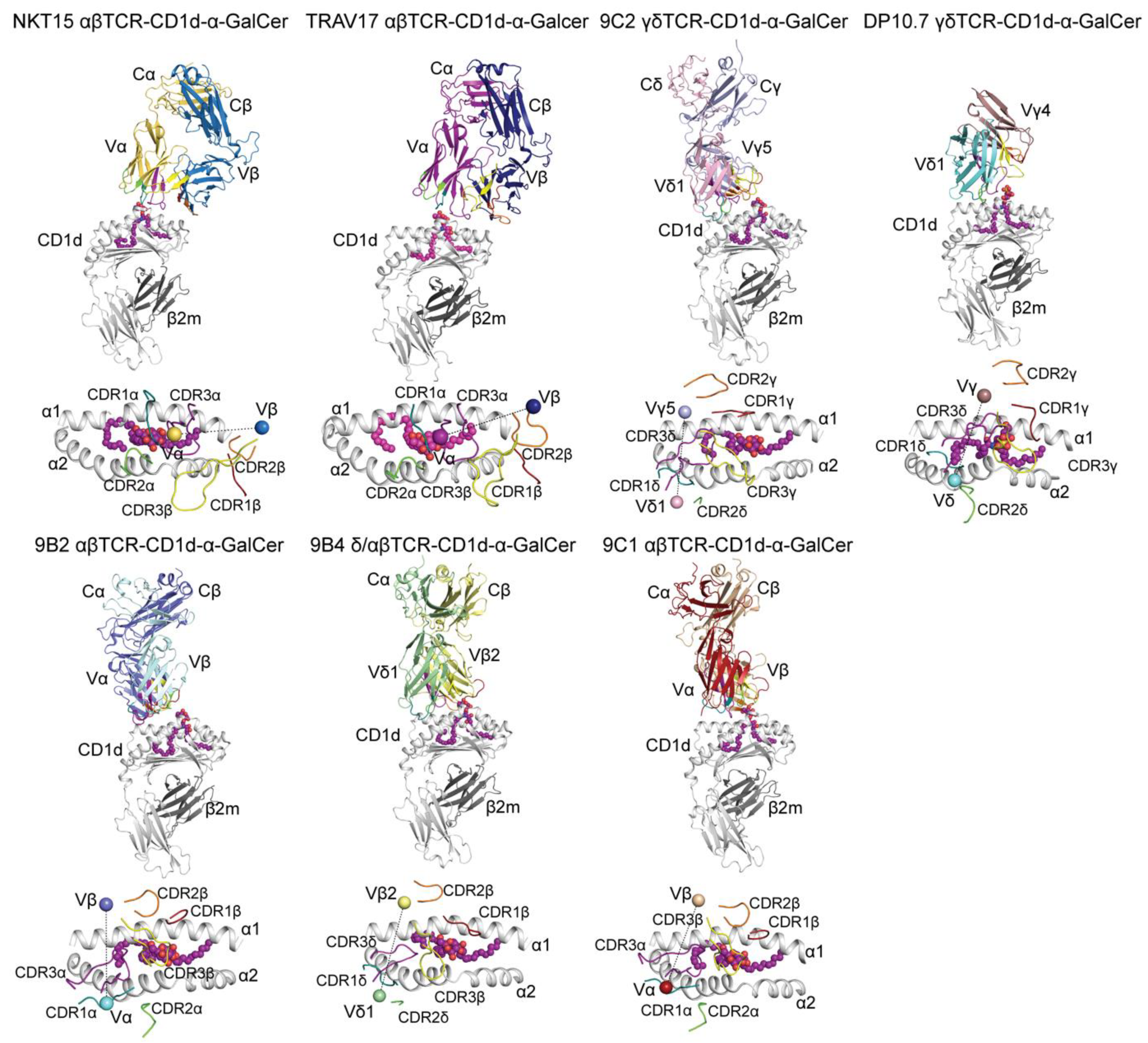
| T Cell Receptor | T Cell Type | PDB ID | References |
|---|---|---|---|
| TRAV17 | αβ | 4EN3 | [14] |
| 9C1 | αβ | 4WW2 | [15] |
| 9B2 | αβ | 4WWK | [15] |
| 9C2 | γδ | 4LHU | [16] |
| DP10.7 | γδ | 4MNG | [17] |
| 9B4 | δ/αβ | 4WO4 | [18] |
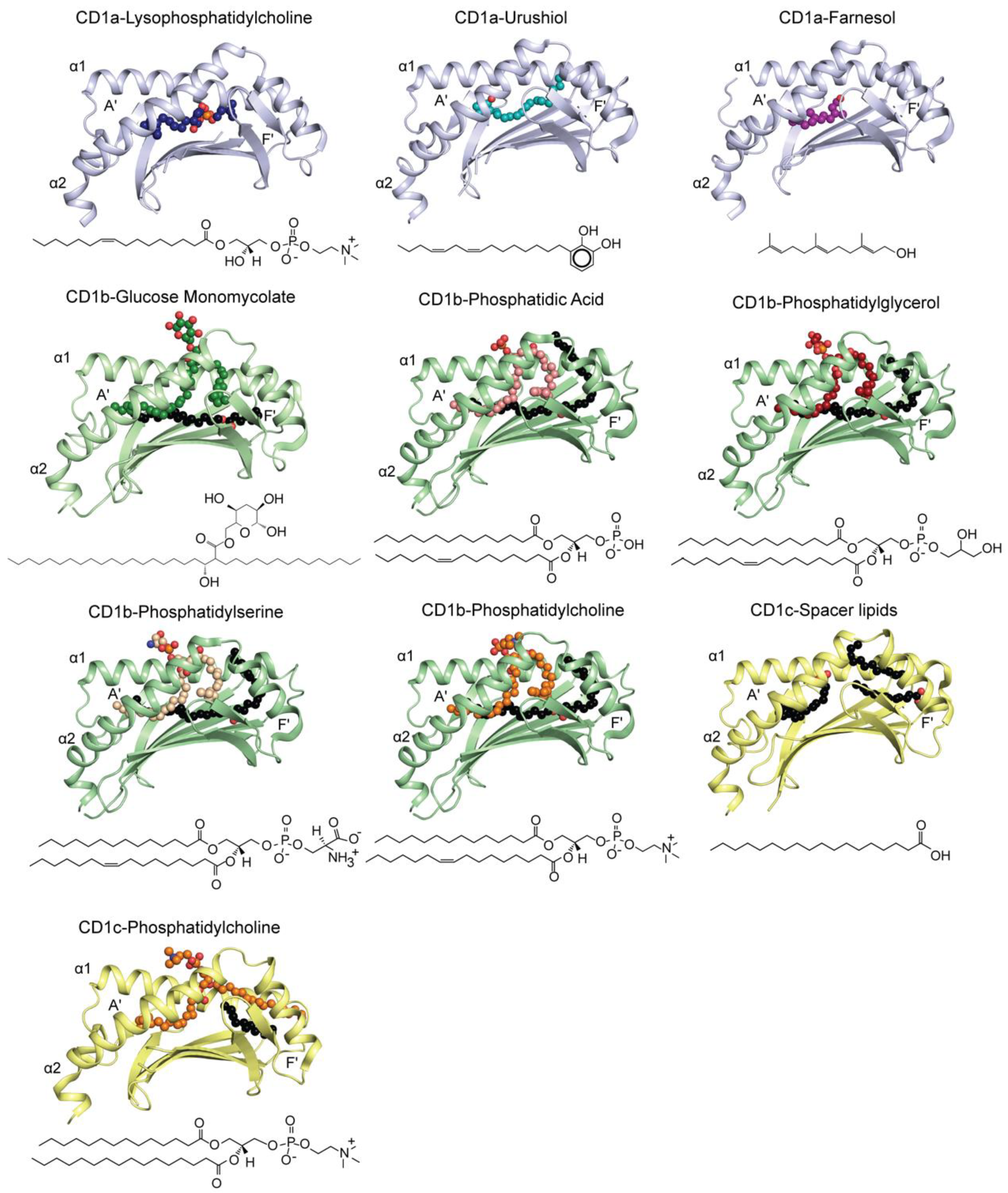
| Lipids | Source | Type | PDB ID | References |
|---|---|---|---|---|
| CD1a | ||||
| Lysophosphatidylcholine | Self (Commercial) | Ternary | 4X6C | [25] |
| Endogenous fatty acids | Self (expression system) | Ternary | 4X6D | [25] |
| Lysophosphatidylcholine | Self (Commercial) | Binary | 4X6E | [25] |
| Sphingomyelin | Self (Commercial) | Binary | 4X6F | [25] |
| Urushiol | Plant (commercial) | Binary | 5JIA | [28] |
| Farnesol | Balsam of Peru (commercial) | Binary | 6NUX | [29] |
| CD1b | ||||
| Glucose monomycolate C36 | Bacterial (purified) | Binary | 5L2J | [32] |
| Glucose monomycolate C36 | Bacterial (purified) | Ternary | 5L2K | [32] |
| Phosphatidylserine | Self (commercial) | Binary | 5WKE | [33] |
| Phosphatidic Acid | Self (commercial) | Binary | 5WKG | [33] |
| Phosphatidylglycerol | Bacterial (commercial) | Ternary | 5WKI | [33] |
| Phosphatidylglycerol | Bacterial (commercial) | Binary | 5WL1 | [33] |
| Phosphatidylcholine | Self (commercial) | Ternary | 6CUG | [34] |
| Phosphatidylcholine | Self (commercial) | Binary | 6D64 | [34] |
| CD1c | ||||
| Spacer lipids | Refolding | Binary | 5C9J | [31] |
| Monoacyl glyceride | Self (expression system) | Ternary | 6C09 | [35] |
| Phosphatidylcholine | Self (expression system) | Binary | 6C15 | [35] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shahine, A.; Wegrecki, M.; Le Nours, J. Novel Molecular Insights into Human Lipid-Mediated T Cell Immunity. Int. J. Mol. Sci. 2021, 22, 2617. https://doi.org/10.3390/ijms22052617
Shahine A, Wegrecki M, Le Nours J. Novel Molecular Insights into Human Lipid-Mediated T Cell Immunity. International Journal of Molecular Sciences. 2021; 22(5):2617. https://doi.org/10.3390/ijms22052617
Chicago/Turabian StyleShahine, Adam, Marcin Wegrecki, and Jérôme Le Nours. 2021. "Novel Molecular Insights into Human Lipid-Mediated T Cell Immunity" International Journal of Molecular Sciences 22, no. 5: 2617. https://doi.org/10.3390/ijms22052617
APA StyleShahine, A., Wegrecki, M., & Le Nours, J. (2021). Novel Molecular Insights into Human Lipid-Mediated T Cell Immunity. International Journal of Molecular Sciences, 22(5), 2617. https://doi.org/10.3390/ijms22052617