
The Importance of Sex in the Discovery of Colorectal Cancer Prognostic Biomarkers

 ,
, 
Abstract
1. Introduction
2. Results
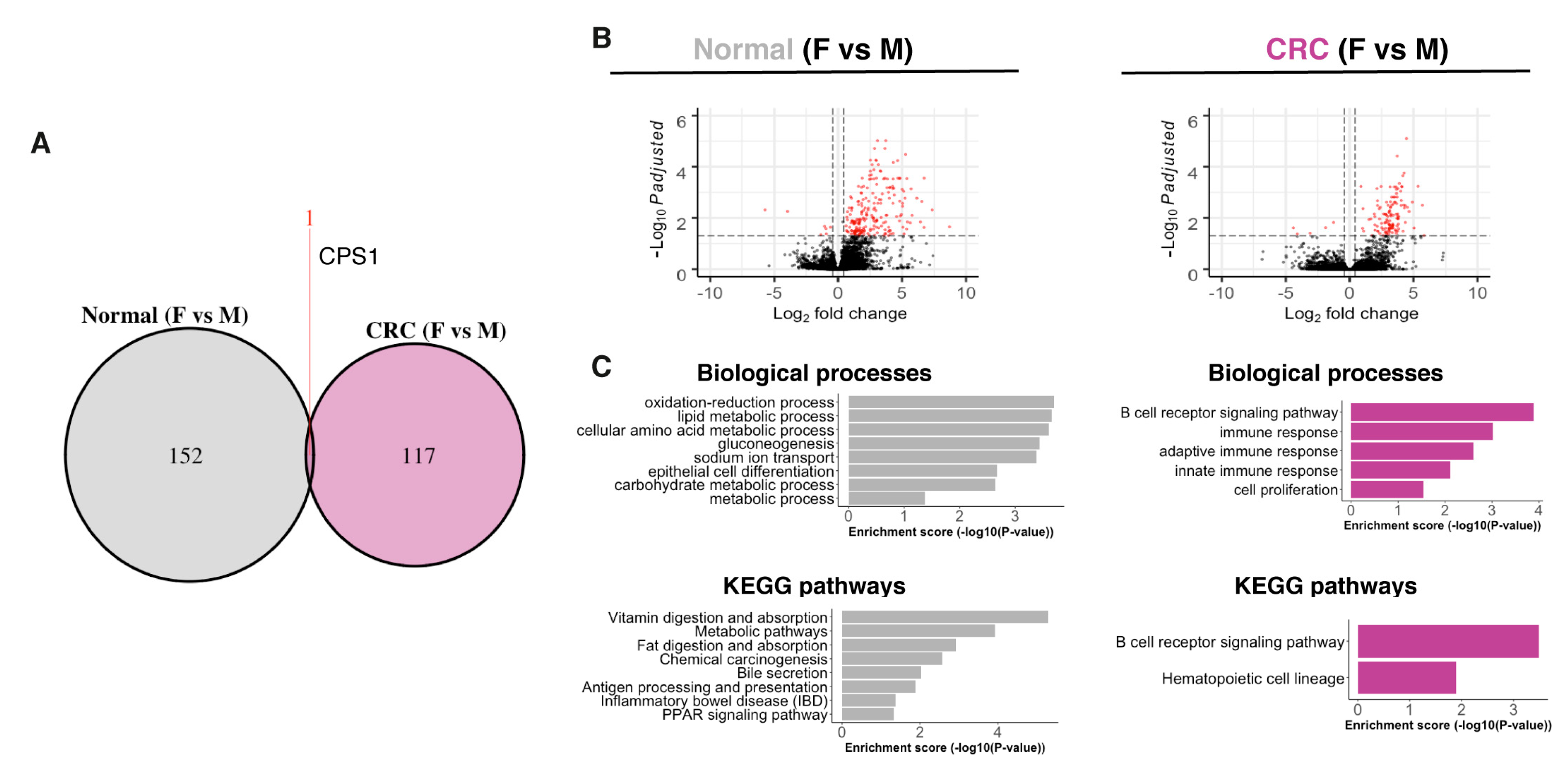
2.1. Normal Colon and CRC Transcriptomes Exhibit Sex-Related Differences
2.2. Transcriptomic Sex Differences Independent of Subtype and Tumor Location
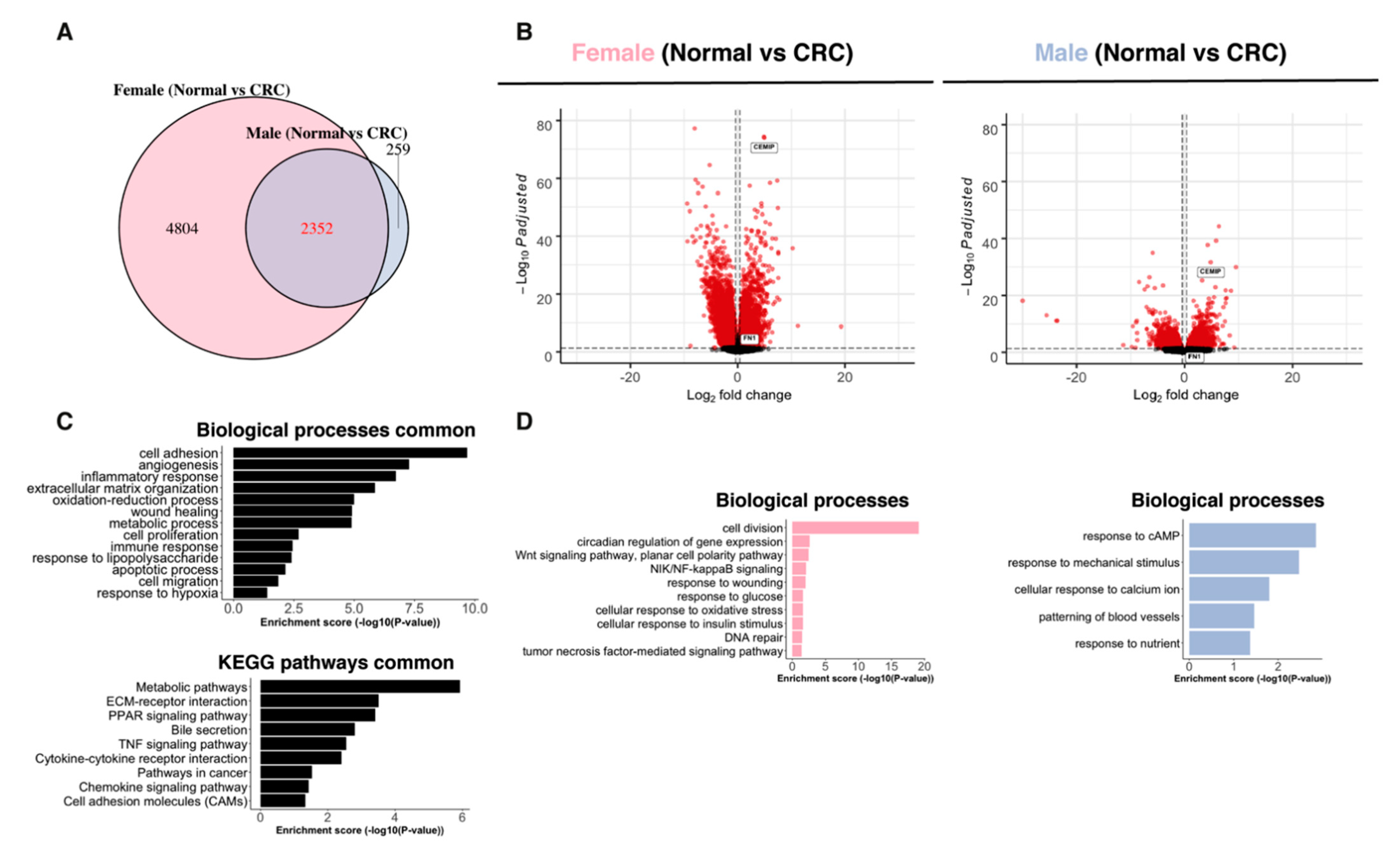
2.3. Paired Normal–CRC Gene Expression Analysis Reveals Sex-Related Differences
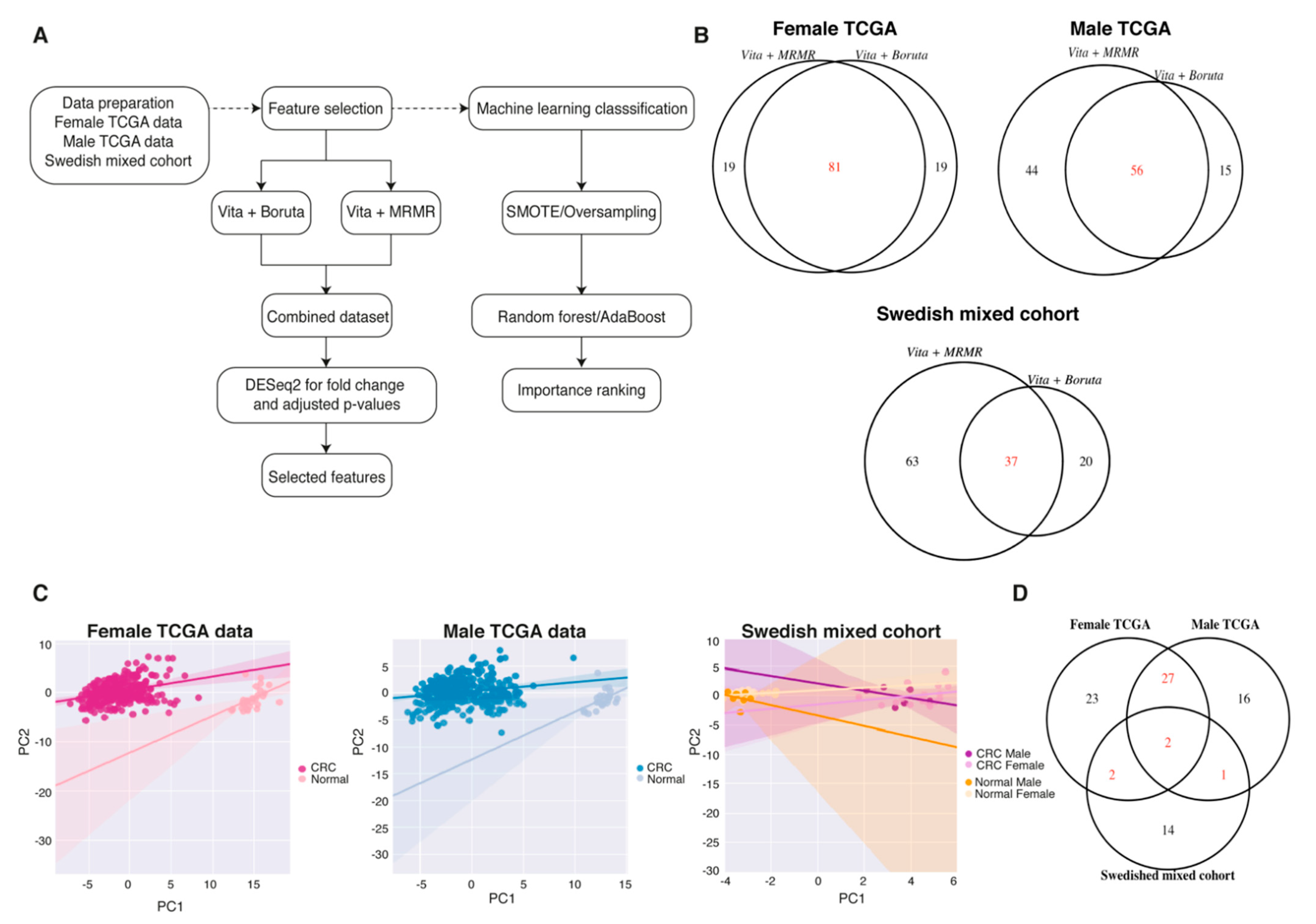
2.4. Sex-Specific Features Independent of the Imbalanced Data
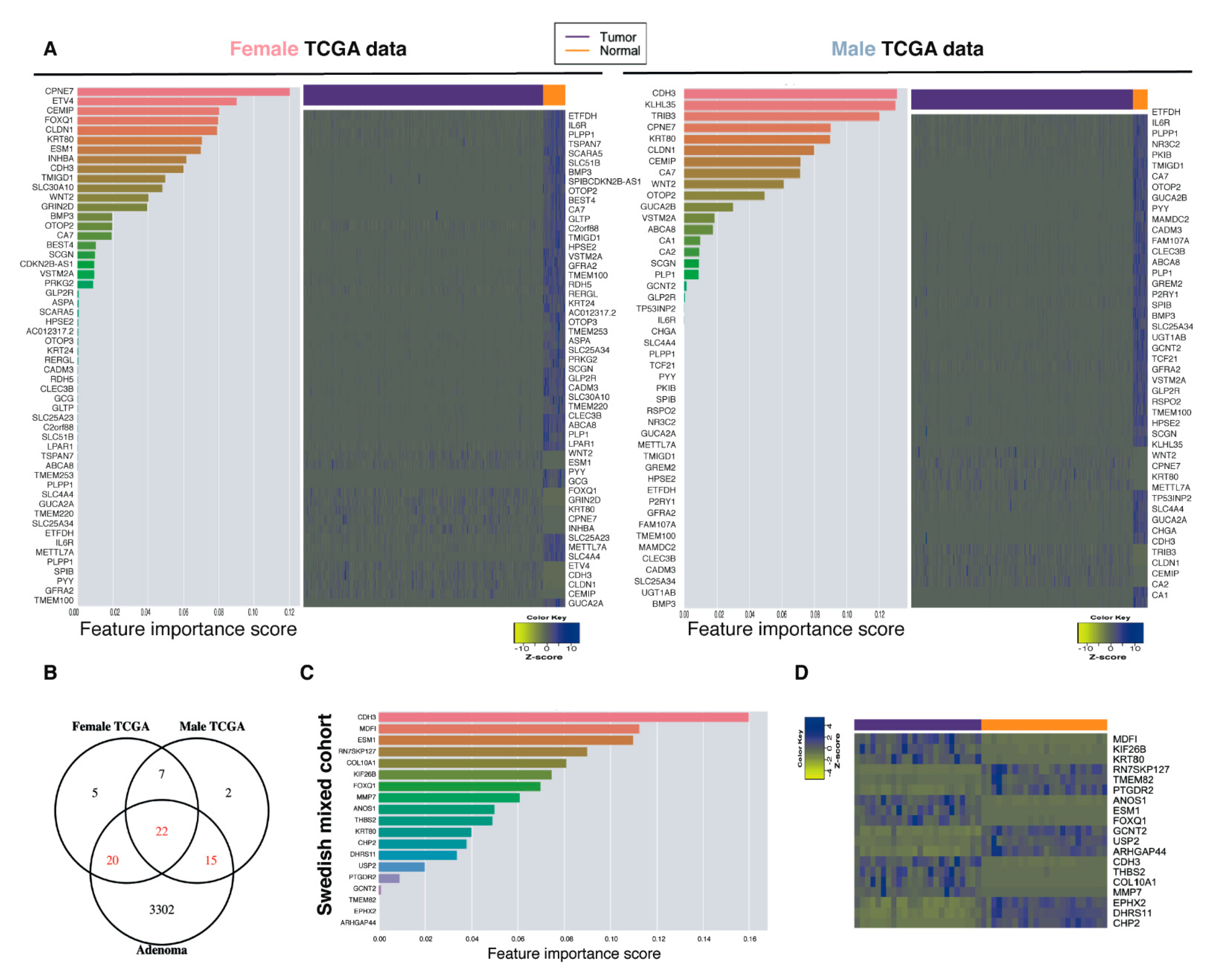
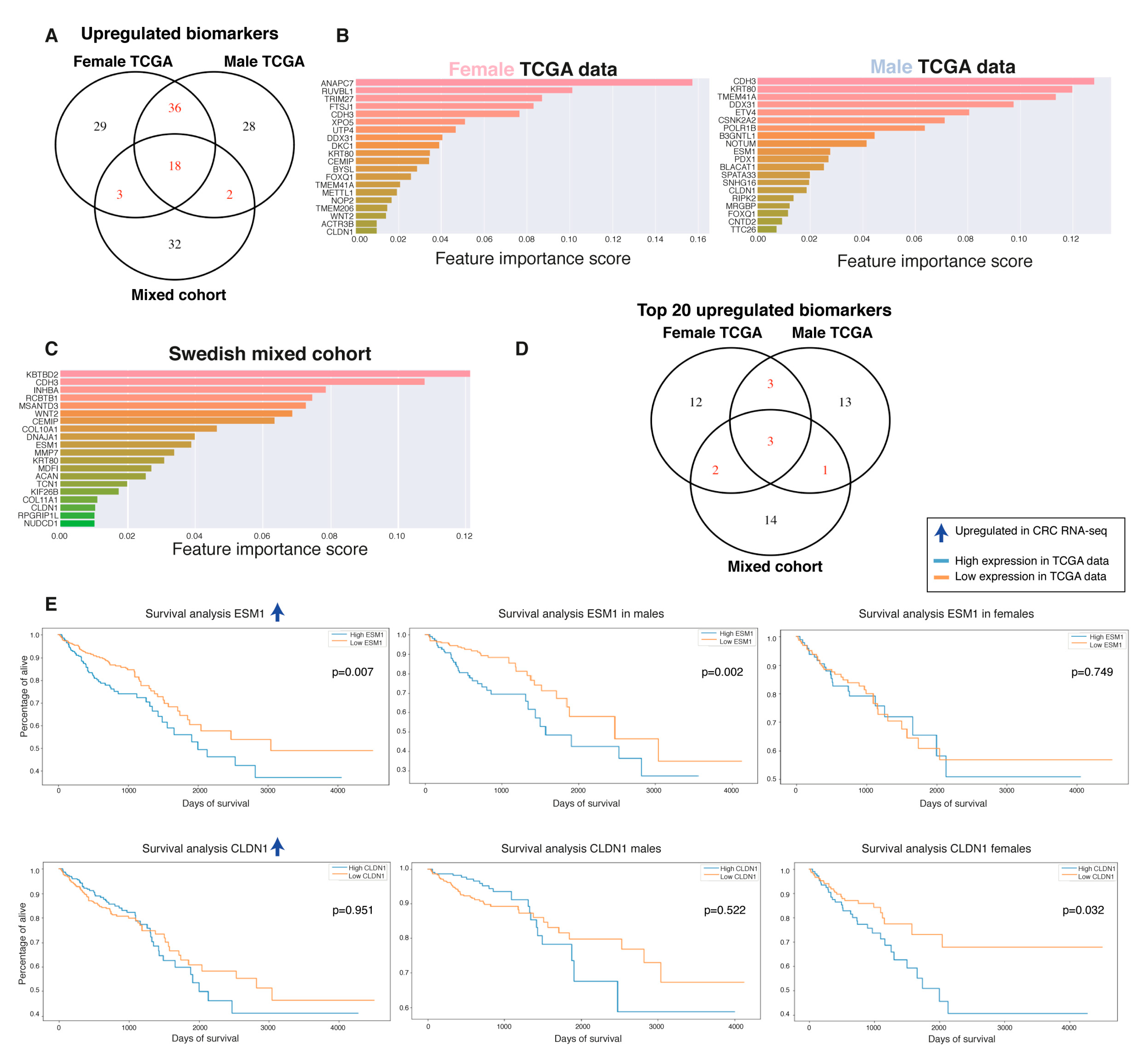
2.5. Biomarker Discovery Revealed Common and Sex-Specific CRC Biomarkers
2.6. Top-Ranked Common and Sex-Specific Biomarkers
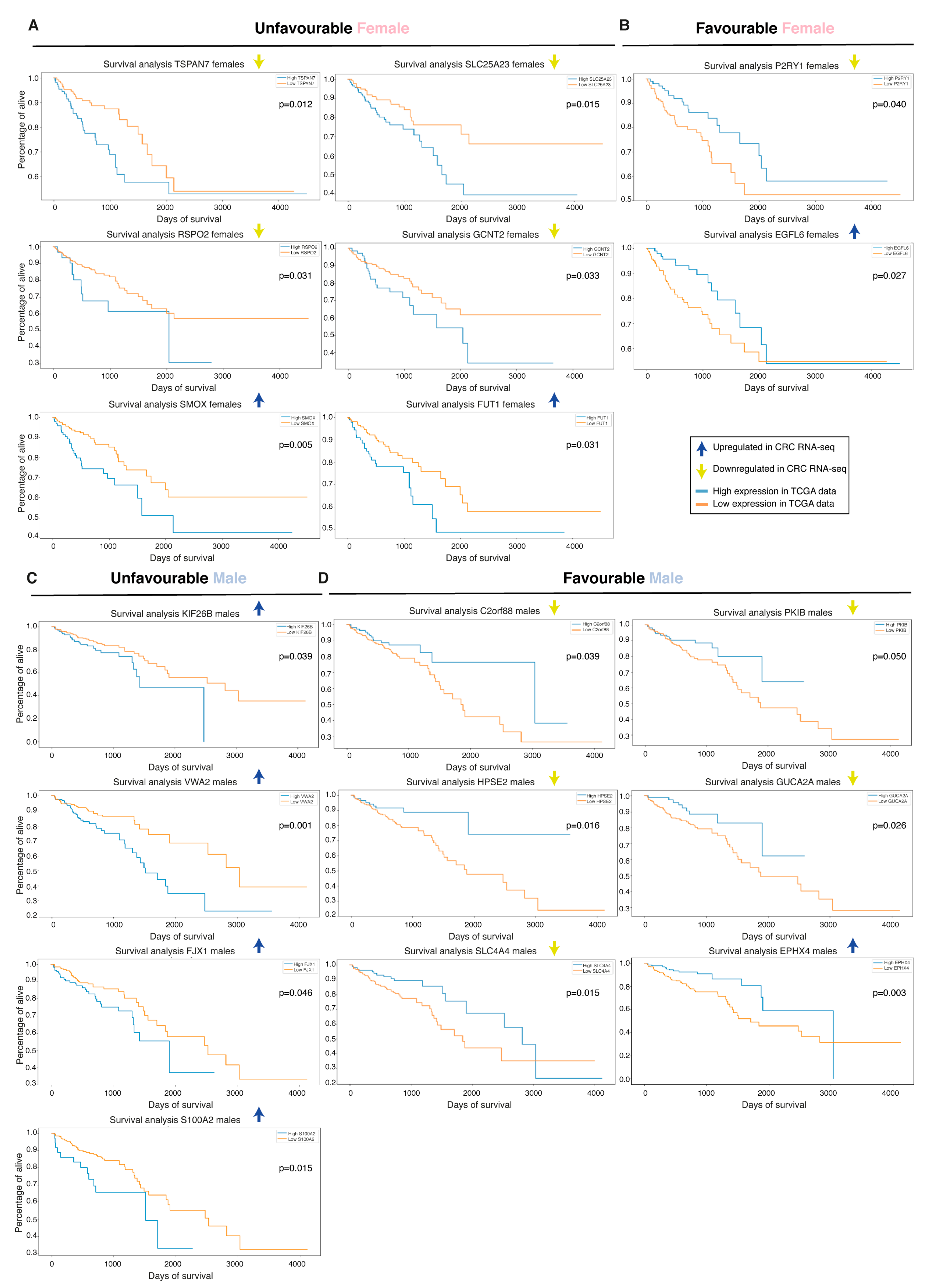
2.7. Biomarkers Have Sex-Specific Prognostic Value
3. Discussion
4. Materials and Methods
Patients and Samples
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Mathers, C.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Estimating the global cancer incidence and mortality in 2018: GLOBOCAN sources and methods. Int. J. Cancer 2019, 144, 1941–1953. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.D.; Nogueira, L.; Mariotto, A.B.; Rowland, J.H.; Yabroff, K.R.; Alfano, C.M.; Jemal, A.; Kramer, J.L.; Siegel, R.L. Cancer treatment and survivorship statistics, 2019. CA Cancer J. Clin. 2019, 69, 363–385. [Google Scholar] [CrossRef]
- Baxter, N.N.; Goldwasser, M.A.; Paszat, L.F.; Saskin, R.; Urbach, D.R.; Rabeneck, L. Association of colonoscopy and death from colorectal cancer. Ann. Intern. Med. 2009, 150, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Adler, A.; Geiger, S.; Keil, A.; Bias, H.; Schatz, P.; deVos, T.; Dhein, J.; Zimmermann, M.; Tauber, R.; Wiedenmann, B. Improving compliance to colorectal cancer screening using blood and stool based tests in patients refusing screening colonoscopy in Germany. BMC Gastroenterol. 2014, 14, 183. [Google Scholar] [CrossRef] [PubMed]
- Long, N.P.; Park, S.; Anh, N.H.; Nghi, T.D.; Yoon, S.J.; Park, J.H.; Lim, J.; Kwon, S.W. High-Throughput Omics and Statistical Learning Integration for the Discovery and Validation of Novel Diagnostic Signatures in Colorectal Cancer. Int. J. Mol. Sci. 2019, 20, 296. [Google Scholar] [CrossRef] [PubMed]
- Long, N.P.; Jung, K.H.; Yoon, S.J.; Anh, N.H.; Nghi, T.D.; Kang, Y.P.; Yan, H.H.; Min, J.E.; Hong, S.S.; Kwon, S.W. Systematic assessment of cervical cancer initiation and progression uncovers genetic panels for deep learning-based early diagnosis and proposes novel diagnostic and prognostic biomarkers. Oncotarget 2017, 8, 109436–109456. [Google Scholar] [CrossRef]
- Kim, S.E.; Paik, H.Y.; Yoon, H.; Lee, J.E.; Kim, N.; Sung, M.K. Sex- and gender-specific disparities in colorectal cancer risk. World J. Gastroenterol. 2015, 21, 5167–5175. [Google Scholar] [CrossRef]
- Hendifar, A.; Yang, D.; Lenz, F.; Lurje, G.; Pohl, A.; Lenz, C.; Ning, Y.; Zhang, W.; Lenz, H.J. Gender disparities in metastatic colorectal cancer survival. Clin. Cancer Res. 2009, 15, 6391–6397. [Google Scholar] [CrossRef]
- Pal, S.K.; Hurria, A. Impact of age, sex, and comorbidity on cancer therapy and disease progression. J. Clin. Oncol. 2010, 28, 4086–4093. [Google Scholar] [CrossRef]
- Hansen, I.O.; Jess, P. Possible better long-term survival in left versus right-sided colon cancer—A systematic review. Dan. Med. J. 2012, 59, A4444. [Google Scholar]
- Tsai, Y.J.; Huang, S.C.; Lin, H.H.; Lin, C.C.; Lan, Y.T.; Wang, H.S.; Yang, S.H.; Jiang, J.K.; Chen, W.S.; Lin, T.C.; et al. Differences in gene mutations according to gender among patients with colorectal cancer. World J. Surg. Oncol. 2018, 16, 128. [Google Scholar] [CrossRef] [PubMed]
- Hases, L.; Archer, A.; Indukuri, R.; Birgersson, M.; Savva, C.; Korach-André, M.; Williams, C. High-fat diet and estrogen impacts the colon and its transcriptome in a sex-dependent manner. Sci. Rep. 2020, 10, 16160. [Google Scholar] [PubMed]
- DeCosse, J.J.; Ngoi, S.S.; Jacobson, J.S.; Cennerazzo, W.J. Gender and colorectal cancer. Eur. J. Cancer Prev. 1993, 2, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Hases, L.; Indukuri, R.; Birgersson, M.; Nguyen-Vu, T.; Lozano, R.; Saxena, A.; Hartman, J.; Frasor, J.; Gustafsson, J.; Katajisto, P.; et al. Intestinal estrogen receptor beta suppresses colon inflammation and tumorigenesis in both sexes. Cancer Lett. 2020, 492, 54–62. [Google Scholar]
- Luo, T.; Wu, S.; Shen, X.; Li, L. Network cluster analysis of protein-protein interaction network identified biomarker for early onset colorectal cancer. Mol. Biol. Rep. 2013, 40, 6561–6568. [Google Scholar] [CrossRef]
- Fink, S.P.; Myeroff, L.L.; Kariv, R.; Platzer, P.; Xin, B.; Mikkola, D.; Lawrence, E.; Morris, N.; Nosrati, A.; Willson, J.K.; et al. Induction of KIAA1199/CEMIP is associated with colon cancer phenotype and poor patient survival. Oncotarget 2015, 6, 30500–30515. [Google Scholar] [CrossRef]
- Phipps, A.I.; Limburg, P.J.; Baron, J.A.; Burnett-Hartman, A.N.; Weisenberger, D.J.; Laird, P.W.; Sinicrope, F.A.; Rosty, C.; Buchanan, D.D.; Potter, J.D.; et al. Association between molecular subtypes of colorectal cancer and patient survival. Gastroenterology 2015, 148, 77–87.e2. [Google Scholar] [CrossRef]
- Jass, J.R. Classification of colorectal cancer based on correlation of clinical, morphological and molecular features. Histopathology 2007, 50, 113–130. [Google Scholar] [CrossRef]
- Cai, Y.; Rattray, N.J.W.; Zhang, Q.; Mironova, V.; Santos-Neto, A.; Hsu, K.S.; Rattray, Z.; Cross, J.R.; Zhang, Y.; Paty, P.B.; et al. Sex Differences in Colon Cancer Metabolism Reveal A Novel Subphenotype. Sci. Rep. 2020, 10, 4905. [Google Scholar] [CrossRef]
- Botteri, E.; Stoer, N.C.; Sakshaug, S.; Graff-Iversen, S.; Vangen, S.; Hofvind, S.; de Lange, T.; Bagnardi, V.; Ursin, G.; Weiderpass, E. Menopausal hormone therapy and colorectal cancer: A linkage between nationwide registries in Norway. BMJ Open 2017, 7, e017639. [Google Scholar] [CrossRef]
- Grodstein, F.; Newcomb, P.A.; Stampfer, M.J. Postmenopausal hormone therapy and the risk of colorectal cancer: A review and meta-analysis. Am. J. Med. 1999, 106, 574–582. [Google Scholar] [CrossRef]
- Lobo, R.A. Hormone-replacement therapy: Current thinking. Nat. Rev. Endocrinol. 2017, 13, 220–231. [Google Scholar] [CrossRef]
- Bianchi, I.; Lleo, A.; Gershwin, M.E.; Invernizzi, P. The X chromosome and immune associated genes. J. Autoimmun. 2012, 38, J187–J192. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, S.; Miyoshi, N.; Ishii, H.; Mimori, K.; Tanaka, F.; Sekimoto, M.; Doki, Y.; Mori, M. Expression of CLDN1 in colorectal cancer: A novel marker for prognosis. Int. J. Oncol. 2011, 39, 791–796. [Google Scholar] [PubMed]
- Cherradi, S.; Ayrolles-Torro, A.; Vezzo-Vie, N.; Gueguinou, N.; Denis, V.; Combes, E.; Boissiere, F.; Busson, M.; Canterel-Thouennon, L.; Mollevi, C.; et al. Antibody targeting of claudin-1 as a potential colorectal cancer therapy. J. Exp. Clin. Cancer Res. 2017, 36, 89. [Google Scholar] [CrossRef]
- Kang, Y.H.; Ji, N.Y.; Han, S.R.; Lee, C.I.; Kim, J.W.; Yeom, Y.I.; Kim, Y.H.; Chun, H.K.; Kim, J.W.; Chung, J.W.; et al. ESM-1 regulates cell growth and metastatic process through activation of NF-kappaB in colorectal cancer. Cell Signal. 2012, 24, 1940–1949. [Google Scholar]
- Kim, J.H.; Park, M.Y.; Kim, C.N.; Kim, K.H.; Kang, H.B.; Kim, K.D.; Kim, J.W. Expression of endothelial cell-specific molecule-1 regulated by hypoxia inducible factor-1alpha in human colon carcinoma: Impact of ESM-1 on prognosis and its correlation with clinicopathological features. Oncol. Rep. 2012, 28, 1701–1708. [Google Scholar] [CrossRef]
- Al-Greene, N.T.; Means, A.L.; Lu, P.; Jiang, A.; Schmidt, C.R.; Chakravarthy, A.B.; Merchant, N.B.; Washington, M.K.; Zhang, B.; Shyr, Y.; et al. Four jointed box 1 promotes angiogenesis and is associated with poor patient survival in colorectal carcinoma. PLoS ONE 2013, 8, e69660. [Google Scholar] [CrossRef]
- Wilson, C.; Lin, J.E.; Li, P.; Snook, A.E.; Gong, J.; Sato, T.; Liu, C.; Girondo, M.A.; Rui, H.; Hyslop, T.; et al. The paracrine hormone for the GUCY2C tumor suppressor, guanylin, is universally lost in colorectal cancer. Cancer Epidemiol. Biomark. Prev. 2014, 23, 2328–2337. [Google Scholar] [CrossRef]
- Zhang, H.; Du, Y.; Wang, Z.; Lou, R.; Wu, J.; Feng, J. Integrated Analysis of Oncogenic Networks in Colorectal Cancer Identifies GUCA2A as a Molecular Marker. Biochem. Res. Int. 2019, 2019, 6469420. [Google Scholar] [CrossRef]
- Li, P.; Schulz, S.; Bombonati, A.; Palazzo, J.P.; Hyslop, T.M.; Xu, Y.; Baran, A.A.; Siracusa, L.D.; Pitari, G.M.; Waldman, S.A. Guanylyl cyclase C suppresses intestinal tumorigenesis by restricting proliferation and maintaining genomic integrity. Gastroenterology 2007, 133, 599–607. [Google Scholar] [CrossRef]
- Bryant, A.P.; Busby, R.W.; Bartolini, W.P.; Cordero, E.A.; Hannig, G.; Kessler, M.M.; Pierce, C.M.; Solinga, R.M.; Tobin, J.V.; Mahajan-Miklos, S.; et al. Linaclotide is a potent and selective guanylate cyclase C agonist that elicits pharmacological effects locally in the gastrointestinal tract. Life Sci. 2010, 86, 760–765. [Google Scholar] [CrossRef]
- Li, C.; Chen, Q.; Zhou, Y.; Niu, Y.; Wang, X.; Li, X.; Zheng, H.; Wei, T.; Zhao, L.; Gao, H. S100A2 promotes glycolysis and proliferation via GLUT1 regulation in colorectal cancer. FASEB J. 2020. [Google Scholar] [CrossRef]
- Masuda, T.; Ishikawa, T.; Mogushi, K.; Okazaki, S.; Ishiguro, M.; Iida, S.; Mizushima, H.; Tanaka, H.; Uetake, H.; Sugihara, K. Overexpression of the S100A2 protein as a prognostic marker for patients with stage II and III colorectal cancer. Int. J. Oncol. 2016, 48, 975–982. [Google Scholar] [CrossRef]
- Niu, B.; Ye, K.; Zhang, Q.; Lu, C.; Xie, M.; McLellan, M.D.; Wendl, M.C.; Ding, L. MSIsensor: Microsatellite instability detection using paired tumor-normal sequence data. Bioinformatics 2014, 30, 1015–1016. [Google Scholar] [CrossRef]
- Robinson, J.T.; Thorvaldsdóttir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative genomics viewer. Nat. Biotechnol. 2011, 29, 24–26. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cohort | Original Feature Numbers | Feature Selection Methods | Selected Features | Features in Common | Differentially Expressed Features |
|---|---|---|---|---|---|
| Female TCGA data | 56,719 | Vita + Boruta | 100 | 81 | 54 |
| Vita + MRMR | 100 | ||||
| Male TCGA data | 56,719 | Vita + Boruta | 71 | 56 | 46 |
| Vita + MRMR | 100 | ||||
| Swedish mixed | 63,678 | Vita + Boruta | 57 | 37 | 19 |
| Vita + MRMR | 100 |
| Biomarker | Cohort 1 | Rank 2 | Regulation 3 | Prognostic Value 4 |
|---|---|---|---|---|
| ESM1 | All | Top20 | Up | Unfavorable in males |
| CLDN1 | All | Top20 | Up | Unfavorable in females |
| TSPAN7 | Female TCGA | 39 | Down | Unfavorable in females |
| SLC25A23 | Female TCGA | 35 | Down | Unfavorable in females |
| C2orf88 | Female TCGA | 36 | Down | Favorable in males |
| PKIB | Male TCGA | 27 | Down | Favorable in males |
| P2RYI | Male TCGA | 37 | Down | Favorable in females |
| RSPO2 | Male TCGA | 29 | Down | Unfavorable in females |
| GCNT2 | Male TCGA and Swedish | Top20 | Down | Unfavorable in females |
| HPSE2 | TCGA | 35 M and 25 F | Down | Favorable in males |
| GUCA2A | TCGA | 31 M and 44 F | Down | Favorable in males |
| SLC4A4 | TCGA | 23 M and 43 F | Down | Favorable in males |
| KIF26B | Swedish | 6 | Up | Unfavorable in males |
| PTGDR2 | Swedish | 15 | Down | Favorable in males and females (combined) |
| ASPA | Female TCGA | 23 | Down | Unfavorable in males and females (combined) |
| BEST4 | Female TCGA | 17 | Down | Favorable in males and females (combined) |
| NR3C2 | Male TCGA | 30 | Down | Favorable in males and females (combined) |
| SMOX | Male TCGA | 37 | Up | Unfavorable in females |
| FUT1 | All | 38 M, 78 F and 27 S | Up | Unfavorable in females |
| EGFL6 | Female TCGA | 27 | Up | Favorable in females |
| VWA2 | Male TCGA | 31 | Up | Unfavorable in males |
| FJX1 | TCGA | 67 M and 83 F | Up | Unfavorable in males |
| S100A2 | TCGA | 64 M and 45 F | Up | Unfavorable in males |
| EPHX4 | Female TCGA | 75 | Up | Favorable in females |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hases, L.; Ibrahim, A.; Chen, X.; Liu, Y.; Hartman, J.; Williams, C. The Importance of Sex in the Discovery of Colorectal Cancer Prognostic Biomarkers. Int. J. Mol. Sci. 2021, 22, 1354. https://doi.org/10.3390/ijms22031354
Hases L, Ibrahim A, Chen X, Liu Y, Hartman J, Williams C. The Importance of Sex in the Discovery of Colorectal Cancer Prognostic Biomarkers. International Journal of Molecular Sciences. 2021; 22(3):1354. https://doi.org/10.3390/ijms22031354
Chicago/Turabian StyleHases, Linnea, Ahmed Ibrahim, Xinsong Chen, Yanghong Liu, Johan Hartman, and Cecilia Williams. 2021. "The Importance of Sex in the Discovery of Colorectal Cancer Prognostic Biomarkers" International Journal of Molecular Sciences 22, no. 3: 1354. https://doi.org/10.3390/ijms22031354
APA StyleHases, L., Ibrahim, A., Chen, X., Liu, Y., Hartman, J., & Williams, C. (2021). The Importance of Sex in the Discovery of Colorectal Cancer Prognostic Biomarkers. International Journal of Molecular Sciences, 22(3), 1354. https://doi.org/10.3390/ijms22031354