
Filifactor alocis and Tumor Necrosis Factor-Alpha Stimulate Synthesis of Visfatin by Human Macrophages

 , , ,
, , ,  , , ,
, , ,
Abstract
1. Introduction
2. Results
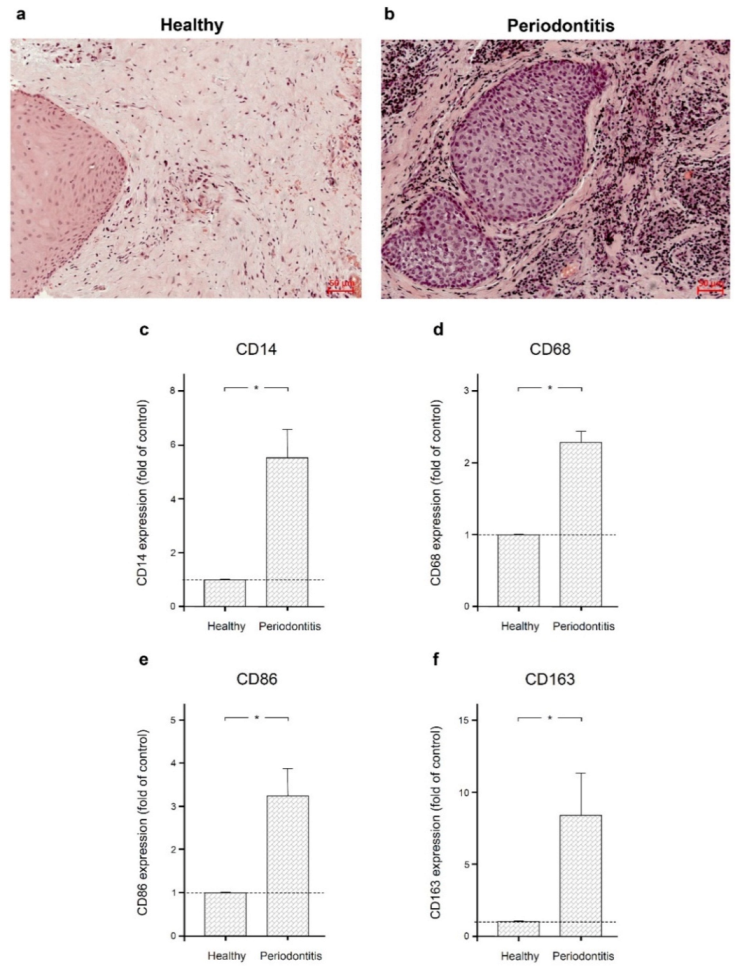
2.1. Macrophage Infiltration in Human Gingival Tissues from Sites of Periodontitis
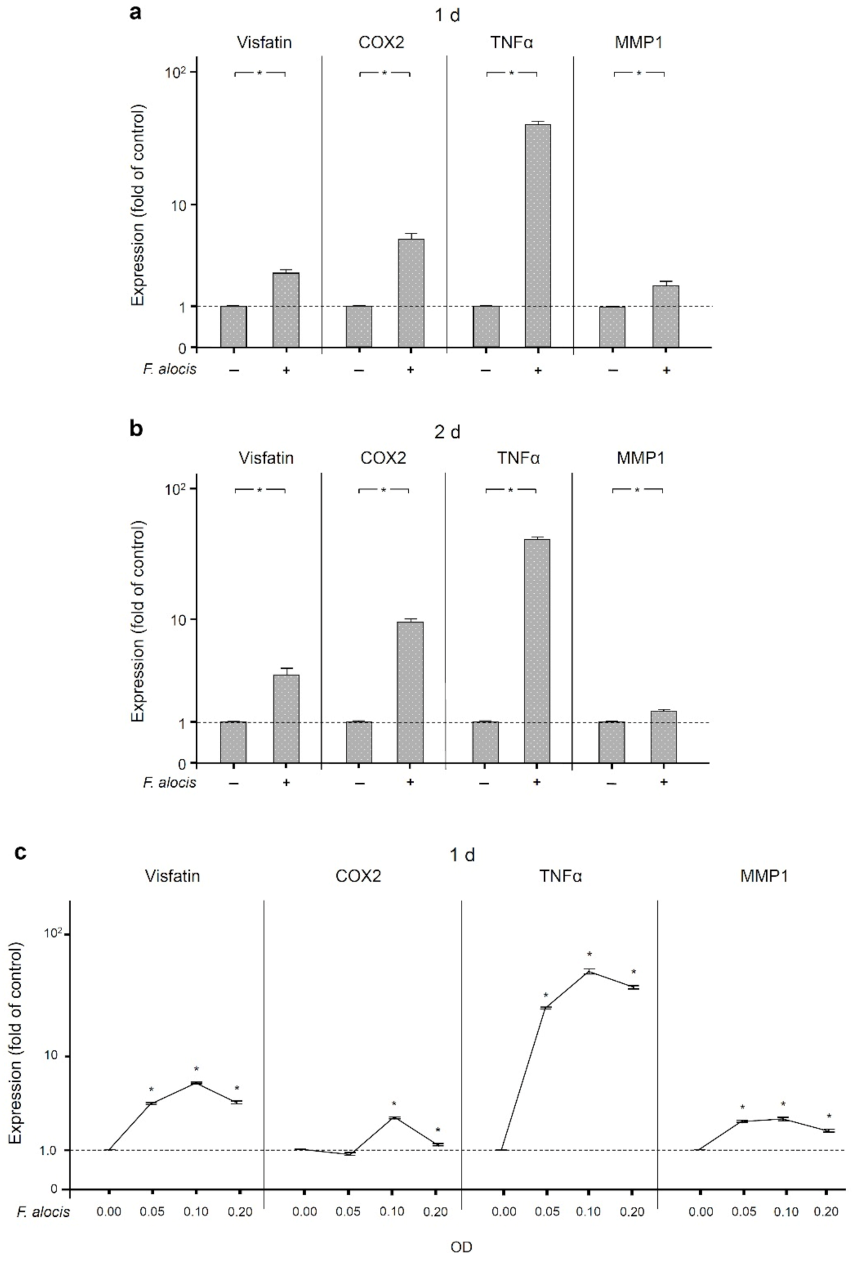
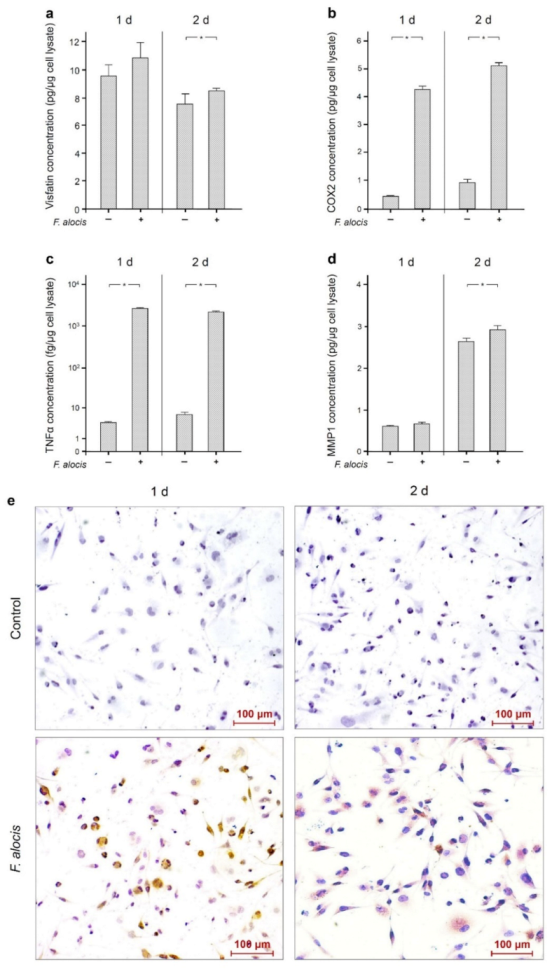
2.2. Stimulatory Effects of F. alocis on Human Macrophages
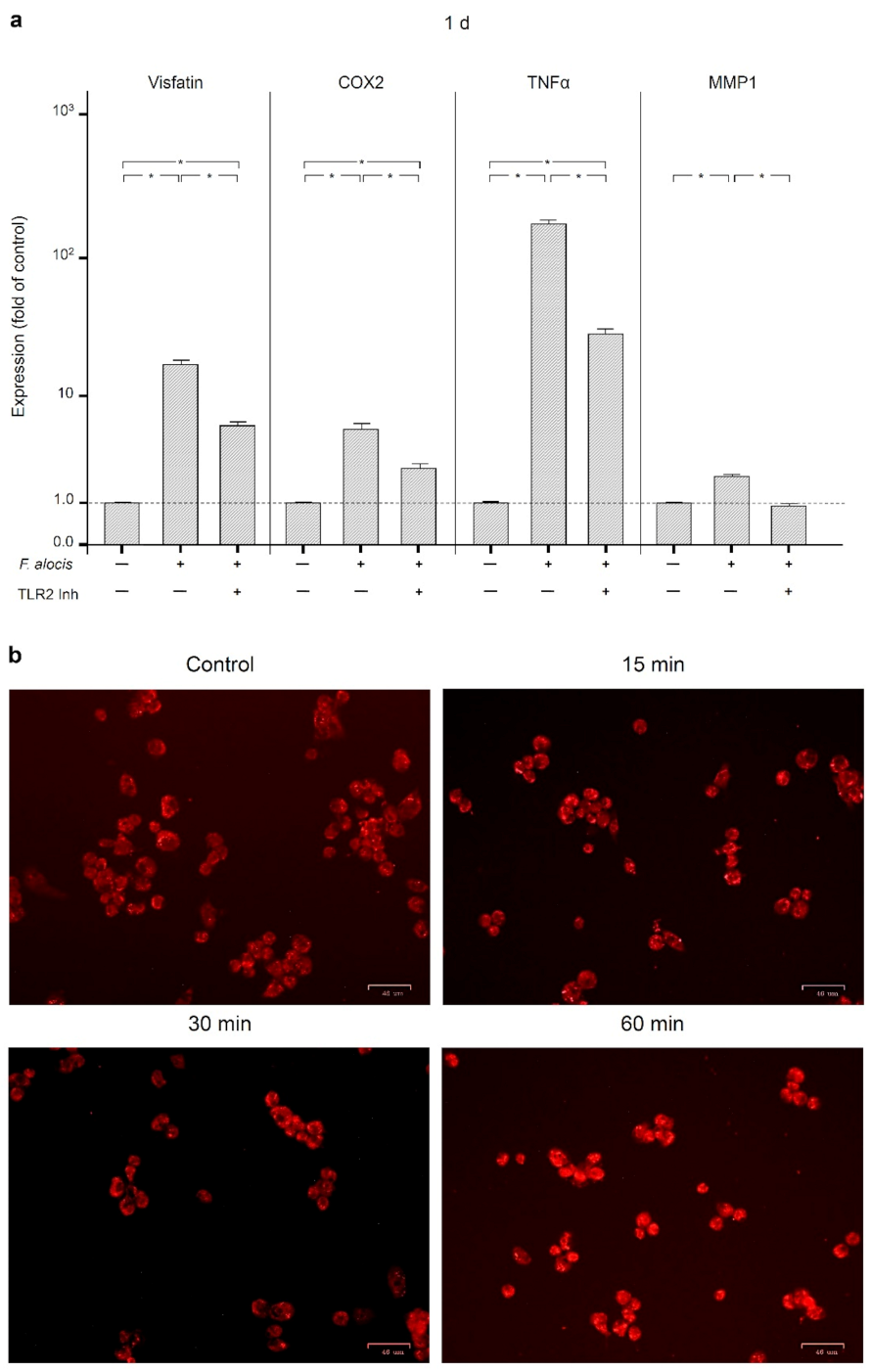
2.3. Involvement of Toll-Like Receptor 2 and Intracellular Pathways in the Actions of F. alocis
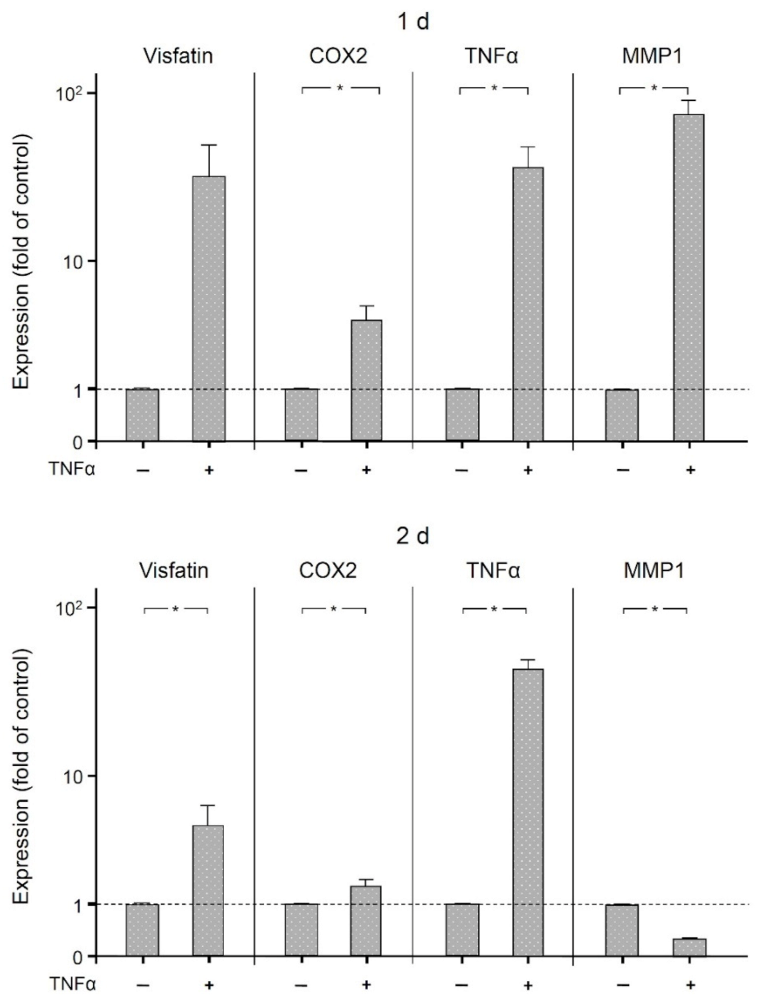
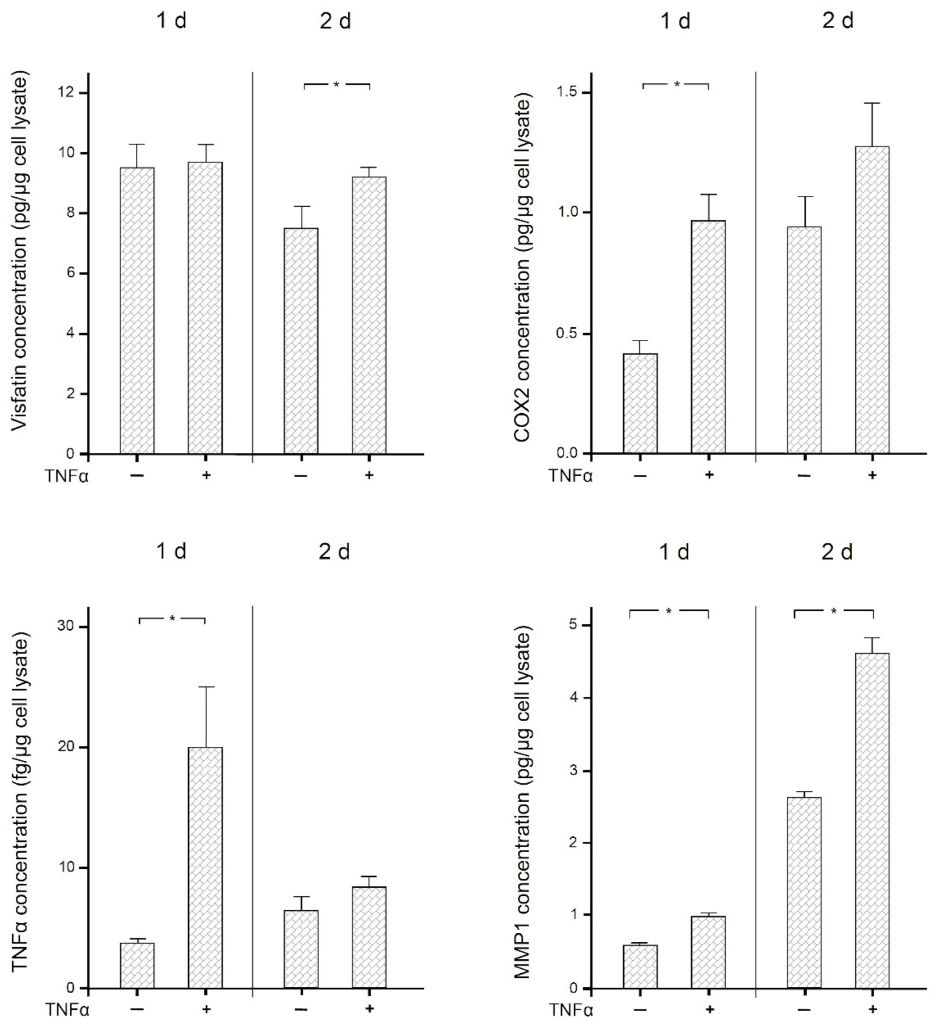
2.4. Stimulatory Effects of TNFα on Human Macrophages
3. Discussion
4. Materials and Methods
4.1. Culture and Treatment of Cells
4.2. Human Gingival Biopsies
4.3. Real-Time PCR
4.4. ELISA
4.5. Immunocytochemistry
4.6. Immunofluorescence
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ATCC | American Type Culture Collection |
| cDNA | complementary desoxyribonucleic acid |
| COX2 | cyclooxygenase-2 |
| ELISA | enzyme-linked immunosorbent assay |
| FBS | fetal bovine serum |
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase |
| GCF | gingival crevicular fluid |
| HRP | horseradish peroxidase |
| JNK | c-Jun N-terminal kinases |
| MAPK | mitogen-activated protein kinase |
| MEK1/2 | MAP/ERK kinases 1/2 |
| MMP1 | matrix metalloproteinase 1 |
| NF-κB | nuclear factor kappa B |
| OD | optical density |
| PCR | polymerase chain reaction |
| PDL | periodontal ligament |
| PGE2 | prostaglandin E2 |
| PMA | phorbol 12-myristate 13-acetate |
| RNA | ribonucleic acid |
| TLR2 | toll-like receptor 2 |
| TNFα | tumor necrosis factor-alpha |
References
- Papapanou, P.N.; Sanz, M.; Buduneli, N.; Dietrich, T.; Feres, M.; Fine, D.H.; Flemmig, T.F.; Garcia, R.; Giannobile, W.V.; Graziani, F.; et al. Periodontitis: Consensus report of workgroup 2 of the 2017 World Workshop on the Classification of Periodontal and Peri-Implant Diseases and Conditions. J. Periodontol. 2018, 89 (Suppl. 1), S173–S182. [Google Scholar] [CrossRef] [PubMed]
- Meyle, J.; Chapple, I. Molecular aspects of the pathogenesis of periodontitis. Periodontology 2000 2015, 69, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Hajishengallis, G.; Chavakis, T.; Lambris, J.D. Current understanding of periodontal disease pathogenesis and targets for host-modulation therapy. Periodontology 2000 2020, 84, 14–34. [Google Scholar] [CrossRef] [PubMed]
- Silva, N.; Abusleme, L.; Bravo, D.; Dutzan, N.; Garcia-Sesnich, J.; Vernal, R.; Hernández, M.; Gamonal, J. Host response mechanisms in periodontal diseases. J. Appl. Oral Sci. 2015, 23, 329–355. [Google Scholar] [CrossRef]
- Sima, C.; Viniegra, A.; Glogauer, M. Macrophage immunomodulation in chronic osteolytic diseases-the case of periodontitis. J Leukoc. Biol. 2019, 105, 473–487. [Google Scholar] [CrossRef]
- Wynn, T.A.; Chawla, A.; Pollard, J.W. Macrophage biology in development, homeostasis and disease. Nature 2013, 496, 445–455. [Google Scholar] [CrossRef]
- Sinder, B.P.; Pettit, A.R.; McCauley, L.K. Macrophages: Their Emerging Roles in Bone. J. Bone Miner Res. 2015, 30, 2140–2149. [Google Scholar] [CrossRef]
- Fortunato, L.; Bennardo, F.; Buffone, C.; Giudice, A. Is the application of platelet concentrates effective in the prevention and treatment of medication-related osteonecrosis of the jaw? A systematic review. J. Craniomaxillofac. Surg. 2020, 48, 268–285. [Google Scholar] [CrossRef]
- Leira, Y.; Seoane, J.; Blanco, M.; Rodríguez-Yáñez, M.; Takkouche, B.; Blanco, J.; Castillo, J. Association between periodontitis and ischemic stroke: A systematic review and meta-analysis. Eur. J. Epidemiol. 2017, 32, 43–53. [Google Scholar] [CrossRef]
- Humphrey, L.L.; Fu, R.; Buckley, D.I.; Freeman, M.; Helfand, M. Periodontal disease and coronary heart disease incidence: A systematic review and meta-analysis. J. Gen. Intern. Med. 2008, 23, 2079–2086. [Google Scholar] [CrossRef]
- Chávarry, N.G.; Vettore, M.V.; Sansone, C.; Sheiham, A. The relationship between diabetes mellitus and destructive periodontal disease: A meta-analysis. Oral Health Prev. Dent. 2009, 7, 107–127. [Google Scholar] [PubMed]
- Chaffee, B.W.; Weston, S.J. Association between chronic periodontal disease and obesity: A systematic review and meta-analysis. J. Periodontol. 2010, 81, 1708–1724. [Google Scholar] [CrossRef] [PubMed]
- Moura-Grec, P.G.; Marsicano, J.A.; Carvalho, C.A.; Sales-Peres, S.H. Obesity and periodontitis: Systematic review and meta-analysis. Cien Saude Colet 2014, 19, 1763–1772. [Google Scholar] [CrossRef] [PubMed]
- Jepsen, S.; Suvan, J.; Deschner, J. The association of periodontal diseases with metabolic syndrome and obesity. Periodontology 2000 2020, 83, 125–153. [Google Scholar] [CrossRef] [PubMed]
- Deschner, J.; Eick, S.; Damanaki, A.; Nokhbehsaim, M. The role of adipokines in periodontal infection and healing. Mol. Oral Microbiol. 2014, 29, 258–269. [Google Scholar] [CrossRef]
- Boutens, L.; Stienstra, R. Adipose tissue macrophages: Going off track during obesity. Diabetologia 2016, 59, 879–894. [Google Scholar] [CrossRef]
- Stastny, J.; Bienertova-Vasku, J.; Vasku, A. Visfatin and its role in obesity development. Diabetes Metab. Syndr. 2012, 6, 120–124. [Google Scholar] [CrossRef]
- Chang, Y.H.; Chang, D.M.; Lin, K.C.; Shin, S.J.; Lee, Y.J. Visfatin in overweight/obesity, type 2 diabetes mellitus, insulin resistance, metabolic syndrome and cardiovascular diseases: A meta-analysis and systemic review. Diabetes Metab. Res. Rev. 2011, 27, 515–527. [Google Scholar] [CrossRef]
- Taşkesen, D.; Kirel, B.; Us, T. Serum visfatin levels, adiposity and glucose metabolism in obese adolescents. J. Clin. Res. Pediatr. Endocrinol. 2012, 4, 76–81. [Google Scholar] [CrossRef]
- Zhang, L.Q.; Heruth, D.P.; Ye, S.Q. Nicotinamide Phosphoribosyltransferase in Human Diseases. J. Bioanal. Biomed. 2011, 3, 13–25. [Google Scholar] [CrossRef]
- Raghavendra, N.M.; Pradeep, A.R.; Kathariya, R.; Sharma, A.; Rao, N.S.; Naik, S.B. Effect of non-surgical periodontal therapy on gingival crevicular fluid and serum visfatin concentration in periodontal health and disease. Dis. Markers 2012, 32, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Damanaki, A.; Nokhbehsaim, M.; Eick, S.; Götz, W.; Winter, J.; Wahl, G.; Jäger, A.; Jepsen, S.; Deschner, J. Regulation of NAMPT in human gingival fibroblasts and biopsies. Mediators Inflamm. 2014, 2014, 912821. [Google Scholar] [CrossRef] [PubMed]
- Tabari, Z.A.; Azadmehr, A.; Nohekhan, A.; Naddafpour, N.; Ghaedi, F.B. Salivary visfatin concentrations in patients with chronic periodontitis. J. Periodontol. 2014, 85, 1081–1085. [Google Scholar] [CrossRef] [PubMed]
- Nokhbehsaim, M.; Eick, S.; Nogueira, A.V.; Hoffmann, P.; Herms, S.; Fröhlich, H.; Jepsen, S.; Jäger, A.; Cirelli, J.A.; Deschner, J. Stimulation of MMP-1 and CCL2 by NAMPT in PDL cells. Mediators Inflamm. 2013, 2013, 437123. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, A.V.; Nokhbehsaim, M.; Eick, S.; Bourauel, C.; Jäger, A.; Jepsen, S.; Cirelli, J.A.; Deschner, J. Regulation of visfatin by microbial and biomechanical signals in PDL cells. Clin. Oral Investig. 2014, 18, 171–178. [Google Scholar] [CrossRef]
- Nokhbehsaim, M.; Keser, S.; Jäger, A.; Jepsen, S.; Deschner, J. Regulation of regenerative periodontal healing by NAMPT. Mediators Inflamm. 2013, 2013, 202530. [Google Scholar] [CrossRef]
- Kilian, M.; Chapple, I.L.; Hannig, M.; Marsh, P.D.; Meuric, V.; Pedersen, A.M.; Tonetti, M.S.; Wade, W.G.; Zaura, E. The oral microbiome-an update for oral healthcare professionals. Br. Dent J. 2016, 221, 657–666. [Google Scholar] [CrossRef]
- Belstrøm, D.; Fiehn, N.E.; Nielsen, C.H.; Kirkby, N.; Twetman, S.; Klepac-Ceraj, V.; Paster, B.J.; Holmstrup, P. Differences in bacterial saliva profile between periodontitis patients and a control cohort. J. Clin. Periodontol. 2014, 41, 104–112. [Google Scholar] [CrossRef]
- Oliveira, R.R.; Fermiano, D.; Feres, M.; Figueiredo, L.C.; Teles, F.R.; Soares, G.M.; Faveri, M. Levels of Candidate Periodontal Pathogens in Subgingival Biofilm. J. Dent Res. 2016, 95, 711–718. [Google Scholar] [CrossRef]
- Aruni, W.; Chioma, O.; Fletcher, H.M. Filifactor alocis: The Newly Discovered Kid on the Block with Special Talents. J. Dent. Res. 2014, 93, 725–732. [Google Scholar] [CrossRef]
- Aruni, A.W.; Roy, F.; Fletcher, H.M. Filifactor alocis has virulence attributes that can enhance its persistence under oxidative stress conditions and mediate invasion of epithelial cells by porphyromonas gingivalis. Infect Immun. 2011, 79, 3872–3886. [Google Scholar] [CrossRef] [PubMed]
- Nokhbehsaim, M.; Nogueira, A.V.B.; Nietzsche, S.; Eick, S.; Deschner, J. Regulation of Cyclooxygenase 2 by Filifactor alocis in Fibroblastic and Monocytic Cells. Mediat. Inflamm. 2020, 2020, 4185273. [Google Scholar] [CrossRef] [PubMed]
- Nokhbehsaim, M.; Nogueira, A.V.B.; Damanaki, A.; Dalagiorgou, G.; Eick, S.; Adamopoulos, C.; Piperi, C.; Basdra, E.K.; Papavassiliou, A.G.; Deschner, J. Regulation of matrix metalloproteinase-1 by Filifactor alocis in human gingival and monocytic cells. Clin Oral Investig 2020, 24, 1987–1995. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Wright, C.J.; Dingming, H.; Uriarte, S.M.; Lamont, R.J. Oral community interactions of Filifactor alocis in vitro. PLoS ONE 2013, 8, e76271. [Google Scholar] [CrossRef]
- Noguchi, K.; Ishikawa, I. The roles of cyclooxygenase-2 and prostaglandin E2 in periodontal disease. Periodontology 2000 2007, 43, 85–101. [Google Scholar] [CrossRef]
- Sorsa, T.; Tjäderhane, L.; Konttinen, Y.T.; Lauhio, A.; Salo, T.; Lee, H.M.; Golub, L.M.; Brown, D.L.; Mäntylä, P. Matrix metalloproteinases: Contribution to pathogenesis, diagnosis and treatment of periodontal inflammation. Ann. Med. 2006, 38, 306–321. [Google Scholar] [CrossRef]
- Zidar, N.; Odar, K.; Glavac, D.; Jerse, M.; Zupanc, T.; Stajer, D. Cyclooxygenase in normal human tissues-is COX-1 really a constitutive isoform, and COX-2 an inducible isoform? J. Cell Mol. Med. 2009, 13, 3753–3763. [Google Scholar] [CrossRef]
- Ricciotti, E.; FitzGerald, G.A. Prostaglandins and inflammation. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 986–1000. [Google Scholar] [CrossRef]
- Hikiji, H.; Takato, T.; Shimizu, T.; Ishii, S. The roles of prostanoids, leukotrienes, and platelet-activating factor in bone metabolism and disease. Prog. Lipid Res. 2008, 47, 107–126. [Google Scholar] [CrossRef]
- Gümüş, P.; Nizam, N.; Nalbantsoy, A.; Özçaka, Ö.; Buduneli, N. Saliva, Serum Levels of Interleukin-21, -33 and Prostaglandin E2 in Patients with Generalised Aggressive or Chronic Periodontitis. Oral Health Prev. Dent. 2017, 15, 385–390. [Google Scholar] [CrossRef]
- Kurgan, Ş.; Fentoğlu, Ö.; Önder, C.; Serdar, M.; Eser, F.; Tatakis, D.N.; Günhan, M. The effects of periodontal therapy on gingival crevicular fluid matrix metalloproteinase-8, interleukin-6 and prostaglandin E2 levels in patients with rheumatoid arthritis. J. Periodontal. Res. 2016, 51, 586–595. [Google Scholar] [CrossRef] [PubMed]
- Mesa, F.; O‘Valle, F.; Rizzo, M.; Cappello, F.; Donos, N.; Parkar, M.; Chaudhary, N.; Carini, F.; Muñoz, R.; Nibali, L. Association between COX-2 rs 6681231 genotype and interleukin-6 in periodontal connective tissue. A pilot study. PLoS ONE 2014, 9, e87023. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.K.; Reddy, N.R.; Babu, M.; Kumar, P.M.; Reddy, V.S.; Chavan, C.V. Estimation of prostaglandin E2 levels in gingival crevicular fluid in periodontal health, disease and after treatment. Contemp. Clin. Dent. 2013, 4, 303–306. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, G.A.; Miozza, V.A.; Delgado, A.; Busch, L. Salivary IL-1β and PGE2 as biomarkers of periodontal status, before and after periodontal treatment. J. Clin. Periodontol. 2013, 40, 1112–1117. [Google Scholar] [CrossRef]
- Preshaw, P.M. Host modulation therapy with anti-inflammatory agents. Periodontology 2000 2018, 76, 131–149. [Google Scholar] [CrossRef]
- Salvi, G.E.; Lang, N.P. Host response modulation in the management of periodontal diseases. J. Clin. Periodontol. 2005, 32 (Suppl. 6), 108–129. [Google Scholar] [CrossRef]
- Buduneli, N.; Vardar, S.; Atilla, G.; Sorsa, T.; Luoto, H.; Baylas, H. Gingival crevicular fluid matrix metalloproteinase-8 levels following adjunctive use of meloxicam and initial phase of periodontal therapy. J. Periodontol. 2002, 73, 103–109. [Google Scholar] [CrossRef]
- Hannas, A.R.; Pereira, J.C.; Granjeiro, J.M.; Tjäderhane, L. The role of matrix metalloproteinases in the oral environment. Acta Odontol. Scand. 2007, 65, 1–13. [Google Scholar] [CrossRef]
- Wang, J.; Yang, D.; Li, C.; Shang, S.; Xiang, J. Expression of extracellular matrix metalloproteinase inducer glycosylation and caveolin-1 in healthy and inflamed human gingiva. J. Periodontal. Res. 2014, 49, 197–204. [Google Scholar] [CrossRef]
- Popat, P.R.; Bhavsar, N.V.; Popat, P.R. Gingival crevicular fluid levels of Matrix Metalloproteinase-1 (MMP-1) and Tissue Inhibitor of Metalloproteinase-1 (TIMP-1) in periodontal health and disease. Singapore Dent J. 2014, 35, 59–64. [Google Scholar] [CrossRef]
- Alfant, B.; Shaddox, L.M.; Tobler, J.; Magnusson, I.; Aukhil, I.; Walker, C. Matrix metalloproteinase levels in children with aggressive periodontitis. J. Periodontol. 2008, 79, 819–826. [Google Scholar] [CrossRef]
- Ghodpage, P.S.; Kolte, R.A.; Kolte, A.P.; Gupta, M. Influence of phase I periodontal therapy on levels of matrix metalloproteinase 1 and tissue inhibitor of metalloproteinase 1. Saudi Dent J. 2014, 26, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Tüter, G.; Kurtiş, B.; Serdar, M. Effects of phase I periodontal treatment on gingival crevicular fluid levels of matrix metalloproteinase-1 and tissue inhibitor of metalloproteinase-1. J. Periodontol. 2002, 73, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Graves, D.T.; Cochran, D. The contribution of interleukin-1 and tumor necrosis factor to periodontal tissue destruction. J. Periodontol. 2003, 74, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B. TNF and Bone Remodeling. Curr. Osteoporos. Rep. 2017, 15, 126–134. [Google Scholar] [CrossRef]
- Madureira, D.F.; Lucas De Abreu Lima, I.; Costa, G.C.; Lages, E.M.B.; Martins, C.C.; Aparecida Da Silva, T. Tumor Necrosis Factor-alpha in Gingival Crevicular Fluid as a Diagnostic Marker for Periodontal Diseases: A Systematic Review. J. Evid. Based Dent. Pract. 2018, 18, 315–331. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, C.L.; Miralda, I.; Neff, A.C.; Tian, S.; Vashishta, A.; Perez, L.; Le, J.; Lamont, R.J.; Uriarte, S.M. Filifactor alocis Promotes Neutrophil Degranulation and Chemotactic Activity. Infect Immun. 2016, 84, 3423–3433. [Google Scholar] [CrossRef]
- Song, M.K.; Kim, H.Y.; Choi, B.K.; Kim, H.H. Filifactor alocis-derived extracellular vesicles inhibit osteogenesis through TLR2 signaling. Mol. Oral Microbiol. 2020, 35, 202–210. [Google Scholar] [CrossRef]
- Lin, X.L.; Hu, H.J.; Liu, Y.B.; Hu, X.M.; Fan, X.J.; Zou, W.W.; Pan, Y.Q.; Zhou, W.Q.; Peng, M.W.; Gu, C.H. Allicin induces the upregulation of ABCA1 expression via PPARγ/LXRα signaling in THP-1 macrophage-derived foam cells. Int. J. Mol. Med. 2017, 39, 1452–1460. [Google Scholar] [CrossRef]
- Cao, H.; Huang, Y.; Wang, L.; Wang, H.; Pang, X.; Li, K.; Dang, W.; Tang, H.; Wei, L.; Su, M.; et al. Leptin promotes migration and invasion of breast cancer cells by stimulating IL-8 production in M2 macrophages. Oncotarget 2016, 7, 65441–65453. [Google Scholar] [CrossRef]
- Lugli, S.M.; Feng, N.; Heim, M.H.; Adam, M.; Schnyder, B.; Etter, H.; Yamage, M.; Eugster, H.P.; Lutz, R.A.; Zurawski, G.; et al. Tumor necrosis factor alpha enhances the expression of the interleukin (IL)-4 receptor alpha-chain on endothelial cells increasing IL-4 or IL-13-induced Stat6 activation. J. Biol. Chem. 1997, 272, 5487–5494. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 Day | 2 Days | |||
|---|---|---|---|---|
| Control | F. alocis | Control | F. alocis | |
| Visfatin (pg/µg) | 9.52 ± 0.77 | 10.81 ± 1.10 | 7.50 ± 0.73 | 8.45 ± 0.17 1 |
| COX2 (pg/µg) | 0.42 ± 0.05 | 4.22 ± 0.11 1 | 0.94 ± 0.13 | 5.05 ± 0.10 1 |
| TNFα (fg/µg) | 3.87 ± 0.27 | 2619.51 ± 58.26 1 | 6.52 ± 1.11 | 2151.72 ± 83.37 1 |
| MMP1 (pg/µg) | 0.61 ± 0.02 | 0.67 ± 0.05 | 2.64 ± 0.07 | 2.92 ± 0.10 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nogueira, A.V.B.; Nokhbehsaim, M.; Damanaki, A.; Eick, S.; Kirschneck, C.; Schröder, A.; Jantsch, J.; Deschner, J. Filifactor alocis and Tumor Necrosis Factor-Alpha Stimulate Synthesis of Visfatin by Human Macrophages. Int. J. Mol. Sci. 2021, 22, 1235. https://doi.org/10.3390/ijms22031235
Nogueira AVB, Nokhbehsaim M, Damanaki A, Eick S, Kirschneck C, Schröder A, Jantsch J, Deschner J. Filifactor alocis and Tumor Necrosis Factor-Alpha Stimulate Synthesis of Visfatin by Human Macrophages. International Journal of Molecular Sciences. 2021; 22(3):1235. https://doi.org/10.3390/ijms22031235
Chicago/Turabian StyleNogueira, Andressa Vilas Boas, Marjan Nokhbehsaim, Anna Damanaki, Sigrun Eick, Christian Kirschneck, Agnes Schröder, Jonathan Jantsch, and James Deschner. 2021. "Filifactor alocis and Tumor Necrosis Factor-Alpha Stimulate Synthesis of Visfatin by Human Macrophages" International Journal of Molecular Sciences 22, no. 3: 1235. https://doi.org/10.3390/ijms22031235
APA StyleNogueira, A. V. B., Nokhbehsaim, M., Damanaki, A., Eick, S., Kirschneck, C., Schröder, A., Jantsch, J., & Deschner, J. (2021). Filifactor alocis and Tumor Necrosis Factor-Alpha Stimulate Synthesis of Visfatin by Human Macrophages. International Journal of Molecular Sciences, 22(3), 1235. https://doi.org/10.3390/ijms22031235