
Restraint upon Embryonic Metatarsal Ex Vivo Growth by Hydrogel Reveals Interaction between Quasi-Static Load and the mTOR Pathway

 , ,
, ,
Abstract
1. Introduction
2. Results
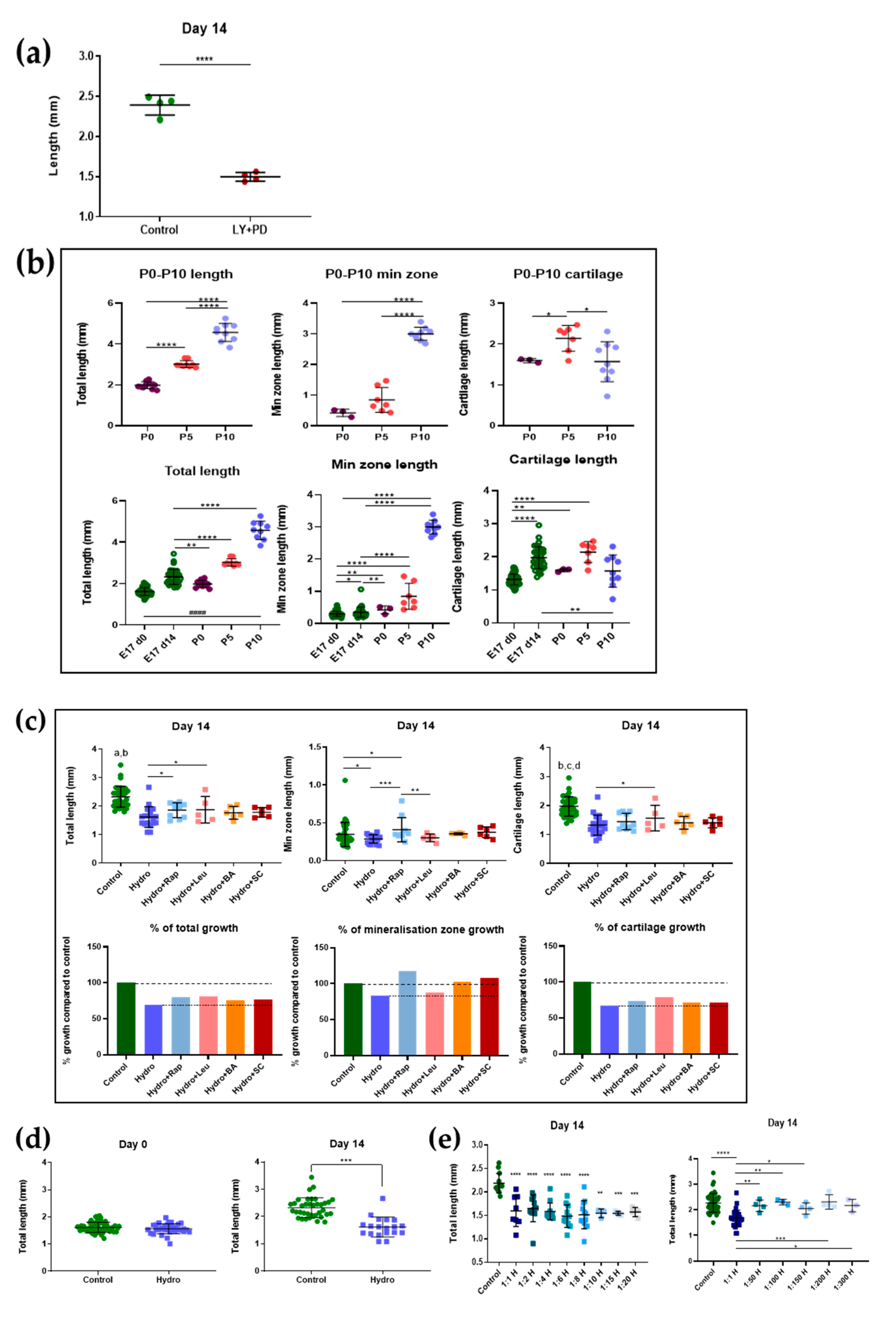
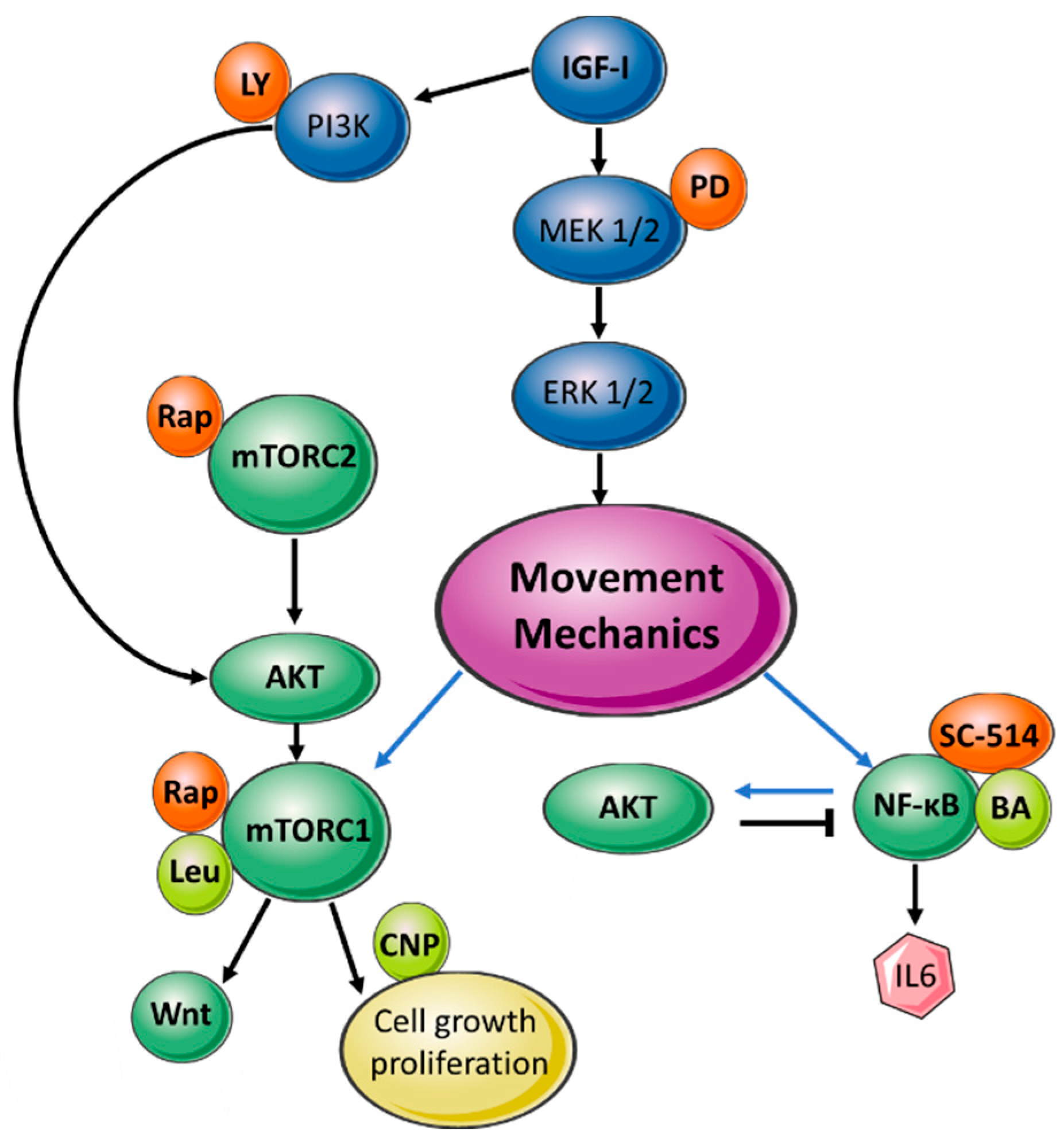
2.1. Mouse Metatarsal Growth Is Relatively Insensitive to mTOR and NF-ĸB Pathway Regulators
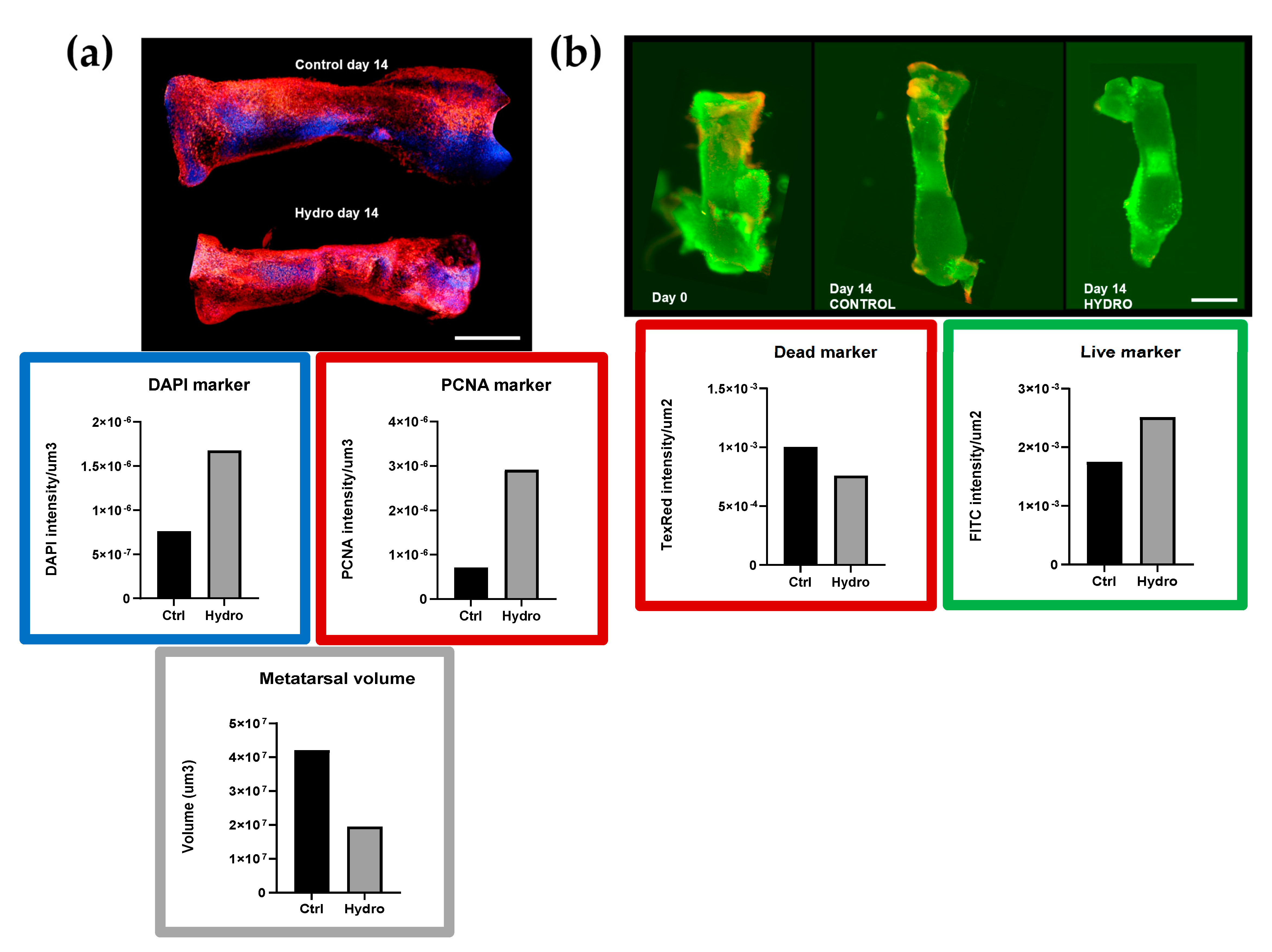
2.2. Interaction between Quasi-Static Loading and mTOR Modulates Endochondral Growth
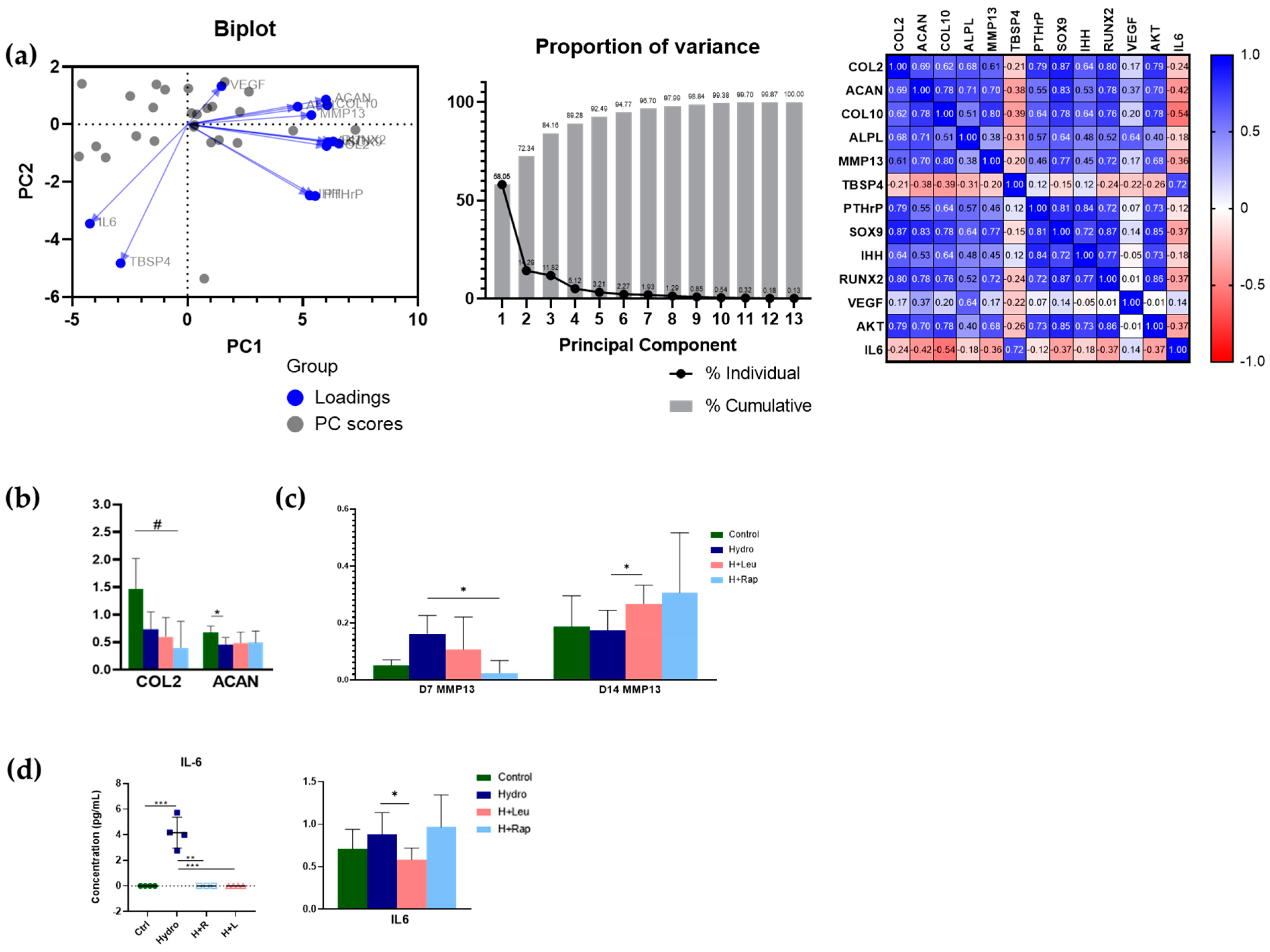
2.3. Transcriptional Analyses Support the Interplay between Quasi-Static Mechanical Load and mTOR Pathway Signalling in the Control of Endochondral Growth
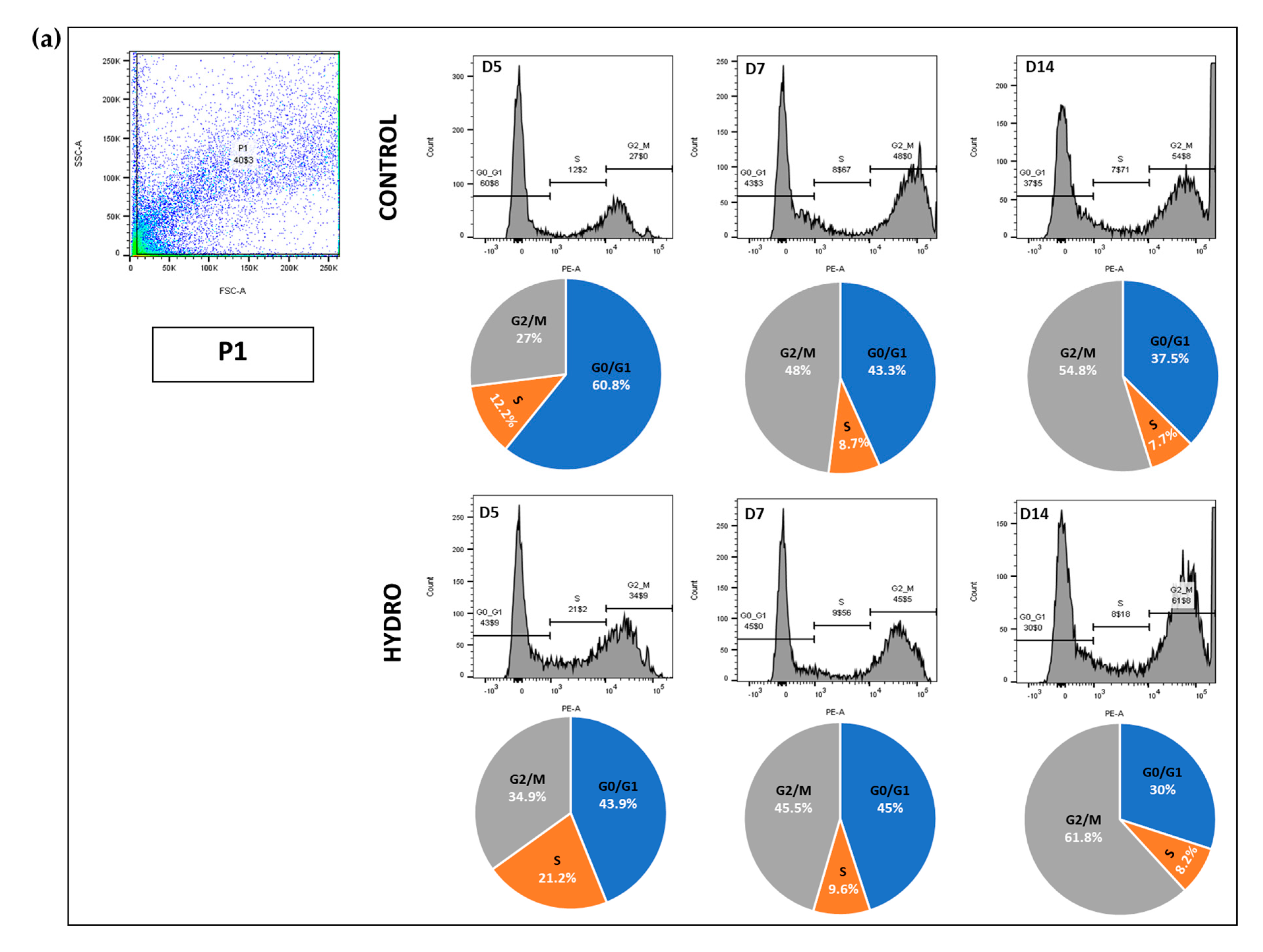
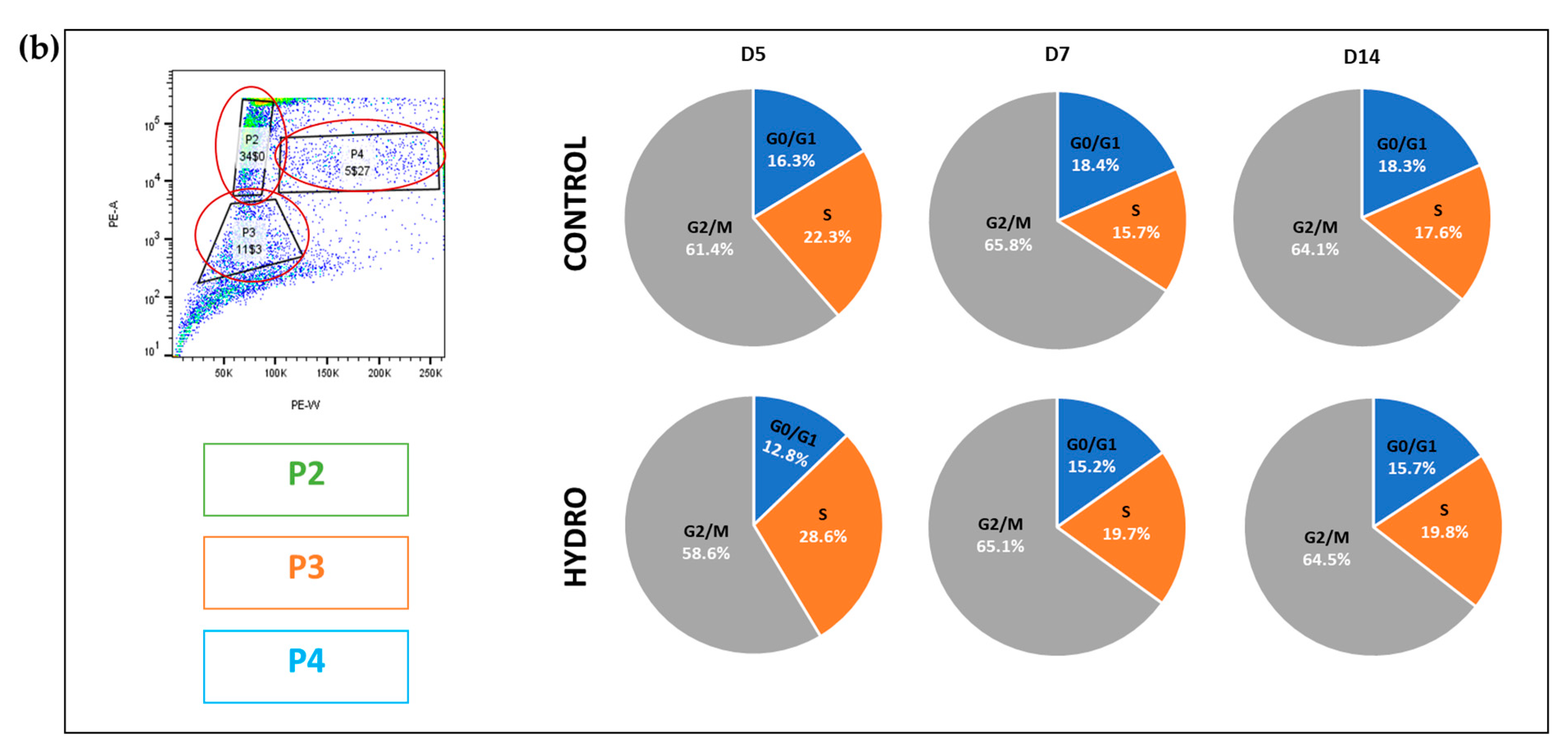
2.4. Quasi-Static Loading Induces Chondrocyte S to G2/M Cell Cycle Arrest
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Mouse Metatarsal Organ Culture Model
4.3. Pharmacological Manipulation of Murine Metatarsi
4.4. Mechanical Manipulation of Murine Metatarsi
4.5. Quantification of Change in Element Length
4.6. Sample Size and Individual Experiments in Metatarsi Length
4.7. Live/Dead Assay
4.8. Confocal Imaging
4.9. RNA Isolation
4.10. Multiplex Real Time Polymerase Chain Reaction
4.11. Assessment of mTOR and NF-ĸB Activity in Culture Supernatant
4.12. Flow Cytometry
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, W.; Chung, U.-I.; Kronenberg, H.M.; de Crombrugghe, B. The chondrogenic transcription factor Sox9 is a target of signaling by the parathyroid hormone-related peptide in the growth plate of endochondral bones. Proc. Natl. Acad. Sci. USA 2001, 98, 160–165. [Google Scholar] [CrossRef]
- Shu, B.; Zhang, M.; Xie, R.; Wang, M.; Jin, H.; Hou, W.; Tang, D.; Harris, S.E.; Mishina, Y.; O’Keefe, R.J.; et al. BMP2, but not BMP4, is crucial for chondrocyte proliferation and maturation during endochondral bone development. J. Cell Sci. 2011, 124, 3428–3440. [Google Scholar] [CrossRef]
- Hutchison, M.R.; Bassett, M.H.; White, P.C. Insulin-like growth factor-I and fibroblast growth factor, but not growth hormone, affect growth plate chondrocyte proliferation. Endocrinology 2007, 148, 3122–3130. [Google Scholar] [CrossRef][Green Version]
- Provot, S.; Schipani, E. Molecular mechanisms of endochondral bone development. Biochem. Biophys. Res. Commun. 2005, 328, 658–665. [Google Scholar] [CrossRef] [PubMed]
- Mackie, E.J.; Ahmed, Y.A.; Tatarczuch, L.; Chen, K.S.; Mirams, M. Endochondral ossification: How cartilage is converted into bone in the developing skeleton. Int. J. Biochem. Cell Biol. 2008, 40, 46–62. [Google Scholar] [CrossRef] [PubMed]
- White, A.; Wallis, G. Endochondral ossification: A delicate balance between growth and mineralisation. Curr. Biology. 2001, 11, R589–R591. [Google Scholar] [CrossRef]
- Gerber, H.P.; Vu, T.H.; Ryan, A.M.; Kowalski, J.; Werb, Z.; Ferrara, N. VEGF couples hypertrophic cartilage remodeling, ossification and angiogenesis during endochondral bone formation. Nat. Med. 1999, 5, 623–628. [Google Scholar] [CrossRef]
- Haeusler, G.; Walter, I.; Helmreich, M.; Egerbacher, M. Localization of matrix metalloproteinases, (MMPs) their tissue inhibitors, and vascular endothelial growth factor (VEGF) in growth plates of children and adolescents indicates a role for MMPs in human postnatal growth and skeletal maturation. Calcif. Tissue Int. 2005, 76, 326–335. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, E.A.; Bouguerra, S.; Londono, I.; Moldovan, F.; Aubin, C.E.; Villemure, I. In situ deformation of growth plate chondrocytes in stress-controlled static vs. dynamic compression. J. Biomech. 2017, 56, 76–82. [Google Scholar] [CrossRef]
- Carroll, S.F.; Buckley, C.T.; Kelly, D.J. Cyclic Tensile Strain Can Play a Role in Directing both Intramembranous and Endochondral Ossification of Mesenchymal Stem Cells. Front Bioeng. Biotechnol. 2017, 5, 73. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, S.; Yoshikawa, H.; Nakata, K. Axial mechanical loading to ex vivo mouse long bone regulates endochondral ossification and endosteal mineralization through activation of the BMP-Smad pathway during postnatal growth. Bone Rep. 2021, 15, 101088. [Google Scholar] [CrossRef]
- Nowlan, N.C.; Sharpe, J.; Roddy, K.A.; Prendergast, P.J.; Murphy, P. Mechanobiology of embryonic skeletal development: Insights from animal models. Birth Defects Res. Part C Embryo Today Rev. 2010, 90, 203–213. [Google Scholar] [CrossRef]
- Roddy, K.A.; Prendergast, P.J.; Murphy, P. Mechanical influences on morphogenesis of the knee joint revealed through morphological, molecular and computational analysis of immobilised embryos. PLoS ONE 2011, 6, e17526. [Google Scholar] [CrossRef]
- Shea, C.A.; Rolfe, R.A.; Murphy, P. The importance of foetal movement for co-ordinated cartilage and bone development in utero. Bone Jt. Res. 2015, 4, 105–116. [Google Scholar] [CrossRef]
- Levillain, A.; Rolfe, R.A.; Huang, Y.; Iatridis, J.C.; Nowlan, N.C. Short-term foetal immobility temporally and progressively affects chick spinal curvature and anatomy and rib development. Eur. Cells Mater. 2019, 37, 23–41. [Google Scholar] [CrossRef] [PubMed]
- Rolfe, R.A.; O’Callaghan, D.S.; Murphy, P. Joint development recovery on resumption of embryonic movement following paralysis. Dis. Models Mech. 2021, 14, dmm048913. [Google Scholar] [CrossRef]
- Houston, D.A.; Staines, K.A.; MacRae, V.E.; Farquharson, C. Culture of Murine Embryonic Metatarsals: A Physiological Model of Endochondral Ossification. J. Vis. Exp. 2016, 118, e54978. [Google Scholar] [CrossRef] [PubMed]
- Marino, S.; Staines, K.A.; Brown, G.; Howard-Jones, R.A.; Adamczyk, M. Models of ex vivo explant cultures: Applications in bone research. Bonekey Rep. 2016, 5, 818. [Google Scholar] [CrossRef] [PubMed]
- Dobie, R.; Ahmed, S.F.; Staines, K.A.; Pass, C.; Jasim, S.; MacRae, V.E.; Farquharson, C. Increased linear bone growth by GH in the absence of SOCS2 is independent of IGF-1. J. Cell. Physiol. 2015, 230, 2796–2806. [Google Scholar] [CrossRef]
- Macrae, V.E.; Ahmed, S.F.; Mushtaq, T.; Farquharson, C. IGF-I signalling in bone growth: Inhibitory actions of dexamethasone and IL-1beta. Growth Horm. IGF Res. 2007, 17, 435–439. [Google Scholar] [CrossRef] [PubMed]
- Rokutanda, S.; Fujita, T.; Kanatani, N.; Yoshida, C.A.; Komori, H.; Liu, W.; Mizuno, A.; Komori, T. Akt regulates skeletal development through GSK3, mTOR, and FoxOs. Dev. Biol. 2009, 328, 78–93. [Google Scholar] [CrossRef] [PubMed]
- Hahn-Windgassen, A.; Nogueira, V.; Chen, C.C.; Skeen, J.E.; Sonenberg, N.; Hay, N. Akt activates the mammalian target of rapamycin by regulating cellular ATP level and AMPK activity. J. Biol. Chem. 2005, 280, 32081–32089. [Google Scholar] [CrossRef] [PubMed]
- Dan, H.C.; Ebbs, A.; Pasparakis, M.; Van Dyke, T.; Basseres, D.S.; Baldwin, A.S. Akt-dependent activation of mTORC1 complex involves phosphorylation of mTOR (mammalian target of rapamycin) by IkappaB kinase alpha (IKKalpha). J. Biol. Chem. 2014, 289, 25227–25240. [Google Scholar] [CrossRef]
- Sun, K.; Luo, J.; Guo, J.; Yao, X.; Jing, X.; Guo, F. The PI3K/AKT/mTOR signaling pathway in osteoarthritis: A narrative review. Osteoarthr. Cartil. 2020, 28, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Pollard, A.S.; Charlton, B.G.; Hutchinson, J.R.; Gustafsson, T.; McGonnell, I.M.; Timmons, J.A.; Pitsillides, A.A. Limb proportions show developmental plasticity in response to embryo movement. Sci. Rep. 2017, 7, 41926. [Google Scholar] [CrossRef]
- Rawlinson, S.C.; Murray, D.H.; Mosley, J.R.; Wright, C.D.; Bredl, J.C.; Saxon, L.K.; Loveridge, N.; Leterrier, C.; Constantin, P.; Farquharson, C.; et al. Genetic selection for fast growth generates bone architecture characterised by enhanced periosteal expansion and limited consolidation of the cortices but a diminution in the early responses to mechanical loading. Bone 2009, 45, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Yang, X.; Yang, W.; Charbonneau, C.; Chen, Q. Mechanical activation of mammalian target of rapamycin pathway is required for cartilage development. FASEB J. 2014, 28, 4470–4481. [Google Scholar] [CrossRef]
- Fitter, S.; Matthews, M.P.; Martin, S.K.; Xie, J.; Ooi, S.S.; Walkley, C.R.; Codrington, J.D.; Ruegg, M.A.; Hall, M.N.; Proud, C.G.; et al. mTORC1 Plays an Important Role in Skeletal Development by Controlling Preosteoblast Differentiation. Mol. Cell. Biol. 2017, 37, e00668-16. [Google Scholar] [CrossRef]
- Chen, J.; Long, F. mTOR signaling in skeletal development and disease. Bone Res. 2018, 6, 1. [Google Scholar] [CrossRef]
- Agirdil, Y. The growth plate: A physiologic overview. EFORT Open Rev. 2020, 5, 498–507. [Google Scholar] [CrossRef]
- Novack, D.V. Role of NF-kappaB in the skeleton. Cell Res. 2011, 21, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Jimi, E.; Fei, H.; Nakatomi, C. NF-kappaB Signaling Regulates Physiological and Pathological Chondrogenesis. Int. J. Mol. Sci. 2019, 20, 6275. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, J.; Sohn, P.; Zeng, X.; Doetschman, T.; Robbins, D.J.; Serra, R. TGFb2 mediates the effects of Hedgehog on hypertrophic differentiation and PTHrP expression. Development 2002, 129, 1913–1924. [Google Scholar] [CrossRef]
- Amano, K.; Ichida, F.; Sugita, A.; Hata, K.; Wada, M.; Takigawa, Y.; Nakanishi, M.; Kogo, M.; Nishimura, R.; Yoneda, T. MSX2 stimulates chondrocyte maturation by controlling Ihh expression. J. Biol. Chem. 2008, 283, 29513–29521. [Google Scholar] [CrossRef]
- Huesa, C.; Houston, D.; Kiffer-Moreira, T.; Yadav, M.M.; Millan, J.L.; Farquharson, C. The Functional co-operativity of Tissue-Nonspecific Alkaline Phosphatase (TNAP) and PHOSPHO1 during initiation of Skeletal Mineralization. Biochem. Biophys. Rep. 2015, 4, 196–201. [Google Scholar] [CrossRef]
- Cai, H.; Liu, A. Spop promotes skeletal development and homeostasis by positively regulating Ihh signaling. Proc. Natl. Acad. Sci. USA 2016, 113, 14751–14756. [Google Scholar] [CrossRef] [PubMed]
- Kurki, P.; Vanderlaan, M.; Dolbeare, F.; Gray, J.; Tan, E.M. Expression of proliferating cell nuclear antigen (PCNA)/cyclin during the cell cycle. Exp. Cell Res. 1986, 166, 209–219. [Google Scholar] [CrossRef]
- Ohashi, N.; Robling, A.G.; Burr, D.B.; Turner, C.H. The effects of dynamic axial loading on the rat growth plate. J. Bone Miner. Res. 2002, 17, 284–292. [Google Scholar] [CrossRef]
- Benoit, A.; Mustafy, T.; Londono, I.; Grimard, G.; Aubin, E.; Villemure, I. In vivo dynamic compression has less detrimental effect than static compression on newly formed bone of a rat caudal vertebra. J. Musculoskelet Neuronal Interact. 2016, 16, 211–220. [Google Scholar]
- Ferrao Blanco, M.N.; Bastiaansen-Jenniskens, Y.M.; Chambers, M.G.; Pitsillides, A.A.; Narcisi, R.; van Osch, G. Effect of Inflammatory Signaling on Human Articular Chondrocyte Hypertrophy: Potential Involvement of Tissue Repair Macrophages. Cartilage 2021, 19476035211021907. [Google Scholar] [CrossRef]
- Marcu, K.B.; Otero, M.; Olivotto, E.; Borzi, R.M.; Goldring, M.B. NF-κB Signaling: Multiple angles to target OA. Curr. Drug Targets 2010, 11, 599–613. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Wang, Z.; Tang, E.; Fan, Z.; McCauley, L.; Franceschi, R.; Guan, K.; Krebsbach, P.H.; Wang, C.Y. Inhibition of osteoblastic bone formation by nuclear factor-kappaB. Nat. Med. 2009, 15, 682–689. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Wu, H.; Chim, S.M.; Zhou, L.; Zhao, J.; Feng, H.; Wei, Q.; Wang, Q.; Zheng, M.H.; Tan, R.X.; et al. SC-514, a selective inhibitor of IKKbeta attenuates RANKL-induced osteoclastogenesis and NF-kappaB activation. Biochem. Pharmacol. 2013, 86, 1775–1783. [Google Scholar] [CrossRef] [PubMed]
- Son, S.M.; Park, S.J.; Lee, H.; Siddiqi, F.; Lee, J.E.; Menzies, F.M.; Rubinsztein, D.C. Leucine Signals to mTORC1 via Its Metabolite Acetyl-Coenzyme A. Cell Metab. 2019, 29, 192–201. [Google Scholar] [CrossRef]
- Condon, K.J.; Sabatini, D.M. Nutrient regulation of mTORC1 at a glance. J. Cell Sci. 2019, 132, jcs222570. [Google Scholar] [CrossRef]
- Kasperczyk, H.; La Ferla-Bruhl, K.; Westhoff, M.A.; Behrend, L.; Zwacka, R.M.; Debatin, K.M.; Fulda, S. Betulinic acid as new activator of NF-kappaB: Molecular mechanisms and implications for cancer therapy. Oncogene 2005, 24, 6945–6956. [Google Scholar] [CrossRef]
- Kroemer, S.F.A.G. Targeting mitochondrial apoptosis by betulinic acid in human cancers. Drug Discov. Today 2009, 14, 885–890. [Google Scholar] [CrossRef]
- Liu, V.W.S.; Yau, W.L.; Tam, C.W.; Yao, K.M.; Shiu, S.Y.W. Melatonin Inhibits Androgen Receptor Splice Variant-7 (AR-V7)-Induced Nuclear Factor-Kappa B (NF-kB) Activation and NF-kB Activator-Induced AR-V7 Expression in Prostate Cancer Cells: Potential Implications for the Use of Melatonin in Castration-Resistant Prostate Cancer (CRPC) Therapy. Int. J. Mol. Sci. 2017, 18, 1130. [Google Scholar] [CrossRef]
- Choi, H.; Jeong, B.C.; Kook, M.S.; Koh, J.T. Betulinic acid synergically enhances BMP2-induced bone formation via stimulating Smad 1/5/8 and p38 pathways. J. Biomed. Sci. 2016, 23, 45. [Google Scholar] [CrossRef]
- Jeong, D.H.; Kwak, S.C.; Lee, M.S.; Yoon, K.H.; Kim, J.Y.; Lee, C.H. Betulinic Acid Inhibits RANKL-Induced Osteoclastogenesis via Attenuating Akt, NF-kappaB, and PLCgamma2-Ca(2+) Signaling and Prevents Inflammatory Bone Loss. J. Nat. Prod. 2020, 83, 1174–1182. [Google Scholar] [CrossRef]
- Bachmann, B.; Spitz, S.; Schadl, B.; Teuschl, A.H.; Redl, H.; Nurnberger, S.; Ertl, P. Stiffness Matters: Fine-Tuned Hydrogel Elasticity Alters Chondrogenic Redifferentiation. Front Bioeng. Biotechnol. 2020, 8, 373. [Google Scholar] [CrossRef]
- Sarrigiannidis, S.O.; Rey, J.M.; Dobre, O.; Gonzalez-Garcia, C.; Dalby, M.J.; Salmeron-Sanchez, M. A tough act to follow: Collagen hydrogel modifications to improve mechanical and growth factor loading capabilities. Mater. Today Bio 2021, 10, 100098. [Google Scholar] [CrossRef]
- Hagel, V.; Haraszti, T.; Boehm, H. Diffusion and interaction in PEG-DA hydrogels. Biointerphases 2013, 8, 36. [Google Scholar] [CrossRef]
- Figueiredo, L.; Pace, R.; D’Arros, C.; Rethore, G.; Guicheux, J.; Le Visage, C.; Weiss, P. Assessing glucose and oxygen diffusion in hydrogels for the rational design of 3D stem cell scaffolds in regenerative medicine. J. Tissue Eng. Regen. Med. 2018, 12, 1238–1246. [Google Scholar] [CrossRef]
- Nam, J.; Aguda, B.D.; Rath, B.; Agarwal, S. Biomechanical thresholds regulate inflammation through the NF-kappaB pathway: Experiments and modeling. PLoS ONE 2009, 4, e5262. [Google Scholar] [CrossRef]
- Segurado, M.; Tercero, J.A. The S-phase checkpoint: Targeting the replication fork. Biol. Cell 2009, 101, 617–627. [Google Scholar] [CrossRef]
- Oeckinghaus, A.; Ghosh, S. The NF-kappaB family of transcription factors and its regulation. Cold Spring Harb. Perspect. Biol. 2009, 1, a000034. [Google Scholar] [CrossRef]
- Bloom, M.J.; Saksena, S.D.; Swain, G.P.; Behar, M.S.; Yankeelov, T.E.; Sorace, A.G. The effects of IKK-beta inhibition on early NF-kappa-B activation and transcription of downstream genes. Cell. Signal. 2019, 55, 17–25. [Google Scholar] [CrossRef]
- Altaf, H.; Revell, P.A. Evidence for active antigen presentation by monocyte/macrophages in response to stimulation with particles: The expression of NFkappaB transcription factors and costimulatory molecules. Inflammopharmacology 2013, 21, 279–290. [Google Scholar] [CrossRef]
- Kishore, N.; Sommers, C.; Mathialagan, S.; Guzova, J.; Yao, M.; Hauser, S.; Huynh, K.; Bonar, S.; Mielke, C.; Albee, L.; et al. A selective IKK-2 inhibitor blocks NF-kappa B-dependent gene expression in interleukin-1 beta-stimulated synovial fibroblasts. J. Biol. Chem. 2003, 278, 32861–32871. [Google Scholar] [CrossRef]
- Jingbo, W.; Aimin, C.; Qi, W.; Xin, L.; Huaining, L. Betulinic acid inhibits IL-1beta-induced inflammation by activating PPAR-gamma in human osteoarthritis chondrocytes. Int. Immunopharmacol. 2015, 29, 687–692. [Google Scholar] [CrossRef]
- Liu, C.; Chen, Y.; Lu, C.; Chen, H.; Deng, J.; Yan, Y.; Xu, Y.Y.; Liu, H.; Huang, H.; Wei, J.; et al. Betulinic acid suppresses Th17 response and ameliorates psoriasis-like murine skin inflammation. Int. Immunopharmacol. 2019, 73, 343–352. [Google Scholar] [CrossRef]
- Li, Z.; Yang, Z.; Passaniti, A.; Lapidus, R.G.; Liu, X.; Cullen, K.J.; Dan, H.C. A positive feedback loop involving EGFR/Akt/mTORC1 and IKK/NF-kB regulates head and neck squamous cell carcinoma proliferation. Oncotarget 2016, 7, 31892–31906. [Google Scholar] [CrossRef]
- Dan, H.C.; Antonia, R.J.; Baldwin, A.S. PI3K/Akt promotes feedforward mTORC2 activation through IKKalpha. Oncotarget 2016, 7, 21064–21075. [Google Scholar] [CrossRef] [PubMed]
- Marr, N.; Hopkinson, M.; Hibbert, A.P.; Pitsillides, A.A.; Thorpe, C.T. Bimodal Whole-Mount Imaging of Tendon Using Confocal Microscopy and X-ray Micro-Computed Tomography. Biol. Proced. Online 2020, 22, 13. [Google Scholar] [CrossRef]
- June, R.K.; Fyhrie, D.P. Enzymatic digestion of articular cartilage results in viscoelasticity changes that are consistent with polymer dynamics mechanisms. Biomed. Eng. Online 2009, 8, 32. [Google Scholar] [CrossRef]
- Kim, K.H.; Sederstrom, J.M. Assaying Cell Cycle Status Using Flow Cytometry. Curr. Protoc. Mol. Biol. 2015, 111, 28.6.1–28.6.11. [Google Scholar] [CrossRef] [PubMed]
- Cecchini, M.J.; Amiri, M.; Dick, F.A. Analysis of cell cycle position in mammalian cells. J. Vis. Exp. 2012, 59, 3491. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Function | Concentration |
|---|---|---|
| Betulinic acid | NF-ĸB activator | 2.5 µM |
| SC-514 | NF-ĸB inhibitor | 20 µM |
| Leucine | mTOR activator | 10 mM |
| Rapamycin | mTOR inhibitor | 100 nM |
| Vitrogel 3D™ (Hydrogel) | Quasi-static loading | 1:1 dilution |
| Control vs. Hydro (%) | ||||
|---|---|---|---|---|
| Day/Cycle Phase | P1 (All Cells) | P2 (Pre-Hypertrophic) | P3 (Proliferative) | P4 (Hypertrophic) |
| Day 5/G0 and G1 | 60.8 vs. 43.9 | 0 | 48.9 vs. 38.3 | 0 |
| 5/S | 12.2 vs. 21.2 | 15.8 vs. 14.6 | 51.1 vs. 61.7 | 0 vs. 9.5 |
| 5/G2 and M | 27 vs. 34.9 | 84.2 vs. 85.4 | 0 | 100 vs. 90.5 |
| Day 7/G0 and G1 | 43.3 vs. 45 | 0 | 55.3 vs. 45.7 | 0 |
| 5/S | 8.7 vs. 9.6 | 2.5 vs. 4.7 | 44.7 vs. 54.3 | 0 |
| 5/G2 and M | 48 vs. 45.5 | 97.5 vs. 95.3 | 0 | 100 |
| Day 14/G0 and G1 | 37.5 vs. 30 | 0 | 54.9 vs. 47.1 | 0 |
| 5/S | 7.7 vs. 8.2 | 2.3 vs. 2.9 | 45.1 vs. 52.9 | 5.5 vs. 3.6 |
| 5/G2 and M | 54.8 vs. 61.8 | 97.7 vs. 97.1 | 0 | 94.5 vs. 96.4 |
| Gene | Sequence (5′-3′) | |
|---|---|---|
| Vegf | F | AGGTGACACTATAGAATAGCTGTGTGTGTGAGTGGCTT |
| R | GTACGACTCACTATAGGGACTCTTTTCTCTGCCTCCGTG | |
| Sox9 | F | AGGTGACACTATAGAATAAGGAAGCTGGCAGACCAGTA |
| R | GTACGACTCACTATAGGGACGTTCTTCACCGACTTCCTC | |
| Runx2 | F | AGGTGACACTATAGAATAACAGTCCCAACTTCCTGTGC |
| R | GTACGACTCACTATAGGGATAGTTCTCATCATTCCCGGC | |
| Alpl | F | AGGTGACACTATAGAATACACTCAAGGGAGAGGTCCAG |
| R | GTACGACTCACTATAGGGACCCAAGAGAGAAACCTGCTG | |
| Ihh | F | AGGTGACACTATAGAATACCGAACCTTCATCTTGGTGT |
| R | GTACGACTCACTATAGGGACCCCGAGAAACATTGGAGTA | |
| Tbsp4 | F | AGGTGACACTATAGAATATTCAGTCCCCAACTCCAAAC |
| R | GTACGACTCACTATAGGGACGTTTCCCGTGTAACCATCT | |
| Il6 | F | AGGTGACACTATAGAATAAGTTGCCTTCTTGGGACTGA |
| R | GTACGACTCACTATAGGGAAGCCTCCGACTTGTGAAGTG | |
| Col10 | F | AGGTGACACTATAGAATAGCAATTGCAGAAAGTCCACA |
| R | GTACGACTCACTATAGGGACTCGATTGAAAGGCACACAA | |
| Mmp13 | F | AGGTGACACTATAGAATACCAGAACTTCCCAACCATGT |
| R | GTACGACTCACTATAGGGAGTCTTCCCCGTGTTCTCAAA | |
| Pthrp | F | AGGTGACACTATAGAATATTCCTGCTCAGCTACTCCGT |
| R | GTACGACTCACTATAGGGAGGTAGCTCTGATTTCGGCTG | |
| Acan | F | AGGTGACACTATAGAATAAGGACTGAAATCAGCGGAGA |
| R | GTACGACTCACTATAGGGATGTCTCTGTAGGGTACCGGG | |
| Col2 | F | AGGTGACACTATAGAATAACACTGGTAAGTGGGGCAAG |
| R | GTACGACTCACTATAGGGATCGCAATGGATTGTGTTGTT | |
| Akt | F | AGGTGACACTATAGAATAGCAGTGGACCACAGTCATTG |
| R | GTACGACTCACTATAGGGACATCGTCTCTTCTTCCTGCC | |
| Gapdh | F | AGGTGACACTATAGAATAGGGTGTGAACCACGAGAAAT |
| R | GTACGACTCACTATAGGGAACTGTGGTCATGAGCCCTTC | |
| Actb | F | AGGTGACACTATAGAATAGTACCACCATGTACCCAGGC |
| R | GTACGACTCACTATAGGGAGTACTTGCGCTCAGGAGGAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caetano-Silva, S.; Simbi, B.H.; Marr, N.; Hibbert, A.; Allen, S.P.; Pitsillides, A.A. Restraint upon Embryonic Metatarsal Ex Vivo Growth by Hydrogel Reveals Interaction between Quasi-Static Load and the mTOR Pathway. Int. J. Mol. Sci. 2021, 22, 13220. https://doi.org/10.3390/ijms222413220
Caetano-Silva S, Simbi BH, Marr N, Hibbert A, Allen SP, Pitsillides AA. Restraint upon Embryonic Metatarsal Ex Vivo Growth by Hydrogel Reveals Interaction between Quasi-Static Load and the mTOR Pathway. International Journal of Molecular Sciences. 2021; 22(24):13220. https://doi.org/10.3390/ijms222413220
Chicago/Turabian StyleCaetano-Silva, Soraia, Bigboy H. Simbi, Neil Marr, Andrew Hibbert, Steve P. Allen, and Andrew A. Pitsillides. 2021. "Restraint upon Embryonic Metatarsal Ex Vivo Growth by Hydrogel Reveals Interaction between Quasi-Static Load and the mTOR Pathway" International Journal of Molecular Sciences 22, no. 24: 13220. https://doi.org/10.3390/ijms222413220
APA StyleCaetano-Silva, S., Simbi, B. H., Marr, N., Hibbert, A., Allen, S. P., & Pitsillides, A. A. (2021). Restraint upon Embryonic Metatarsal Ex Vivo Growth by Hydrogel Reveals Interaction between Quasi-Static Load and the mTOR Pathway. International Journal of Molecular Sciences, 22(24), 13220. https://doi.org/10.3390/ijms222413220