
Bacillus subtilis ANSB168 Producing d-alanyl-d-alanine Carboxypeptidase Could Alleviate the Immune Injury and Inflammation Induced by Ochratoxin A



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
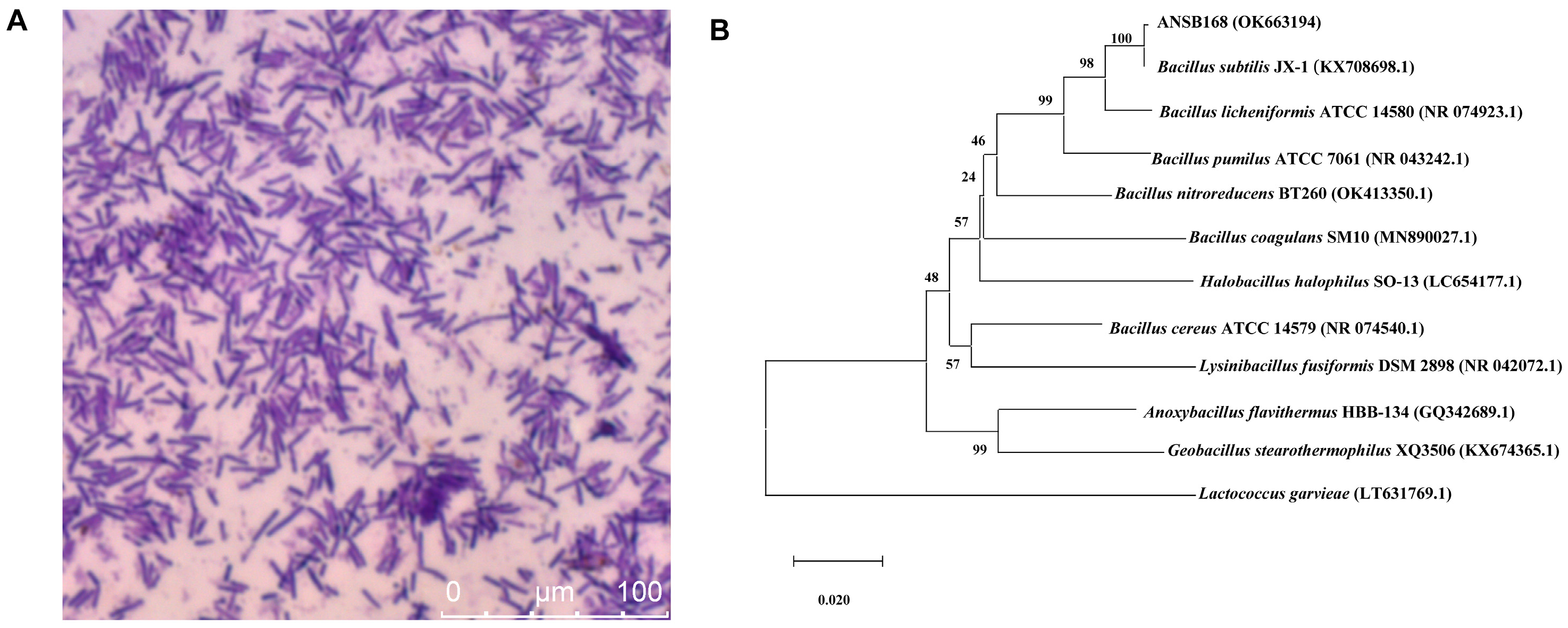
2.1. Isolation and Identity of OTA-Degrading Bacteria ANSB168
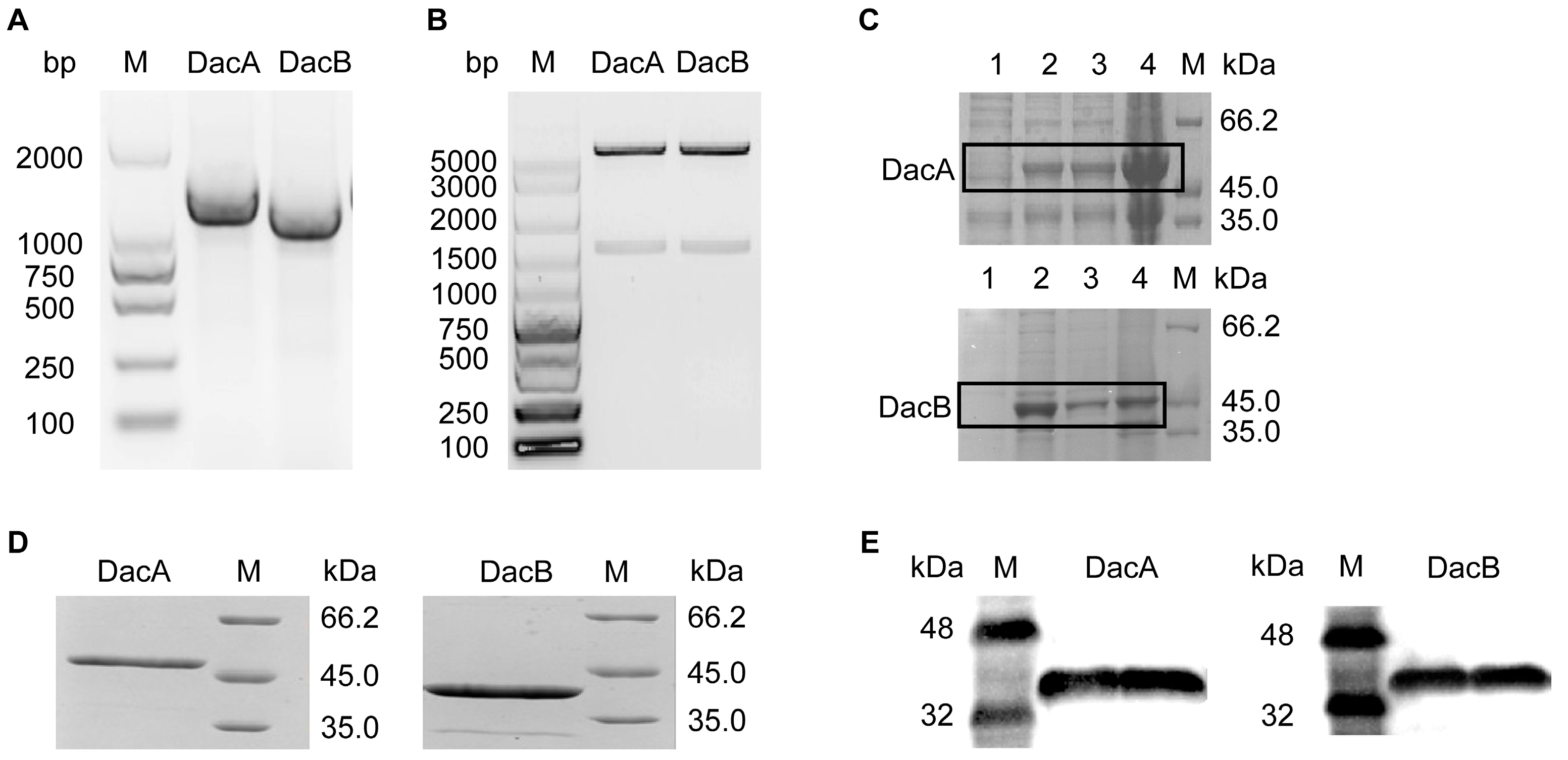
2.2. Cloning and Expression of DacA and DacB in E. coli
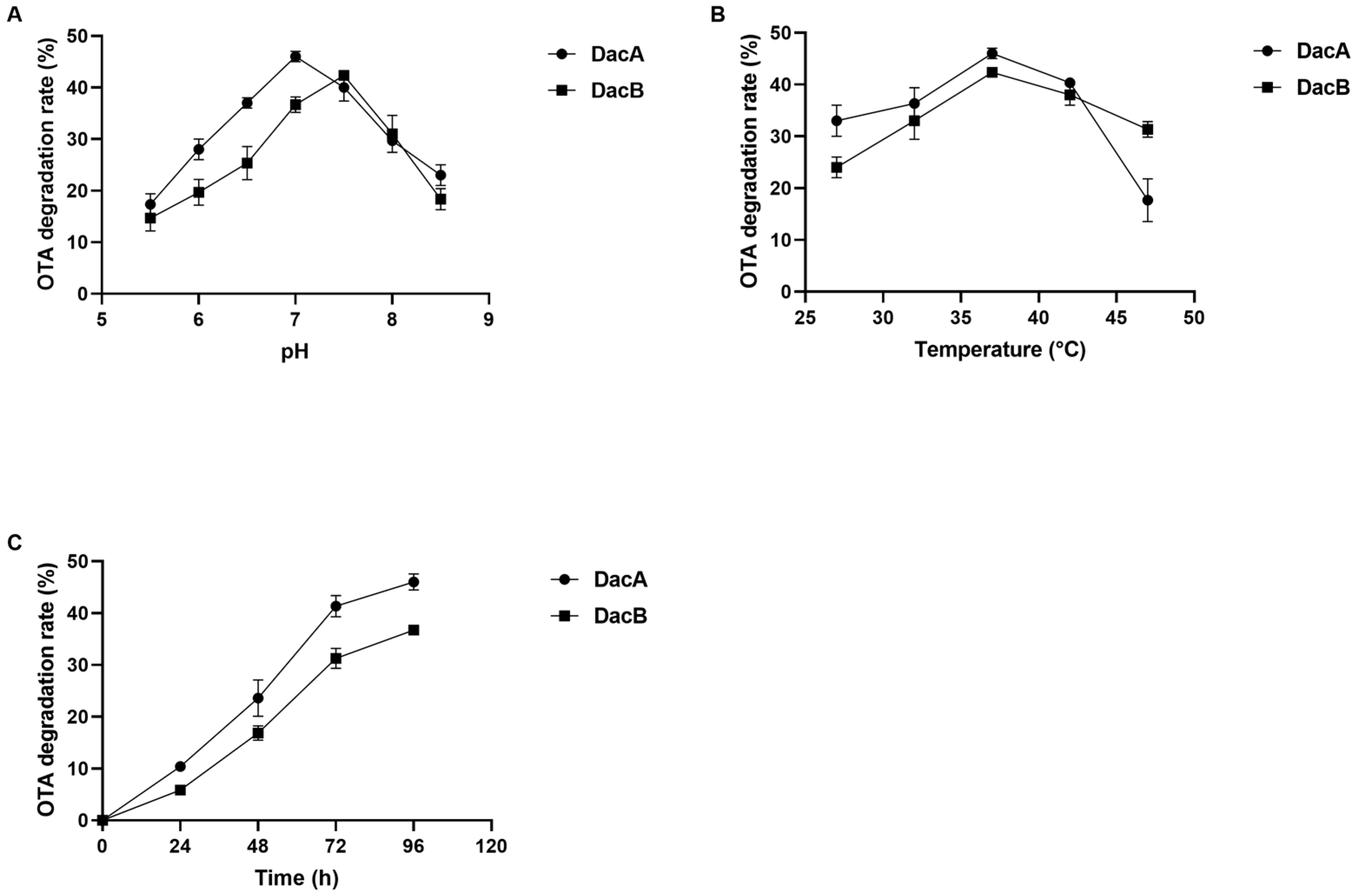
2.3. d-alanyl-d-alanine Carboxypeptidase DacA and DacB Degradation Activity and Biochemical Characteristics
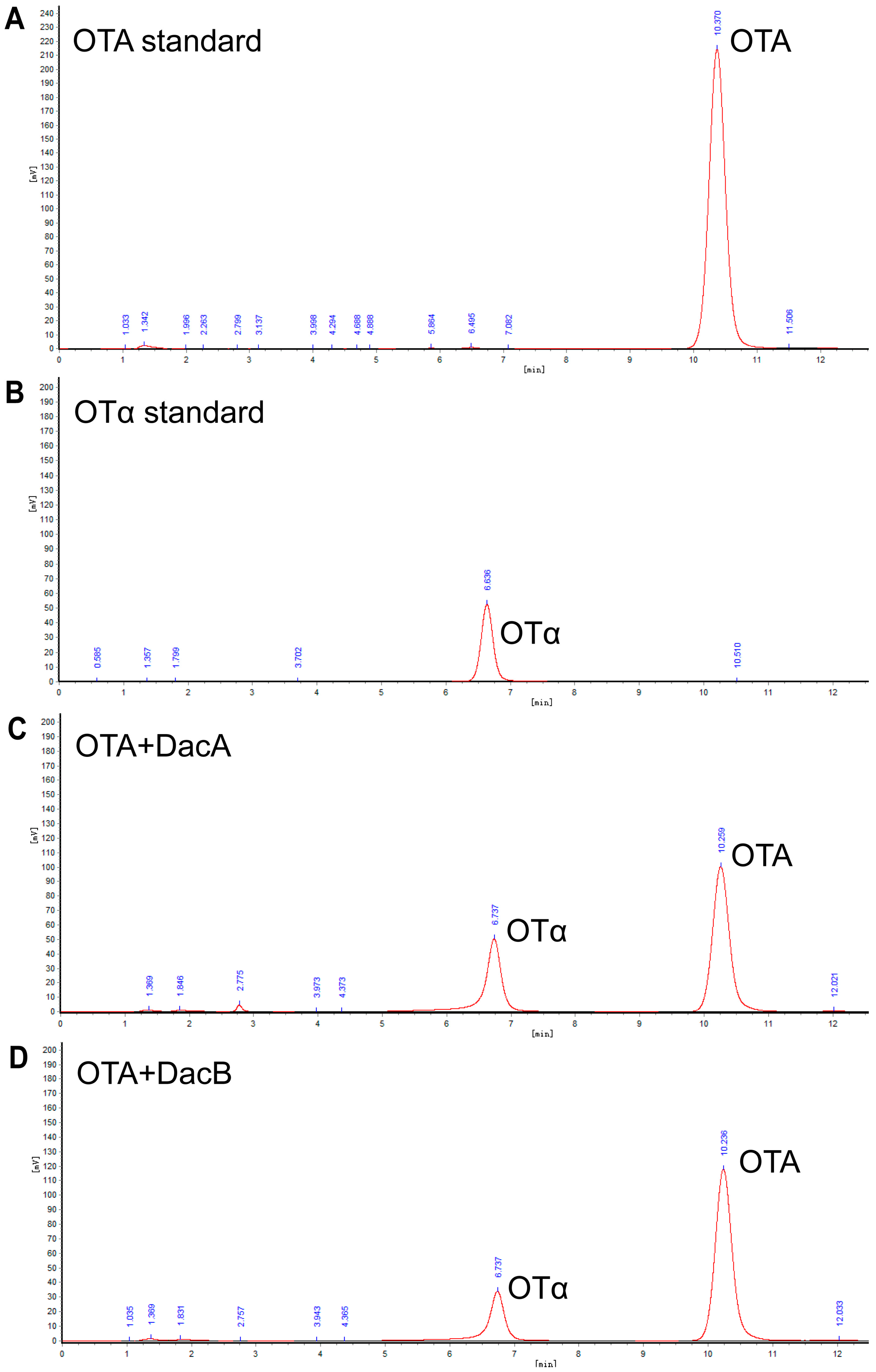
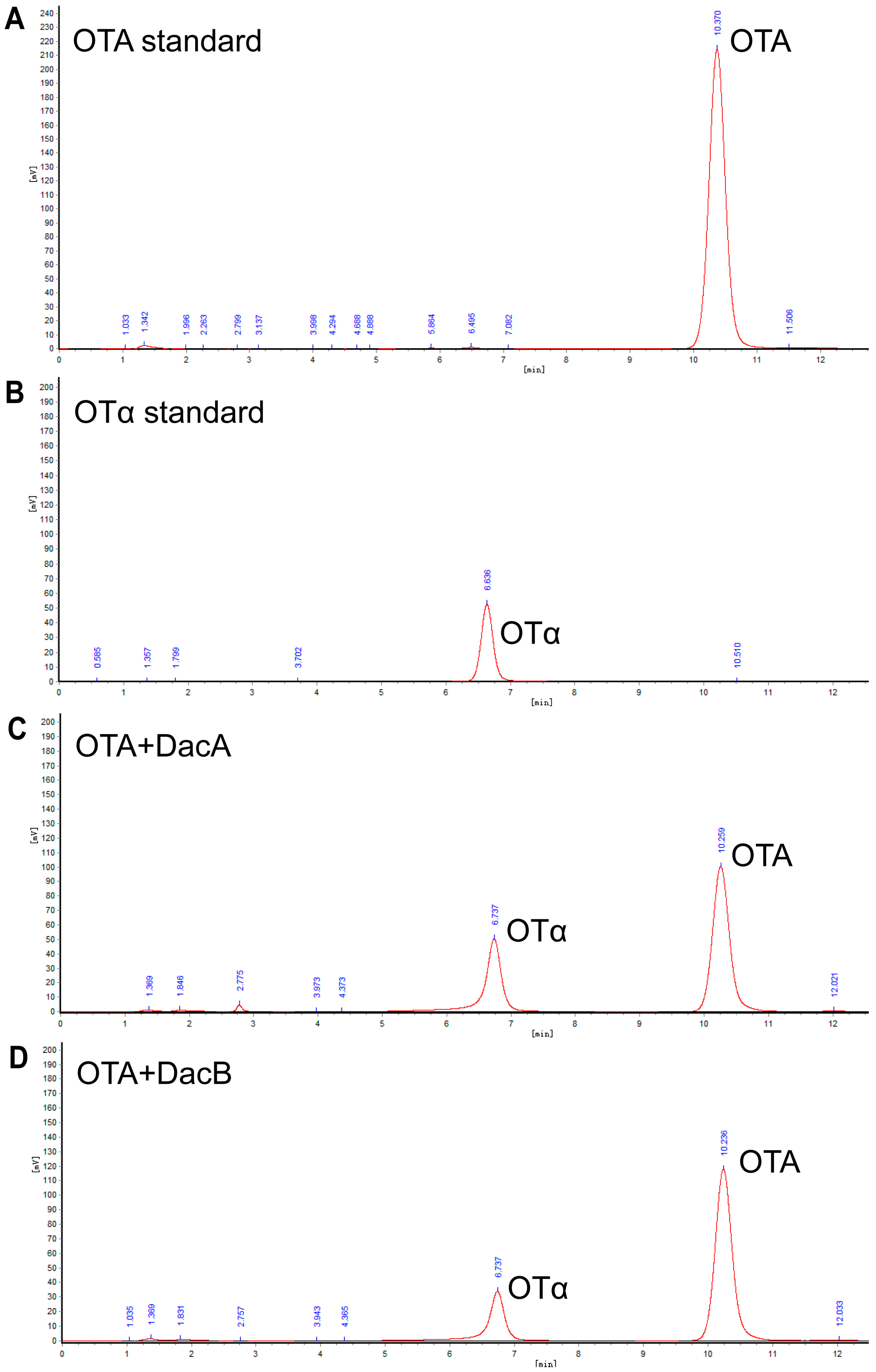
2.4. Degraded Product Identification of DacA and DacB
2.5. Ameliorative Effects of Bacillus subtilis ANSB168 in Laying Hens
2.5.1. Production Performance
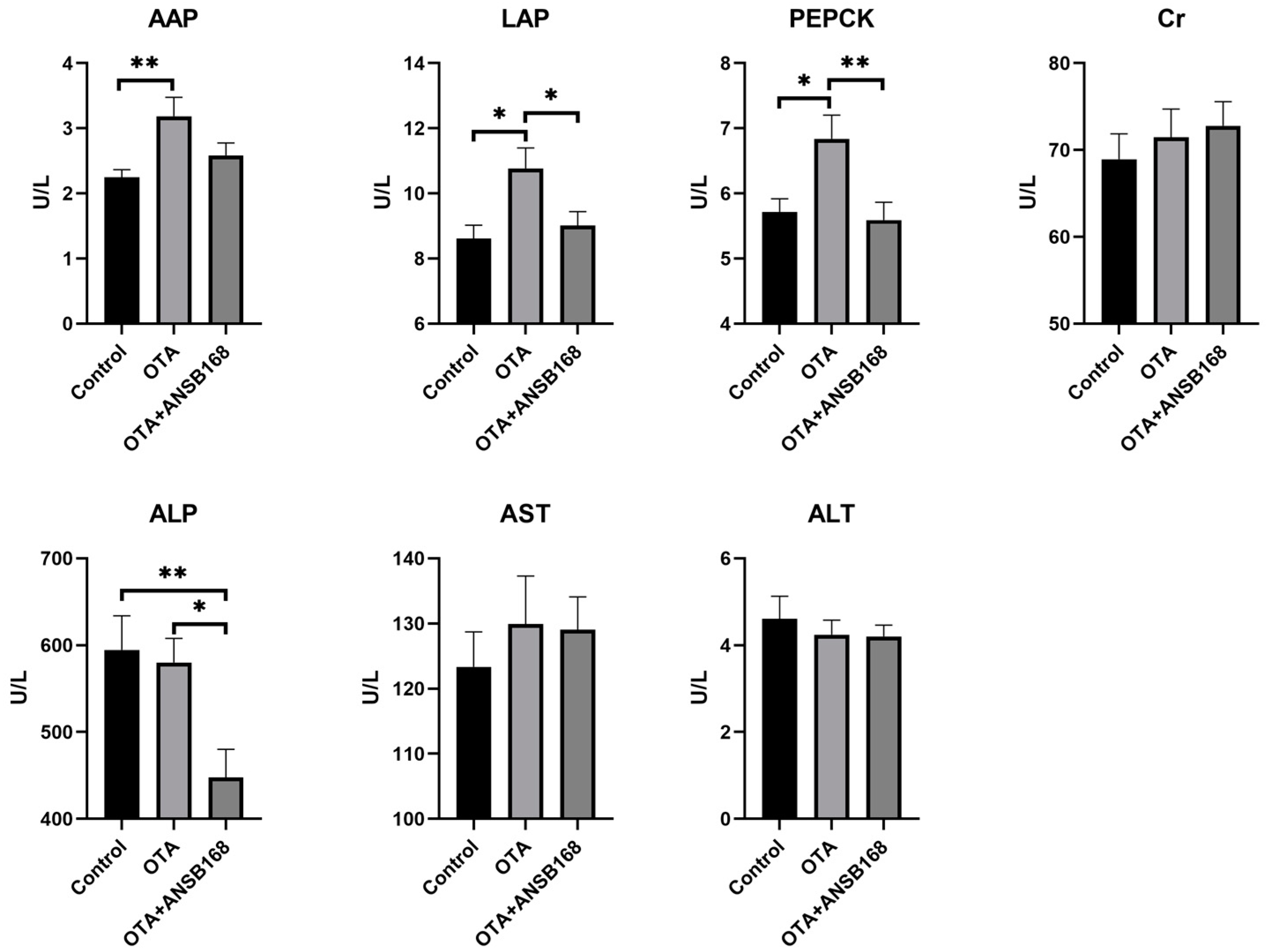
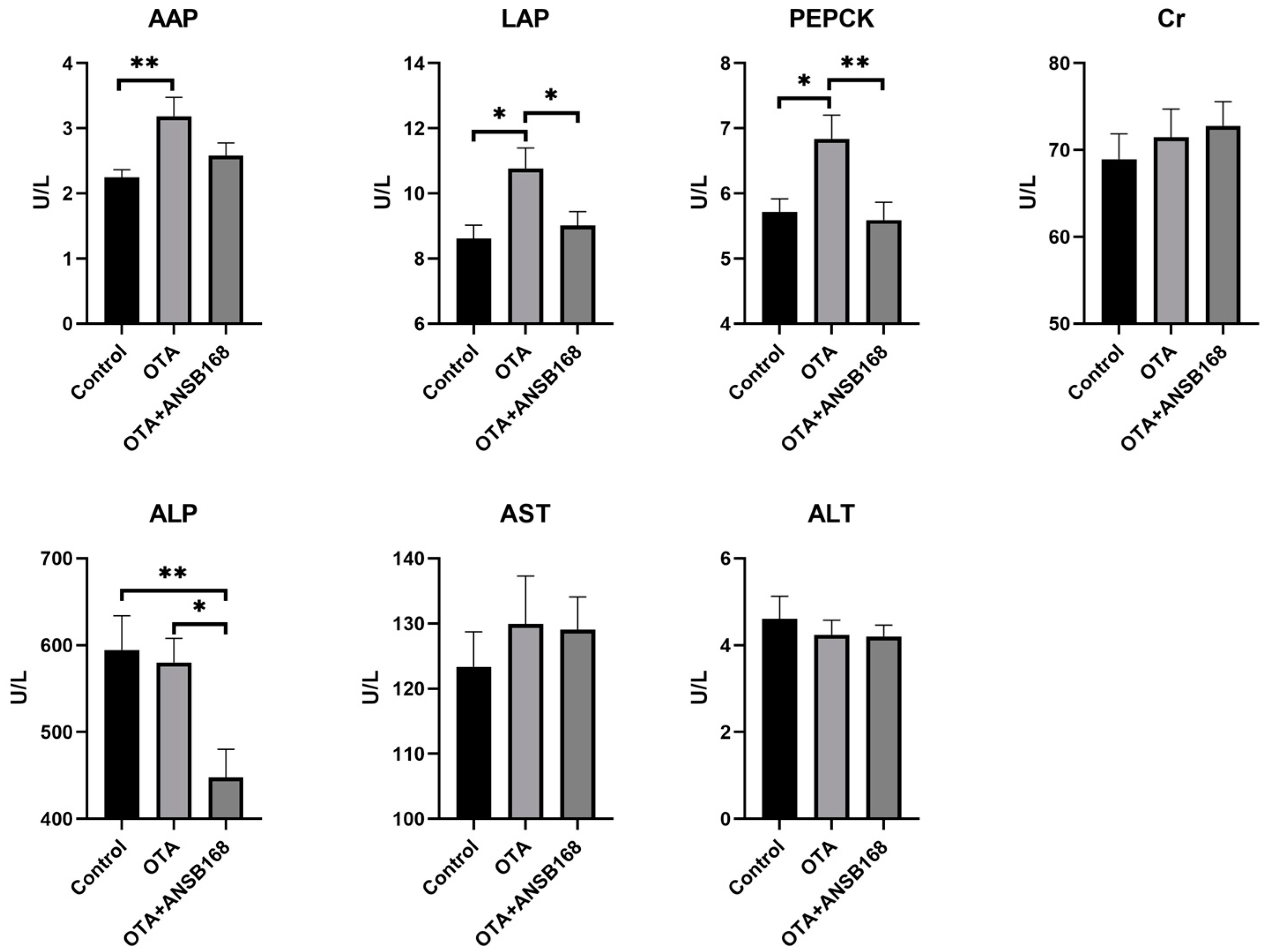
2.5.2. Serum Biochemical Parameters
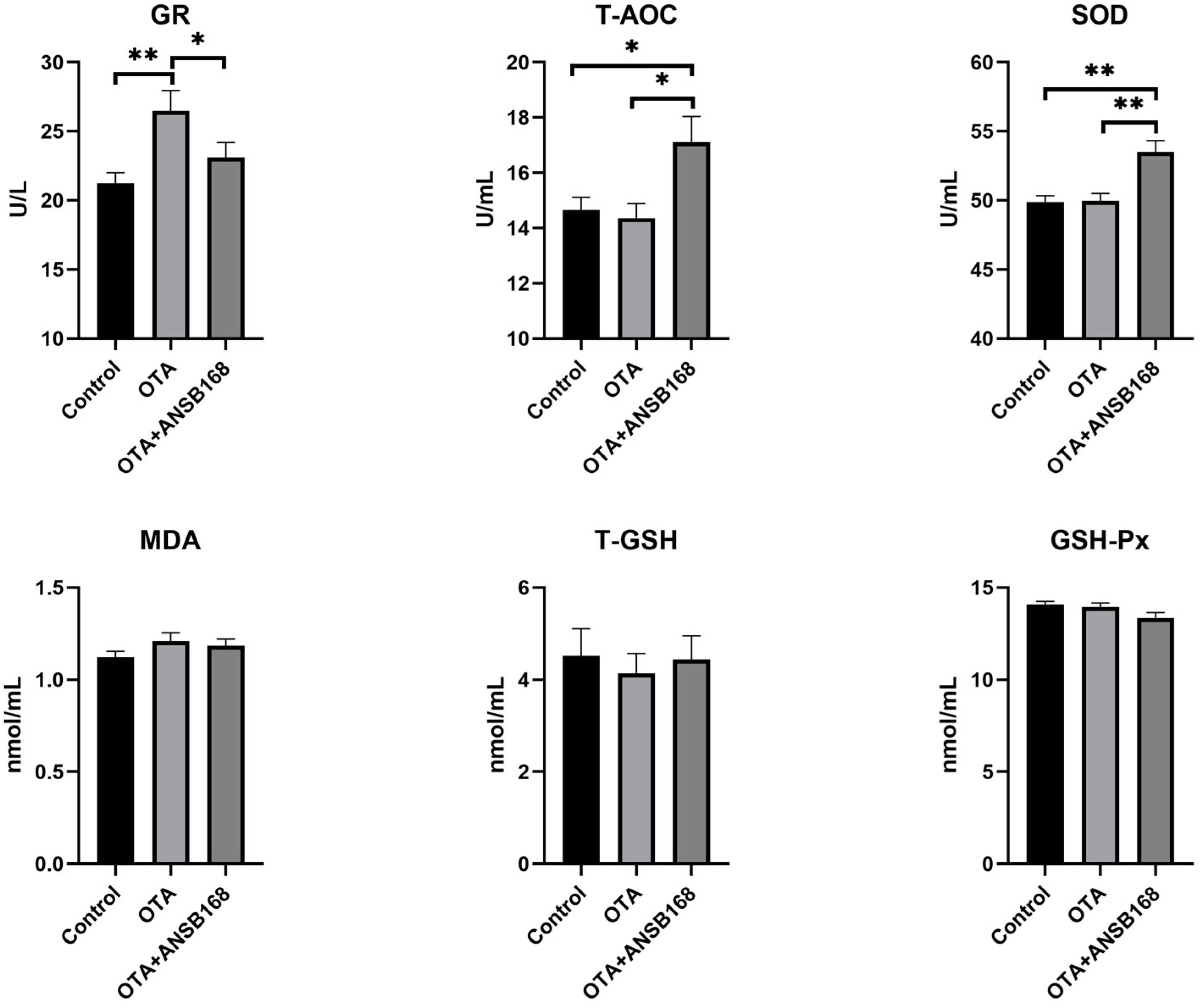
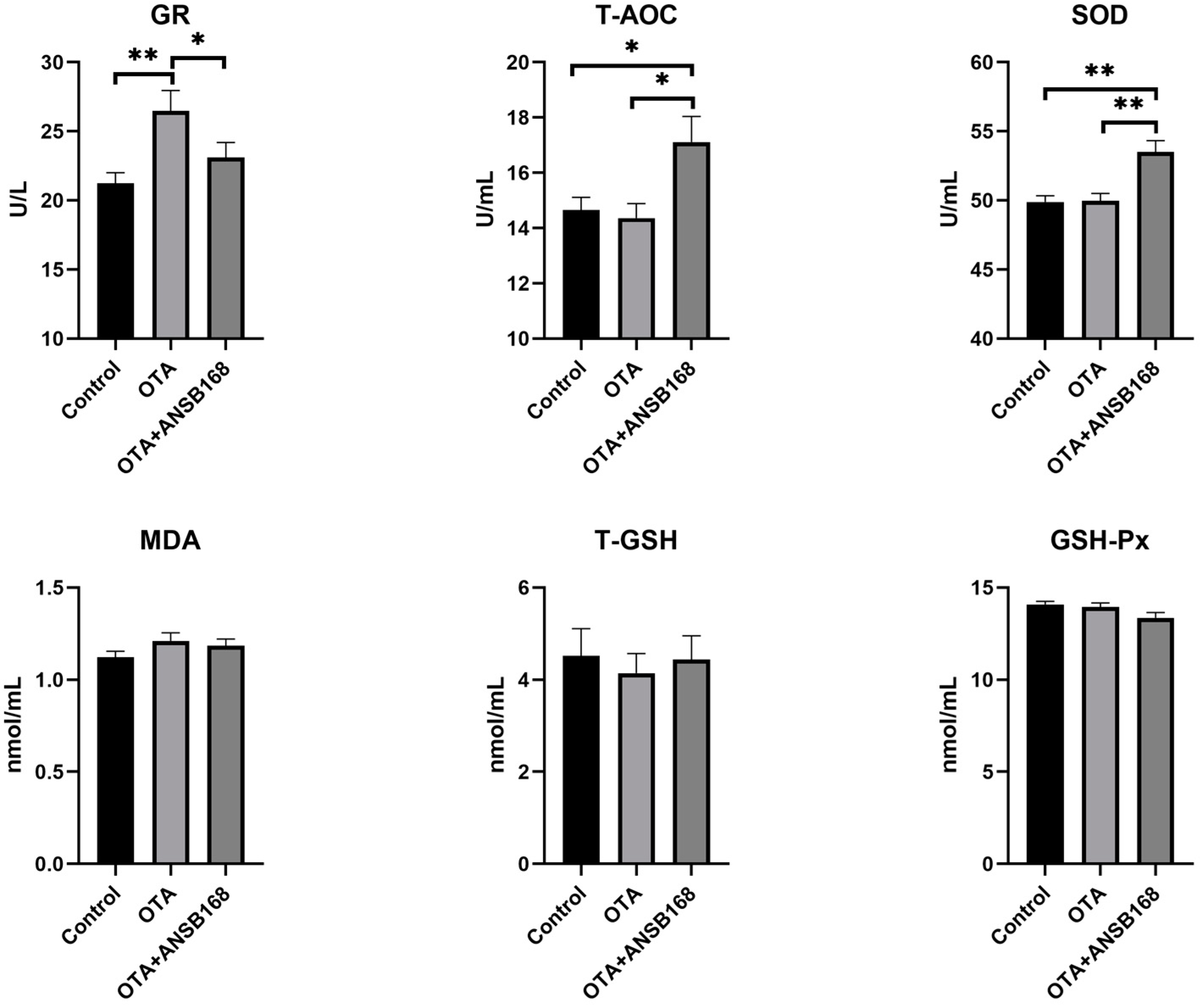
2.5.3. Oxidative Stress and Antioxidant Status
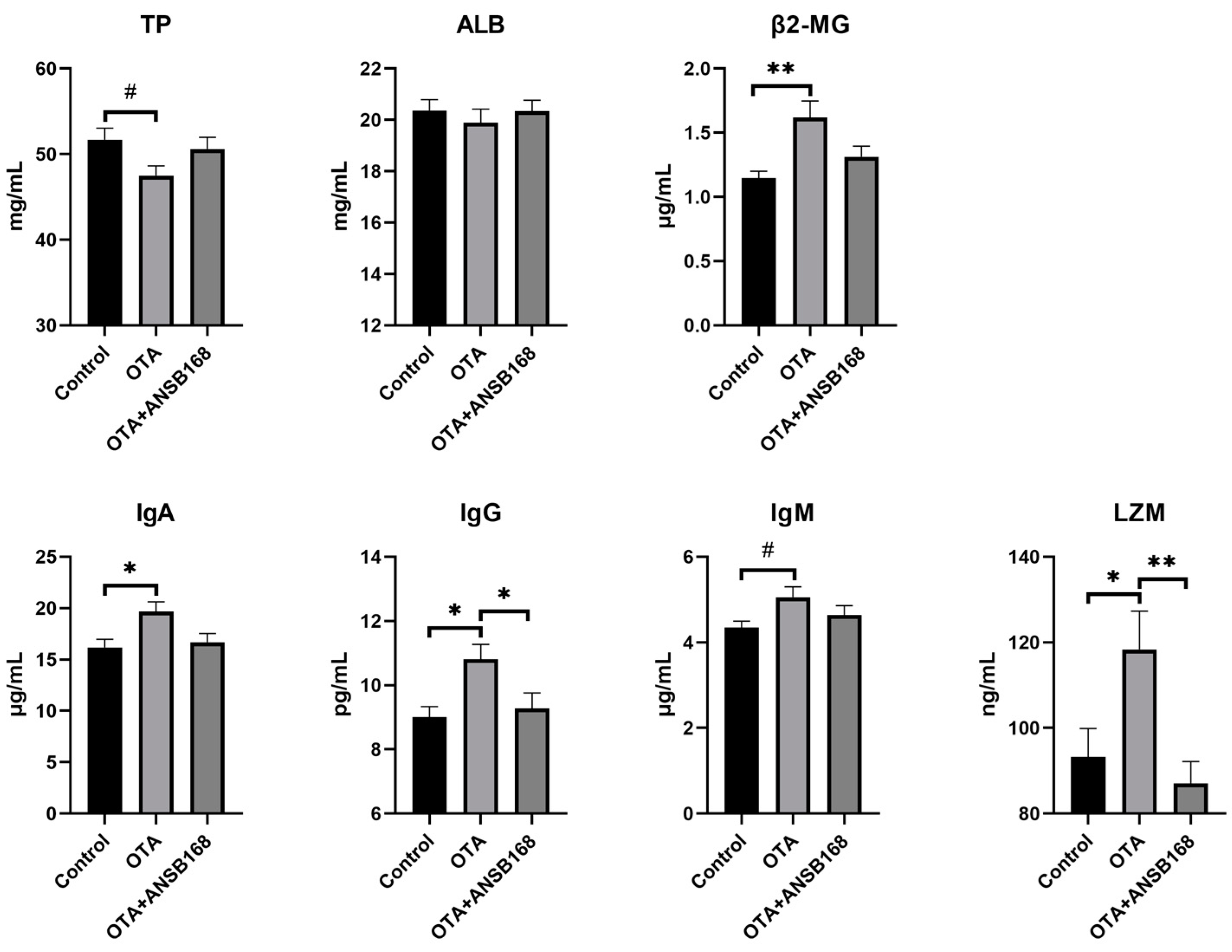
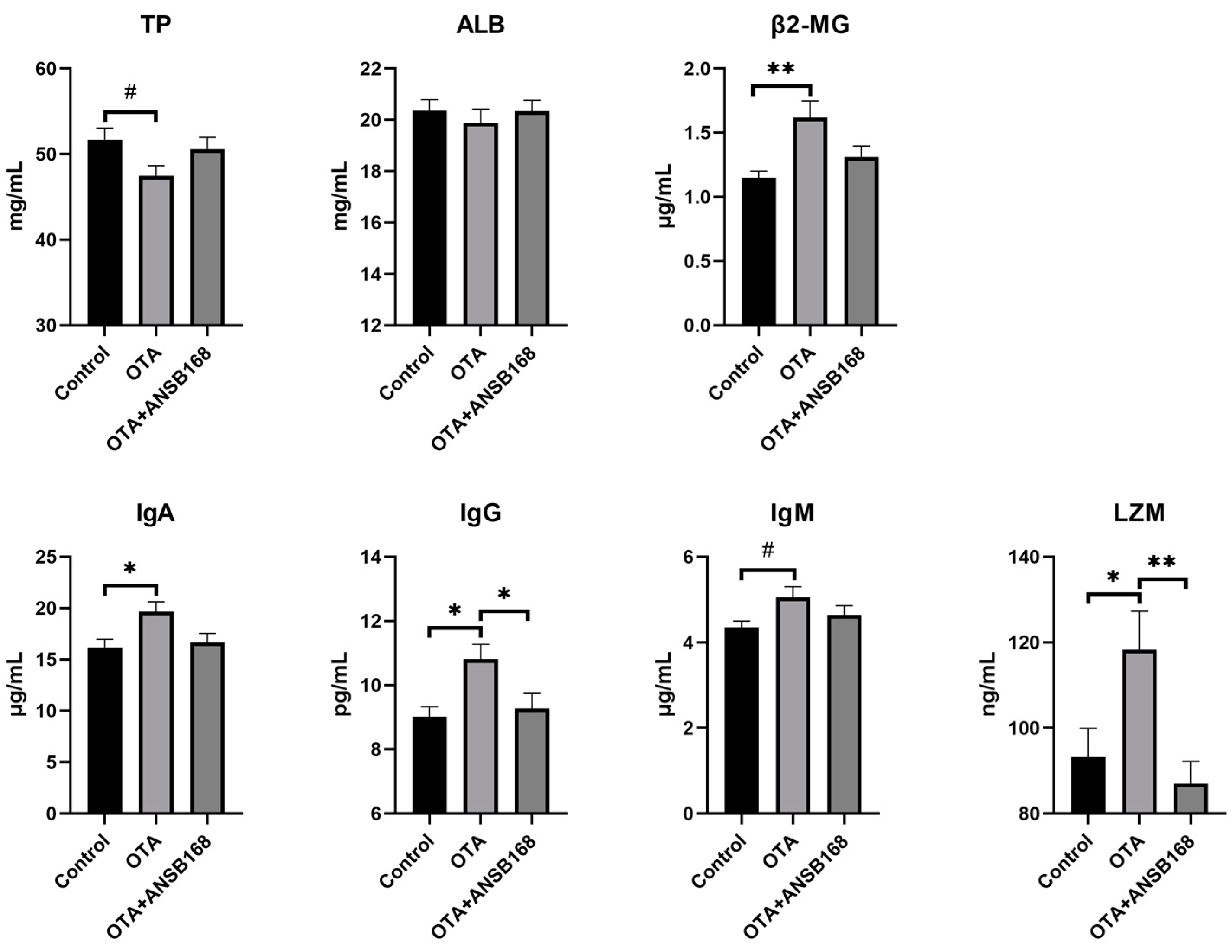
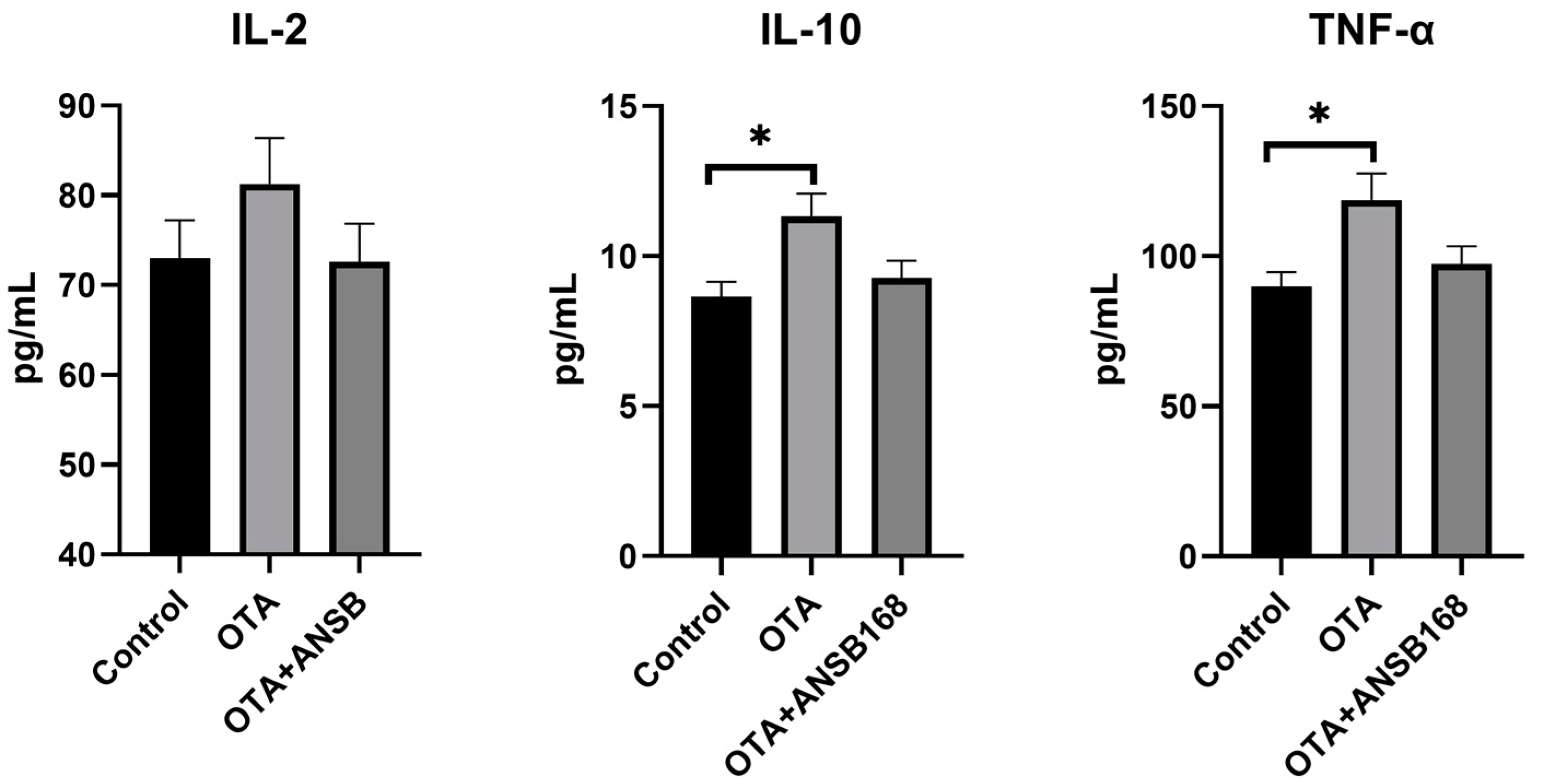
2.5.4. Immune and Inflammatory Response
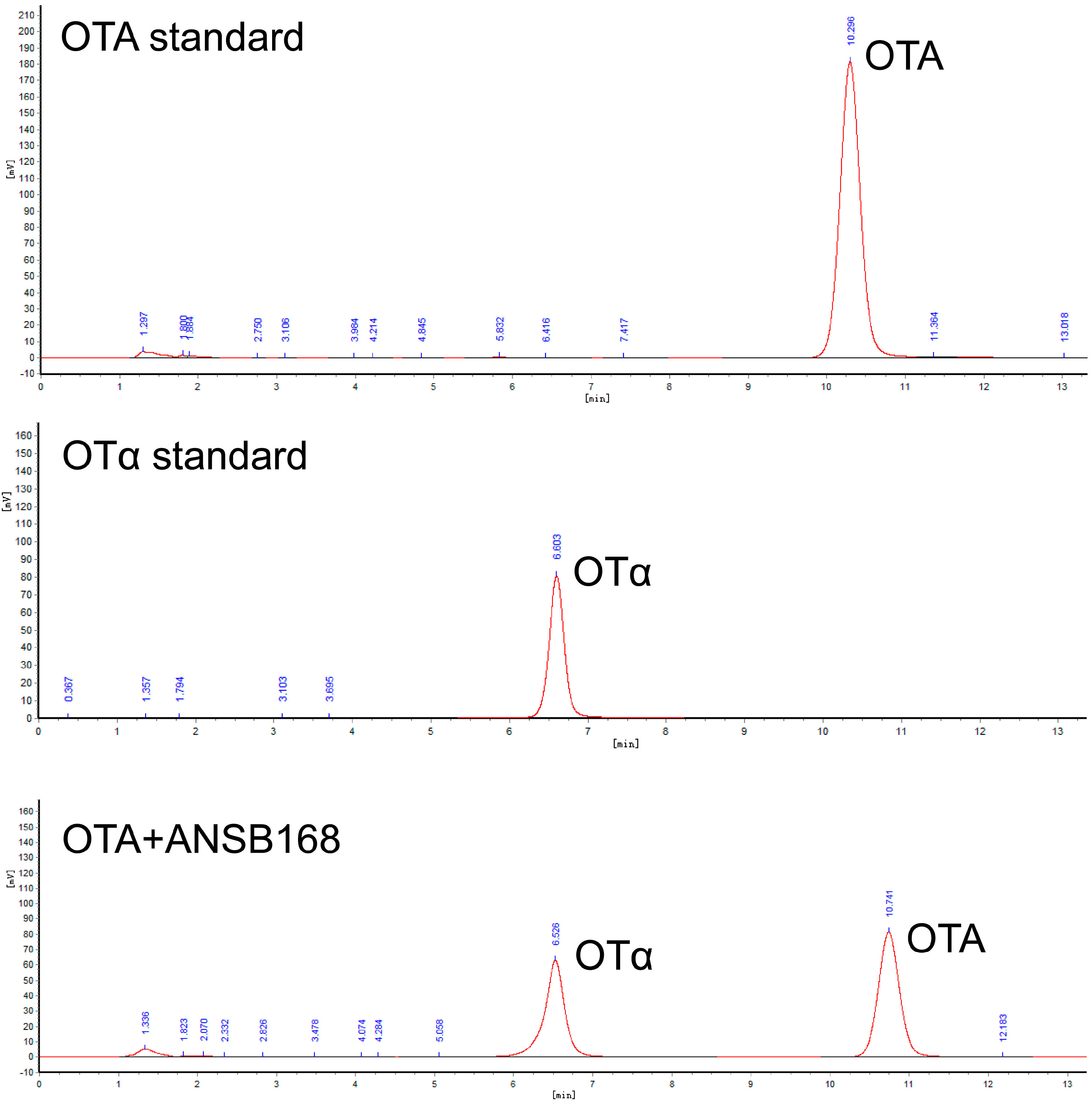
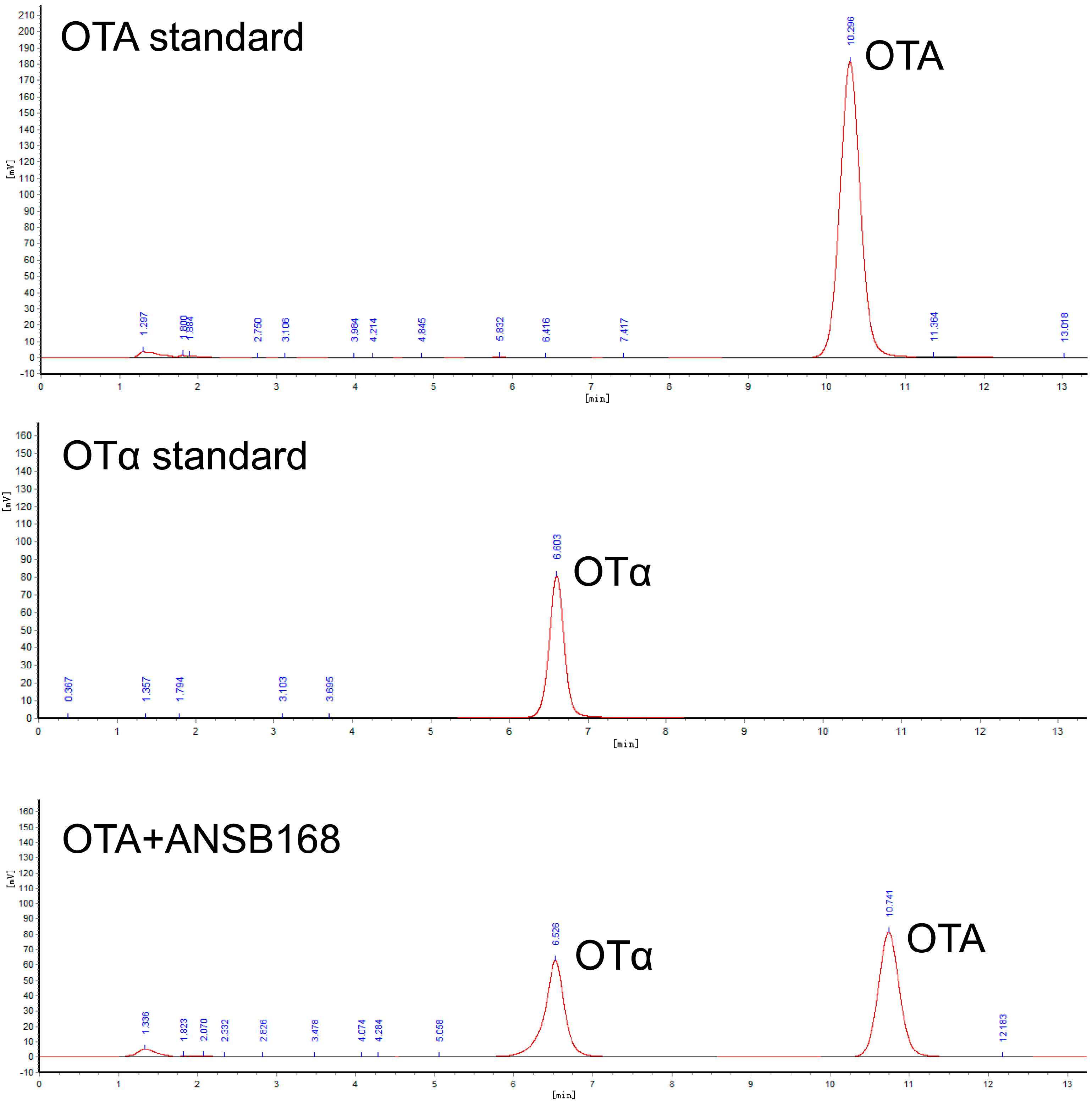
2.5.5. Residues of OTA and OTα in Eggs
3. Discussion
4. Materials and Methods
4.1. Chemicals and Strains
4.2. Enrichment and Isolation of OTA-Degrading Bacteria from Donkey Cecum
4.3. Cloning of d-alanyl-d-alanine Carboxypeptidase DacA and DacB
4.4. Heterologous Expression and Protein Purification
4.5. SDS-PAGE and Western Blot Analysis
4.6. OTA Degradation Activity and Relevant Enzyme Characteristics
4.7. Dietary Treatments of Animal Trial
4.8. Animal Trial in Layers
4.9. Analysis of Serum Parameters
4.10. Extration and Clean-Up of OTA in Feed and Eggs
4.11. High-Performance Liquid Chromatography
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Van Der Merwe, K.J.; Steyn, P.S.; Fourie, L.; Scott, D.B.; Theron, J.J. Ochratoxin A, a Toxic Metabolite produced by Aspergillus ochraceus Wilh. Nature 1965, 205, 1112–1113. [Google Scholar] [CrossRef] [PubMed]
- Abarca, M.L.; Bragulat, M.R.; Castella, G.; Cabañes, F.J. Ochratoxin A production by strains of Aspergillus niger var. niger. Appl. Environ. Microbiol. 1994, 60, 2650–2652. [Google Scholar] [CrossRef] [Green Version]
- Téren, J.; Varga, J.; Hamari, Z.; Rinyu, E.; Kevei, F. Immunochemical detection of ochratoxin A in black Aspergillus strains. Mycopathologia 1996, 134, 171–176. [Google Scholar] [CrossRef]
- Visagie, C.M.; Varga, J.; Houbraken, J.; Meijer, M.; Kocsubé, S.; Yilmaz, N.; Fotedar, R.; Seifert, K.A.; Frisvad, J.C.; Samson, R.A. Ochratoxin production and taxonomy of the yellow aspergilli (Aspergillus section Circumdati). Stud. Mycol. 2014, 78, 1–61. [Google Scholar] [CrossRef] [Green Version]
- Larsen, T.O.; Svendsen, A.; Smedsgaard, J. Biochemical characterization of ochratoxin a-producing strains of the genus penicillium. Appl. Environ. Microbiol. 2001, 67, 3630–3635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabañes, F.J.; Bragulat, M.R.; Castellá, G. Ochratoxin A Producing Species in the Genus Penicillium. Toxins 2010, 2, 1111–1120. [Google Scholar] [CrossRef]
- Akpinar, H.A.; Kahraman, H.; Yaman, I. Ochratoxin a sequentially activates autophagy and the ubiquitin-proteasome System. Toxins 2019, 11, 615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imaoka, T.; Yang, J.; Wang, L.; McDonald, M.G.; Afsharinejad, Z.; Bammler, T.K.; Van Ness, K.; Yeung, C.K.; Rettie, A.E.; Himmelfarb, J.; et al. Microphysiological system modeling of ochratoxin A-associated nephrotoxicity. Toxicology 2020, 444, 152582. [Google Scholar] [CrossRef]
- Li, P.; Li, K.; Zou, C.; Tong, C.; Sun, L.; Cao, Z.; Yang, S.; Lyu, Q. Selenium Yeast Alleviates Ochratoxin A-Induced Hepatotoxicity via Modulation of the PI3K/AKT and Nrf2/Keap1 Signaling Pathways in Chickens. Toxins 2020, 12, 143. [Google Scholar] [CrossRef] [Green Version]
- Le, G.; Yuan, X.; Hou, L.; Ge, L.; Liu, S.; Muhmood, A.; Liu, K.; Lin, Z.; Liu, D.; Gan, F.; et al. Ochratoxin A induces glomerular injury through activating the ERK/NF-κB signaling pathway. Food Chem. Toxicol. 2020, 143, 111516. [Google Scholar] [CrossRef]
- Damiano, S.; Andretta, E.; Longobardi, C.; Prisco, F.; Paciello, O.; Squillacioti, C.; Mirabella, N.; Florio, S.; Ciarcia, R. Effects of Curcumin on the Renal Toxicity Induced by Ochratoxin A in Rats. Antioxidants 2020, 9, 332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marin, D.E.; Pistol, G.; Gras, M.A.; Palade, M.L.; Taranu, I. Comparative effect of ochratoxin A on inflammation and oxidative stress parameters in gut and kidney of piglets. Regul. Toxicol. Pharmacol. 2017, 89, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Marin, D.E.; Motiu, M.; Pistol, G.C.; Gras, M.A.; Israel-Roming, F.; Calin, L.; Stancu, M.; Taranu, I. Diet contaminated with ochratoxin A at the highest level allowed by EU recommendation disturbs liver metabolism in weaned piglets. World Mycotoxin J. 2016, 9, 587–596. [Google Scholar] [CrossRef]
- Hameed, M.R.; Khan, M.Z.; Saleemi, M.K.; Khan, A.; Akhtar, M.; Hassan, Z.-U.; Hussain, Z. Study of ochratoxin A (OTA)-induced oxidative stress markers in broiler chicks. Toxin Rev. 2017, 36, 270–274. [Google Scholar] [CrossRef]
- Khatoon, A.; Khan, M.Z.; Abidin, Z.U.; Bhatti, S.A. Effects of feeding bentonite clay upon ochratoxin A–induced immunosuppression in broiler chicks. Food Addit. Contam. Part A 2018, 35, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Raters, M.; Matissek, R. Thermal stability of aflatoxin B1 and ochratoxin A. Mycotoxin Res. 2008, 24, 130–134. [Google Scholar] [CrossRef]
- Varga, J.; Rigo, K.; Teren, J.; Mesterhazy, A. Recent advances in ochratoxin research—I. Production, detection and occurrence of ochratoxins. Cereal Res. Commun. 2001, 29, 85–92. [Google Scholar] [CrossRef]
- Ameer Sumbal, G.; Hussain Shar, Z.; Hussain Sherazi, S.T.; Sirajuddin; Nizamani, S.M.; Mahesar, S.A. Decontamination of poultry feed from ochratoxin A by UV and sunlight radiations. J. Sci. Food Agric. 2016, 96, 2668–2673. [Google Scholar] [CrossRef]
- Rajendran, R.M.; Umesh, B.; Chirakkal, H. Assessment of H-β zeolite as an ochratoxin binder for poultry. Poult. Sci. 2020, 99, 76–88. [Google Scholar] [CrossRef]
- Zeidan, R.; Ul-Hassan, Z.; Al-Thani, R.; Migheli, Q.; Jaoua, S. In-Vitro Application of a Qatari Burkholderia cepacia strain (QBC03) in the Biocontrol of Mycotoxigenic Fungi and in the Reduction of Ochratoxin A biosynthesis by Aspergillus carbonarius. Toxins 2019, 11, 700. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, H.; Reveron, I.; Doria, F.; Costantini, A.; Rivas, B.D.L.; Muňoz, R.; Garcia-Moruno, E. Degradation of Ochratoxin A by Brevibacterium Species. J. Agric. Food Chem. 2011, 59, 10755–10760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Wang, Y.; Zhao, C.; Wang, J.; Zhang, X. Biodegradation of ochratoxin A by Alcaligenes faecalisisolated from soil. J. Appl. Microbiol. 2017, 123, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Xiong, K.; Zhi, H.-W.; Liu, J.-Y.; Wang, X.-Y.; Zhao, Z.-Y.; Pei, P.-G.; Deng, L.; Xiong, S.-Y. Detoxification of Ochratoxin A by a novel Aspergillus oryzae strain and optimization of its biodegradation. Rev. Argentina Microbiol. 2021, 53, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Varga, J.; Rigó, K.; Téren, J. Degradation of ochratoxin A by Aspergillus species. Int. J. Food Microbiol. 2000, 59, 1–7. [Google Scholar] [CrossRef]
- Jiang, C.; Shi, J.; Liu, Y.; Zhu, C. Inhibition of Aspergillus carbonarius and fungal contamination in table grapes using Bacillus subtilis. Food Control 2014, 35, 41–48. [Google Scholar] [CrossRef]
- Ammar, H.; El-Desouky, T. Green synthesis of nanosilver particles by Aspergillus terreus HA1N and Penicillium expansum HA2N and its antifungal activity against mycotoxigenic fungi. J. Appl. Microbiol. 2016, 121, 89–100. [Google Scholar] [CrossRef] [Green Version]
- Hadidane, R.; Bacha, H.; Creppy, E.; Hammami, M.; Ellouze, F.; Dirheimer, G. Isolation and structure determination of natural analogues of the mycotoxin ochratoxin A produced by Aspergillus ochraceus. Toxicology 1992, 76, 233–243. [Google Scholar] [CrossRef]
- Marín, S.; Ramos, A.J.; Cano-Sancho, G.; Sanchis, V. Mycotoxins: Occurrence, toxicology, and exposure assessment. Food Chem. Toxicol. 2013, 60, 218–237. [Google Scholar] [CrossRef]
- Parker, R.W.; Phillips, T.D.; Kubena, L.F.; Russell, L.H.; Heidelbaugh, N.D. Inhibition of pancreatic carboxypeptidase A: A possible mechanism of interaction between penicillic acid and ochratoxin A. J. Environ. Sci. Health Part B 1982, 17, 77–91. [Google Scholar] [CrossRef]
- Xiong, L.; Peng, M.; Zhao, M.; Liang, Z. Truncated expression of a carboxypeptidase a from bovine improves its enzymatic properties and detoxification efficiency of ochratoxin A. Toxins 2020, 12, 680. [Google Scholar] [CrossRef]
- Ma, L.; Qiu, X.; Li, Y.; Tang, S.; Shen, W.; Xing, C.; Kong, D.; Sheng, J. Carboxypeptidase A immobilization with zeolitic imidazolate framework for enhancement of ochratoxin A degradation ability. Food Agric. Immunol. 2020, 31, 587–599. [Google Scholar] [CrossRef]
- Dellafiora, L.; Gonaus, C.; Streit, B.; Galaverna, G.; Moll, W.-D.; Vogtentanz, G.; Schatzmayr, G.; Dall’Asta, C.; Prasad, S. An in silico target fishing approach to identify novel ochratoxin a hydrolyzing enzyme. Toxins 2020, 12, 258. [Google Scholar] [CrossRef]
- Chang, X.; Wu, Z.; Wu, S.; Dai, Y.; Sun, C. Degradation of ochratoxin A by Bacillus amyloliquefaciens ASAG1. Food Addit. Contam. Part A 2015, 32, 564–571. [Google Scholar] [CrossRef]
- Hu, H.; Jia, X.; Wang, Y.; Xiong, L.; Peng, M.; Liang, Z. Detoxification of ochratoxin a by an expressed carboxypeptidase and some isolated peptides from Bacillus subtilis CW14. Toxicon 2019, 158, S67. [Google Scholar] [CrossRef]
- Wei, W.; Qian, Y.; Wu, Y.; Chen, Y.; Peng, C.; Luo, M.; Xu, J.; Zhou, Y. Detoxification of ochratoxin A by Lysobacter sp. CW239 and characteristics of a novel degrading gene carboxypeptidase cp4. Environ. Pollut. 2020, 258, 113677. [Google Scholar] [CrossRef]
- Petruzzi, L.; Bevilacqua, A.; Baiano, A.; Beneduce, L.; Corbo, M.R.; Sinigaglia, M. In vitro removal of ochratoxin A by two strains of Saccharomyces cerevisiae and their performances under fermentative and stressing conditions. J. Appl. Microbiol. 2014, 116, 60–70. [Google Scholar] [CrossRef]
- Ferenczi, S.; Cserháti, M.; Krifaton, C.; Szoboszlay, S.; Kukolya, J.; Szőke, Z.; Kőszegi, B.; Albert, M.; Barna, T.; Mézes, M.; et al. A New Ochratoxin A Biodegradation Strategy Using Cupriavidus basilensis Őr16 Strain. PLoS ONE 2014, 9, e109817. [Google Scholar] [CrossRef]
- Fuchs, S.; Sontag, G.; Stidl, R.; Ehrlich, V.; Kundi, M.; Knasmüller, S. Detoxification of patulin and ochratoxin A, two abundant mycotoxins, by lactic acid bacteria. Food Chem. Toxicol. 2008, 46, 1398–1407. [Google Scholar] [CrossRef]
- Liuzzi, V.C.; Fanelli, F.; Tristezza, M.; Haidukowski, M.; Picardi, E.; Manzari, C.; Lionetti, C.; Grieco, F.; Logrieco, A.F.; Thon, M.R.; et al. Transcriptional Analysis of Acinetobacter sp. neg1 Capable of Degrading Ochratoxin A. Front. Microbiol. 2017, 7, 2162. [Google Scholar] [CrossRef] [Green Version]
- Petchkongkaew, A.; Taillandier, P.; Gasaluck, P.; Lebrihi, A. Isolation of Bacillus spp. from Thai fermented soybean (Thuanao): Screening for aflatoxin B1and ochratoxin A detoxification. J. Appl. Microbiol. 2008, 104, 1495–1502. [Google Scholar] [CrossRef] [Green Version]
- Seddi, R.; Chaix, J.-C.; Puigserver, A.; Guo, X.-J. Expression of a soluble and activatable form of bovine procarboxypeptidase A in Escherichia coli. Protein Expr. Purif. 2003, 27, 220–228. [Google Scholar] [CrossRef]
- Michaelis, L.; Menten, M.L.; Johnson, K.A.; Goody, R.S. The Original Michaelis Constant: Translation of the 1913 Michaelis–Menten Paper. Biochemistry 2011, 50, 8264–8269. [Google Scholar] [CrossRef] [Green Version]
- Abrunhosa, L.; Santos, L.; Venâncio, A. Degradation of Ochratoxin A by Proteases and by a Crude Enzyme of Aspergillus niger. Food Biotechnol. 2006, 20, 231–242. [Google Scholar] [CrossRef] [Green Version]
- Stoev, S.D. Studies on some feed additives and materials giving partial protection against the suppressive effect of ochratoxin A on egg production of laying hens. Res. Vet. Sci. 2010, 88, 486–491. [Google Scholar] [CrossRef]
- Pozzo, L.; Salamano, G.; Mellia, E.; Gennero, M.S.; Doglione, L.; Cavallarin, L.; Tarantola, M.; Forneris, G.; Schiavone, A. Feeding a diet contaminated with ochratoxin A for chickens at the maximum level recommended by the EU for poultry feeds (0.1 mg/kg). 1. Effects on growth and slaughter performance, haematological and serum traits. J. Anim. Physiol. Anim. Nutr. 2013, 97, 13–22. [Google Scholar] [CrossRef]
- Zahoor ul, H.; Khan, M.Z.; Khan, A.; Javed, I. Pathological Responses of White Leghorn Breeder Hens Kept on Ochratoxin A Contaminated Feed. Pak. Vet. J. 2010, 30, 118–123. [Google Scholar]
- Wang, W.; Zhai, S.; Xia, Y.; Wang, H.; Ruan, D.; Zhou, T.; Zhu, Y.; Zhang, H.; Zhang, M.; Ye, H.; et al. Ochratoxin A induces liver inflammation: Involvement of intestinal microbiota. Microbiome 2019, 7, 1–14. [Google Scholar] [CrossRef]
- Denli, M.; Blandon, J.C.; Guynot, M.E.; Salado, S.; Perez, J.F. Efficacy of a new ochratoxin-binding agent (OcraTox) to counteract the deleterious effects of ochratoxin a in laying hens. Poult. Sci. 2008, 87, 2266–2272. [Google Scholar] [CrossRef]
- Armorini, S.; Al-Qudah, K.M.; Altafini, A.; Zaghini, A.; Roncada, P. Biliary ochratoxin A as a biomarker of ochratoxin exposure in laying hens: An experimental study after administration of contaminated diets. Res. Veter Sci. 2015, 100, 265–270. [Google Scholar] [CrossRef]
- Giancarlo, B.; Elisabetta, B.; Edmondo, C.; Valeriana, C.; Giuseppina, T. Determination of ochratoxin A in eggs and target tissues of experimentally drugged hens using HPLC–FLD. Food Chem. 2011, 126, 1278–1282. [Google Scholar] [CrossRef]
- Piskorska-Pliszczyńska, J.; Juszkiewicz, T. Tissue deposition and passage into eggs of ochratoxin A in Japanese quail. J. Environ. Pathol. Toxicol. Oncol. 1990, 10, 8–10. [Google Scholar] [PubMed]
- Gao, C.-Q.; Shi, H.-Q.; Xie, W.-Y.; Zhao, L.-H.; Zhang, J.-Y.; Ji, C.; Ma, Q.-G. Dietary supplementation with acidifiers improves the growth performance, meat quality and intestinal health of broiler chickens. Anim. Nutr. 2021, 7, 762–769. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Zhao, F.; Chen, J.; Zou, Y.; Wang, Y.M.; Liu, S.B.; Tan, H.Z. Research Note: Effect of dietary cottonseed meal and soybean oil concentration on digesta passage time and amino acids digestibility in roosters. Poult. Sci. 2021, 100, 101446. [Google Scholar] [CrossRef] [PubMed]
- Van der Klis, J.D.; Verstegen, M.; De Wit, W. Absorption of minerals and retention time of dry matter in the gastrointestinal tract of broilers. Poult. Sci. 1990, 69, 2185–2194. [Google Scholar] [CrossRef]
- Haridhasapavalan, K.K.; Sundaravadivelu, P.K.; Thummer, R.P. Codon optimization, cloning, expression, purification, and secondary structure determination of human ETS2 Transcription Factor. Mol. Biotechnol. 2020, 62, 485–494. [Google Scholar] [CrossRef]
- Sizoo, E.A.; Van Egmond, H.P. Analysis of duplicate 24-hour diet samples for aflatoxin B1, aflatoxin M1and ochratoxin A. Food Addit. Contam. 2005, 22, 163–172. [Google Scholar] [CrossRef]
- Hassan, Z.U.; Khan, M.Z.; Khan, A.; Javed, I.; Hussain, Z. Effects of individual and combined administration of ochratoxin A and aflatoxin B1 in tissues and eggs of White Leghorn breeder hens. J. Sci. Food Agric. 2012, 92, 1540–1544. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qing, H.; Huo, X.; Huang, S.; Zhao, L.; Zhang, J.; Ji, C.; Ma, Q. Bacillus subtilis ANSB168 Producing d-alanyl-d-alanine Carboxypeptidase Could Alleviate the Immune Injury and Inflammation Induced by Ochratoxin A. Int. J. Mol. Sci. 2021, 22, 12059. https://doi.org/10.3390/ijms222112059
Qing H, Huo X, Huang S, Zhao L, Zhang J, Ji C, Ma Q. Bacillus subtilis ANSB168 Producing d-alanyl-d-alanine Carboxypeptidase Could Alleviate the Immune Injury and Inflammation Induced by Ochratoxin A. International Journal of Molecular Sciences. 2021; 22(21):12059. https://doi.org/10.3390/ijms222112059
Chicago/Turabian StyleQing, Hanrui, Xueting Huo, Shimeng Huang, Lihong Zhao, Jianyun Zhang, Cheng Ji, and Qiugang Ma. 2021. "Bacillus subtilis ANSB168 Producing d-alanyl-d-alanine Carboxypeptidase Could Alleviate the Immune Injury and Inflammation Induced by Ochratoxin A" International Journal of Molecular Sciences 22, no. 21: 12059. https://doi.org/10.3390/ijms222112059
APA StyleQing, H., Huo, X., Huang, S., Zhao, L., Zhang, J., Ji, C., & Ma, Q. (2021). Bacillus subtilis ANSB168 Producing d-alanyl-d-alanine Carboxypeptidase Could Alleviate the Immune Injury and Inflammation Induced by Ochratoxin A. International Journal of Molecular Sciences, 22(21), 12059. https://doi.org/10.3390/ijms222112059