
Simulated Microgravity Subtlety Changes Monoamine Function across the Rat Brain



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
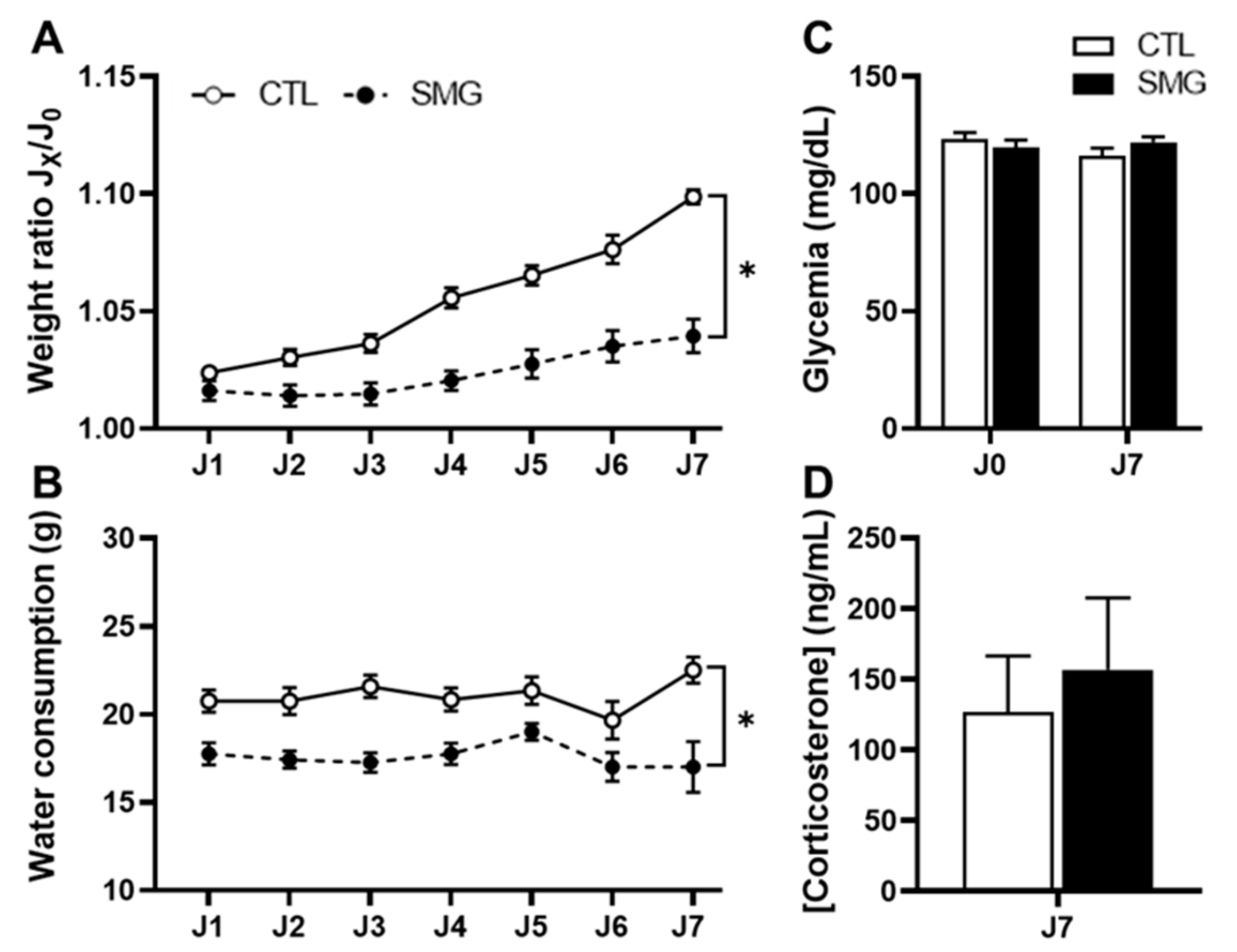
2.1. Effect of Simulated Microgravity on Physiological Parameters
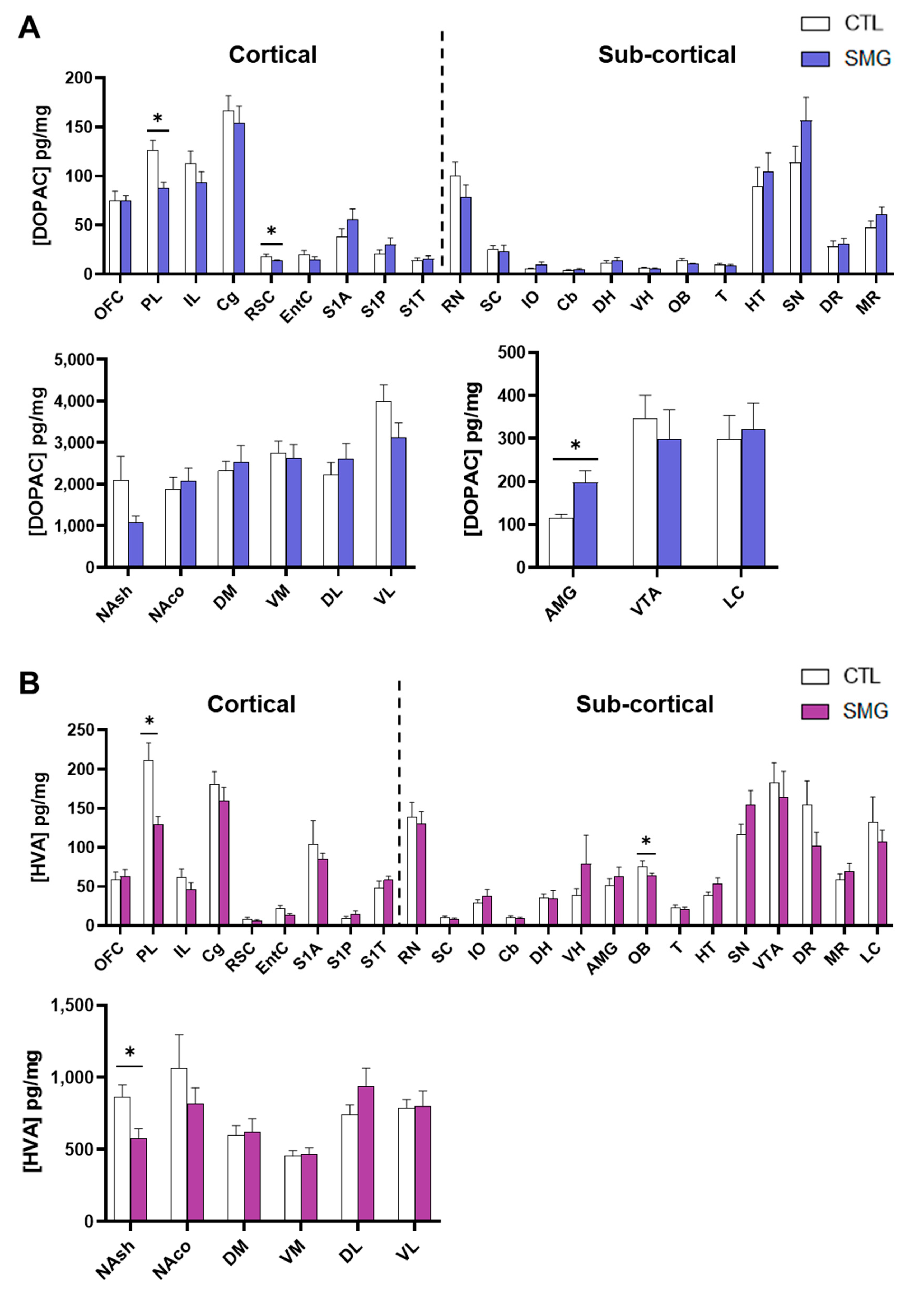
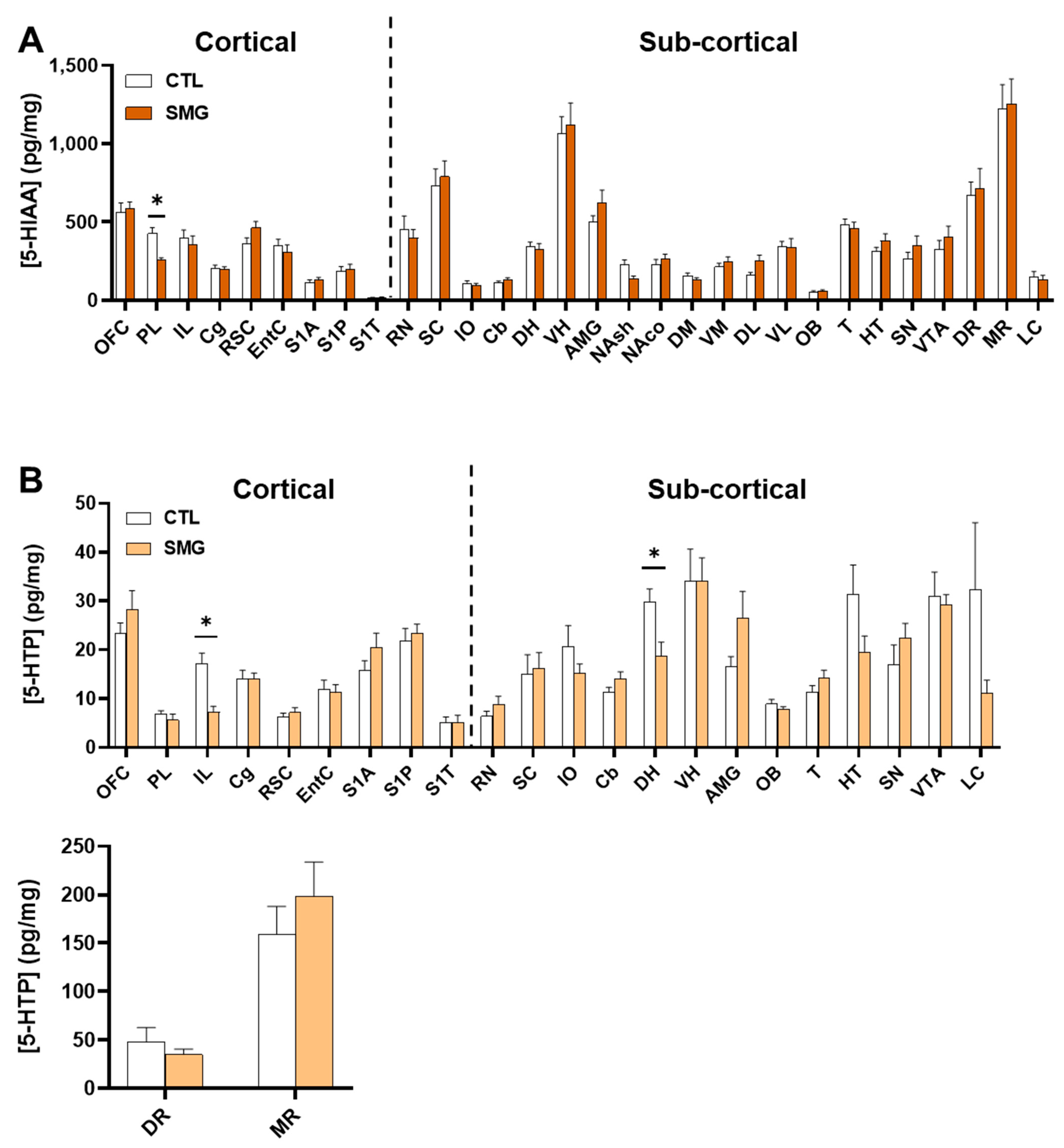
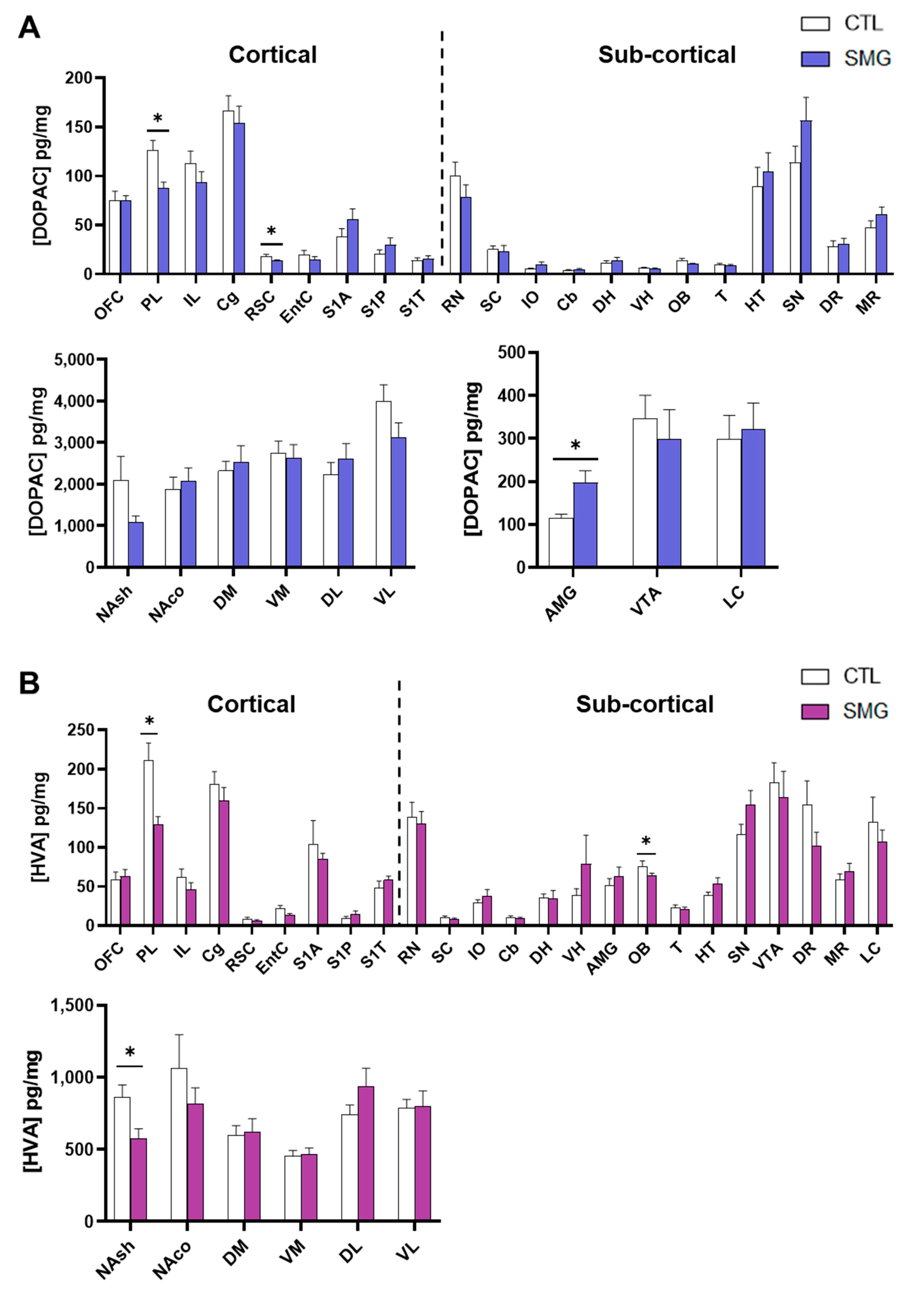
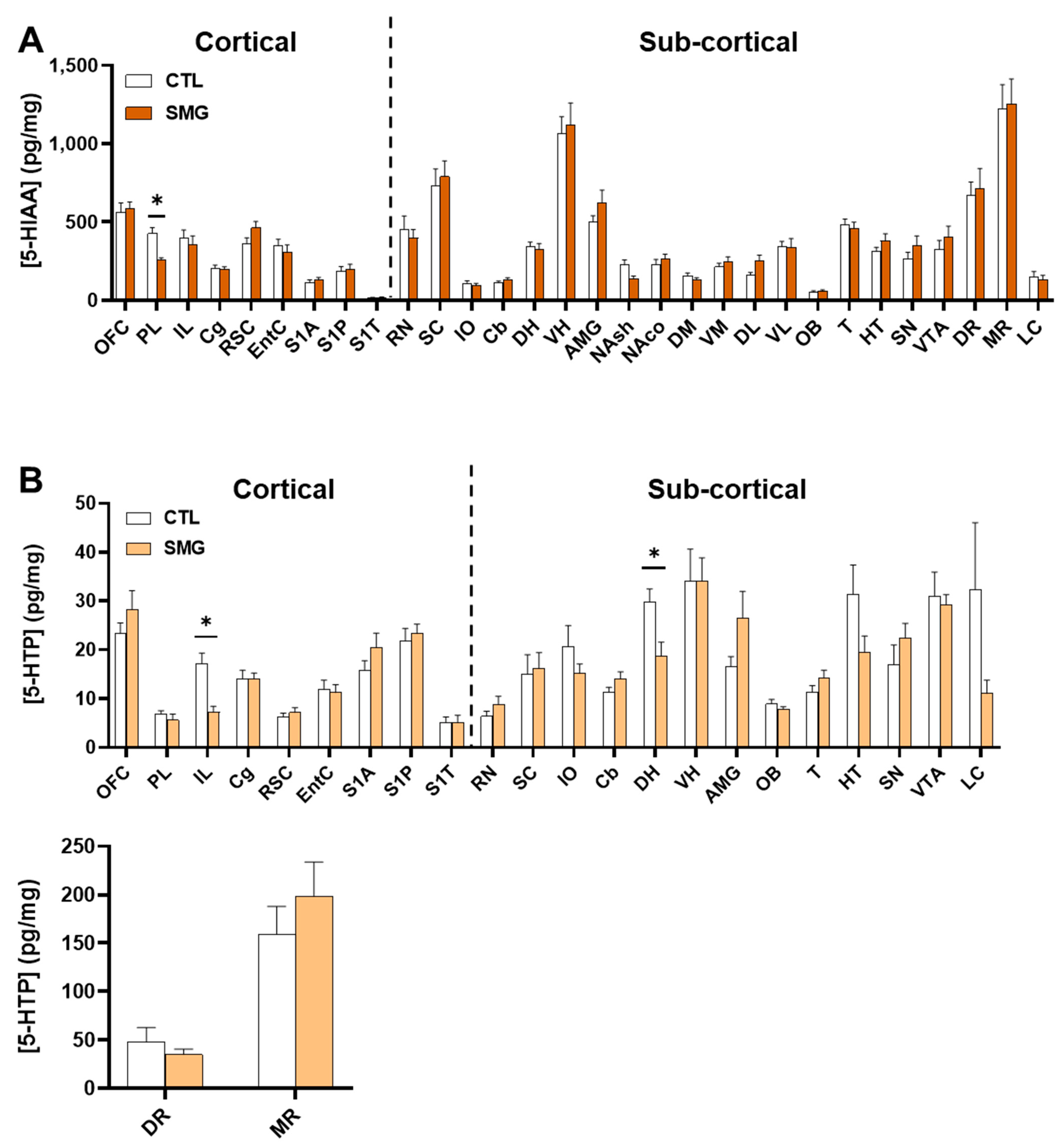
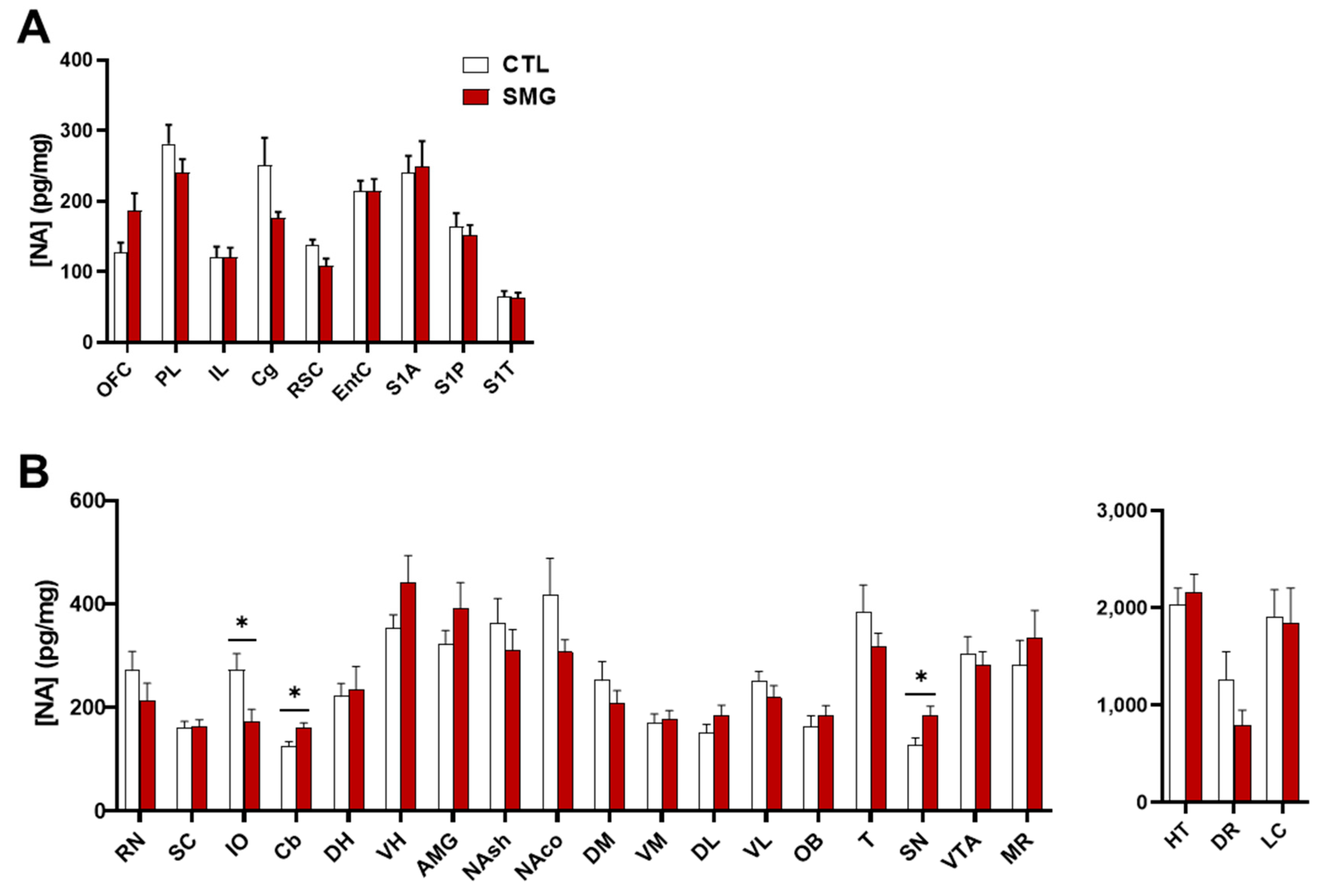
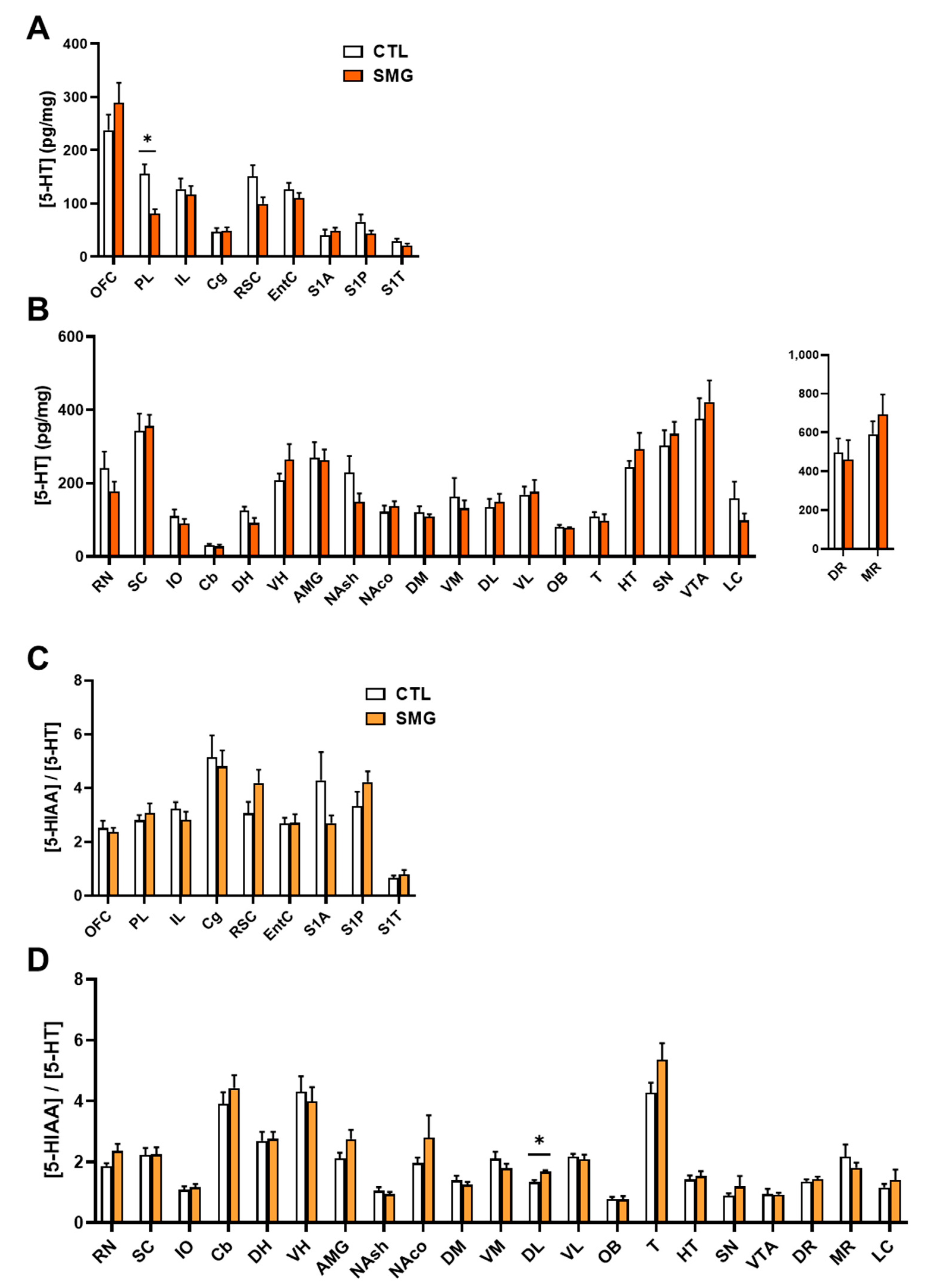
2.2. Effect of Simulated Microgravity on Tissue Levels of Monoamines and Derivatives across the Brain
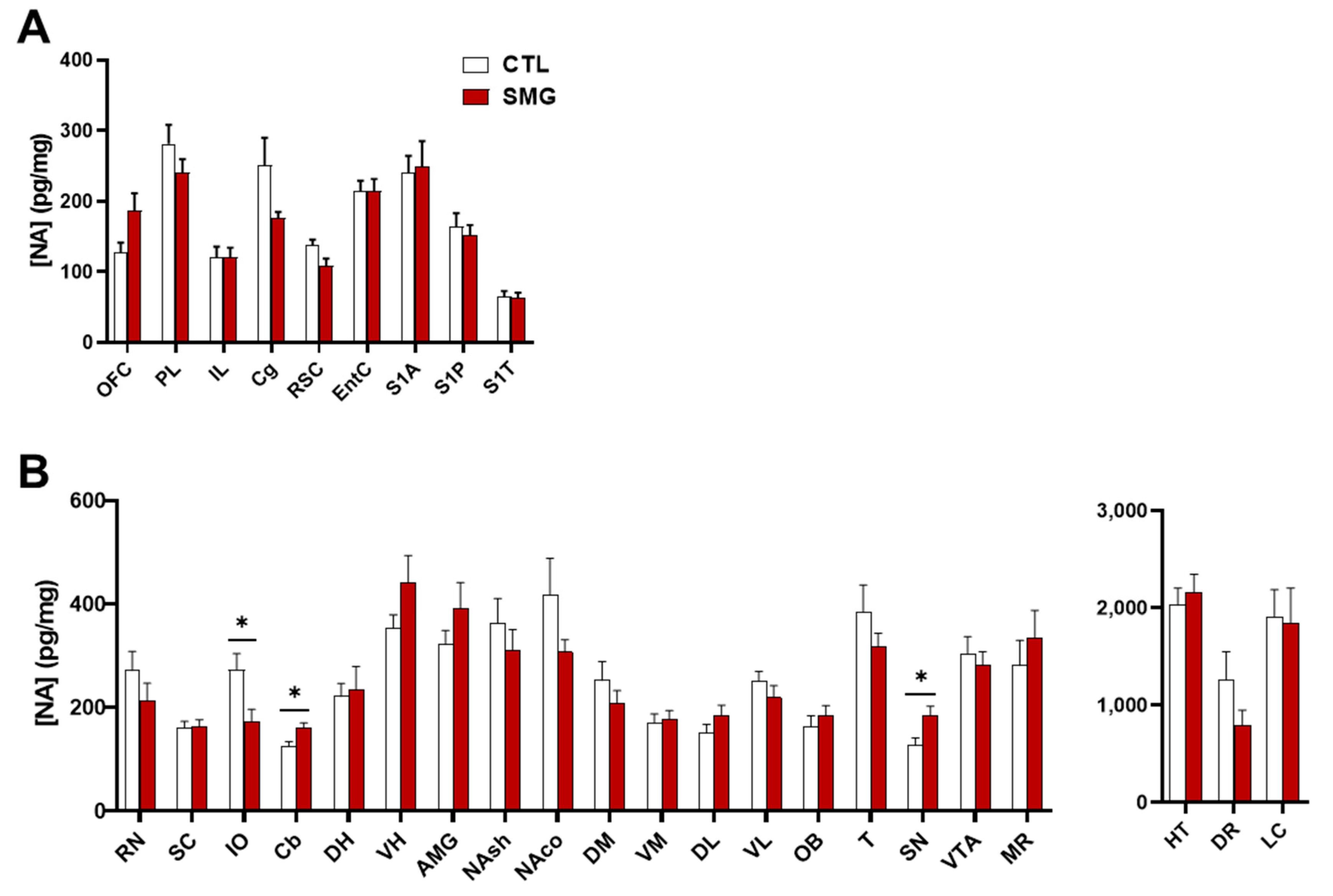
2.2.1. NA System
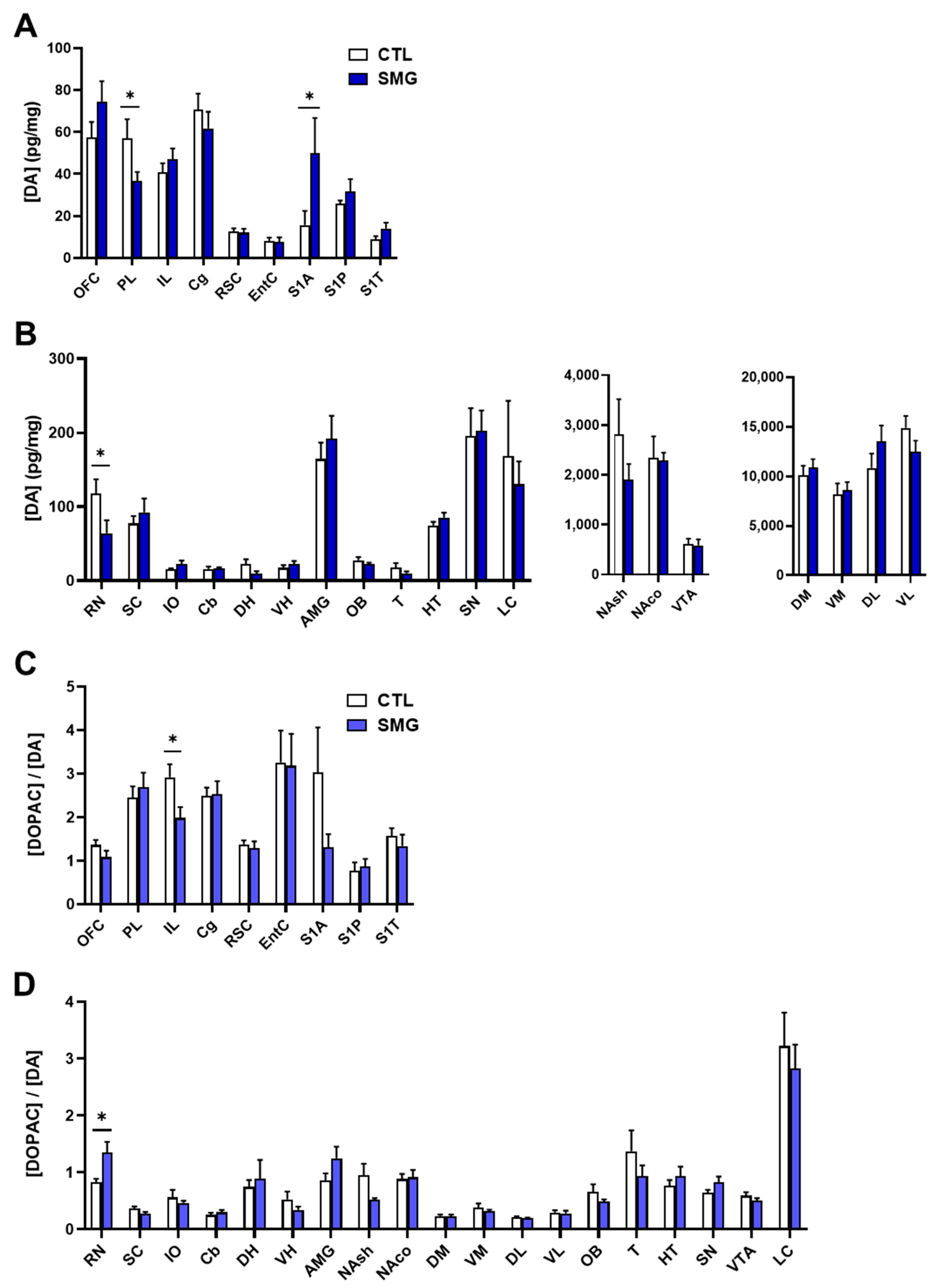
2.2.2. DA System
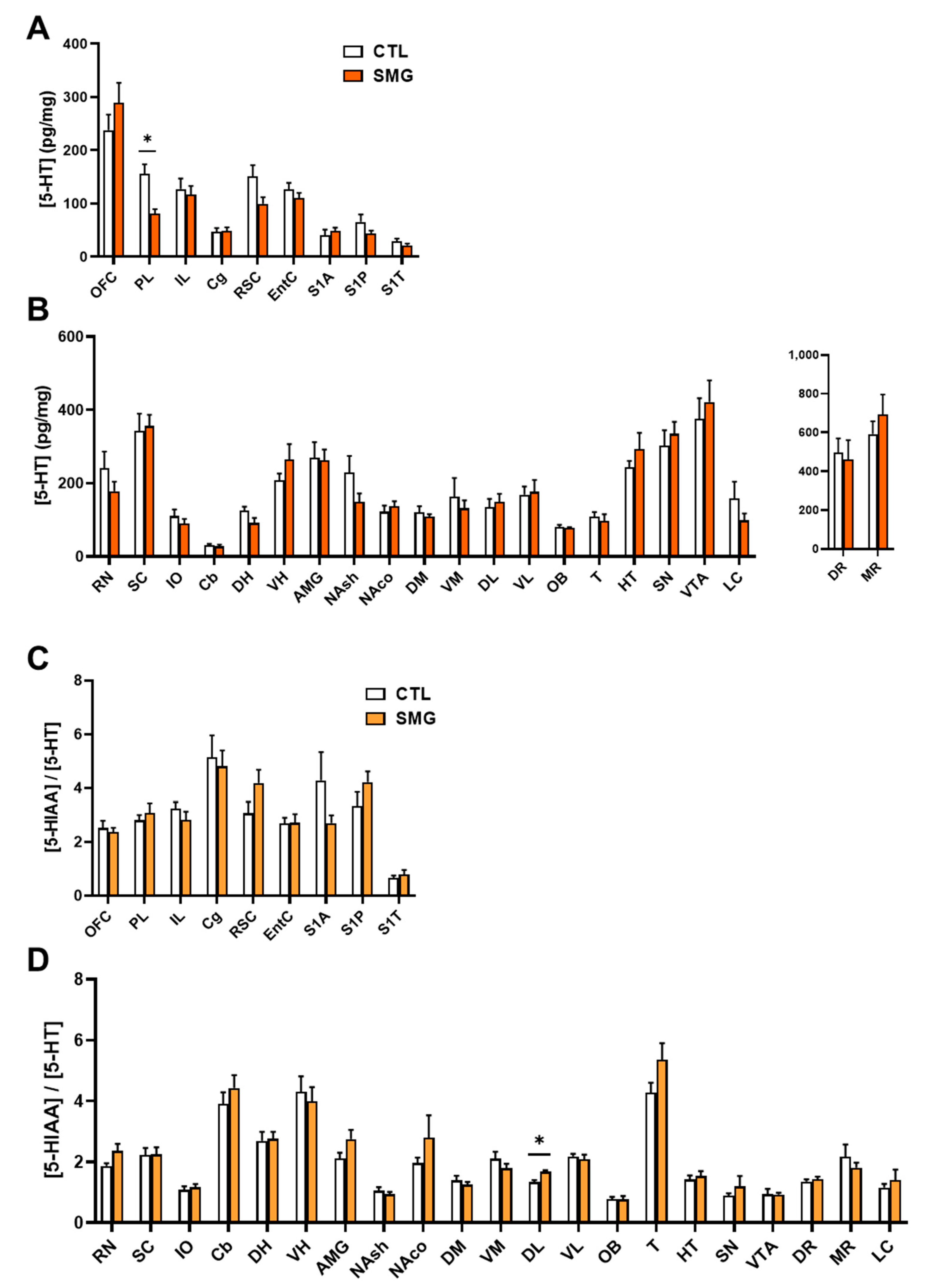
2.2.3. 5-HT System
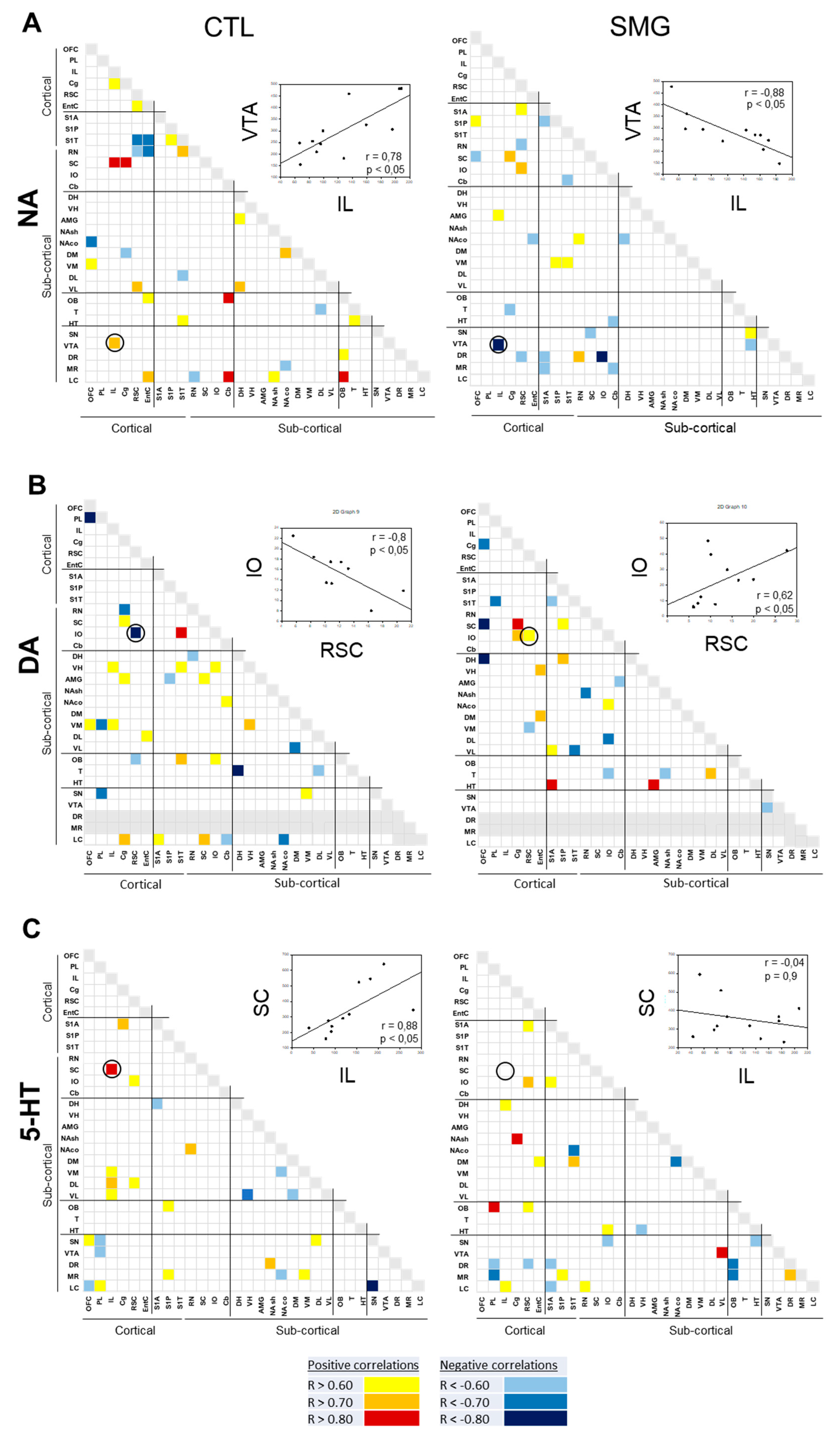
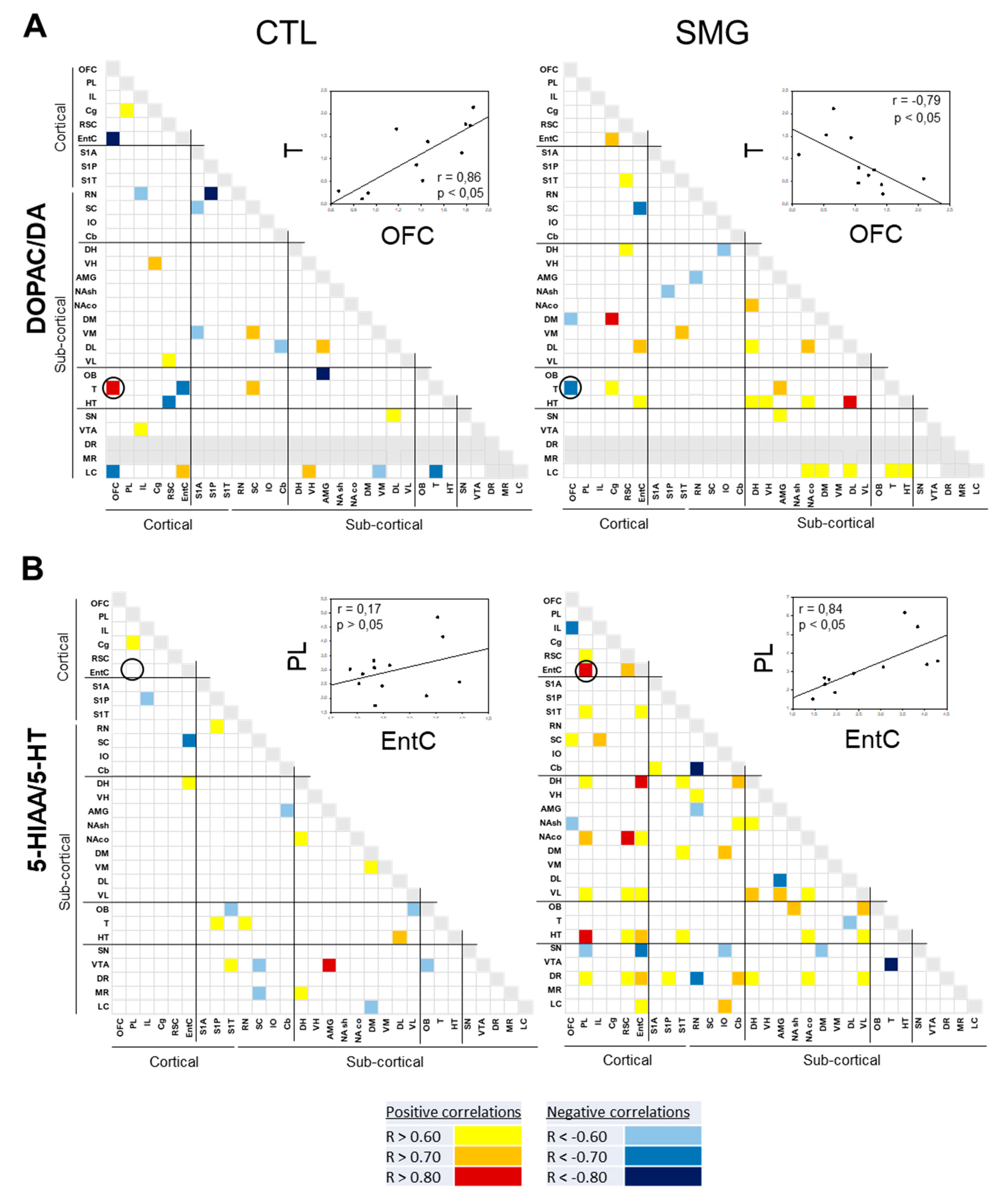
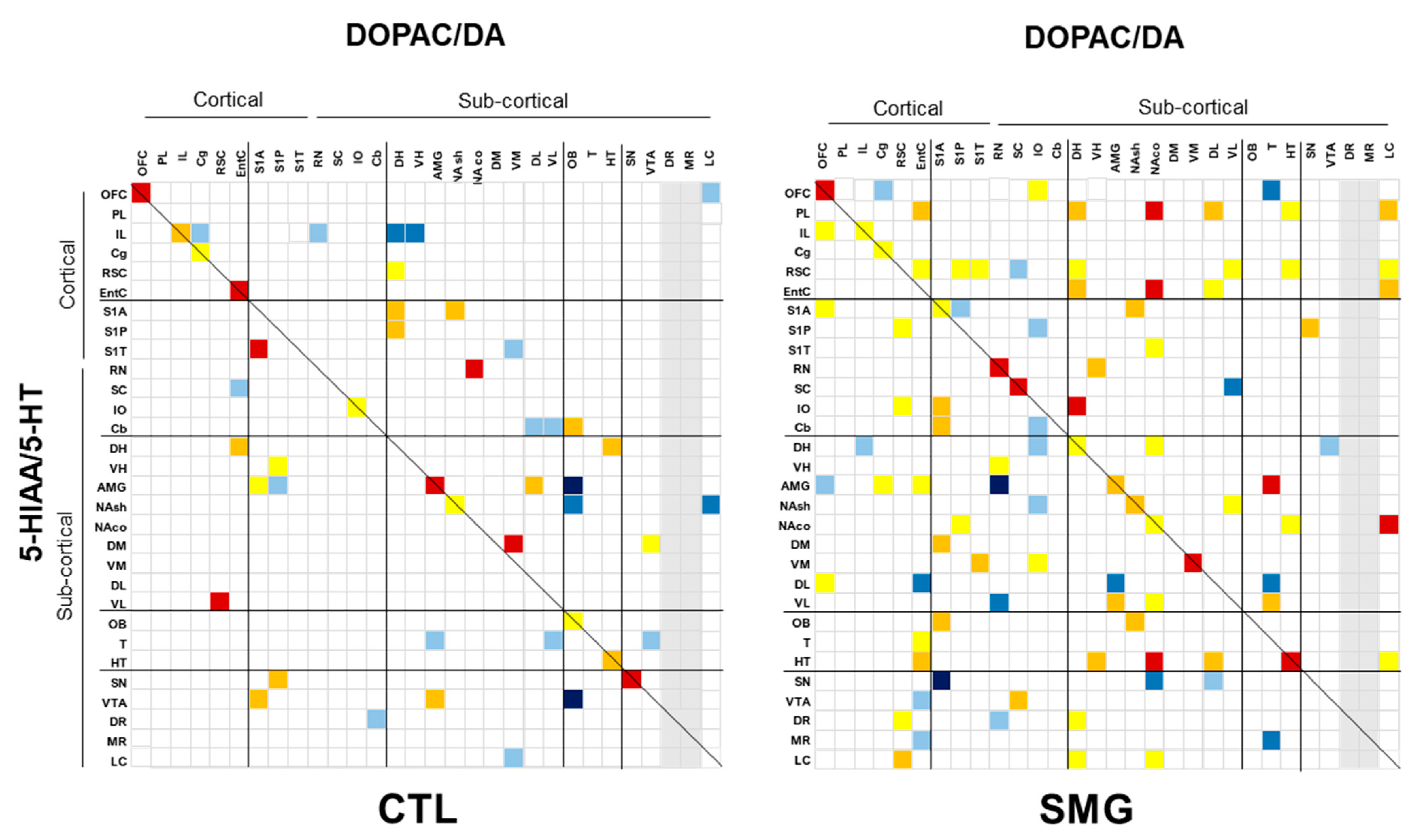
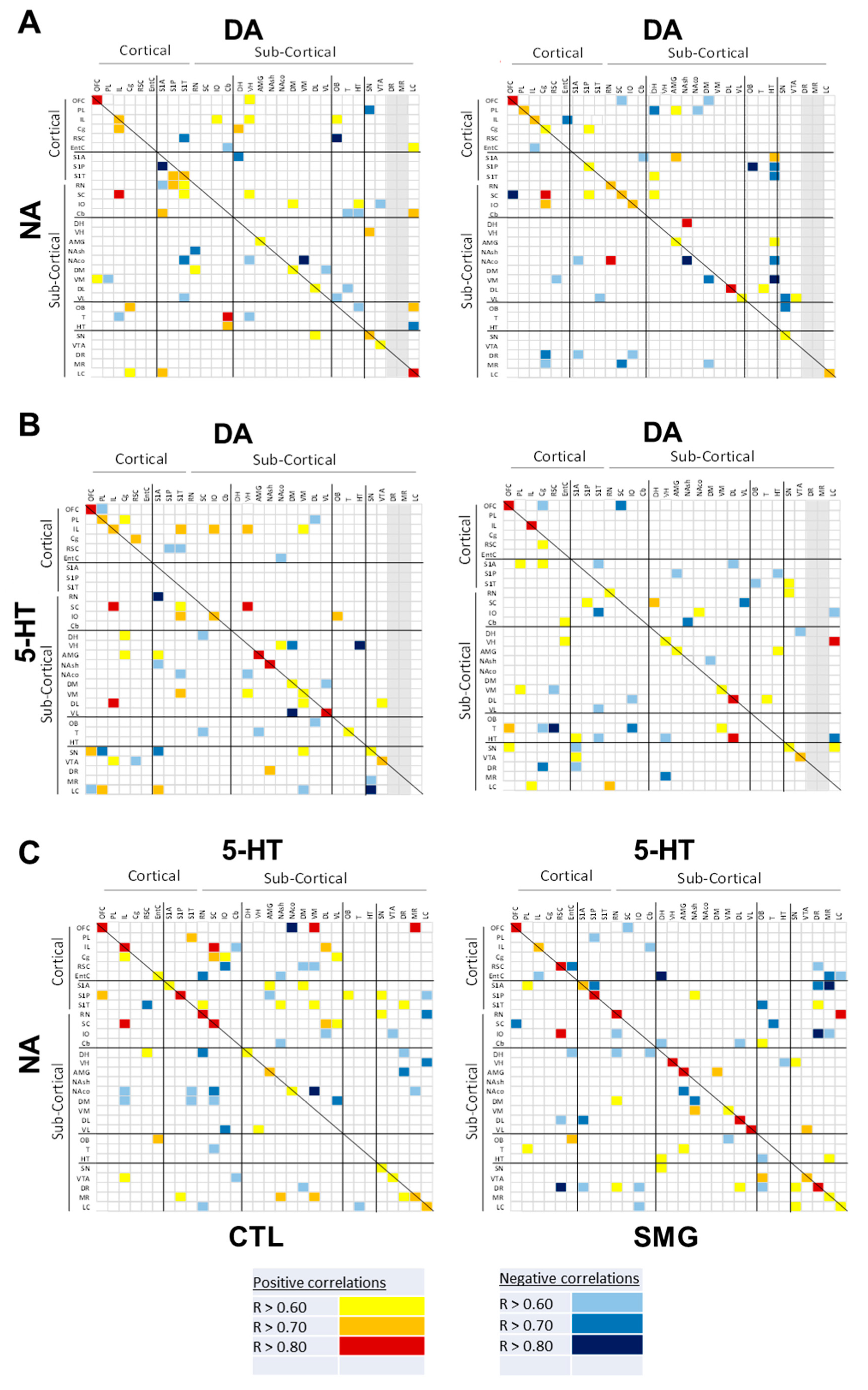
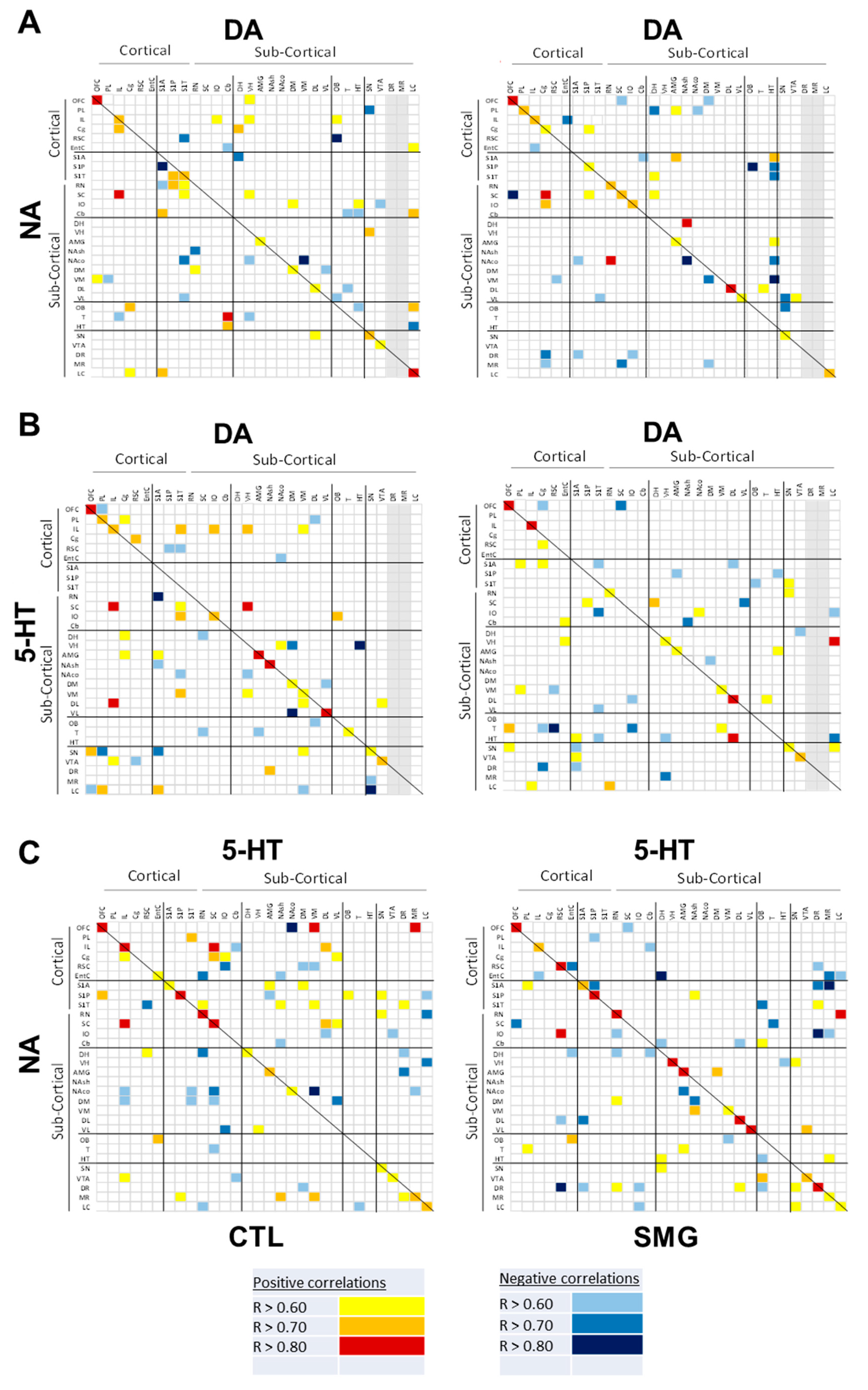
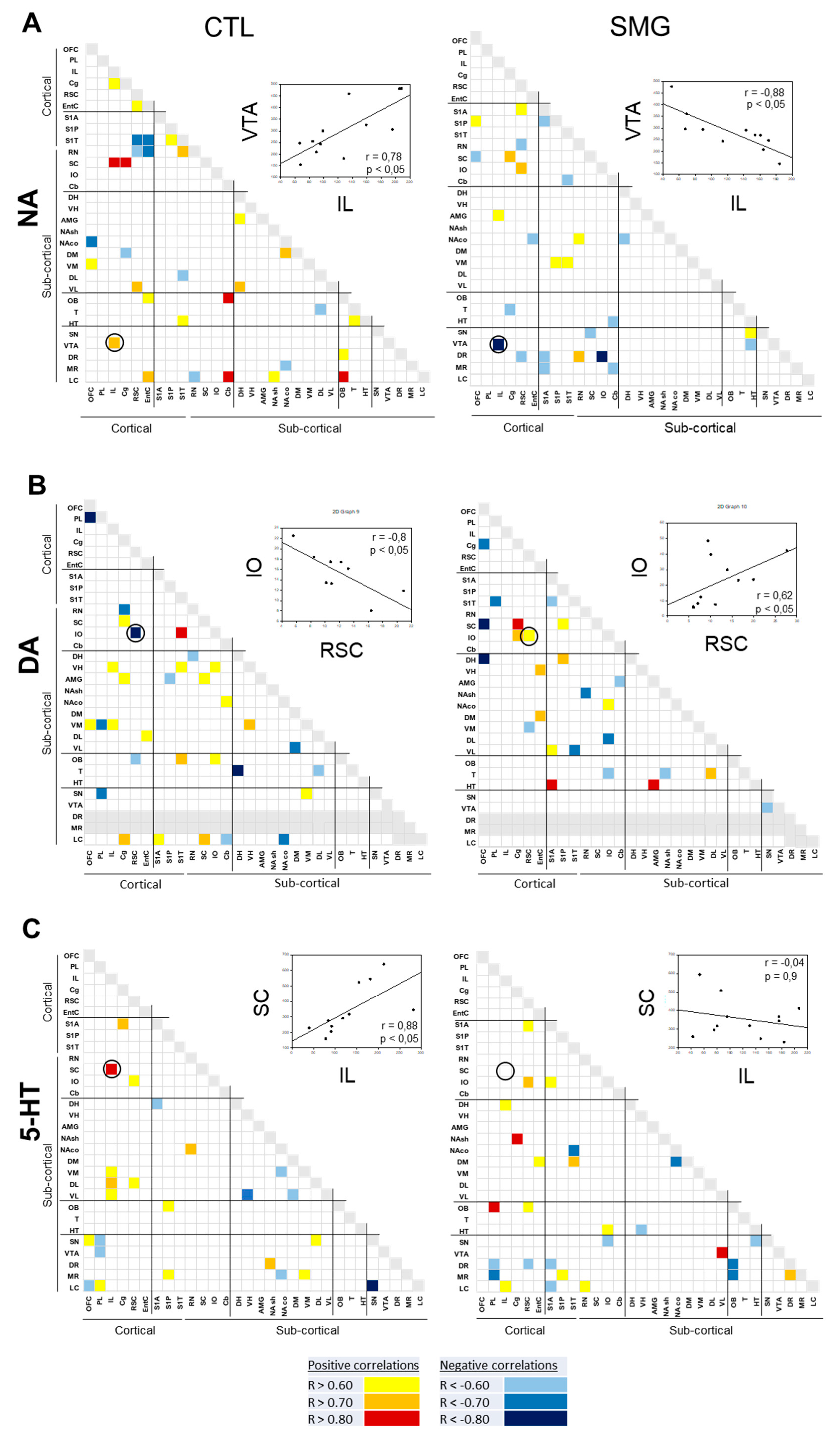
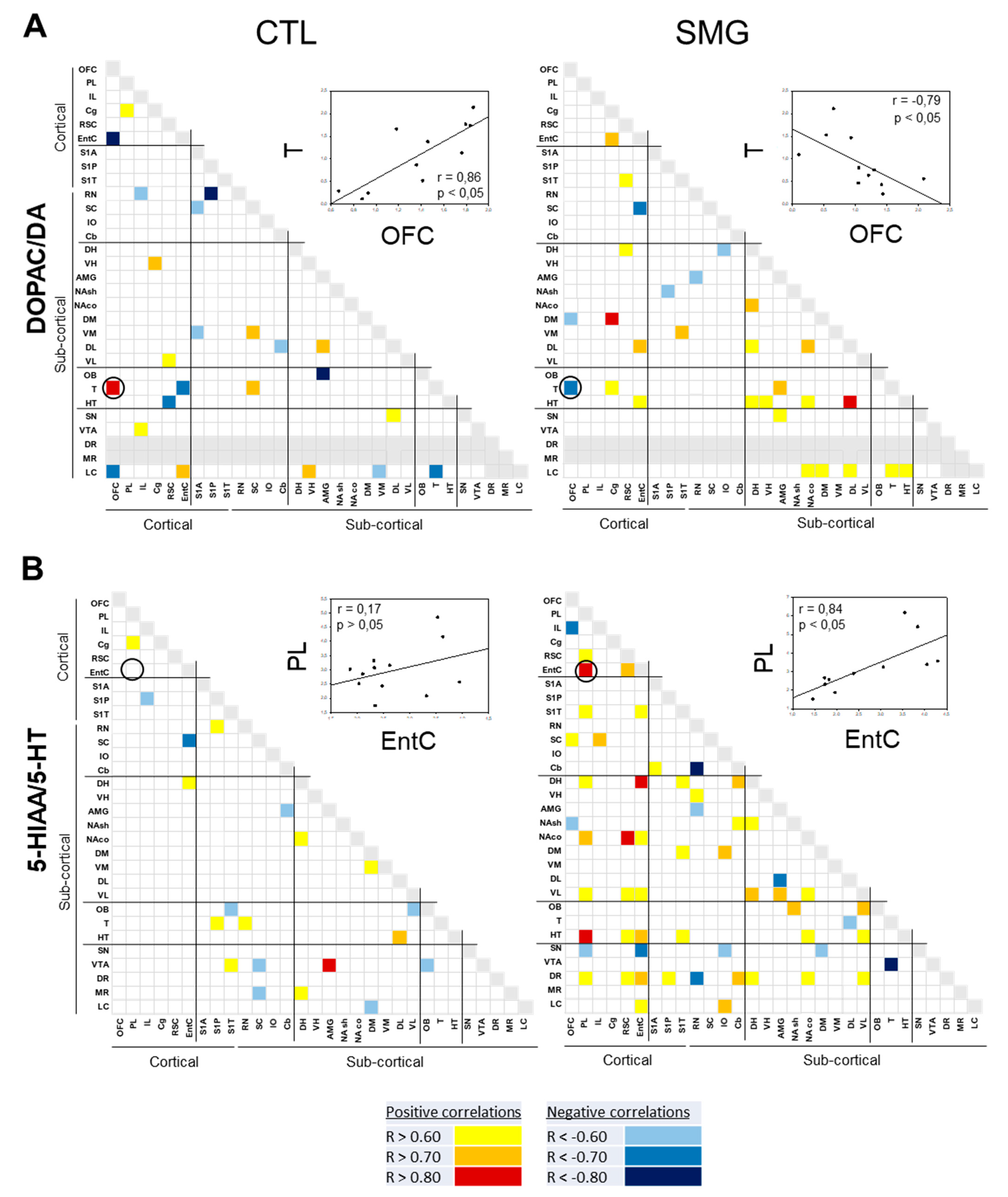
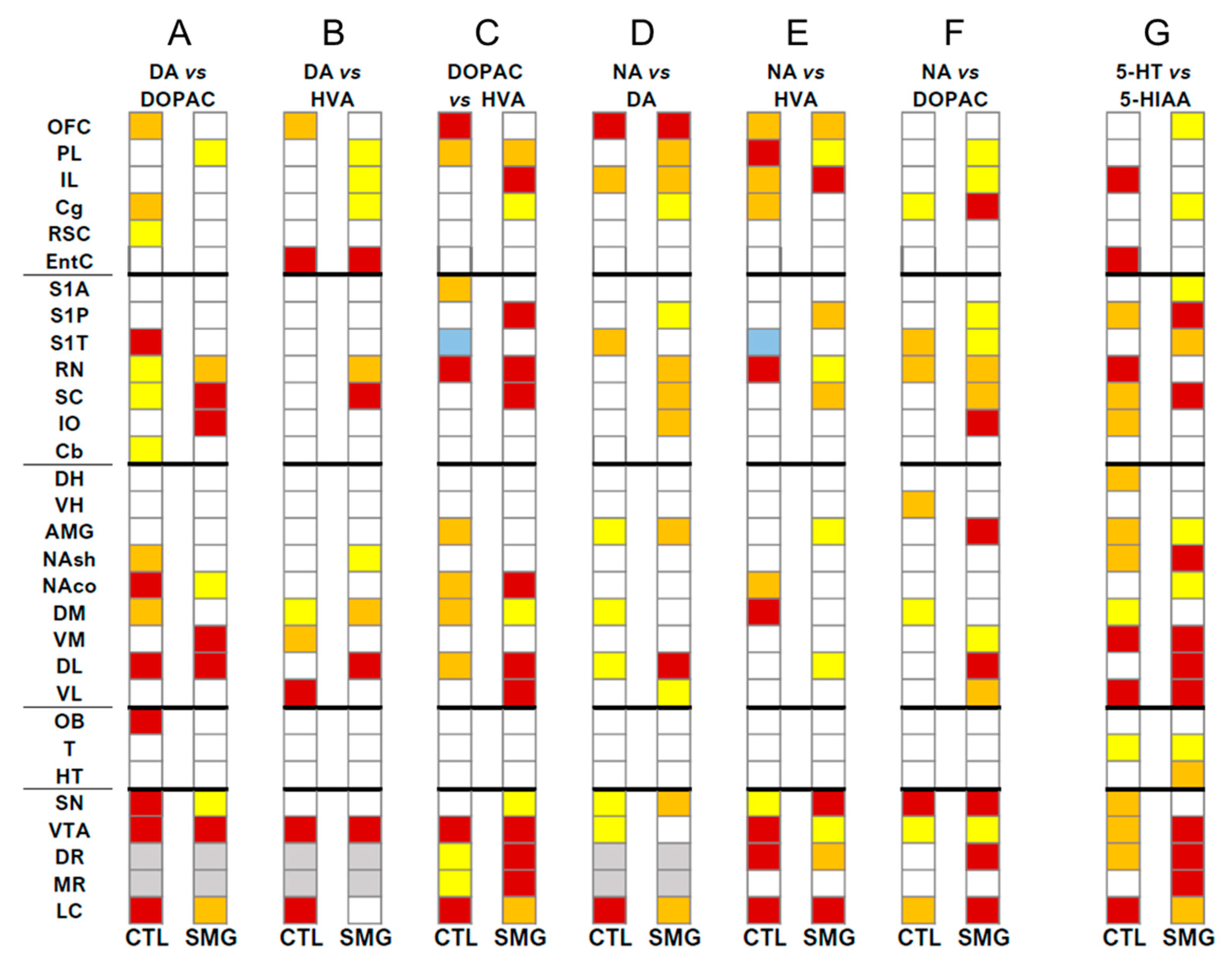
2.3. Effect of Simulated Microgravity on the Qualitative Distribution of Monoamines and Derivatives across the Brain
2.3.1. Within a Single Monoaminergic Modality
2.3.2. Between Indexes across the Brain
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Procedure for Simulated Microgravity
4.3. Physiological Measurements
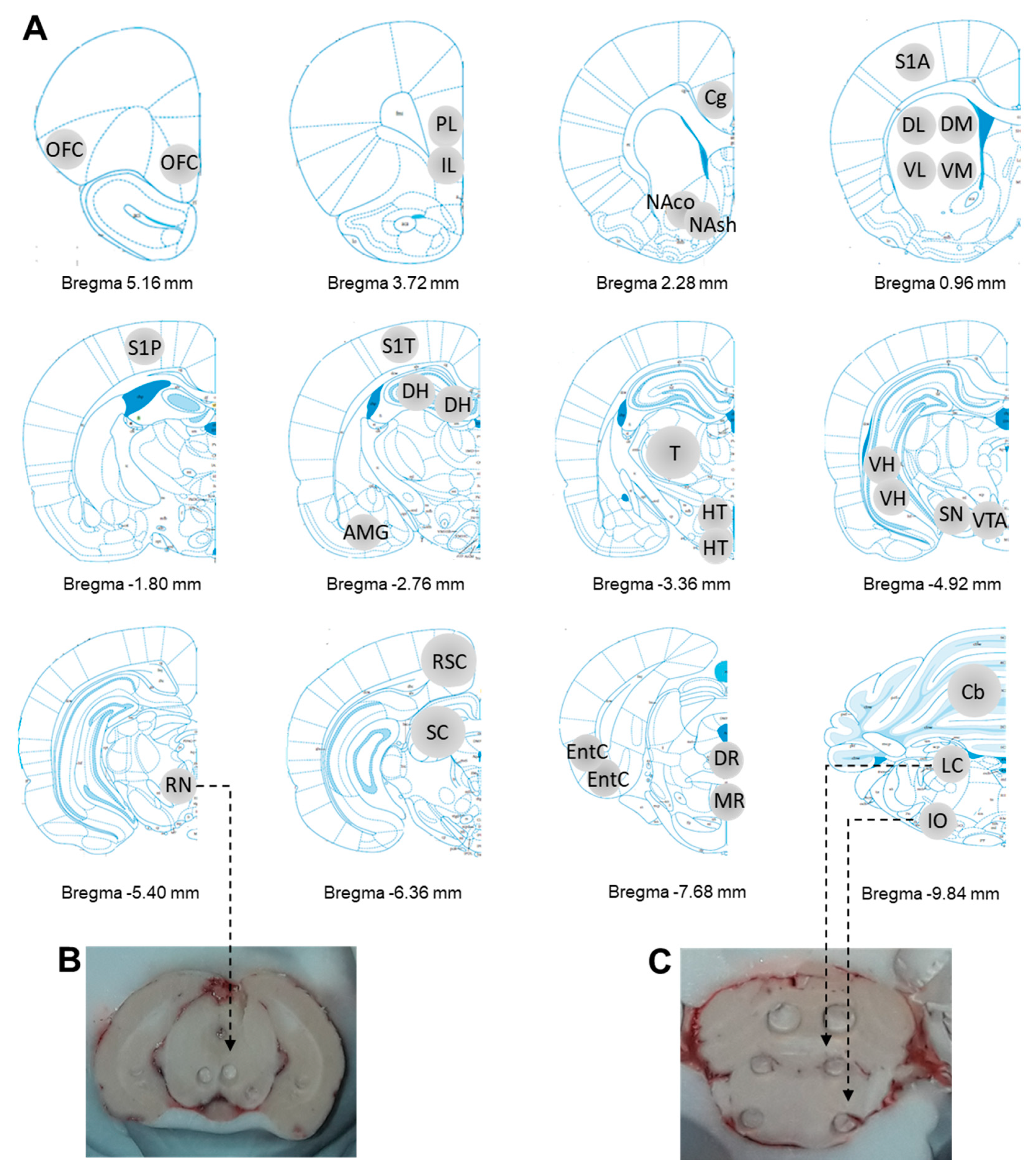
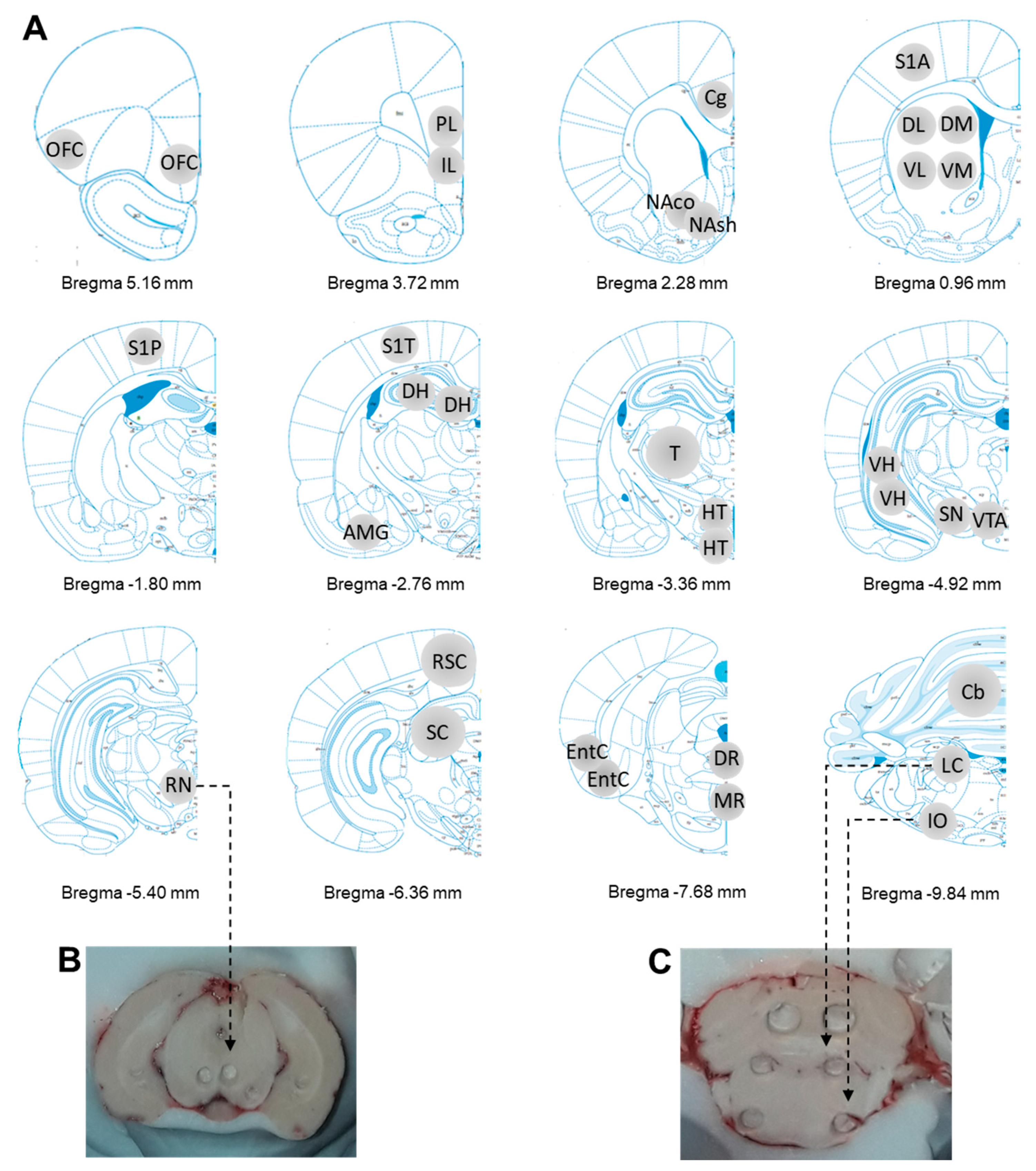
4.4. Tissue Collection of Brain Regions
4.5. Tissue Processing and Neurochemical Analysis
4.6. Chromatographic Analysis
4.7. Statistical Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix B
Appendix C
References
- Hupfeld, K.E.; McGregor, H.R.; Reuter-Lorenz, P.A.; Seidler, R.D. Microgravity effects on the human brain and behavior: Dysfunction and adaptive plasticity. Neurosci. Biobehav. Rev. 2021, 122, 176–189. [Google Scholar] [CrossRef]
- Oluwafemi, F.A.; Abdelbaki, R.; Lai, J.C.; Mora-Almanza, J.G.; Afolayan, E.M. A review of astronaut mental health in manned missions: Potential interventions for cognitive and mental health challenges. Life Sci. Space Res. 2021, 28, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Strangman, G.E.; Sipes, W.; Beven, G. Human cognitive performance in spaceflight and analogue environments. Aviat. Space Environ. Med. 2014, 85, 1033–1048. [Google Scholar] [CrossRef] [PubMed]
- Aston-Jones, G.; Rajkowski, J.; Cohen, J. Role of locus coeruleus in attention and behavioral flexibility. Biol. Psychiatry 1999, 46, 1309–1320. [Google Scholar] [CrossRef]
- Bacque-Cazenave, J.; Bharatiya, R.; Barriere, G.; Delbecque, J.P.; Bouguiyoud, N.; Di Giovanni, G.; Cattaert, D.; De Deurwaerdere, P. Serotonin in Animal Cognition and Behavior. Int. J. Mol. Sci. 2020, 21, 1649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Moal, M.; Simon, H. Mesocorticolimbic dopaminergic network: Functional and regulatory roles. Physiol. Rev. 1991, 71, 155–234. [Google Scholar] [CrossRef]
- Dalley, J.W.; Everitt, B.J.; Robbins, T.W. Impulsivity, compulsivity, and top-down cognitive control. Neuron 2011, 69, 680–694. [Google Scholar] [CrossRef] [Green Version]
- Di Giovanni, G.; Svob Strac, D.; Sole, M.; Unzeta, M.; Tipton, K.F.; Muck-Seler, D.; Bolea, I.; Della Corte, L.; Nikolac Perkovic, M.; Pivac, N.; et al. Monoaminergic and Histaminergic Strategies and Treatments in Brain Diseases. Front. Neurosci. 2016, 10, 541. [Google Scholar] [CrossRef] [Green Version]
- Buoite Stella, A.; Ajčević, M.; Furlanis, G.; Manganotti, P. Neurophysiological adaptations to spaceflight and simulated microgravity. Clin. Neurophysiol. 2021, 132, 498–504. [Google Scholar] [CrossRef]
- Jillings, S.; Van Ombergen, A.; Tomilovskaya, E.; Rumshiskaya, A.; Litvinova, L.; Nosikova, I.; Pechenkova, E.; Rukavishnikov, I.; Kozlovskaya, I.B.; Manko, O.; et al. Macro- and microstructural changes in cosmonauts’ brains after long-duration spaceflight. Sci. Adv. 2020, 6, eaaz9488. [Google Scholar] [CrossRef] [PubMed]
- Pechenkova, E.; Nosikova, I.; Rumshiskaya, A.; Litvinova, L.; Rukavishnikov, I.; Mershina, E.; Sinitsyn, V.; Van Ombergen, A.; Jeurissen, B.; Jillings, S.; et al. Alterations of Functional Brain Connectivity After Long-Duration Spaceflight as Revealed by fMRI. Front. Physiol. 2019, 10, 761. [Google Scholar] [CrossRef] [PubMed]
- Roy-O’Reilly, M.; Mulavara, A.; Williams, T. A review of alterations to the brain during spaceflight and the potential relevance to crew in long-duration space exploration. NPJ Microgravity 2021, 7, 5. [Google Scholar] [CrossRef]
- Culman, J.; Kvetnansky, T.; Serova, L.V.; Tigranjan, R.A.; Macho, L. Serotonin in individual hypothalamic nuclei of rats after space flight on biosatellite Cosmos 1129. Acta Astronaut. 1985, 12, 373–376. [Google Scholar] [CrossRef]
- Kvetnansky, R.; Culman, J.; Serova, L.V.; Tigranjan, R.A.; Torda, T.; Macho, L. Catecholamines and their enzymes in discrete brain areas of rats after space flight on biosatellites Cosmos. Acta Astronaut. 1983, 10, 295–300. [Google Scholar] [CrossRef]
- Miller, J.D.; McMillen, B.A.; McConnaughey, M.M.; Williams, H.L.; Fuller, C.A. Effects of microgravity on brain neurotransmitter receptors. Eur. J. Pharmacol. 1989, 161, 165–171. [Google Scholar] [CrossRef]
- Blanc, S.; Somody, L.; Gharib, A.; Gauquelin, G.; Gharib, C.; Sarda, N. Counteraction of spaceflight-induced changes in the rat central serotonergic system by adrenalectomy and corticosteroid replacement. Neurochem. Int. 1998, 33, 375–382. [Google Scholar] [CrossRef]
- Popova, N.K.; Kulikov, A.V.; Kondaurova, E.M.; Tsybko, A.S.; Kulikova, E.A.; Krasnov, I.B.; Shenkman, B.S.; Bazhenova, E.Y.; Sinyakova, N.A.; Naumenko, V.S. Risk neurogenes for long-term spaceflight: Dopamine and serotonin brain system. Mol. Neurobiol. 2015, 51, 1443–1451. [Google Scholar] [CrossRef]
- Morey-Holton, E.R.; Globus, R.K. Hindlimb unloading rodent model: Technical aspects. J. Appl. Physiol. 2002, 92, 1367–1377. [Google Scholar] [CrossRef] [PubMed]
- Globus, R.K.; Morey-Holton, E. Hindlimb unloading: Rodent analog for microgravity. J. Appl. Physiol. 2016, 120, 1196–1206. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Li, D.; Liu, J.; Diao, L.; Ling, S.; Li, Y.; Gao, J.; Fan, Q.; Sun, W.; Li, Q.; et al. Dammarane Sapogenins Ameliorates Neurocognitive Functional Impairment Induced by Simulated Long-Duration Spaceflight. Front. Pharmacol. 2017, 8, 315. [Google Scholar] [CrossRef] [Green Version]
- Kokhan, V.S.; Lebedeva-Georgievskaya, K.B.; Kudrin, V.S.; Bazyan, A.S.; Maltsev, A.V.; Shtemberg, A.S. An investigation of the single and combined effects of hypogravity and ionizing radiation on brain monoamine metabolism and rats’ behavior. Life Sci. Space Res. 2019, 20, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Kulikova, E.A.; Kulikov, V.A.; Sinyakova, N.A.; Kulikov, A.V.; Popova, N.K. The effect of long-term hindlimb unloading on the expression of risk neurogenes encoding elements of serotonin-, dopaminergic systems and apoptosis; comparison with the effect of actual spaceflight on mouse brain. Neurosci. Lett. 2017, 640, 88–92. [Google Scholar] [CrossRef]
- Feng, L.; Yue, X.F.; Chen, Y.X.; Liu, X.M.; Wang, L.S.; Cao, F.R.; Wang, Q.; Liao, Y.H.; Pan, R.L.; Chang, Q. LC/MS-based metabolomics strategy to assess the amelioration effects of ginseng total saponins on memory deficiency induced by simulated microgravity. J. Pharm. Biomed. Anal. 2016, 125, 329–338. [Google Scholar] [CrossRef]
- Kokhan, V.S.; Matveeva, M.I.; Bazyan, A.S.; Kudrin, V.S.; Mukhametov, A.; Shtemberg, A.S. Combined effects of antiorthostatic suspension and ionizing radiation on the behaviour and neurotransmitters changes in different brain structures of rats. Behav. Brain Res. 2017, 320, 473–483. [Google Scholar] [CrossRef]
- Wang, T.; Chen, H.; Lv, K.; Ji, G.; Zhang, Y.; Wang, Y.; Li, Y.; Qu, L. iTRAQ-based proteomics analysis of hippocampus in spatial memory deficiency rats induced by simulated microgravity. J. Proteom. 2017, 160, 64–73. [Google Scholar] [CrossRef]
- Dellu-Hagedorn, F.; Fitoussi, A.; De Deurwaerdere, P. Correlative analysis of dopaminergic and serotonergic metabolism across the brain to study monoaminergic function and interaction. J. Neurosci. Methods 2017, 280, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Di Giovanni, G.; Bharatiya, R.; Puginier, E.; Ramos, M.; De Deurwaerdère, S.; Chagraoui, A.; De Deurwaerdère, P. Lorcaserin Alters Serotonin and Noradrenaline Tissue Content and Their Interaction with Dopamine in the Rat Brain. Front. Pharmacol. 2020, 11, 962. [Google Scholar] [CrossRef]
- Fitoussi, A.; Dellu-Hagedorn, F.; De Deurwaerdere, P. Monoamines tissue content analysis reveals restricted and site-specific correlations in brain regions involved in cognition. Neuroscience 2013, 255, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Puginier, E.; Bharatiya, R.; Chagraoui, A.; Manem, J.; Cho, Y.H.; Garret, M.; De Deurwaerdere, P. Early neurochemical modifications of monoaminergic systems in the R6/1 mouse model of Huntington’s disease. Neurochem. Int. 2019, 128, 186–195. [Google Scholar] [CrossRef]
- Hupfeld, K.E.; McGregor, H.R.; Koppelmans, V.; Beltran, N.E.; Kofman, I.S.; De Dios, Y.E.; Riascos, R.F.; Reuter-Lorenz, P.A.; Wood, S.J.; Bloomberg, J.J.; et al. Brain and Behavioral Evidence for Reweighting of Vestibular Inputs with Long-Duration Spaceflight. Cereb. Cortex 2021, bhab239. [Google Scholar] [CrossRef] [PubMed]
- Carriot, J.; Jamali, M.; Cullen, K.E. Rapid adaptation of multisensory integration in vestibular pathways. Front. Syst. Neurosci. 2015, 9, 59. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, J.A.; Johnston, C.A. Rapid, concurrent analysis of dopamine, 5-hydroxytryptamine, their precursors and metabolites utilizing high performance liquid chromatography with electrochemical detection: Analysis of brain tissue and cerebrospinal fluid. Life Sci. 1982, 31, 2847–2856. [Google Scholar] [CrossRef]
- Bharatiya, R.; Chagraoui, A.; De Deurwaerdere, S.; Argiolas, A.; Melis, M.R.; Sanna, F.; De Deurwaerdere, P. Chronic Administration of Fipronil Heterogeneously Alters the Neurochemistry of Monoaminergic Systems in the Rat Brain. Int. J. Mol. Sci. 2020, 21, 5711. [Google Scholar] [CrossRef]
- Chowdhury, P.; Akel, N.; Jamshidi-Parsian, A.; Gaddy, D.; Griffin, R.J.; Yadlapalli, J.S.; Dobretsov, M. Degenerative Tissue Responses to Space-like Radiation Doses in a Rodent Model of Simulated Microgravity. Ann. Clin. Lab. Sci. 2016, 46, 190–197. [Google Scholar] [PubMed]
- Chowdhury, P.; Long, A.; Harris, G.; Soulsby, M.E.; Dobretsov, M. Animal model of simulated microgravity: A comparative study of hindlimb unloading via tail versus pelvic suspension. Physiol. Rep. 2013, 1, e00012. [Google Scholar] [CrossRef] [PubMed]
- Morel, J.L.; Dabertrand, F.; Porte, Y.; Prevot, A.; Macrez, N. Up-regulation of ryanodine receptor expression increases the calcium-induced calcium release and spontaneous calcium signals in cerebral arteries from hindlimb unloaded rats. Pflugers Arch. 2014, 466, 1517–1528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, R.T.; Branscum, A.J.; Wong, C.P.; Iwaniec, U.T.; Morey-Holton, E. Studies in microgravity, simulated microgravity and gravity do not support a gravitostat. J. Endocrinol. 2020, 247, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Ulanova, A.; Gritsyna, Y.; Salmov, N.; Lomonosova, Y.; Belova, S.; Nemirovskaya, T.; Shenkman, B.; Vikhlyantsev, I. Effect of L-Arginine on Titin Expression in Rat Soleus Muscle After Hindlimb Unloading. Front. Physiol. 2019, 10, 1221. [Google Scholar] [CrossRef] [Green Version]
- Edgell, H.; Kaufman, S. Effect of hindlimb unloading on salt and water intake and output in male and female rats. Med. Sci. Sports Exerc. 2008, 40, 1249–1254. [Google Scholar] [CrossRef]
- Sullivan, M.J.; Hasser, E.M.; Moffitt, J.A.; Bruno, S.B.; Cunningham, J.T. Rats exhibit aldosterone-dependent sodium appetite during 24 h hindlimb unloading. J. Physiol. 2004, 557, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Macho, L.; Kvetnansky, R.; Vigas, M.; Nemeth, S.; Popova, I.; Tigranian, R.A.; Noskov, V.B.; Serova, L.; Grigoriev, I.A. Effect of space flights on plasma hormone levels in man and in experimental animal. Acta Astronaut. 1991, 23, 117–121. [Google Scholar] [CrossRef]
- Tanaka, Y.; Nakano, J.; Hamaue, Y.; Sekino, Y.; Sakamoto, J.; Kataoka, H.; Okita, M. Hindlimb suspension does not influence mechanical sensitivity, epidermal thickness, and peripheral nerve density in the glabrous skin of the rat hind paw. Physiol. Res. 2013, 62, 119–123. [Google Scholar] [CrossRef]
- Bigbee, A.J.; Grindeland, R.E.; Roy, R.R.; Zhong, H.; Gosselink, K.L.; Arnaud, S.; Edgerton, V.R. Basal and evoked levels of bioassayable growth hormone are altered by hindlimb unloading. J. Appl. Physiol. 2006, 100, 1037–1042. [Google Scholar] [CrossRef] [PubMed]
- Kempadoo, K.A.; Mosharov, E.V.; Choi, S.J.; Sulzer, D.; Kandel, E.R. Dopamine release from the locus coeruleus to the dorsal hippocampus promotes spatial learning and memory. Proc. Natl. Acad. Sci. USA 2016, 113, 14835–14840. [Google Scholar] [CrossRef] [Green Version]
- Devoto, P.; Flore, G.; Saba, P.; Bini, V.; Gessa, G.L. The dopamine beta-hydroxylase inhibitor nepicastat increases dopamine release and potentiates psychostimulant-induced dopamine release in the prefrontal cortex. Addict. Biol. 2014, 19, 612–622. [Google Scholar] [CrossRef] [PubMed]
- Devoto, P.; Flore, G.; Saba, P.; Fa, M.; Gessa, G.L. Co-release of noradrenaline and dopamine in the cerebral cortex elicited by single train and repeated train stimulation of the locus coeruleus. BMC Neurosci. 2005, 6, 31. [Google Scholar] [CrossRef] [Green Version]
- Devoto, P.; Flore, G.; Saba, P.; Scheggi, S.; Mulas, G.; Gambarana, C.; Spiga, S.; Gessa, G.L. Noradrenergic terminals are the primary source of α(2)-adrenoceptor mediated dopamine release in the medial prefrontal cortex. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 90, 97–103. [Google Scholar] [CrossRef]
- De Deurwaerdere, P.; Gaetani, S.; Vaughan, R.A. Old neurochemical markers, new functional directions?: An Editorial for ‘Distinct gradients of various neurotransmitter markers in caudate nucleus and putamen of the human brain’ on page 650. J. Neurochem. 2020, 152, 623–626. [Google Scholar] [CrossRef]
- Salamone, J.D.; Correa, M. The mysterious motivational functions of mesolimbic dopamine. Neuron 2012, 76, 470–485. [Google Scholar] [CrossRef] [Green Version]
- Chagraoui, A.; Whitestone, S.; Baassiri, L.; Manem, J.; Di Giovanni, G.; De Deurwaerdere, P. Neurochemical impact of the 5-HT2C receptor agonist WAY-163909 on monoamine tissue content in the rat brain. Neurochem. Int. 2019, 124, 245–255. [Google Scholar] [CrossRef]
- De Deurwaerdere, P.; Ramos, M.; Bharatiya, R.; Puginier, E.; Chagraoui, A.; Manem, J.; Cuboni, E.; Pierucci, M.; Deidda, G.; Casarrubea, M.; et al. Lorcaserin bidirectionally regulates dopaminergic function site-dependently and disrupts dopamine brain area correlations in rats. Neuropharmacology 2020, 166, 107915. [Google Scholar] [CrossRef] [PubMed]
- Marquis, K.L.; Sabb, A.L.; Logue, S.F.; Brennan, J.A.; Piesla, M.J.; Comery, T.A.; Grauer, S.M.; Ashby, C.R., Jr.; Nguyen, H.Q.; Dawson, L.A.; et al. WAY-163909 [(7bR,10aR)-1,2,3,4,8,9,10,10a-octahydro-7bH-cyclopenta-[b][1,4]diazepino[6,7,1hi ]indole]: A novel 5-hydroxytryptamine 2C receptor-selective agonist with preclinical antipsychotic-like activity. J. Pharmacol. Exp. Ther. 2007, 320, 486–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dellu-Hagedorn, F.; Rivalan, M.; Fitoussi, A.; De Deurwaerdere, P. Inter-individual differences in the impulsive/compulsive dimension: Deciphering related dopaminergic and serotonergic metabolisms at rest. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2018, 373, 20170154. [Google Scholar] [CrossRef]
- Fitoussi, A.; Le Moine, C.; De Deurwaerdere, P.; Laqui, M.; Rivalan, M.; Cador, M.; Dellu-Hagedorn, F. Prefronto-subcortical imbalance characterizes poor decision-making: Neurochemical and neural functional evidences in rats. Brain Struct. Funct. 2015, 220, 3485–3496. [Google Scholar] [CrossRef] [PubMed]
- Duda, N.J.; Moore, K.E. Simultaneous determination of 5-hydroxytryptophan and 3,4-dihydroxyphenylalanine in rat brain by HPLC with electrochemical detection following electrical stimulation of the dorsal raphe nucleus. J. Neurochem. 1985, 44, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Bigelow, R.T.; Agrawal, Y. Vestibular involvement in cognition: Visuospatial ability, attention, executive function, and memory. J. Vestib Res. 2015, 25, 73–89. [Google Scholar] [CrossRef]
- Krupina, N.A.; Khlebnikova, N.N.; Narkevich, V.B.; Naplekova, P.L.; Kudrin, V.S. The Levels of Monoamines and Their Metabolites in the Brain Structures of Rats Subjected to Two- and Three-Month-Long Social Isolation. Bull. Exp. Biol. Med. 2020, 168, 605–609. [Google Scholar] [CrossRef]
- Miachon, S.; Rochet, T.; Mathian, B.; Barbagli, B.; Claustrat, B. Long-term isolation of Wistar rats alters brain monoamine turnover, blood corticosterone, and ACTH. Brain Res. Bull. 1993, 32, 611–614. [Google Scholar] [CrossRef]
- Morey-Holton, E.; Globus, R.K.; Kaplansky, A.; Durnova, G. The hindlimb unloading rat model: Literature overview, technique update and comparison with space flight data. Adv. Space Biol. Med. 2005, 10, 7–40. [Google Scholar] [CrossRef] [PubMed]
- Chagraoui, A.; Boulain, M.; Juvin, L.; Anouar, Y.; Barriere, G.; Deurwaerdere, P. L-DOPA in Parkinson’s Disease: Looking at the “False” Neurotransmitters and Their Meaning. Int. J. Mol. Sci. 2019, 21, 294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motulsky, H.J.; Brown, R.E. Detecting outliers when fitting data with nonlinear regression—A new method based on robust nonlinear regression and the false discovery rate. BMC Bioinform. 2006, 7, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gros, A.; Lavenu, L.; Morel, J.-L.; De Deurwaerdère, P. Simulated Microgravity Subtlety Changes Monoamine Function across the Rat Brain. Int. J. Mol. Sci. 2021, 22, 11759. https://doi.org/10.3390/ijms222111759
Gros A, Lavenu L, Morel J-L, De Deurwaerdère P. Simulated Microgravity Subtlety Changes Monoamine Function across the Rat Brain. International Journal of Molecular Sciences. 2021; 22(21):11759. https://doi.org/10.3390/ijms222111759
Chicago/Turabian StyleGros, Alexandra, Léandre Lavenu, Jean-Luc Morel, and Philippe De Deurwaerdère. 2021. "Simulated Microgravity Subtlety Changes Monoamine Function across the Rat Brain" International Journal of Molecular Sciences 22, no. 21: 11759. https://doi.org/10.3390/ijms222111759
APA StyleGros, A., Lavenu, L., Morel, J.-L., & De Deurwaerdère, P. (2021). Simulated Microgravity Subtlety Changes Monoamine Function across the Rat Brain. International Journal of Molecular Sciences, 22(21), 11759. https://doi.org/10.3390/ijms222111759