Abstract
Most patients with ovarian cancer (OvCA) present peritoneal disseminated disease at the time of diagnosis. During peritoneal metastasis, cancer cells detach from the primary tumor and disseminate through the intraperitoneal fluid. The peritoneal mesothelial cell (PMC) monolayer that lines the abdominal cavity is the first barrier encountered by OvCA cells. Subsequent progression of tumors through the peritoneum leads to the accumulation into the peritoneal stroma of a sizeable population of carcinoma-associated fibroblasts (CAFs), which is mainly originated from a mesothelial-to-mesenchymal transition (MMT) process. A common characteristic of OvCA patients is the intraperitoneal accumulation of ascitic fluid, which is composed of cytokines, chemokines, growth factors, miRNAs, and proteins contained in exosomes, as well as tumor and mesothelial suspended cells, among other components that vary in proportion between patients. Exosomes are small extracellular vesicles that have been shown to mediate peritoneal metastasis by educating a pre-metastatic niche, promoting the accumulation of CAFs via MMT, and inducing tumor growth and chemoresistance. This review summarizes and discusses the pivotal role of exosomes and MMT as mediators of OvCA peritoneal colonization and as emerging diagnostic and therapeutic targets.
1. Introduction
Worldwide, 314,000 new cases of ovarian cancer (OvCA) were diagnosed in 2020, with over 207,000 disease-related deaths. OvCA is the fifth leading cause of cancer-related deaths among women, and the second one amongst gynecologic cancers (following cervical cancer) [1]. When considering industrialized countries, OvCA is the leading cause of death due to gynecological cancer. The World Health Organization (WHO) categorizes OvCA according to the origin of the cancer cell type: coelomic surface epithelial cells, stromal cells, and germ cells [2]. Epithelial OvCA (EOC), also known as ovarian carcinoma, is the most common type, accounting for over 90% of the ovarian malignancies [3]. Histologically, EOC is divided into 5 main subtypes: high-grade serous, low-grade serous, clear cell, endometrioid, and mucinous OvCA; these differ not just in their histologic features but also in their molecular characteristics, natural behavior, prognosis, and, therefore, therapeutic options [4]. High-grade serous ovarian carcinoma (HGSOC) is the most common subtype, accounting for 70–75% of EOCs. Most HGSOC patients experience non-specific symptoms, and, usually at diagnosis, the tumor presents peritoneal extension [5]. The 5-year survival rate is only 29% for these patients with clinically advanced disease [6]. Cytoreductive surgery and platinum-based chemotherapy are the keystone therapy for advanced stage OvCA [7]. However, multidrug-resistant disease is still a major problem for the overall survival of these patients, critically needing new and extending windows of therapeutic opportunities [8].
In contrast to other cancers, which metastasize via hematogenous or lymphatic routes, OvCA mostly disseminates intraperitoneally due to the anatomic location of the primary tumor [9]. In fact, the peritoneum is sometimes the only site of subsequent relapses, and patients invariably die due to complications derived from peritoneal disease [10]. OvCA cells detach from the primary tumor and are transported by the peritoneal fluid, where they spread by colonizing the pelvic and abdominal peritoneum [11]. The membrane that lines the abdominal cavity and all peritoneal organs is formed by a monolayer of peritoneal mesothelial cells (PMCs) with epithelial characteristics that rests on an underlying stroma composed of extracellular matrix (ECM) and connective tissue with few capillaries and resident fibroblasts [12]. The accumulation of a sizeable population of carcinoma-associated fibroblasts (CAFs), which can derive from the PMCs through a mesothelial-to-mesenchymal transition (MMT) process, is an important effect of tumor nesting in the peritoneal membrane [13,14,15]. During MMT, PMCs first dissociate from each other in the monolayer, then lose their apical-basolateral polarity, and reorganize their actin cytoskeleton to progressively acquire migratory and invasive properties [16,17]. The mesothelial cell conversion into CAFs is the result of a complex cellular reprogramming, where diverse pathways can be triggered by multiple promoting stimuli. The profibrotic transforming growth factor-β1 (TGF-β1) is considered as a prototypical inducer of MMT [18]. The receptor-mediated signaling in response to TGF-β1 can trigger the activation of a complex network of intracellular effectors, such as Smad 2/3, integrin-linked kinase (ILK), nuclear factor-𝜅B (NF-𝜅B), extracellular-signal regulated kinases 1/2 (ERKs1/2), phosphatidylinositol 3-kinase (PI3-K)/Akt pathway, c-jun-N terminal kinase (JNK), and TGF- 𝛽-activated kinase-1 (TAK-1) (reviewed in Reference [19]). On the other hand, the accumulation of large volumes of ascitic fluid in patients with OvCA peritoneal carcinomatosis has been linked to alterations of mechanical properties in the peritoneum, which, in turn, regulate the morphological and functional properties of cancer cells [20]. In fact, MMT markers overlapped with TGF-β1-dependent signaling, caveolin-1, and Yes-associated protein (YAP) activation in peritoneal biopsies of OvCA patients, supporting a cooperation between biochemical and biomechanical signal pathways in the triggering of MMT [21]. As a result of MMT, CAFs derived from PMCs synthesize ECM and secrete a variety of cytokines and growth factors that collectively promote tumor implantation, invasion, vascularization, and growth in the peritoneal stroma [13,14,15] (Figure 1).
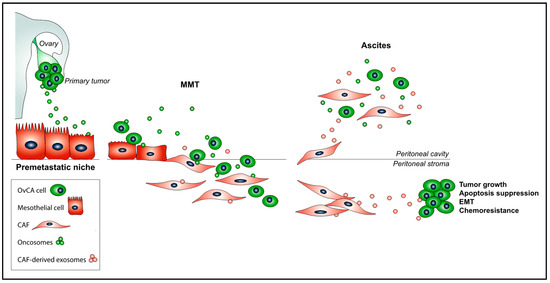
Figure 1.
The promoting role of exosomes impinges on crucial steps of the OvCA peritoneal metastasis process: (i) Primary tumor-derived oncosomes educate a pre-metastatic peritoneal niche; (ii) during MMT, exosomes participate in the processes of adherence of OvCA cells to the mesothelium and co-invasion of OvCA cells and PMC-derived CAFs into the peritoneal stroma; and (iii) exosomes derived from CAFs induce EMT in tumor cells and suppress cancer cell apoptosis, as well as confer tumor growth and chemoresistance. Finally, OvCA cells, PMC-derived CAFs, and their, respectively, secreted exosomes are accumulated in the intraperitoneal ascitic fluid.
OvCA is often accompanied by intraperitoneal accumulation of ascitic fluid, which is associated with poor prognosis [11]. Malignant ascites is the result of leakiness of microvasculature, as well as obstruction of lymphatic vessels, and is frequently a sign of peritoneal affectation [22,23,24]. Within this intraperitoneal fluidic microenvironment, tumor cells, mesothelial-derived CAFs, and infiltrating leukocytes produce a multitude of factors, including but not limited to cytokines, chemokines, and growth factors [15,23,25,26,27,28]. These autocrine and paracrine soluble molecules form complex signaling networks that govern, in part, tumor-peritoneum interactions [14]. However, large quantities of both, tumor-produced exosomes (termed “oncosomes”) and CAF-secreted exosomes, have been found in malignant ascites from OvCA patients [29]. In fact, more and more studies point to exosomes as principal mediators of tumor-stroma crosstalk and suggest that these small extracellular vesicles play an important role in favoring peritoneal metastasis, through facilitating cell adhesion, invasion, angiogenesis, proliferation, immune evasion, and chemoresistance in OvCA (reviewed in Reference [30]).
Exosomes are a subtype of 30–150-nm-sized extracellular vesicles with endocytic origin that are released to the extracellular space upon fusion of intracellular multivesicular bodies with the plasma membrane [31]. Although the content of exosomes shows specificity to the cell of origin and depends, as well, on the functional state and regulated sorting mechanisms of the cell, common components have been described for exosomes released by different cells (reviewed in Reference [32]). Typical exosomal proteins include those related to their biogenesis, such as ESCORT, ALIX, and TSG101, but also membrane proteins, such as adhesion molecules, integrins, transport and fusion proteins, heat shock proteins, cytoskeleton proteins, and the tetraspanins CD9, CD63, and CD81, which are often used as exosome detection markers [31,33,34]. In addition, exosomes are enriched in lipids, which mainly derive from the plasma membrane of the cell of origin, including cholesterol, sphingomyelin, ceramide, and phosphatidylserine (reviewed in Reference [35]). Exosomes also transport functional RNA molecules, among which are mRNAs and non-coding RNAs, such as microRNAs (miRNAs) and long non-codingRNAs (lncRNAs) [36,37]. Moreover, single and double-stranded DNA [38], as well as mitochondrial DNA, are also contained in exosomes [39,40].
Exosomes are formed via the endocytic pathway. The first step in exosome biogenesis is the formation of early endosomes through the fusion of endocytic vesicles in the cytosol. Early endosome proteins can return to the plasma membrane through recycling vesicles or, alternatively, they can mature into a specialized form of late endosomes [41], termed multivesicular bodies (MVBs), that contain intraluminal vesicles formed by the inward budding from the membrane into the lumen of MVBs [42]. The biogenesis mechanism of exosomes affects their cargo, which, in turn, determines how exosomes communicate with target cells, as well as the processes that will be regulated [43]. Three routes of exosome biogenesis have been proposed: the endosomal sorting complex for transport (ESCRT)-dependent pathway [44,45,46]; the lipid-mediated endocytosis [47,48]; and the tetraspanin-mediated mechanism [49,50,51]. Exosomes are ultimately released to the extracellular environment by the fusion of MVBs with the plasma membrane, where they can interact locally with other cells or be transported through the blood or lymph to distant sites. It has been also shown that some exosomes remain fused to the plasma membrane of the cell of origin, where they could work as signaling platforms [52,53,54].
Exosomes are important vehicles of intercellular communication through the transfer of their cargo of proteins, nucleic acids, and lipids between donor and recipient cells [55,56]. Different possibilities for the interaction between exosomes and their target cells have been proposed, including: binding of exosomes to the cell surface through ligand-receptor pairs of specific adhesion molecules; direct fusion between exosomal and cellular membranes; and internalization of exosomes into endocytic compartments through receptor-mediated endocytosis, such as the caveolin- [57] or clathrin-dependent pathways [57,58,59], through a mechanism based on lipid rafts [60], or by phagocytosis [61] or micropinocitosis [62]. The interaction with exosomes can induce direct stimulation of target cells, the transfer of membrane receptors, or the intracellular reception and integration of molecular information carried by exosomes in recipient cells.
This review focuses on providing novel insights to understand how exosomes participate in OvCA progression through the peritoneum. The new knowledge related to exosomes as potential biomarkers and therapeutic tools for peritoneal metastasis in OvCA will be also briefly discussed.
2. The Role of Exosomes in Ovarian Cancer Peritoneal Metastasis
Tumors originating in the abdominal cavity, such as ovarian, endometrial, pancreatic, gastric, and colorectal cancers, frequently colonize the peritoneum [9]. Interestingly, exosome-related peritoneal metastasis mechanisms have been described for these types of cancer [30,63,64]. Exosomes can be found in almost all biological fluids, including serum, saliva, urine, amniotic fluid, breast milk, and seminal fluid [65,66,67,68,69]. In recent years, the detection of exosomes in serum samples of oncological patients has raised great interest, since they have been found to play crucial roles in tumorigenesis, progression, and metastasis in different cancers that mainly disseminate through the hematogenous or lymphatic routes [70,71,72,73]. However, in the context of peritoneal metastasis, the abundance of exosomes in intra-abdominal ascitic fluid acquires a special relevance. On this note, exosomes show up to 3–4-fold increased concentrations in the malignant ascites of ovarian carcinoma patients as compared to the peritoneal fluid of non-oncological individuals [74,75]. In OvCA, exosomes exert important roles, acting directly on cancer cells, facilitating their shedding from the primary tumor, promoting their survival in the peritoneal fluid, and favoring their attachment to the PMC monolayer and subsequent invasion into the underlying peritoneal stroma [76]. Additionally, exosomes participate in the process of peritoneal metastasis by mediating complex networks of intercellular communication between OvCA cells and resident cells of the peritoneal microenvironment. In this regard, exosomes participate in the formation of a peritoneal pre-metastatic niche susceptible of being subsequently metastasized through different mechanisms, including the conversion of PMCs into CAFs via MMT, inducing immunosuppression, and promoting tumor vascularization [30] (Figure 1). On the other hand, an increasing number of studies point to exosomes as promising tools to improve OvCA outcome by reducing rates of peritoneal metastatic lesions, by facilitating early diagnosis and by interfering with tumor chemoresistance mechanisms (reviewed in References [77,78]).
3. Oncosomes and Their Recipient Target Cells in the Peritoneum
3.1. Peritoneal Mesothelial Cells
At the initial steps of peritoneal metastasis, OvCA cells directly encounter the monolayer formed by PMCs. Until recent years, it was believed that PMCs only acted as a passive mechanical barrier, avoiding tumor cell adhesion and invasion in the peritoneum and, as a consequence, preventing the formation of secondary tumor nodules into the submesothelial peritoneal stroma [79]. However, more recently, it has been reported that PMCs exert an active role in establishing a pre-metastatic niche required for the subsequent colonization of the peritoneum [80]. As in any distant metastatic process, peritoneal colonization requires the previous education of a pre-metastatic niche, a peritoneal microenvironment that favors the subsequent OvCA cell invasion through the submesothelium. PMCs are considered the principal recipient target cells for a wide range of molecules packed in oncosomes, which are initially released to the peritoneal cavity from the primary tumor site. On this note, Yokoi et al. proposed a mechanism of apoptotic PMC death via OvCA-produced extracellular vesicles carrying MMP1 mRNA [74]. Undoubtedly, the destruction of the peritoneal mesothelium barrier facilitates the establishment of metastatic implants into the peritoneal stroma. Nevertheless, in the context of peritoneal metastasis, PMCs can be converted into CAFs through an MMT process [13,14,15]. In this regard, an increasing number of reports point to oncosomes as key mediators of peritoneal metastasis through the mesenchymal reprograming of PMCs [81,82]. In fact, Wei et al. revealed the expression of specific fibroblast markers, including fibroblast activation protein (FAP) and alpha-smooth muscle actin (α-SMA), in PMCs upon in vitro and in vivo treatments with malignant ascites-derived exosomes [81].
The MMT is a consequence of a sequential process [14], and oncosome-containing proteins have been noticed to play an important role in many MMT-related steps. On this note, TGF-β1 has been found to be overexpressed in malignant ascites-derived exosomes, therefore being proposed as the principal inducer of mesenchymal conversion in the peritoneum [81]. On the other hand, the molecule CD44, a cell surface glycoprotein, has been found to be enriched in EOC-derived exosomes [82]. Interestingly, CD44 has an important role in many cellular functions, such as cell-cell interaction, adhesion, migration, and metastasis [83,84,85,86,87]. CD44 mediates tumor cell adhesion to the mesothelial monolayer through its interaction with hyaluronic acid, and, indeed, this interaction partly mediates the adhesion of OvCA cells to the peritoneal membrane [86]. In OvCA peritoneal metastasis, CD44 is transferred in oncosomes to PMCs. As a consequence, PMCs are induced to secrete MMP9, promoting ECM remodeling, clearing the mesothelial barrier, and participating in OvCA cell invasion through the peritoneal membrane [82].
In addition to proteins, ascites-isolated exosomes contain a unique miRNA signature specific to OvCA cells [88,89,90]. In this regard, it has been described that miR-99a-5p is up-regulated in oncosomes and transferred to PMCs, where, in turn, it up-regulates the expression of ECM components, such as fibronectin and vitronectin [91]. Interestingly, these two matrix proteins have been involved in OvCA cell adhesion to, and invasion through, the mesothelial monolayer that lines the peritoneal cavity [92,93]. lncRNAs have also been found to take part in OvCA progression. For example, the lncRNA SPOCD1-AS, embedded in OvCA-secreted extracellular vesicles, is transported to recipient PMCs, inducing MMT-related changes via interacting with G3BP1 protein and enhancing peritoneal colonization [94]. Besides miRNAs and lncRNAs, the exosomal circular RNA (circRNA) circPUM1 has been recently reported to participate in the peritoneal progression of OvCA. CircPUM1 can exert its tumorigenic effects by acting directly on cancer cells, but it can also be released in oncosomes and transferred to PMCs, where it up-regulates both MMP2 and NF-κB expression [95]. Zong et al. described how the circRNA circWHSC1 induces EOC metastasis by acting on the peritoneal mesothelium. CircWHSC1 is secreted by OvCA cells contained in exosomes and is taken up by PMCs, inducing up-regulation of MUC1 expression and MMT, which favors peritoneal tumor implantation [96] (Figure 1).
3.2. Other Oncosome-Target Cells in the Peritoneal Stroma
It is known that a crosstalk exists between tumor cells and the tumoral niche that is crucial to the development of the OvCA disease. Moreover, tumor cells can produce and release different biomolecules to their microenvironment with relevant effects on the local stroma, causing its remodeling and transformation into a pre-metastatic niche that favors ovarian tumor growth and metastasis [97,98]. Some of these biomolecules can be transferred through oncosomes from producing tumor cells to a variety of target cells, including not only the PMCs as described above but also endothelial, immune and other tumor cells, regulating gene expression and altering the phenotype and functions in these recipient cells [99,100,101,102] (Figure 2).
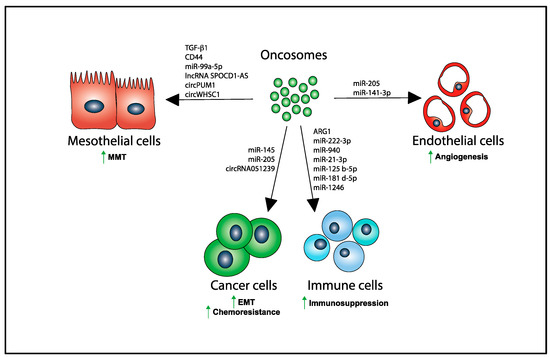
Figure 2.
Several biomolecules are transferred via oncosomes from producing OvCA cells to their recipient target cells in the peritoneum inducing MMT in PMCs, EMT and chemoresistance in tumor cells, immunosuppression in immune cells, and angiogenesis in endothelial cells. Upward green arrows denote increase in the indicated biological process.
3.2.1. Effects of Oncosomes on Immune Cells
OvCA oncosomes can exert both direct and indirect effects on innate and adaptive immune cells, promoting tumor-induced immunosuppression and evasion from immunosurveillance (reviewed in References [30,76,78]). For instance, it has been observed that they can inhibit T cell activation through their receptor (TCR) by means of different proteins expressed on their surface, including the ganglioside GD3 [103]. They can also promote apoptosis of different immune cells, including dendritic cells (DCs), peripheral blood lymphocytes, and hematopoietic stem cells, using the Fas ligand (FasL) expressed on their surface, leading to immunosuppression [104,105,106]. Another mechanism for immune suppression is the presence of arginase-1 (ARG1); these ARG1-expressing exosomes are taken up by DCs which inhibit the proliferation of CD4+ and CD8+ T-cells [107]. OvCA exosomes could also potentially down-regulate the activity of NK cells through NKG2D, similar to what has been found for other tumors [108].
Macrophages are another immune cell that plays an important role in the establishment of the pre-tumoral niche, and, in fact, they are the major type of immune cell present in the tumor environment. They can display either a pro-inflammatory (M1) or an anti-inflammatory phenotype (M2), and many studies have shown that tumor-associated macrophages are typically polarized toward an M2-like phenotype and play a crucial role in the progression of the tumor [109,110]. These cells can produce and release several molecules, such as TGFβ-1, VEGFA, IL-4, IL-5, or IL-6, that suppress the adaptive immune response and promote tumor cell survival and proliferation, invasion, and metastasis. Some studies have demonstrated that several miRNAs contained in oncosomes, such as miR-222-3p, miR-940, miR-21-3p, miR-125 b-5p, miR-181 d-5p, or miR-1246, induce the polarization toward M2 macrophages [109,110,111].
3.2.2. Effects of Oncosomes on Ovarian Cancer Cells
Exosomes released by some OvCA cells can be taken up by other cancer cells, provoking changes in their phenotypic and functional properties. In this regard, the miRNA content in OvCA oncosomes has been observed to promote changes, known as epithelial-to-mesenchymal transition (EMT), in recipient tumor cells. For example, miR-145 seems to be down-regulated in OvCA oncosomes, accordingly, resulting in reduced suppression of its direct target gene CTGF (connective tissue growth factor), which is involved in tumor cell migration and adhesion [112]. A recent study shows that the amount of circRNA051239, another non-coding RNA, is increased in oncosomes and can regulate the expression of many genes in target tumor cells. This seems to have consequences in the progression of the disease by promoting cell proliferation and migration [113].
3.2.3. Effects of Oncosomes on Endothelial Cells
The biomolecular cargo in OvCA oncosomes can also facilitate tumor progression by targeting, directly or indirectly, angiogenic factors. These factors can be encapsulated in exosomes and transported to endothelial cells, inducing angiogenesis and promoting metastasis [114,115]. miR-205 is one of the molecules that has been reported to be involved in the development of metastasis through the induction of tumor angiogenesis. This miRNA, which is up-regulated in OvCA patients, can promote angiogenesis by being transported to the recipient endothelial cells. On the other hand, it can also induce angiogenesis through the PTEN-AKT pathway [114]. This latter effect of miRNA-205 is thought to be mediated by targeting the VEGF pathway. Furthermore, the overexpression of miR-205 also seems to be able to promote EMT in OvCA [90]. A recent study has shown that oncosomes are also capable to up-regulate the pro-angiogenic factor VEGFA pathway in endothelial cells, driving their proliferation and migration [116].
The RNA and protein cargo in oncosomes modulate the angiogenesis process in vitro and in vivo [100]. For example, a recent study has shown that miR-141-3p in oncosomes can promote the angiogenesis process in vitro by the modulation of SOCS5, which is considered a negative regulator of the JAK-STAT and the VEGFR-2 signaling [117].
4. CAFs Generated via MMT Produce Exosomes That Impact on Recipient Target Ovarian Cancer Cells
While most studies are focused on oncosomes, little is known about exosomes released by cells of the surrounding tumor microenvironment and their effects in tumor progression at secondary metastatic sites.
Solid tumors are complex and unstructured organs that, in addition to cancer cells, also contain stromal cell types. It is known that CAFs represent an important population in the tumor microenvironment and participate in providing a suitable ECM and blood vessel formation to support tumor cell survival at secondary metastatic sites [118]. Furthermore, in the last few years, a number of studies have provided critical evidence regarding the significance of exosome-mediated intercellular crosstalk between CAFs and cancer cells for tumor progression [119]. For instance, in OvCA, it has been reported that CAF-derived exosomal miR-98-5p increases tumor cell proliferation and cell cycle entry, as well as confer cisplatin resistance, by targeting CDKN1A [120].
The origin of peritoneal CAFs associated with OvCA metastasis has been the subject of intense debate. However, our group demonstrated, for the first time, that an important proportion of CAFs, in peritoneal OvCA tumor implants, derives from PMCs as a consequence of an MMT process [13,14,15]. Regardless their origin, peritoneal CAFs can produce and secrete exosomes containing molecules that can be transferred, in turn, to tumor cells. On this note, it has been observed that omental CAF-derived exosomes are enriched in TGF-β1, which can be transferred to OvCA cells, triggering the acquisition of a more aggressive tumoral phenotype through undergoing EMT-related changes [121]. Interestingly, TGF-β1 has been found to be significantly up-regulated in MMT-derived CAFs isolated from the ascitic fluid of OvCA patients as compared to normal PMCs, suggesting that targeting exosomes secreted by PMCs undergoing MMT could be a potential mechanism to be interfered in the treatment of peritoneal metastasis [15]. On the other hand, Au Yeung et al. showed that miR21, a very recently identified cargo biomolecule in CAF-derived exosomes [119], is transferred from neighboring stromal cells in the omental tumor microenvironment (including CAFs and cancer-associated adipocytes) to cancer cells, where it suppresses OvCA apoptosis and confers chemoresistance by binding to its direct target APAF1 [122]. Accordingly, miR-21 has been identified as one of the most abundant miRNAs in PMCs, exhibiting mesenchymal changes upon TGF-β1 stimulation, thus providing a novel approach in the context of peritoneal carcinomatosis [123] (Figure 1).
5. Molecules That Mediate Specific Interactions and Uptake of Exosomes by Recipient Target Cells
As indicated above in Section 3 and Section 4, in order to act as efficient vehicles of intercellular communication, oncosomes and exosomes produced by mesothelial cells must be able to deliver their cargo of biomolecules to a variety of different target cells. The molecules that dictate these specific interactions and the subsequent uptake of exosomes by recipient cells are only beginning to be identified, and they have been reviewed by Mulcahy et al. and French et al. [124,125]. The involvement of specific molecules in these processes is frequently inferred from the use of antibodies that block exosome uptake. In this regard, different members of the integrin and immunoglobulin families of adhesion receptors were amongst the proteins consistently found to mediate these interactions, together with proteoglycans, such as CD44, and extracellular matrix proteins, such as fibronectin. Additionally, members of the tetraspanin family, such as CD9 and CD81, were also found to participate as regulators of exosome-recipient cell interactions. Our group has recently reported that the interaction of cellular integrin α5β1 with exosomal ADAM17 mediates the binding and uptake of colorectal carcinoma exosomes by recipient PMCs and cancer cells, which may bear relevance in the process of peritoneal dissemination [126]. Furthermore, exosomal tetraspanin CD9 was found to negatively regulate these interactions between cellular integrin α5β1 and its exosomal ligand ADAM17.
The question of whether cancer cells can release different subpopulations of exosomes with unique biological functions which could be targeting distinct recipient cells has not been fully resolved (reviewed in Reference [73]). However, some evidence would support that notion; for instance, neuroblastoma cells secrete different exosome populations, which differ in their cargoes and target different cells, such as neurons or glial cells [127]. Furthermore, Hoshino et al. reported, in their seminal paper, that exosomes produced by a variety of cancer cells display different integrin cargoes, which direct their selective uptake by distinct target cells, thus dictating the metastatic organotropism [52]. These findings are of upmost relevance for understanding the specific roles played by different subpopulations of cancer-produced exosomes, and, clearly, more research is still needed to further advance knowledge on this topic.
6. Exosomes in the Diagnosis, Prognosis, and Therapy of Ovarian Cancer Peritoneal Metastasis
The majority of women with EOC present peritoneal metastasis at the time of diagnosis. The metastatic process, however, starts long before secondary cancer implants are detected. Exosomes derived from the primary tumor prepare a cancer-favorable microenvironment in the pre-metastatic niche before the target organ is already colonized [128]. On this note, OvCA-secreted exosomes from the primary site could represent a unique opportunity to assist patients in the early detection of peritoneal dissemination. As an example, oncosomes isolated from OvCA patients carried TGF-β1, which distinguished OvCA patients from those with benign lesions [129]. Interestingly, despite their elevated TGF-β1 production, this factor has limited effects in OvCA cells, being that its contribution to peritoneal metastasis is mainly mediated through activation of Smad3-dependent TGF-β1-signaling in surrounding PMC-derived CAFs [15,130]. Moreover, high levels of oncosomal CA125 and claudin-4 have been detected in OvCA patients, significantly contributing to improved diagnosis [131]. Im et al. developed a nano-plasmonic sensor to identify oncosomes expressing CD24 and EpCAM in malignant ascites samples from OvCA patients, highlighting their potential for diagnostics [29]. Alternatively, a large battery of miRNAs has been described to be highly dysregulated in exosomes of patients with EOC [88,89,132,133,134]. Therefore, the oncosomal miRNA profiling could also be highly informative for the early diagnosis of OvCA peritoneal metastasis.
On the other hand, OvCA malignant ascites-derived exosomes display a cargo of tumor progression related proteins, such as L1CAM, CD24, ADAM10, and EMMPRIN, which have been found to correlate with worse prognosis [135]. After completion of first-line treatment, chemoresistance frequently develops, and recurrent peritoneal malignant disease is subsequently observed; this development of chemoresistance by tumor cells is a major hurdle in the treatment of OvCA. In this regard, oncosomal cargoes could also have the potential to serve as prognostic biomarkers of chemoresistance in patients with peritoneal carcinomatosis as exosomes have been proposed to play a pivotal role in the acquisition of chemotherapy resistance by OvCA cells. They have been found to mediate the acquisition of the chemoresistant phenotype in OvCA cells through multiple mechanisms, including inhibition of apoptosis, enhanced DNA repair, increased drug effluxion through the transfer of multidrug resistance (MDR) transporters, and by reducing the cellular concentration of chemotherapeutic drugs in tumoral donor cells through their expulsion in these vesicles (reviewed in References [78,136,137]. Several proteins have been found to be overexpressed in exosomes produced by chemoresistant OvCA cells, including Annexin A3 [138,139], cisplatin export transporters (MRP2 and ATP7A/B) [140], DNA methyltransferase 1 (DNMT1) [141], EpCAM [142,143], and MAGE3/6 [129]. In addition, acquired SMAD4 mutations enhance the chemoresistance profile of epithelial OvCA cells, representing a mechanism in which exchange of tumor-derived exosomes perpetuates an EMT phenotype, leading to the development of subpopulations of platinum-refractory tumor cells [144]. In addition, some miRNas have also been found to be overexpressed in OvCA tumor chemoresistance, including miR-21-3p [145], miR21 [122], miR-433 [146], miR-1246, and miR-223 [147], which could bear potential diagnostic and prognostic value for patients [145].
The singular condition of the peritoneal cavity microenvironment not only affects the chemoresistant oncosome profile but also the amount of CAF-secreted exosomes, and their cargo could be particularly relevant from a prognostic standpoint [29]. Little is known about the value of exosomes produced by MMT-derived CAFs to predict peritoneal tumor progression or therapeutic response to chemotherapy in patients with advanced OvCA. Intriguingly, Rafii et al. isolated from ascites of OvCA patients a particular type of cells with common characteristics to MCs, referred to as “Hospicells”. These cells represent a differentiated stromal subset of mesenchymal stem cells with expression of multi-drug resistance proteins. Hospicells preferentially interact with EOC cells, inducing their chemoresistance to platin and taxanes through the capture of stromal cell membrane patches by a process termed onco-trogocytosis [148]. This work led us to speculate that PMC-derived CAFs could transfer information to OvCA cells by an exosome-dependent mechanism in order to confer them a chemo-resistant phenotype. Accordingly, miR-21 is transferred in exosomes from peritoneal CAFs to cancer cells, where it suppresses OvCA apoptosis and confers chemoresistance, as it is mentioned before [122]. On this note, miR-21, known for its pro-oncogenic and pro-fibrotic activities, is highly present in OvCA-associated acites [149]. Effusion fluid-derived exosomes containing miR-21 have been associated to TGF-β-related pathways, extracellular matrix-receptor interaction, mesothelial clearance and worse prognosis value in metastatic OvCA [132]. Therefore, the detection of exosomes containing miR-21 could improve prognosis in OvCA peritoneal metastasis.
Exosomes are continuously being investigated for their applications in the therapeutic field, and, increasingly, novel options for exploiting exosomes in the treatment of OvCA peritoneal metastases are emerging [77,150]. For example, interfering with exosomal secretion or uptake mechanisms could represent an important target for therapeutical intervention. On this note, drug-resistant OvCA cells abnormally sort some lysosomal proteins showing enhanced exosomal export of cisplatin, thus this being a characteristic to be explored as a target in advanced OvCA patients [140]. Samuel et al. described that cisplatin treatment of OvCA cells led to the release of extracellular vesicles that could induce invasion and increased resistance via p38 and JNK signaling when taken up by neighboring unstressed tumor cell populations. In addition, extracellular vesicle uptake inhibitors prevented this extracellular vesicle-mediated crosstalk and, thus, sensitized cancer cells to the effects of chemotherapy [151,152]. Alternatively, removal of exosomes from malignant ascites could also contribute to improve OvCA clinical outcome. De la Fuente et al. employed exosomes purified from the ascitic fluid of OvCA patients in a murine model of peritoneal metastasis as traps to interfere with tumor cell peritoneal attachment [153]. On the other hand, interfering with the exosome-mediated MMT process could be highly advantageous in the context of peritoneal metastasis. On this note, hepatocyte growth factor (HGF) has been validated as an exosome-contained protein of interest in HGSOC patients [154]. In addition, OvCA-produced HGF is known to transform the peritoneum via MMT into a more suitable niche for subsequent tumor invasion [14,155,156]. Interestingly, siRNA against HGF packed in exosomes has been described to be transported into tumor cells metastasizing peritoneum, suppressing proliferation and migration [157]. These data lead us to speculate that exosomes delivering MMT-blocking drugs could have potential therapeutic value in OvCA peritoneal metastasis.
7. Conclusions
Development of peritoneal carcinomatosis is a frequent outcome in OvCA patients, which still today represents mostly a deadly incurable stage of this disease, despite the improved surgical and chemotherapeutic approaches resulting in increased progression-free disease intervals achieved in these patients over the past 30 years. A better understanding of the precise roles played by peritoneal exosomes released by tumor and stromal cells and of the mechanisms by which these extracellular vesicles deliver their biomolecular cargoes and alter the properties of recipient target cells is urgently needed. Furthermore, exosomes in OvCA are increasingly becoming recognized as key players in the conversion of PMCs into tumor-promoting CAFs through an MMT reprogramming process, which has important implications in the pathogenesis of the disease. This new knowledge on exosomes in OvCA will undoubtedly lead to the development of novel disease biomarkers, leading to earlier diagnostic procedures, and will open novel and more effective therapeutic avenues, which will collectively improve the clinical management of these women and the survival rates of this disease.
Author Contributions
Conceptualization, L.P.-A., B.C., C.C. and P.S.; writing—original draft preparation, L.P.-A., B.C., C.C. and P.S.; writing—review and editing, L.P.-A., B.C., R.S.d.l.C., L.G.-C., M.L.-C., P.S. and C.C.; visualization, P.S., B.C. and L.P.-A.; supervision, C.C. and P.S.; funding acquisition, M.L.-C. and C.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Spanish Ministry of Science and Innovation/Fondo Europeo de Desarrollo Regional (MICINN/FEDER), grant numbers PID2019-110132RB-I00/AEI/10.13039/501100011033 (to M.L.-C.) and SAF2016-77096-R (to C.C.), and by Consejo Superior de Investigaciones Científicas (CSIC), grant number 2020AEP018 (to C.C.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.T.; Tavassoli, F.A.; Prat, J. Tumors of the ovary and peritoneum. In World Health Organization Classification of Tumours: Pathology and Genetics of Tumours of the Breast and Female Genital Organs, 3rd ed.; IARC Press: Lyon, France, 2003; Volume 4, p. 117. [Google Scholar]
- Heintz, A.P.; Odicino, F.; Maisonneuve, P.; Beller, U.; Benedet, J.L.; Creasman, W.T.; Ngan, H.Y.; Sideri, M.; Pecorelli, S. Carcinoma of the ovary. J. Epidemiol. Biostat. 2001, 6, 107–138. [Google Scholar] [PubMed]
- Prat, J. Ovarian carcinomas: Five distinct diseases with different origins, genetic alterations, and clinicopathological features. Virchows Arch. 2012, 460, 237–249. [Google Scholar] [CrossRef]
- Kobel, M.; Kalloger, S.E.; Huntsman, D.G.; Santos, J.L.; Swenerton, K.D.; Seidman, J.D.; Gilks, C.B. Cheryl Brown Ovarian Cancer Outcomes Unit of the British Columbia Cancer Agency, V.B.C. Differences in tumor type in low-stage versus high-stage ovarian carcinomas. Int. J. Gynecol. Pathol. 2010, 29, 203–211. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenhauer, E.L.; Abu-Rustum, N.R.; Sonoda, Y.; Aghajanian, C.; Barakat, R.R.; Chi, D.S. The effect of maximal surgical cytoreduction on sensitivity to platinum-taxane chemotherapy and subsequent survival in patients with advanced ovarian cancer. Gynecol. Oncol. 2008, 108, 276–281. [Google Scholar] [CrossRef]
- McGee, J.; Bookman, M.; Harter, P.; Marth, C.; McNeish, I.; Moore, K.N.; Poveda, A.; Hilpert, F.; Hasegawa, K.; Bacon, M.; et al. Fifth Ovarian Cancer Consensus Conference: Individualized therapy and patient factors. Ann. Oncol. 2017, 28, 702–710. [Google Scholar] [CrossRef]
- Desai, J.P.; Moustarah, F. Peritoneal Metastasis. In StatPearls; StatPearls: Treasure Island, FL, USA, 2021. [Google Scholar]
- Kurnit, K.C.; Fleming, G.F.; Lengyel, E. Updates and New Options in Advanced Epithelial Ovarian Cancer Treatment. Obs. Gynecol. 2021, 137, 108–121. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.S.; Agarwal, R.; Kaye, S.B. Mechanisms of transcoelomic metastasis in ovarian cancer. Lancet Oncol. 2006, 7, 925–934. [Google Scholar] [CrossRef]
- Di Paolo, N.; Sacchi, G. Atlas of peritoneal histology. Perit. Dial. Int. 2000, 20 (Suppl. 3), S5–S96. [Google Scholar] [PubMed]
- Sandoval, P.; Jimenez-Heffernan, J.A.; Rynne-Vidal, A.; Perez-Lozano, M.L.; Gilsanz, A.; Ruiz-Carpio, V.; Reyes, R.; Garcia-Bordas, J.; Stamatakis, K.; Dotor, J.; et al. Carcinoma-associated fibroblasts derive from mesothelial cells via mesothelial-to-mesenchymal transition in peritoneal metastasis. J. Pathol. 2013, 231, 517–531. [Google Scholar] [CrossRef] [Green Version]
- Rynne-Vidal, A.; Jimenez-Heffernan, J.A.; Fernandez-Chacon, C.; Lopez-Cabrera, M.; Sandoval, P. The Mesothelial Origin of Carcinoma Associated-Fibroblasts in Peritoneal Metastasis. Cancers 2015, 7, 1994–2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rynne-Vidal, A.; Au-Yeung, C.L.; Jimenez-Heffernan, J.A.; Perez-Lozano, M.L.; Cremades-Jimeno, L.; Barcena, C.; Cristobal-Garcia, I.; Fernandez-Chacon, C.; Yeung, T.L.; Mok, S.C.; et al. Mesothelial-to-mesenchymal transition as a possible therapeutic target in peritoneal metastasis of ovarian cancer. J. Pathol. 2017, 242, 140–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanez-Mo, M.; Lara-Pezzi, E.; Selgas, R.; Ramirez-Huesca, M.; Dominguez-Jimenez, C.; Jimenez-Heffernan, J.A.; Aguilera, A.; Sanchez-Tomero, J.A.; Bajo, M.A.; Alvarez, V.; et al. Peritoneal dialysis and epithelial-to-mesenchymal transition of mesothelial cells. N. Engl. J. Med. 2003, 348, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Carpio, V.; Sandoval, P.; Aguilera, A.; Albar-Vizcaino, P.; Perez-Lozano, M.L.; Gonzalez-Mateo, G.T.; Acuna-Ruiz, A.; Garcia-Cantalejo, J.; Botias, P.; Bajo, M.A.; et al. Genomic reprograming analysis of the Mesothelial to Mesenchymal Transition identifies biomarkers in peritoneal dialysis patients. Sci. Rep. 2017, 7, 44941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loureiro, J.; Aguilera, A.; Selgas, R.; Sandoval, P.; Albar-Vizcaino, P.; Perez-Lozano, M.L.; Ruiz-Carpio, V.; Majano, P.L.; Lamas, S.; Rodriguez-Pascual, F.; et al. Blocking TGF-beta1 protects the peritoneal membrane from dialysate-induced damage. J. Am. Soc. Nephrol. 2011, 22, 1682–1695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Cabrera, M. Mesenchymal Conversion of Mesothelial Cells Is a Key Event in the Pathophysiology of the Peritoneum during Peritoneal Dialysis. Adv. Med. 2014, 2014, 473134. [Google Scholar] [CrossRef]
- McKenzie, A.J.; Hicks, S.R.; Svec, K.V.; Naughton, H.; Edmunds, Z.L.; Howe, A.K. The mechanical microenvironment regulates ovarian cancer cell morphology, migration, and spheroid disaggregation. Sci. Rep. 2018, 8, 7228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strippoli, R.; Sandoval, P.; Moreno-Vicente, R.; Rossi, L.; Battistelli, C.; Terri, M.; Pascual-Anton, L.; Loureiro, M.; Matteini, F.; Calvo, E.; et al. Caveolin1 and YAP drive mechanically induced mesothelial to mesenchymal transition and fibrosis. Cell Death Dis. 2020, 11, 647. [Google Scholar] [CrossRef]
- Ahmed, N.; Stenvers, K.L. Getting to know ovarian cancer ascites: Opportunities for targeted therapy-based translational research. Front. Oncol. 2013, 3, 256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kipps, E.; Tan, D.S.; Kaye, S.B. Meeting the challenge of ascites in ovarian cancer: New avenues for therapy and research. Nat. Rev. Cancer 2013, 13, 273–282. [Google Scholar] [CrossRef] [Green Version]
- Sangisetty, S.L.; Miner, T.J. Malignant ascites: A review of prognostic factors, pathophysiology and therapeutic measures. World J. Gastrointest. Surg. 2012, 4, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Peterson, V.M.; Castro, C.M.; Chung, J.; Miller, N.C.; Ullal, A.V.; Castano, M.D.; Penson, R.T.; Lee, H.; Birrer, M.J.; Weissleder, R. Ascites analysis by a microfluidic chip allows tumor-cell profiling. Proc. Natl. Acad. Sci. USA 2013, 110, E4978–E4986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Q.; Yang, Z.; Xu, S.; Li, X.; Yang, X.; Jin, P.; Liu, Y.; Zhou, X.; Zhang, T.; Gong, C.; et al. Heterotypic CAF-tumor spheroids promote early peritoneal metastatis of ovarian cancer. J. Exp. Med. 2019, 216, 688–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Q.; Huang, B.; Huang, Z.; Cai, J.; Gong, L.; Zhang, Y.; Jiang, J.; Dong, W.; Wang, Z. Tumor cellfibroblast heterotypic aggregates in malignant ascites of patients with ovarian cancer. Int. J. Mol. Med. 2019, 44, 2245–2255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kan, T.; Wang, W.; Ip, P.P.; Zhou, S.; Wong, A.S.; Wang, X.; Yang, M. Single-cell EMT-related transcriptional analysis revealed intra-cluster heterogeneity of tumor cell clusters in epithelial ovarian cancer ascites. Oncogene 2020, 39, 4227–4240. [Google Scholar] [CrossRef] [PubMed]
- Im, H.; Shao, H.; Park, Y.I.; Peterson, V.M.; Castro, C.M.; Weissleder, R.; Lee, H. Label-free detection and molecular profiling of exosomes with a nano-plasmonic sensor. Nat. Biotechnol. 2014, 32, 490–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, W.; Dean, D.C.; Hornicek, F.J.; Shi, H.; Duan, Z. Exosomes promote pre-metastatic niche formation in ovarian cancer. Mol. Cancer 2019, 18, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanez-Mo, M.; Siljander, P.R.; Andreu, Z.; Zavec, A.B.; Borras, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, Y.; Liu, H.; Tang, W.H. Exosomes: Biogenesis, biologic function and clinical potential. Cell Biosci. 2019, 9, 19. [Google Scholar] [CrossRef] [PubMed]
- Mashouri, L.; Yousefi, H.; Aref, A.R.; Ahadi, A.M.; Molaei, F.; Alahari, S.K. Exosomes: Composition, biogenesis, and mechanisms in cancer metastasis and drug resistance. Mol. Cancer 2019, 18, 75. [Google Scholar] [CrossRef]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Record, M.; Carayon, K.; Poirot, M.; Silvente-Poirot, S. Exosomes as new vesicular lipid transporters involved in cell-cell communication and various pathophysiologies. Biochim. Biophys. Acta 2014, 1841, 108–120. [Google Scholar] [CrossRef]
- Gezer, U.; Ozgur, E.; Cetinkaya, M.; Isin, M.; Dalay, N. Long non-coding RNAs with low expression levels in cells are enriched in secreted exosomes. Cell Biol. Int. 2014, 38, 1076–1079. [Google Scholar] [CrossRef]
- Valadi, H.; Ekstrom, K.; Bossios, A.; Sjostrand, M.; Lee, J.J.; Lotvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakur, B.K.; Zhang, H.; Becker, A.; Matei, I.; Huang, Y.; Costa-Silva, B.; Zheng, Y.; Hoshino, A.; Brazier, H.; Xiang, J.; et al. Double-stranded DNA in exosomes: A novel biomarker in cancer detection. Cell Res. 2014, 24, 766–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guescini, M.; Guidolin, D.; Vallorani, L.; Casadei, L.; Gioacchini, A.M.; Tibollo, P.; Battistelli, M.; Falcieri, E.; Battistin, L.; Agnati, L.F.; et al. C2C12 myoblasts release micro-vesicles containing mtDNA and proteins involved in signal transduction. Exp. Cell Res. 2010, 316, 1977–1984. [Google Scholar] [CrossRef] [PubMed]
- Sansone, P.; Savini, C.; Kurelac, I.; Chang, Q.; Amato, L.B.; Strillacci, A.; Stepanova, A.; Iommarini, L.; Mastroleo, C.; Daly, L.; et al. Packaging and transfer of mitochondrial DNA via exosomes regulate escape from dormancy in hormonal therapy-resistant breast cancer. Proc. Natl. Acad. Sci. USA 2017, 114, E9066–E9075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huotari, J.; Helenius, A. Endosome maturation. EMBO J. 2011, 30, 3481–3500. [Google Scholar] [CrossRef]
- Minciacchi, V.R.; Freeman, M.R.; Di Vizio, D. Extracellular vesicles in cancer: Exosomes, microvesicles and the emerging role of large oncosomes. Semin. Cell Dev. Biol. 2015, 40, 41–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isola, A.L.; Chen, S. Exosomes: The Messengers of Health and Disease. Curr. Neuropharmacol. 2017, 15, 157–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teis, D.; Saksena, S.; Emr, S.D. Ordered assembly of the ESCRT-III complex on endosomes is required to sequester cargo during MVB formation. Dev. Cell 2008, 15, 578–589. [Google Scholar] [CrossRef] [Green Version]
- Raiborg, C.; Bache, K.G.; Gillooly, D.J.; Madshus, I.H.; Stang, E.; Stenmark, H. Hrs sorts ubiquitinated proteins into clathrin-coated microdomains of early endosomes. Nat. Cell Biol. 2002, 4, 394–398. [Google Scholar] [CrossRef]
- Teo, H.; Gill, D.J.; Sun, J.; Perisic, O.; Veprintsev, D.B.; Vallis, Y.; Emr, S.D.; Williams, R.L. ESCRT-I core and ESCRT-II GLUE domain structures reveal role for GLUE in linking to ESCRT-I and membranes. Cell 2006, 125, 99–111. [Google Scholar] [CrossRef] [Green Version]
- Trajkovic, K.; Hsu, C.; Chiantia, S.; Rajendran, L.; Wenzel, D.; Wieland, F.; Schwille, P.; Brugger, B.; Simons, M. Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science 2008, 319, 1244–1247. [Google Scholar] [CrossRef] [PubMed]
- Strauss, K.; Goebel, C.; Runz, H.; Mobius, W.; Weiss, S.; Feussner, I.; Simons, M.; Schneider, A. Exosome secretion ameliorates lysosomal storage of cholesterol in Niemann-Pick type C disease. J. Biol. Chem. 2010, 285, 26279–26288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Niel, G.; Charrin, S.; Simoes, S.; Romao, M.; Rochin, L.; Saftig, P.; Marks, M.S.; Rubinstein, E.; Raposo, G. The tetraspanin CD63 regulates ESCRT-independent and -dependent endosomal sorting during melanogenesis. Dev. Cell 2011, 21, 708–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuffers, S.; Sem Wegner, C.; Stenmark, H.; Brech, A. Multivesicular endosome biogenesis in the absence of ESCRTs. Traffic 2009, 10, 925–937. [Google Scholar] [CrossRef] [PubMed]
- Verweij, F.J.; van Eijndhoven, M.A.; Hopmans, E.S.; Vendrig, T.; Wurdinger, T.; Cahir-McFarland, E.; Kieff, E.; Geerts, D.; van der Kant, R.; Neefjes, J.; et al. LMP1 association with CD63 in endosomes and secretion via exosomes limits constitutive NF-kappaB activation. EMBO J. 2011, 30, 2115–2129. [Google Scholar] [CrossRef]
- Hoshino, A.; Costa-Silva, B.; Shen, T.L.; Rodrigues, G.; Hashimoto, A.; Tesic Mark, M.; Molina, H.; Kohsaka, S.; Di Giannatale, A.; Ceder, S.; et al. Tumour exosome integrins determine organotropic metastasis. Nature 2015, 527, 329–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verweij, F.J.; Bebelman, M.P.; Jimenez, C.R.; Garcia-Vallejo, J.J.; Janssen, H.; Neefjes, J.; Knol, J.C.; de Goeij-de Haas, R.; Piersma, S.R.; Baglio, S.R.; et al. Quantifying exosome secretion from single cells reveals a modulatory role for GPCR signaling. J. Cell Biol. 2018, 217, 1129–1142. [Google Scholar] [CrossRef]
- Edgar, J.R.; Manna, P.T.; Nishimura, S.; Banting, G.; Robinson, M.S. Tetherin is an exosomal tether. Elife 2016, 5, e17180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kowal, J.; Tkach, M.; Thery, C. Biogenesis and secretion of exosomes. Curr. Opin. Cell Biol. 2014, 29, 116–125. [Google Scholar] [CrossRef] [Green Version]
- Mathieu, M.; Martin-Jaular, L.; Lavieu, G.; Thery, C. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat. Cell Biol. 2019, 21, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Nanbo, A.; Kawanishi, E.; Yoshida, R.; Yoshiyama, H. Exosomes derived from Epstein-Barr virus-infected cells are internalized via caveola-dependent endocytosis and promote phenotypic modulation in target cells. J. Virol. 2013, 87, 10334–10347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa Verdera, H.; Gitz-Francois, J.J.; Schiffelers, R.M.; Vader, P. Cellular uptake of extracellular vesicles is mediated by clathrin-independent endocytosis and macropinocytosis. J. Control. Release 2017, 266, 100–108. [Google Scholar] [CrossRef]
- Escrevente, C.; Keller, S.; Altevogt, P.; Costa, J. Interaction and uptake of exosomes by ovarian cancer cells. BMC Cancer 2011, 11, 108. [Google Scholar] [CrossRef] [Green Version]
- Svensson, K.J.; Christianson, H.C.; Wittrup, A.; Bourseau-Guilmain, E.; Lindqvist, E.; Svensson, L.M.; Morgelin, M.; Belting, M. Exosome uptake depends on ERK1/2-heat shock protein 27 signaling and lipid Raft-mediated endocytosis negatively regulated by caveolin-1. J. Biol. Chem. 2013, 288, 17713–17724. [Google Scholar] [CrossRef] [Green Version]
- Feng, D.; Zhao, W.L.; Ye, Y.Y.; Bai, X.C.; Liu, R.Q.; Chang, L.F.; Zhou, Q.; Sui, S.F. Cellular internalization of exosomes occurs through phagocytosis. Traffic 2010, 11, 675–687. [Google Scholar] [CrossRef]
- Tian, T.; Zhu, Y.L.; Zhou, Y.Y.; Liang, G.F.; Wang, Y.Y.; Hu, F.H.; Xiao, Z.D. Exosome uptake through clathrin-mediated endocytosis and macropinocytosis and mediating miR-21 delivery. J. Biol. Chem. 2014, 289, 22258–22267. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Li, S.; Xu, Q.; Zhang, X.; Huang, M.; Dai, X.; Liu, L. Exosomes Promote Pre-Metastatic Niche Formation in Gastric Cancer. Front. Oncol. 2021, 11, 652378. [Google Scholar] [CrossRef] [PubMed]
- Serrati, S.; Porcelli, L.; Fragassi, F.; Garofoli, M.; Di Fonte, R.; Fucci, L.; Iacobazzi, R.M.; Palazzo, A.; Margheri, F.; Cristiani, G.; et al. The Interaction between Reactive Peritoneal Mesothelial Cells and Tumor Cells via Extracellular Vesicles Facilitates Colorectal Cancer Dissemination. Cancers 2021, 13, 2505. [Google Scholar] [CrossRef]
- Caby, M.P.; Lankar, D.; Vincendeau-Scherrer, C.; Raposo, G.; Bonnerot, C. Exosomal-like vesicles are present in human blood plasma. Int. Immunol. 2005, 17, 879–887. [Google Scholar] [CrossRef] [Green Version]
- Lasser, C.; Eldh, M.; Lotvall, J. Isolation and characterization of RNA-containing exosomes. J. Vis. Exp. 2012, 59, e3037. [Google Scholar] [CrossRef] [PubMed]
- Pisitkun, T.; Shen, R.F.; Knepper, M.A. Identification and proteomic profiling of exosomes in human urine. Proc. Natl. Acad. Sci. USA 2004, 101, 13368–13373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poliakov, A.; Spilman, M.; Dokland, T.; Amling, C.L.; Mobley, J.A. Structural heterogeneity and protein composition of exosome-like vesicles (prostasomes) in human semen. Prostate 2009, 69, 159–167. [Google Scholar] [CrossRef]
- Keller, S.; Ridinger, J.; Rupp, A.K.; Janssen, J.W.; Altevogt, P. Body fluid derived exosomes as a novel template for clinical diagnostics. J. Transl. Med. 2011, 9, 86. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.A.; Park, H.; Lim, E.H.; Lee, K.W. Exosomes from breast cancer cells can convert adipose tissue-derived mesenchymal stem cells into myofibroblast-like cells. Int. J. Oncol. 2012, 40, 130–138. [Google Scholar] [CrossRef] [Green Version]
- Nazarenko, I.; Rana, S.; Baumann, A.; McAlear, J.; Hellwig, A.; Trendelenburg, M.; Lochnit, G.; Preissner, K.T.; Zoller, M. Cell surface tetraspanin Tspan8 contributes to molecular pathways of exosome-induced endothelial cell activation. Cancer Res. 2010, 70, 1668–1678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, A.; Thakur, B.K.; Weiss, J.M.; Kim, H.S.; Peinado, H.; Lyden, D. Extracellular Vesicles in Cancer: Cell-to-Cell Mediators of Metastasis. Cancer Cell 2016, 30, 836–848. [Google Scholar] [CrossRef] [Green Version]
- Willms, E.; Cabanas, C.; Mager, I.; Wood, M.J.A.; Vader, P. Extracellular Vesicle Heterogeneity: Subpopulations, Isolation Techniques, and Diverse Functions in Cancer Progression. Front. Immunol. 2018, 9, 738. [Google Scholar] [CrossRef] [Green Version]
- Carollo, E.; Paris, B.; Samuel, P.; Pantazi, P.; Bartelli, T.F.; Dias-Neto, E.; Brooks, S.A.; Pink, R.C.; Carter, D.R.F. Detecting ovarian cancer using extracellular vesicles: Progress and possibilities. Biochem. Soc. Trans. 2019, 47, 295–304. [Google Scholar] [CrossRef]
- Li, X.; Wang, X. The emerging roles and therapeutic potential of exosomes in epithelial ovarian cancer. Mol. Cancer 2017, 16, 92. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, K.; Sawada, K.; Kobayashi, M.; Miyamoto, M.; Shimizu, A.; Yamamoto, M.; Kinose, Y.; Kimura, T. Role of the Exosome in Ovarian Cancer Progression and Its Potential as a Therapeutic Target. Cancers 2019, 11, 1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorayappan, K.D.P.; Wallbillich, J.J.; Cohn, D.E.; Selvendiran, K. The biological significance and clinical applications of exosomes in ovarian cancer. Gynecol. Oncol. 2016, 142, 199–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, J.; Zhu, X.; Fei, J.; Shi, P.; Yu, S.; Zhou, J. Advances of exosome in the development of ovarian cancer and its diagnostic and therapeutic prospect. Onco. Targets Ther. 2018, 11, 2831–2841. [Google Scholar] [CrossRef] [Green Version]
- Lengyel, E. Ovarian cancer development and metastasis. Am. J. Pathol. 2010, 177, 1053–1064. [Google Scholar] [CrossRef] [PubMed]
- Kenny, H.A.; Nieman, K.M.; Mitra, A.K.; Lengyel, E. The first line of intra-abdominal metastatic attack: Breaching the mesothelial cell layer. Cancer Discov. 2011, 1, 100–102. [Google Scholar] [CrossRef] [Green Version]
- Wei, M.; Yang, T.; Chen, X.; Wu, Y.; Deng, X.; He, W.; Yang, J.; Wang, Z. Malignant ascites-derived exosomes promote proliferation and induce carcinoma-associated fibroblasts transition in peritoneal mesothelial cells. Oncotarget 2017, 8, 42262–42271. [Google Scholar] [CrossRef]
- Nakamura, K.; Sawada, K.; Kinose, Y.; Yoshimura, A.; Toda, A.; Nakatsuka, E.; Hashimoto, K.; Mabuchi, S.; Morishige, K.I.; Kurachi, H.; et al. Exosomes Promote Ovarian Cancer Cell Invasion through Transfer of CD44 to Peritoneal Mesothelial Cells. Mol. Cancer Res. 2017, 15, 78–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lessan, K.; Aguiar, D.J.; Oegema, T.; Siebenson, L.; Skubitz, A.P. CD44 and beta1 integrin mediate ovarian carcinoma cell adhesion to peritoneal mesothelial cells. Am. J. Pathol. 1999, 154, 1525–1537. [Google Scholar] [CrossRef]
- Strobel, T.; Swanson, L.; Cannistra, S.A. In vivo inhibition of CD44 limits intra-abdominal spread of a human ovarian cancer xenograft in nude mice: A novel role for CD44 in the process of peritoneal implantation. Cancer Res. 1997, 57, 1228–1232. [Google Scholar]
- Kim, Y.; Lee, Y.S.; Choe, J.; Lee, H.; Kim, Y.M.; Jeoung, D. CD44-epidermal growth factor receptor interaction mediates hyaluronic acid-promoted cell motility by activating protein kinase C signaling involving Akt, Rac1, Phox, reactive oxygen species, focal adhesion kinase, and MMP-2. J. Biol. Chem. 2008, 283, 22513–22528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardner, M.J.; Catterall, J.B.; Jones, L.M.; Turner, G.A. Human ovarian tumour cells can bind hyaluronic acid via membrane CD44: A possible step in peritoneal metastasis. Clin. Exp. Metastasis 1996, 14, 325–334. [Google Scholar] [CrossRef]
- Bourguignon, L.Y.; Zhu, H.; Zhou, B.; Diedrich, F.; Singleton, P.A.; Hung, M.C. Hyaluronan promotes CD44v3-Vav2 interaction with Grb2-p185(HER2) and induces Rac1 and Ras signaling during ovarian tumor cell migration and growth. J. Biol. Chem. 2001, 276, 48679–48692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, D.D.; Gercel-Taylor, C. MicroRNA signatures of tumor-derived exosomes as diagnostic biomarkers of ovarian cancer. Gynecol. Oncol. 2008, 110, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Iorio, M.V.; Visone, R.; Di Leva, G.; Donati, V.; Petrocca, F.; Casalini, P.; Taccioli, C.; Volinia, S.; Liu, C.G.; Alder, H.; et al. MicroRNA signatures in human ovarian cancer. Cancer Res. 2007, 67, 8699–8707. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Wu, L.R.; Li, C.; Zhou, X.; Liu, P.; Jia, X.; Chen, Y.; Zhu, W. Five serum microRNAs for detection and predicting of ovarian cancer. Eur. J. Obs. Gynecol. Reprod. Biol. X 2019, 3, 100017. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, A.; Sawada, K.; Nakamura, K.; Kinose, Y.; Nakatsuka, E.; Kobayashi, M.; Miyamoto, M.; Ishida, K.; Matsumoto, Y.; Kodama, M.; et al. Exosomal miR-99a-5p is elevated in sera of ovarian cancer patients and promotes cancer cell invasion by increasing fibronectin and vitronectin expression in neighboring peritoneal mesothelial cells. BMC Cancer 2018, 18, 1065. [Google Scholar] [CrossRef] [PubMed]
- Kenny, H.A.; Chiang, C.Y.; White, E.A.; Schryver, E.M.; Habis, M.; Romero, I.L.; Ladanyi, A.; Penicka, C.V.; George, J.; Matlin, K.; et al. Mesothelial cells promote early ovarian cancer metastasis through fibronectin secretion. J. Clin. Investig. 2014, 124, 4614–4628. [Google Scholar] [CrossRef] [Green Version]
- Heyman, L.; Kellouche, S.; Fernandes, J.; Dutoit, S.; Poulain, L.; Carreiras, F. Vitronectin and its receptors partly mediate adhesion of ovarian cancer cells to peritoneal mesothelium in vitro. Tumour Biol. 2008, 29, 231–244. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wang, J.; Shen, X.; Li, M.; Yue, Y.; Cheng, X.; Lu, W.; Wang, X.; Xie, X. LncRNA SPOCD1-AS from ovarian cancer extracellular vesicles remodels mesothelial cells to promote peritoneal metastasis via interacting with G3BP1. J. Exp. Clin. Cancer Res. 2021, 40, 101. [Google Scholar] [CrossRef]
- Guan, X.; Zong, Z.H.; Liu, Y.; Chen, S.; Wang, L.L.; Zhao, Y. CircPUM1 Promotes Tumorigenesis and Progression of Ovarian Cancer by Sponging miR-615-5p and miR-6753-5p. Mol. Ther. Nucleic Acids 2019, 18, 882–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zong, Z.H.; Du, Y.P.; Guan, X.; Chen, S.; Zhao, Y. CircWHSC1 promotes ovarian cancer progression by regulating MUC1 and hTERT through sponging miR-145 and miR-1182. J. Exp. Clin. Cancer Res. 2019, 38, 437. [Google Scholar] [CrossRef]
- Lee, A.H.; Ghosh, D.; Quach, N.; Schroeder, D.; Dawson, M.R. Ovarian Cancer Exosomes Trigger Differential Biophysical Response in Tumor-Derived Fibroblasts. Sci. Rep. 2020, 10, 8686. [Google Scholar] [CrossRef]
- Lee, W.; Ko, S.Y.; Mohamed, M.S.; Kenny, H.A.; Lengyel, E.; Naora, H. Neutrophils facilitate ovarian cancer premetastatic niche formation in the omentum. J. Exp. Med. 2019, 216, 176–194. [Google Scholar] [CrossRef] [PubMed]
- Millimaggi, D.; Mari, M.; D’Ascenzo, S.; Carosa, E.; Jannini, E.A.; Zucker, S.; Carta, G.; Pavan, A.; Dolo, V. Tumor vesicle-associated CD147 modulates the angiogenic capability of endothelial cells. Neoplasia 2007, 9, 349–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, H.; Ye, J.; Yang, X.M.; Zhang, L.W.; Zhang, Z.G.; Chen, Y.P. High-grade ovarian cancer secreting effective exosomes in tumor angiogenesis. Int. J. Clin. Exp. Pathol. 2015, 8, 5062–5070. [Google Scholar]
- Bretz, N.P.; Ridinger, J.; Rupp, A.K.; Rimbach, K.; Keller, S.; Rupp, C.; Marme, F.; Umansky, L.; Umansky, V.; Eigenbrod, T.; et al. Body fluid exosomes promote secretion of inflammatory cytokines in monocytic cells via Toll-like receptor signaling. J. Biol. Chem. 2013, 288, 36691–36702. [Google Scholar] [CrossRef] [Green Version]
- Kelleher, R.J., Jr.; Balu-Iyer, S.; Loyall, J.; Sacca, A.J.; Shenoy, G.N.; Peng, P.; Iyer, V.; Fathallah, A.M.; Berenson, C.S.; Wallace, P.K.; et al. Extracellular Vesicles Present in Human Ovarian Tumor Microenvironments Induce a Phosphatidylserine-Dependent Arrest in the T-cell Signaling Cascade. Cancer Immunol. Res. 2015, 3, 1269–1278. [Google Scholar] [CrossRef] [Green Version]
- Shenoy, G.N.; Loyall, J.; Berenson, C.S.; Kelleher, R.J., Jr.; Iyer, V.; Balu-Iyer, S.V.; Odunsi, K.; Bankert, R.B. Sialic Acid-Dependent Inhibition of T Cells by Exosomal Ganglioside GD3 in Ovarian Tumor Microenvironments. J. Immunol. 2018, 201, 3750–3758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, D.D.; Gercel-Taylor, C.; Lyons, K.S.; Stanson, J.; Whiteside, T.L. T-cell apoptosis and suppression of T-cell receptor/CD3-zeta by Fas ligand-containing membrane vesicles shed from ovarian tumors. Clin. Cancer Res. 2003, 9, 5113–5119. [Google Scholar] [PubMed]
- Taylor, D.D.; Gercel-Taylor, C. Tumour-derived exosomes and their role in cancer-associated T-cell signalling defects. Br. J. Cancer 2005, 92, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Peng, P.; Yan, Y.; Keng, S. Exosomes in the ascites of ovarian cancer patients: Origin and effects on anti-tumor immunity. Oncol. Rep. 2011, 25, 749–762. [Google Scholar] [CrossRef] [Green Version]
- Czystowska-Kuzmicz, M.; Sosnowska, A.; Nowis, D.; Ramji, K.; Szajnik, M.; Chlebowska-Tuz, J.; Wolinska, E.; Gaj, P.; Grazul, M.; Pilch, Z.; et al. Small extracellular vesicles containing arginase-1 suppress T-cell responses and promote tumor growth in ovarian carcinoma. Nat. Commun. 2019, 10, 3000. [Google Scholar] [CrossRef]
- Lundholm, M.; Schroder, M.; Nagaeva, O.; Baranov, V.; Widmark, A.; Mincheva-Nilsson, L.; Wikstrom, P. Prostate tumor-derived exosomes down-regulate NKG2D expression on natural killer cells and CD8+ T cells: Mechanism of immune evasion. PLoS ONE 2014, 9, e108925. [Google Scholar] [CrossRef] [PubMed]
- Ying, X.; Wu, Q.; Wu, X.; Zhu, Q.; Wang, X.; Jiang, L.; Chen, X.; Wang, X. Epithelial ovarian cancer-secreted exosomal miR-222-3p induces polarization of tumor-associated macrophages. Oncotarget 2016, 7, 43076–43087. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Zhou, J.; Li, X.; Wang, X.; Lin, Y.; Wang, X. Exosomes derived from hypoxic epithelial ovarian cancer cells deliver microRNAs to macrophages and elicit a tumor-promoted phenotype. Cancer Lett. 2018, 435, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.W.; Yu, T.J.; Zhang, J.; Li, Y.; Chen, H.L.; Yang, G.F.; Yu, W.; Liu, Y.Z.; Liu, X.X.; Duan, C.F.; et al. CYP4A in tumor-associated macrophages promotes pre-metastatic niche formation and metastasis. Oncogene 2017, 36, 5045–5057. [Google Scholar] [CrossRef] [Green Version]
- Hang, W.; Feng, Y.; Sang, Z.; Yang, Y.; Zhu, Y.; Huang, Q.; Xi, X. Downregulation of miR-145-5p in cancer cells and their derived exosomes may contribute to the development of ovarian cancer by targeting CT. Int. J. Mol. Med. 2019, 43, 256–266. [Google Scholar] [CrossRef] [Green Version]
- Ma, R.; Ye, X.; Cheng, H.; Cui, H.; Chang, X. Tumor-derived exosomal circRNA051239 promotes proliferation and migration of epithelial ovarian cancer. Am. J. Transl. Res. 2021, 13, 1125–1139. [Google Scholar]
- He, L.; Zhu, W.; Chen, Q.; Yuan, Y.; Wang, Y.; Wang, J.; Wu, X. Ovarian cancer cell-secreted exosomal miR-205 promotes metastasis by inducing angiogenesis. Theranostics 2019, 9, 8206–8220. [Google Scholar] [CrossRef]
- Satapathy, S.; Kumar, C.; Singh, R.K. MicroRNAs as Key Regulators of Ovarian Cancers. Cell Med. 2019, 11, 2155179019873849. [Google Scholar] [CrossRef]
- Ghorbanian, M.; Babashah, S.; Ataei, F. The effects of ovarian cancer cell-derived exosomes on vascular endothelial growth factor expression in endothelial cells. EXCLI J. 2019, 18, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Masoumi-Dehghi, S.; Babashah, S.; Sadeghizadeh, M. microRNA-141-3p-containing small extracellular vesicles derived from epithelial ovarian cancer cells promote endothelial cell angiogenesis through activating the JAK/STAT3 and NF-kappaB signaling pathways. J. Cell Commun. Signal. 2020, 14, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Tao, L.; Huang, G.; Song, H.; Chen, Y.; Chen, L. Cancer associated fibroblasts: An essential role in the tumor microenvironment. Oncol. Lett. 2017, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, T.; Zhang, P.; Zhao, F.; Zhang, S. Exosomal MicroRNAs Mediating Crosstalk Between Cancer Cells With Cancer-Associated Fibroblasts and Tumor-Associated Macrophages in the Tumor Microenvironment. Front. Oncol. 2021, 11, 631703. [Google Scholar] [CrossRef]
- Guo, H.; Ha, C.; Dong, H.; Yang, Z.; Ma, Y.; Ding, Y. Cancer-associated fibroblast-derived exosomal microRNA-98-5p promotes cisplatin resistance in ovarian cancer by targeting CDKN1A. Cancer Cell Int. 2019, 19, 347. [Google Scholar] [CrossRef]
- Li, W.; Zhang, X.; Wang, J.; Li, M.; Cao, C.; Tan, J.; Ma, D.; Gao, Q. TGFbeta1 in fibroblasts-derived exosomes promotes epithelial-mesenchymal transition of ovarian cancer cells. Oncotarget 2017, 8, 96035–96047. [Google Scholar] [CrossRef] [Green Version]
- Au Yeung, C.L.; Co, N.N.; Tsuruga, T.; Yeung, T.L.; Kwan, S.Y.; Leung, C.S.; Li, Y.; Lu, E.S.; Kwan, K.; Wong, K.K.; et al. Exosomal transfer of stroma-derived miR21 confers paclitaxel resistance in ovarian cancer cells through targeting APAF1. Nat. Commun. 2016, 7, 11150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Anton, M.; Lambie, M.; Lopez-Cabrera, M.; Schmitt, C.P.; Ruiz-Carpio, V.; Bartosova, M.; Schaefer, B.; Davies, S.; Stone, T.; Jenkins, R.; et al. miR-21 Promotes Fibrogenesis in Peritoneal Dialysis. Am. J. Pathol. 2017, 187, 1537–1550. [Google Scholar] [CrossRef] [Green Version]
- Mulcahy, L.A.; Pink, R.C.; Carter, D.R. Routes and mechanisms of extracellular vesicle uptake. J. Extracell. Vesicles 2014, 3, 24641. [Google Scholar] [CrossRef] [Green Version]
- French, K.C.; Antonyak, M.A.; Cerione, R.A. Extracellular vesicle docking at the cellular port: Extracellular vesicle binding and uptake. Semin. Cell Dev. Biol. 2017, 67, 48–55. [Google Scholar] [CrossRef]
- Cardenes, B.; Clares, I.; Toribio, V.; Pascual, L.; Lopez-Martin, S.; Torres-Gomez, A.; Sainz de la Cuesta, R.; Lafuente, E.M.; Lopez-Cabrera, M.; Yanez-Mo, M.; et al. Cellular Integrin alpha5beta1 and Exosomal ADAM17 Mediate the Binding and Uptake of Exosomes Produced by Colorectal Carcinoma Cells. Int. J. Mol. Sci. 2021, 22, 9938. [Google Scholar] [CrossRef]
- Laulagnier, K.; Javalet, C.; Hemming, F.J.; Chivet, M.; Lachenal, G.; Blot, B.; Chatellard, C.; Sadoul, R. Amyloid precursor protein products concentrate in a subset of exosomes specifically endocytosed by neurons. Cell Mol. Life Sci. 2018, 75, 757–773. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, M.; Haider, A.; Rashid, S.; Al-Nabet, A. Paget’s “Seed and Soil” Theory of Cancer Metastasis: An Idea Whose Time has Come. Adv. Anat. Pathol. 2019, 26, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Szajnik, M.; Derbis, M.; Lach, M.; Patalas, P.; Michalak, M.; Drzewiecka, H.; Szpurek, D.; Nowakowski, A.; Spaczynski, M.; Baranowski, W.; et al. Exosomes in Plasma of Patients with Ovarian Carcinoma: Potential Biomarkers of Tumor Progression and Response to Therapy. Gynecol. Obs. 2013, (Suppl. 4), 3. [Google Scholar] [CrossRef] [Green Version]
- Yeung, T.L.; Leung, C.S.; Wong, K.K.; Samimi, G.; Thompson, M.S.; Liu, J.; Zaid, T.M.; Ghosh, S.; Birrer, M.J.; Mok, S.C. TGF-beta modulates ovarian cancer invasion by upregulating CAF-derived versican in the tumor microenvironment. Cancer Res. 2013, 73, 5016–5028. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Sherman-Baust, C.A.; Tsai-Turton, M.; Bristow, R.E.; Roden, R.B.; Morin, P.J. Claudin-containing exosomes in the peripheral circulation of women with ovarian cancer. BMC Cancer 2009, 9, 244. [Google Scholar] [CrossRef] [Green Version]
- Vaksman, O.; Trope, C.; Davidson, B.; Reich, R. Exosome-derived miRNAs and ovarian carcinoma progression. Carcinogenesis 2014, 35, 2113–2120. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.H.L.; Yue, C.; Du, K.Y.; Salem, M.; O’Brien, J.; Peng, C. The Role of microRNAs in Epithelial Ovarian Cancer Metastasis. Int. J. Mol. Sci. 2020, 21, 7093. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Shoorei, H.; Taheri, M. miRNA profile in ovarian cancer. Exp. Mol. Pathol. 2020, 113, 104381. [Google Scholar] [CrossRef]
- Keller, S.; Konig, A.K.; Marme, F.; Runz, S.; Wolterink, S.; Koensgen, D.; Mustea, A.; Sehouli, J.; Altevogt, P. Systemic presence and tumor-growth promoting effect of ovarian carcinoma released exosomes. Cancer Lett. 2009, 278, 73–81. [Google Scholar] [CrossRef]
- Alharbi, M.; Zuniga, F.; Elfeky, O.; Guanzon, D.; Lai, A.; Rice, G.E.; Perrin, L.; Hooper, J.; Salomon, C. The potential role of miRNAs and exosomes in chemotherapy in ovarian cancer. Endocr. Relat. Cancer 2018, 25, R663–R685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinbichler, T.B.; Dudas, J.; Skvortsov, S.; Ganswindt, U.; Riechelmann, H.; Skvortsova, I.I. Therapy resistance mediated by exosomes. Mol. Cancer 2019, 18, 58. [Google Scholar] [CrossRef]
- Yan, X.; Yin, J.; Yao, H.; Mao, N.; Yang, Y.; Pan, L. Increased expression of annexin A3 is a mechanism of platinum resistance in ovarian cancer. Cancer Res. 2010, 70, 1616–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, J.; Yan, X.; Yao, X.; Zhang, Y.; Shan, Y.; Mao, N.; Yang, Y.; Pan, L. Secretion of annexin A3 from ovarian cancer cells and its association with platinum resistance in ovarian cancer patients. J. Cell Mol. Med. 2012, 16, 337–348. [Google Scholar] [CrossRef]
- Safaei, R.; Larson, B.J.; Cheng, T.C.; Gibson, M.A.; Otani, S.; Naerdemann, W.; Howell, S.B. Abnormal lysosomal trafficking and enhanced exosomal export of cisplatin in drug-resistant human ovarian carcinoma cells. Mol. Cancer Ther. 2005, 4, 1595–1604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Y.L.; Zhuang, T.; Xing, B.H.; Li, N.; Li, Q. Exosomal DNMT1 mediates cisplatin resistance in ovarian cancer. Cell Biochem. Funct. 2017, 35, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Runz, S.; Keller, S.; Rupp, C.; Stoeck, A.; Issa, Y.; Koensgen, D.; Mustea, A.; Sehouli, J.; Kristiansen, G.; Altevogt, P. Malignant ascites-derived exosomes of ovarian carcinoma patients contain CD24 and EpCAM. Gynecol. Oncol. 2007, 107, 563–571. [Google Scholar] [CrossRef]
- Tayama, S.; Motohara, T.; Narantuya, D.; Li, C.; Fujimoto, K.; Sakaguchi, I.; Tashiro, H.; Saya, H.; Nagano, O.; Katabuchi, H. The impact of EpCAM expression on response to chemotherapy and clinical outcomes in patients with epithelial ovarian cancer. Oncotarget 2017, 8, 44312–44325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crow, J.; Atay, S.; Banskota, S.; Artale, B.; Schmitt, S.; Godwin, A.K. Exosomes as mediators of platinum resistance in ovarian cancer. Oncotarget 2017, 8, 11917–11936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pink, R.C.; Samuel, P.; Massa, D.; Caley, D.P.; Brooks, S.A.; Carter, D.R. The passenger strand, miR-21-3p, plays a role in mediating cisplatin resistance in ovarian cancer cells. Gynecol. Oncol. 2015, 137, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Weiner-Gorzel, K.; Dempsey, E.; Milewska, M.; McGoldrick, A.; Toh, V.; Walsh, A.; Lindsay, S.; Gubbins, L.; Cannon, A.; Sharpe, D.; et al. Overexpression of the microRNA miR-433 promotes resistance to paclitaxel through the induction of cellular senescence in ovarian cancer cells. Cancer Med. 2015, 4, 745–758. [Google Scholar] [CrossRef]
- Zhu, X.; Shen, H.; Yin, X.; Yang, M.; Wei, H.; Chen, Q.; Feng, F.; Liu, Y.; Xu, W.; Li, Y. Macrophages derived exosomes deliver miR-223 to epithelial ovarian cancer cells to elicit a chemoresistant phenotype. J. Exp. Clin. Cancer Res. 2019, 38, 81. [Google Scholar] [CrossRef] [PubMed]
- Rafii, A.; Mirshahi, P.; Poupot, M.; Faussat, A.M.; Simon, A.; Ducros, E.; Mery, E.; Couderc, B.; Lis, R.; Capdet, J.; et al. Oncologic trogocytosis of an original stromal cells induces chemoresistance of ovarian tumours. PLoS ONE 2008, 3, e3894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cappellesso, R.; Tinazzi, A.; Giurici, T.; Simonato, F.; Guzzardo, V.; Ventura, L.; Crescenzi, M.; Chiarelli, S.; Fassina, A. Programmed cell death 4 and microRNA 21 inverse expression is maintained in cells and exosomes from ovarian serous carcinoma effusions. Cancer Cytopathol. 2014, 122, 685–693. [Google Scholar] [CrossRef]
- Thibault, B.; Castells, M.; Delord, J.P.; Couderc, B. Ovarian cancer microenvironment: Implications for cancer dissemination and chemoresistance acquisition. Cancer Metastasis Rev. 2014, 33, 17–39. [Google Scholar] [CrossRef] [PubMed]
- Samuel, P.; Mulcahy, L.A.; Furlong, F.; McCarthy, H.O.; Brooks, S.A.; Fabbri, M.; Pink, R.C.; Carter, D.R.F. Cisplatin induces the release of extracellular vesicles from ovarian cancer cells that can induce invasiveness and drug resistance in bystander cells. Philos. Trans. R Soc. Lond. B Biol. Sci. 2018, 373, 20170065. [Google Scholar] [CrossRef] [PubMed]
- Bewicke-Copley, F.; Mulcahy, L.A.; Jacobs, L.A.; Samuel, P.; Akbar, N.; Pink, R.C.; Carter, D.R.F. Extracellular vesicles released following heat stress induce bystander effect in unstressed populations. J. Extracell. Vesicles 2017, 6, 1340746. [Google Scholar] [CrossRef]
- de la Fuente, A.; Alonso-Alconada, L.; Costa, C.; Cueva, J.; Garcia-Caballero, T.; Lopez-Lopez, R.; Abal, M. M-Trap: Exosome-Based Capture of Tumor Cells as a New Technology in Peritoneal Metastasis. J. Natl. Cancer Inst. 2015, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorayappan, K.D.P.; Gardner, M.L.; Hisey, C.L.; Zingarelli, R.A.; Smith, B.Q.; Lightfoot, M.D.S.; Gogna, R.; Flannery, M.M.; Hays, J.; Hansford, D.J.; et al. A Microfluidic Chip Enables Isolation of Exosomes and Establishment of Their Protein Profiles and Associated Signaling Pathways in Ovarian Cancer. Cancer Res. 2019, 79, 3503–3513. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Ono, Y.J.; Kanemura, M.; Tanaka, T.; Hayashi, M.; Terai, Y.; Ohmichi, M. Hepatocyte growth factor secreted by ovarian cancer cells stimulates peritoneal implantation via the mesothelial-mesenchymal transition of the peritoneum. Gynecol. Oncol. 2015, 139, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Matte, I.; Lane, D.; Laplante, C.; Garde-Granger, P.; Rancourt, C.; Piche, A. Ovarian cancer ascites enhance the migration of patient-derived peritoneal mesothelial cells via cMet pathway through HGF-dependent and -independent mechanisms. Int. J. Cancer 2015, 137, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, Y.; Bai, M.; Wang, J.; Zhu, K.; Liu, R.; Ge, S.; Li, J.; Ning, T.; Deng, T.; et al. Exosomes serve as nanoparticles to suppress tumor growth and angiogenesis in gastric cancer by delivering hepatocyte growth factor siRNA. Cancer Sci. 2018, 109, 629–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
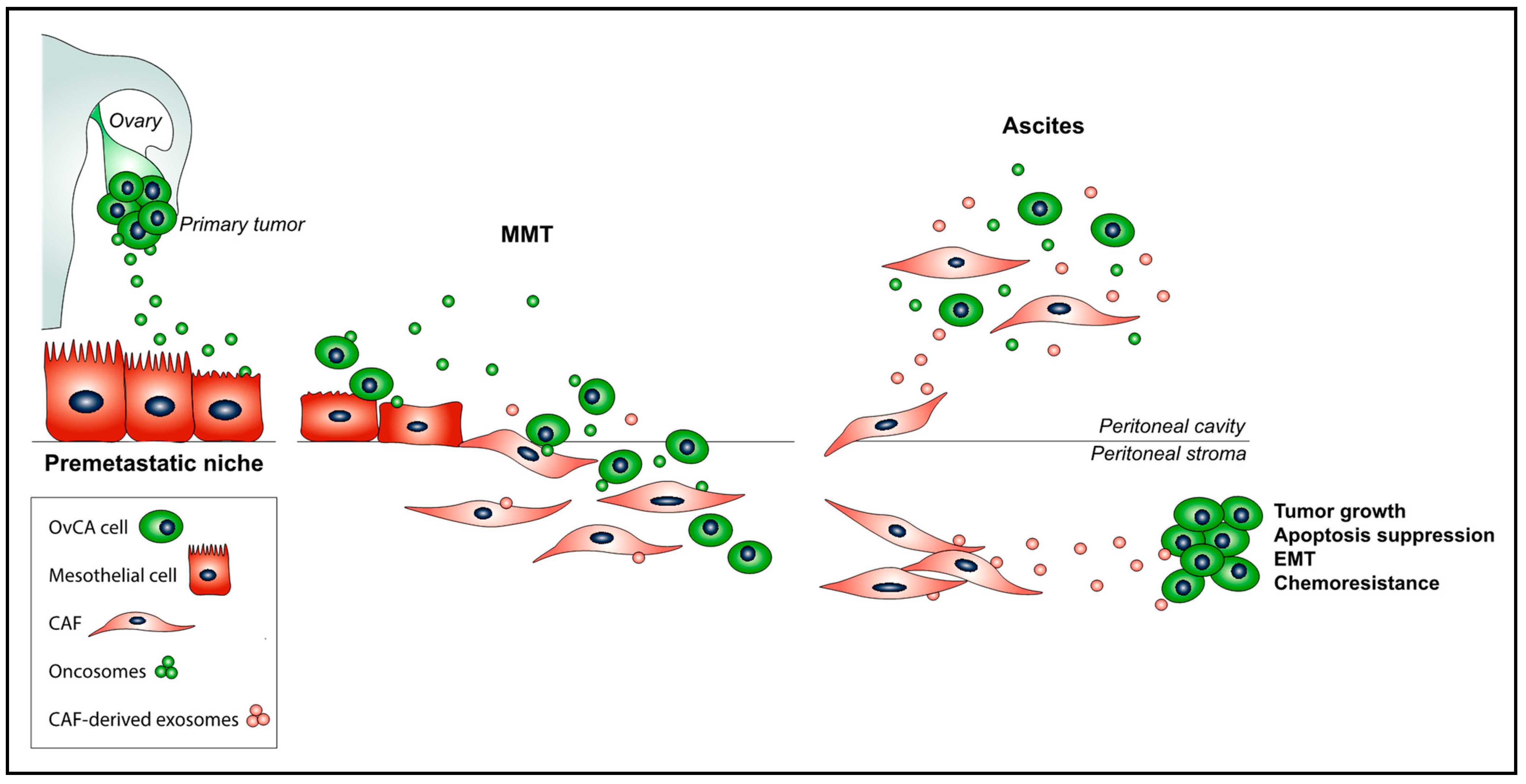
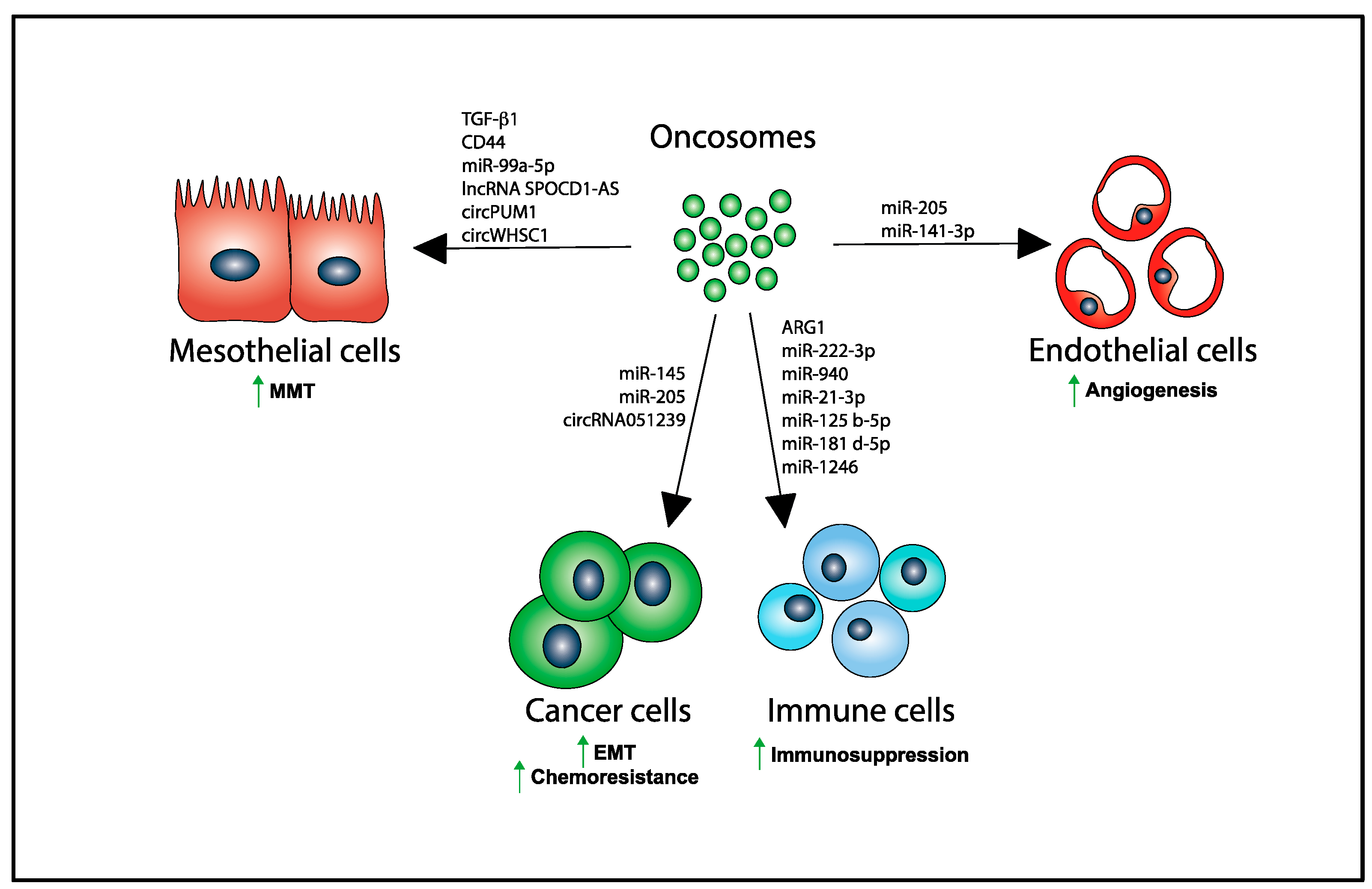
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).