
Short Peptides Make a Big Difference: The Role of Botany-Derived AMPs in Disease Control and Protection of Human Health


Abstract
1. Introduction
2. BAMP Diversity and Mechanisms of Action in Plants
2.1. Plant Defensins: Interacting with Fungal Membrane Components and Targeting Intracellular Pathways
2.2. CAPE Peptides (PR1): Binding to Sterols on Pathogen Membrane and Inhibiting Programmed Cell Death
2.3. Thionins (PR13): Binding to Phospholipids and Disrupting Membrane Permeability
2.4. Lipid Transfer Proteins (LTPs, PR14): Nonspecifically Transporting Lipids
2.5. Proteinase-Inhibitor (PIs, PR6): Inhibiting Proteinase Activity That Is Essential for Pathogen Growth and Pathogenicity

{kind=link}
{kind=link}
{kind=link}
| Family | Representative Peptide | Sources | Mode of Action | References |
|---|---|---|---|---|
| Defensin (PR12) | DmAMP1 | Dahlia merkii | DmAMP1 binds to M(IP)2C in the membrane, resulting in potassium efflux, calcium uptake, membrane permeability change, and CWI pathway activation. | [25,28,29] |
| RsAFP2 | Raphanus sativa | RsAFP2 binds to GlcCer in the cell wall and plasma membrane, resulting in ROS production, apoptosis, ion fluxes, and CWI pathway activation. | [30,31] | |
| MsDef1 | Medicago sativa | MsDef1 interacts with GlcCer in the cell wall and membrane, resulting in the activation of MAPK cascade in the CWI pathway, and disruption of Ca2+ signaling and homeostasis, contributing to fungal cell death. | [32,33,98] | |
| MtDef4 | Mendicago trucatula | MtDef4 binds to PA and is internalized into the fungal cell, resulting in the disruption of Ca2+ signaling and homeostasis in a different way to MsDef1. An interaction with unknown intracellular targets has been proposed. | [34] | |
| NaD1 | Nicotiana alata | NaD1 binds to PI(4,5)P2 and dimerizes in the membrane. The dimer is internalized into the cytoplasm and interacts with intracellular targets to trigger the ROS and NO production. | [33,35] | |
| HsAFP1 | Heuchera sanguinea | HsAFP1 binds to the fungal cell wall and plasma membrane via loop 5. It moves into the cytoplasm and targets mitochondria, produces ROS, and induces programmed cell death. | [36] | |
| Psd1 | Pisum sativum | Psd1 binds to GlcCer in the plasma membrane. It moves to the cytoplasm and interacts with cyclin F in the nucleus, which results in cell cycle arrest and fungal cell death. | [37] | |
| PR1 | PR1a | Nicotiana tabacum | PR1 binds to sterol and sequesters it from pathogens. It inhibits programmed cell death at the pathogen infection sites, and induces the expression of host defense genes by releasing CAPE1 peptide. | [43,45,46,47] |
| P14c | Solanum lycopersicum | |||
| Thionins (PR13) | α1-purothionin β-purothionin | Triticum aestivum | Upon application of purothionin, there is a depolarization of the membrane and Ca2+ ion permeability increases. β-purothionin interacts with dimyristoyl-phosphatidylglycerol, and inhibits protein kinase C. | [59,99] |
| Thionin | Pyrularia pubera | Thionin leads to membrane depolarization, influx of exogenous Ca2+, and activation of PLA2 and adenylate cyclase. | [62] | |
| viscotoxins A3 viscotoxins B | Viscum album | Viscotoxins directly interact with DNA and RNA, interfering with nucleic acid synthesis. | [100] | |
| Lipid transfer proteins (LTPs, PR14) | Ca-LTP1 | Capsicum annuum | Ca-LTP1 penetrates the plasma membrane and causes morphological damage, accompanied by pseudo-mycelia formation. | [77] |
| Ha-AP10 | Helianthus annuus | Ha-AP10 interacts with phospholipids and produces a direct cytotoxic effect on fungal cells mediated by membrane permeabilization. | [78] | |
| Proteinase inhibitor (PIs, PR6) | HyPep | Capsicum annuum | HyPep inhibits α-amylase and serine proteinases, and causes cell aggregation and pseudo-mycelia formation. | [94] |
3. Antimicrobial Activity of BAMPs against Targeted Organisms and Cancer Cells
3.1. Antifungal Activity
3.2. Antiviral Activity
3.3. Anticancer Activity
| Classification | Representative Peptide | Anticancer Activity | Mode of Action | References |
|---|---|---|---|---|
| BAMPs | Lunasin | Skin, colon, prostate, and breast cancers | Lunasin binds directly to deacetylated histones, inhibits acetylation, and turns off the transcription. | [137] |
| Cycloviolacin O2 (CyO2) | Breast cancer and lymphoma cells | CyO2 causes tumor cell death by membrane permeabilization. | [140] | |
| MCo-PMI | Adenocarcinoma | MCo-PMI inhibits tumor proliferation by activating the p53 tumor suppressor pathway. | [141] | |
| HB7 | Pancreatic cancer | HB7 inhibits the proliferation and migration of tumors by membrane permeabilization. | [142] | |
| NaD1 | Monocyte lymphoma | NaD1 inhibits the proliferation of monocyte lymphoma by directly binding to the plasma membrane phosphatidylinositol 4,5-diphosphate. | [143] | |
| Viscotoxin B2 | Osteogenic sarcoma | Viscotoxin B2 inhibits tumor cells by membrane lysis. | [144] | |
| Ligatoxin B | Lymphoma and adenocarcinoma | Ligatoxin B inhibits the proliferation of tumor cells by inhibiting nucleic acid and protein synthesis. | [145] | |
| NTP | Promyelocytic leukemia cells (HL-60) | NTP mediates cancer-related signal transduction cascades by regulating the distribution of lipids in cell membranes, thereby inhibiting tumor cell invasion and metastasis. | [16,146] | |
| Phoratoxins C-F | Different types of solid tumor cells and hematologic tumors | Inhibiting tumor proliferation, while the mechanism of action is unknown. | [147] | |
| Thi2.1 | Tumor cells McF-7, A549, and HeLa | [148] | ||
| Sesquin | McF-7 and leukemia M1 cells | [149] | ||
| Limenin | Leukemia cells | [150] | ||
| Purple pole defensin | HepG2, McF-7, and HT-29 cells | [162] | ||
| Coccinin | HL60 and L1210 cells | [151] | ||
| Other small peptides | Cn-AMP1 | Caco-2 cells | Cn-AMP1 reduces cancer cell viability without causing hemolysis. | [152] |
| Cr-ACP | Hep2 cells | Cr-ACP induces cell cycle arrest in G0–G1 phase. | [153] | |
| Cherimolacyclopeptide C | KB cells | Cherimolacyclopeptide C shows in vitro cytotoxicity to KB cells. | [154] | |
| Cyclosaplin | Breast cancer | Cyclosaplin inhibits cancer cell proliferation in a dose- and time-dependent manner. | [155] | |
| Poca A and B | Breast cancer | Poca inhibits cancer cell migration. | [156] | |
| IbACP | Panc-1, a pancreatic cancer line | IbACP regulates cellular proliferation by inducing and promoting apoptosis through the mitochondrial apoptotic pathway. | [157] | |
| GLTSK | HCT116, human colorectal cancer cells | GLTSK decreases angiotensin II-dependent proliferation in HCT116 through the blockade of the renin-angiotensin system. | [163] |
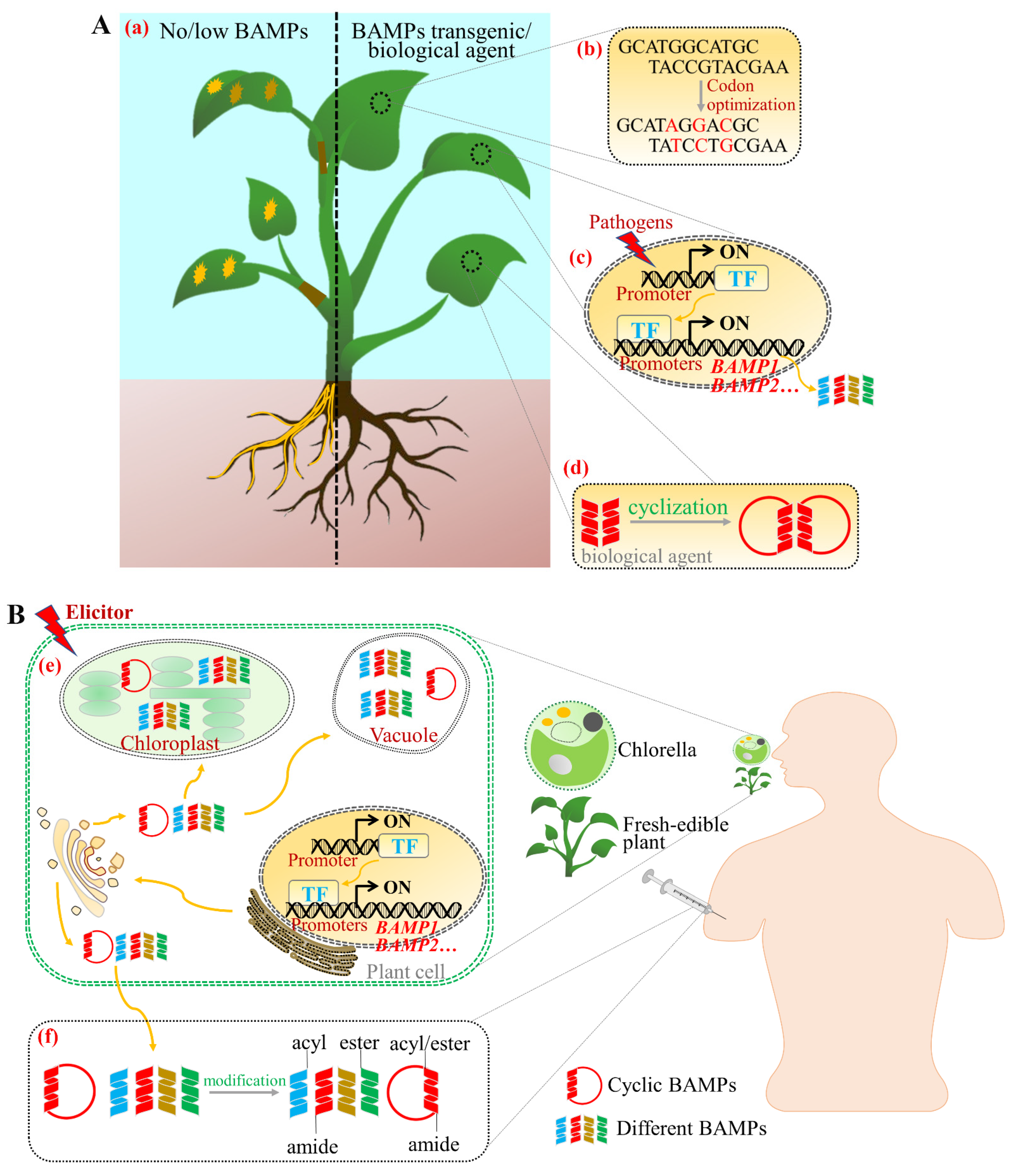
4. Applications of BAMPs for Agricultural Purposes
5. Applications of BAMPs for Human Health
6. Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goyal, R.K.; Mattoo, A.K. Multitasking antimicrobial peptides in plant development and host defense against biotic/abiotic stress. Plant Sci. 2014, 228, 135–149. [Google Scholar] [CrossRef]
- van der Weerden, N.L.; Bleackley, M.R.; Anderson, M.A. Properties and mechanisms of action of naturally occurring antifungal peptides. Cell. Mol. Life Sci. 2013, 70, 3545–3570. [Google Scholar] [CrossRef]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Yount, N.Y.; Yeaman, M.R. Immunocontinuum: Perspectives in antimicrobial peptide mechanisms of action and resistance. Protein Pept. Lett. 2005, 12, 49–67. [Google Scholar] [CrossRef] [PubMed]
- Alfred, R.L.; Palombo, E.A.; Panozzo, J.F.; Bhave, M. The antimicrobial domains of wheat puroindolines are cell-penetrating peptides with possible intracellular mechanisms of action. PLoS ONE 2013, 8, e75488. [Google Scholar] [CrossRef] [PubMed]
- Nanni, V.; Schumacher, J.; Giacomelli, L.; Brazzale, D.; Sbolci, L.; Moser, C.; Tudzynski, P.; Baraldi, E. VvAMP2, a grapevine flower-specific defensin capable of inhibiting Botrytis cinerea growth: Insights into its mode of action. Plant Pathol. 2014, 63, 899–910. [Google Scholar] [CrossRef]
- Struyfs, C.; Cammue, B.P.A.; Thevissen, K. Membrane-interacting antifungal peptides. Front. Cell Dev. Biol. 2021, 9, 706. [Google Scholar] [CrossRef]
- Rahnamaeian, M. Antimicrobial peptides modes of mechanism, modulation of defense responses. Plant Signal. Behav. 2011, 6, 1325–1332. [Google Scholar] [CrossRef]
- Hammami, R.; Ben Hamida, J.; Vergoten, G.; Fliss, I. PhytAMP: A database dedicated to antimicrobial plant peptides. Nucleic Acids Res. 2009, 37, D963–D968. [Google Scholar] [CrossRef]
- Candido, E.d.S.; Silva Cardoso, M.H.e.; Sousa, D.A.; Viana, J.C.; de Oliveira-Junior, N.G.; Miranda, V.; Franco, O.L. The use of versatile plant antimicrobial peptides in agribusiness and human health. Peptides 2014, 55, 65–78. [Google Scholar] [CrossRef]
- Tam, J.P.; Wang, S.; Wong, K.H.; Tan, W.L. Antimicrobial peptides from plants. Pharmaceuticals 2015, 8, 711–757. [Google Scholar] [CrossRef]
- Lay, F.T.; Anderson, M.A. Defensins-components of the innate immune system in plants. Curr. Protein Pept. Sci. 2005, 6, 85–101. [Google Scholar] [CrossRef]
- Terras, F.R.G.; Eggermont, K.; Kovaleva, V.; Raikhel, N.V.; Osborn, R.W.; Kester, A.; Rees, S.B.; Torrekens, S.; Vanleuven, F.; Vanderleyden, J.; et al. SMALL cysteine-rich antifungal proteins from radish-their role in host-defense. Plant Cell 1995, 7, 573–588. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhou, M.; Zhang, Z.; Ren, L.; Du, L.; Zhang, B.; Xu, H.; Xin, Z. Expression of a radish defensin in transgenic wheat confers increased resistance to Fusarium graminearum and Rhizoctonia cerealis. Funct. Integr. Genom. 2011, 11, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Ghag, S.B.; Shekhawat, U.K.S.; Ganapathi, T.R. Petunia floral defensins with unique prodomains as novel candidates for development of Fusarium wilt resistance in transgenic banana plants. PLoS ONE 2012, 7, e39557. [Google Scholar] [CrossRef] [PubMed]
- Ooi, L.S.M.; Tian, L.; Su, M.; Ho, W.S.; Sun, S.S.M.; Chung, H.Y.; Wong, H.N.C.; Ooi, V.E.C. Isolation, characterization, molecular cloning and modeling of a new lipid transfer protein with antiviral and antiproliferative activities from Naycissus tazetta. Peptides 2008, 29, 2101–2109. [Google Scholar] [CrossRef] [PubMed]
- Ilyas, H.; Datta, A.; Bhunia, A. An approach towards structure based antimicrobial peptide design for use in development of transgenic plants: A strategy for plant disease management. Curr. Med. Chem. 2017, 24, 1350–1364. [Google Scholar] [CrossRef]
- van Loon, L.C.; Rep, M.; Pieterse, C.M.J. Significance of inducible defense-related proteins in infected plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [PubMed]
- Parisi, K.; Shafee, T.M.A.; Quimbar, P.; van der Weerden, N.L.; Bleackley, M.R.; Anderson, M.A. The evolution, function and mechanisms of action for plant defensins. Semin. Cell Dev. Biol. 2019, 88, 107–118. [Google Scholar] [CrossRef]
- Lay, F.T.; Poon, S.; McKenna, J.A.; Connelly, A.A.; Barbeta, B.L.; McGinness, B.S.; Fox, J.L.; Daly, N.L.; Craik, D.J.; Heath, R.L.; et al. The C-terminal propeptide of a plant defensin confers cytoprotective and subcellular targeting functions. BMC Plant Biol. 2014, 14, 55–67. [Google Scholar] [CrossRef]
- van der Weerden, N.L.; Anderson, M.A. Plant defensins: Common fold, multiple functions. Fungal Biol. Rev. 2013, 26, 121–131. [Google Scholar] [CrossRef]
- Wilmes, M.; Cammue, B.P.A.; Sahl, H.G.; Thevissen, K. Antibiotic activities of host defense peptides: More to it than lipid bilayer perturbation. Cheminform 2011, 28, 1350–1358. [Google Scholar] [CrossRef]
- Schneider, T.; Kruse, T.; Wimmer, R.; Wiedemann, I.; Sass, V.; Pag, U.; Jansen, A.; Nielsen, A.K.; Mygind, P.H.; Ravents, D.S.; et al. Plectasin, a fungal defensin, targets the bacterial cell wall precursor lipid II. Science 2010, 328, 1168–1172. [Google Scholar] [CrossRef] [PubMed]
- Thevissen, K.; Franois, I.E.J.A.; Takemoto, J.Y.; Ferket, K.K.A.; Meert, E.M.K.; Cammue, B.P.A. DmAMP1, an antifungal plant defensin from dahlia (Dahlia merckii), interacts with sphingolipids from Saccharomyces cerevisiae. FEMS Microbiol. Lett. 2003, 226, 169–173. [Google Scholar] [CrossRef]
- Thevissen, K.; Ferket, K.K.A.; Franois, I.E.J.A.; Cammue, B.P.A. Interactions of antifungal plant defensins with fungal membrane components. Peptides 2003, 24, 1705–1712. [Google Scholar] [CrossRef] [PubMed]
- Payne, J.A.E.; Bleackley, M.R.; Lee, T.H.; Shafee, T.M.A.; Poon, I.K.H.; Hulett, M.D.; Aguilar, M.I.; van der Weerden, N.L.; Anderson, M.A. The plant defensin NaD1 introduces membrane disorder through a specific interaction with the lipid, phosphatidylinositol 4,5 bisphosphate. Biomembranes 2016, 33, 1099–1109. [Google Scholar] [CrossRef]
- Poon, I.K.; Baxter, A.A.; Lay, F.T.; Mills, G.D.; Hulett, M.D. Phosphoinositide-mediated oligomerization of a defensin induces cell lysis. Elife Sci. 2014, 3, e01808. [Google Scholar] [CrossRef]
- Thevissen, K.; Cammue, B.; Lemaire, K.; Winderickx, J.; Dickson, R.; Lester, R.; Ferket, K.; van Even, F.; Parret, A.; Broekaert, W. A gene encoding a sphingolipid biosynthesis enzyme determines the sensitivity of Saccharomyces cerevisiae to an antifungal plant defensin from dahlia (Dahlia merckii). Proc. Natl. Acad. Sci. USA 2000, 97, 9531–9536. [Google Scholar] [CrossRef]
- Thevissen, K. Defensins from insects and plants interact with fungal glucosylceramides. J. Biol. Chem. 2004, 279, 3900–3905. [Google Scholar] [CrossRef]
- Aerts, A.M.; Fran, I.E.J.A.; Meert, E.M.K.; Li, Q.T.; Cammue, B.P.A.; Thevissen, K. The antifungal activity of RsAFP2, a plant defensin from Raphanus sativus, involves the induction of reactive oxygen species in Candida albicans. J. Mol. Microbiol. Biotechnol. 2007, 13, 243–247. [Google Scholar] [CrossRef]
- Thevissen, K.; Tavares, P.D.M.; Xu, D.; Blankenship, J.; Vandenbosch, D.; Idkowiak-Baldys, J.; Govaert, G.; Bink, A.; Rozental, S.; Groot, P.W.J.D. The plant defensin RsAFP2 induces cell wall stress, septin mislocalization and accumulation of ceramides in Candida albicans. Mol. Microbiol. 2012, 84, 166–180. [Google Scholar] [CrossRef] [PubMed]
- Vriens, K.; Cammue, B.; Thevissen, K. Antifungal plant defensins: Mechanisms of action and production. Molecules 2014, 19, 12280–12303. [Google Scholar] [CrossRef]
- Spelbrink, R.G. Differential antifungal and calcium channel-blocking activity among structurally related plant defensins. Plant Physiol. 2004, 135, 2055–2067. [Google Scholar] [CrossRef] [PubMed]
- Ramamoorthy, V.; Cahoon, E.B.; Li, J.; Thokala, M.; Shah, D.M. Glucosylceramide synthase is essential for alfalfa defensin-mediated growth inhibition but not for pathogenicity of Fusarium graminearum. Mol. Microbiol. 2010, 66, 771–786. [Google Scholar] [CrossRef] [PubMed]
- van der Weerden, N.L.; Hancock, R.E.W.; Anderson, M.A. Permeabilization of fungal hyphae by the plant defensin NaD1 occurs through a cell wall-dependent process. J. Biol. Chem. 2010, 285, 37513–37520. [Google Scholar] [CrossRef]
- Aerts, A.M.; Leen, B.; Gilmer, G.; Didac, C.G.; Frank, M.; Cammue, B.P.A.; Karin, T. The antifungal plant defensin HsAFP1 from Heuchera sanguinea induces apoptosis in Candida albicans. Front. Microbiol. 2011, 2, 47. [Google Scholar] [CrossRef]
- Lobo, D.S.; Pereira, I.B.; Fragel-Madeira, L.; Medeiros, L.N.; Kurtenbach, E. Antifungal Pisum sativum defensin 1 interacts with Neurospora crassa cyclin F related to the cell cycle. Biochemistry 2007, 46, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Loon, L.C.V.; Pierpoint, W.S.; Boller, T.; Conejero, V. Recommendations for naming plant pathogenesis-related proteins. Plant Mol. Biol. Rep. 1994, 12, 245–264. [Google Scholar] [CrossRef]
- Loon, L.C.V.; Strien, E.A.V. The families of pathogenesis-related proteins, their activities, and comparative analysis of PR-1 type proteins. Physiol. Mol. Plant Pathol. 2002, 55, 85–97. [Google Scholar] [CrossRef]
- Sung, Y.C.; Outram, M.A.; Breen, S.; Wang, C.; Dagvadorj, B.; Winterberg, B.; Kobe, B.; Williams, S.J.; Solomon, P.S. PR1-mediated defence via C-terminal peptide release is targeted by a fungal pathogen effector. New Phytol. 2021, 229, 3467–3480. [Google Scholar] [CrossRef]
- Alexander, D.; Goodman, R.M.; Gut-Rella, M.; Glascock, C.; Weymann, K.; Friedrich, L.; Maddox, D.; Ahl-Goy, P.; Luntz, T.; Ward, E. Increased tolerance to two oomycete pathogens in transgenic tobacco expressing pathogenesis-related protein 1a. Proc. Natl. Acad. Sci. USA 1993, 90, 7327–7331. [Google Scholar] [CrossRef]
- Niderman, T.; Genetet, I.; Bruyere, T.; Gees, R.; Stintzi, A.; Legrand, M.; Fritig, B.; Mosinger, E. Pathogenesis-Related PR-1 proteins are antifungal (isolation and characterization of three 14-kilodalton proteins of tomato and of a basic PR-1 of tobacco with inhibitory activity against Phytophthora infestans). Plant Physiol. 1995, 108, 17–27. [Google Scholar] [CrossRef]
- Gamir, J.; Darwiche, R.; van‘t Hof, P.; Choudhary, V.; Stumpe, M.; Schneiter, R.; Mauch, F. The sterol-binding activity of pathogenesis-related protein 1 reveals the mode of action of an antimicrobial protein. Plant J. 2017, 89, 502–509. [Google Scholar] [CrossRef]
- Choudhary, V.; Schneiter, R. Pathogen-Related Yeast (PRY) proteins and members of the CAP superfamily are secreted sterol-binding proteins. Proc. Natl. Acad. Sci. USA 2012, 109, 16882–16887. [Google Scholar] [CrossRef]
- Lincoln, J.E.; Sanchez, J.P.; Zumstein, K.; Gilchrist, D.G. Plant and animal PR1 family members inhibit programmed cell death and suppress bacterial pathogens in plant tissues. Mol. Plant Pathol. 2018, 19, 2111–2123. [Google Scholar] [CrossRef]
- Chien, P.S.; Nam, H.G.; Chen, Y.R. A salt-regulated peptide derived from the CAP superfamily protein negatively regulates salt-stress tolerance in Arabidopsis. J. Exp. Bot. 2015, 66, 5301–5313. [Google Scholar] [CrossRef]
- Chen, Y.L.; Lee, C.Y.; Cheng, K.T.; Chang, W.H.; Huang, R.N.; Nam, H.G.; Chen, Y.R. Quantitative peptidomics study reveals that a wound-induced peptide from PR-1 regulates immune signaling in tomato. Plant Cell 2014, 26, 4135–4148. [Google Scholar] [CrossRef]
- Lu, S.; Faris, J.D.; Sherwood, R.; Friesen, T.L.; Edwards, M.C. A dimeric PR-1-type pathogenesis-related protein interacts with ToxA and potentially mediates ToxA-induced necrosis in sensitive wheat. Mol. Plant Pathol. 2014, 15, 650–663. [Google Scholar] [CrossRef] [PubMed]
- Breen, S.; Williams, S.J.; Winterberg, B.; Kobe, B.; Solomon, P.S. Wheat PR-1 proteins are targeted by necrotrophic pathogen effector proteins. Plant J. 2016, 88, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Tang, L.; Gong, Y.; Xie, J.; Fu, Y.; Jiang, D.; Li, G.; Collinge, D.B.; Chen, W.; Cheng, J. A cerato-platanin protein SsCP1 targets plant PR1 and contributes to virulence of Sclerotinia sclerotiorum. New Phytol. 2018, 217, 739–755. [Google Scholar] [CrossRef] [PubMed]
- Bohlmann, H.; Apel, K. Thionins. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1991, 42, 227–240. [Google Scholar] [CrossRef]
- Stec, B. Plant thionins-the structural perspective. Cell. Mol. Life Sci. 2006, 63, 1370–1385. [Google Scholar] [CrossRef]
- Colilla, F.J.; Rocher, A.; Mendez, E. gamma-Purothionins: Amino acid sequence of two polypeptides of a new family of thionins from wheat endosperm. FEBS Lett. 1990, 270, 191–194. [Google Scholar] [CrossRef]
- Romero, A.; Alamillo, J.M.; Garcia-Olmedo, F. Processing of thionin precursors in barley leaves by a vacuolar proteinase. Eur. J. Biochem. 1997, 243, 202–208. [Google Scholar] [CrossRef]
- Epple, P.; Apel, K.; Bohlmann, H. Overexpression of an endogenous thionin enhances resistance of Arabidopsis against Fusarium oxysporum. Plant Cell 1997, 9, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.L.; Prasad, V.; Sanjaya; Chen, K.H.; Liu, P.C.; Chan, M.T.; Chiu-Ping, C. Transgenic tomato plants expressing an Arabidopsis thionin (Thi2.1) driven by fruit-inactive promoter battle against phytopathogenic attack. Planta 2005, 221, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Carmona, M.J.; Molina, A.; Fernandez, J.A.; Lopezfando, J.J.; Garciaolmedo, F. Expression of the alpha-thionin gene from barley in tobacco confers enhanced resistance to bacterial pathogens. Plant J. 1993, 3, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Iwai, T.; Kaku, H.; Honkura, R.; Nakamura, S.; Ochiai, H.; Sasaki, T.; Ohashi, Y. Enhanced resistance to seed-transmitted bacterial diseases in transgenic rice plants overproducing an oat cell-wall-bound thionin. Mol. Plant Microbe Interact. 2002, 15, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Richard, J.A.; Kelly, I.; Marion, D.; Pezolet, M.; Auger, M. Interaction between beta-purothionin and dimyristoylphosphatidylglycerol: A P-31-NMR and infrared spectroscopic study. Biophys. J. 2002, 83, 2074–2083. [Google Scholar] [CrossRef]
- Thevissen, K.; Terras, F.R.G.; Broekaert, W.F. Permeabilization of fungal membranes by plant defensins inhibits fungal growth. Appl. Environ. Microbiol. 1999, 65, 5451–5458. [Google Scholar] [CrossRef]
- Molina, A.; Segura, A.; Garciaolmedo, F. Lipid transfer proteins (NSLTPS) from barley and maize leaves are potent inhibitors of bacterial and fungal plant-pathogens. FEBS Lett. 1993, 316, 119–122. [Google Scholar] [CrossRef]
- Evans, J.; Wang, Y.D.; Shaw, K.P.; Vernon, L.P. Cellular responses to Pyrularia thionin are mediated by Ca2+ influx and phospholipase A2 activation and are inhibited by thionin tyrosine iodination. Proc. Natl. Acad. Sci. USA 1989, 86, 5849–5853. [Google Scholar] [CrossRef]
- Stein, G.M.; Pfuller, U.; Schietzel, M.; Bussing, A. Toxic proteins from European mistletoe (Viscum album L.: Increase of intracellular IL-4 but decrease of IFN-gamma in apoptotic cells. Anticancer Res. 2000, 20, 1673–1678. [Google Scholar]
- Richard, J.A.; Kelly, I.; Marion, D.; Auger, M.; Pezolet, M. Structure of beta-purothionin in membranes: A two-dimensional infrared correlation spectroscopy study. Biochemistry 2005, 44, 52–61. [Google Scholar] [CrossRef]
- Salminen, T.A.; Blomqvist, K.; Edqvist, J. Lipid transfer proteins: Classification, nomenclature, structure, and function. Planta 2016, 244, 971–997. [Google Scholar] [CrossRef]
- Castro, M.S.; Fontes, W. Plant defense and antimicrobial peptides. Protein Pept. Lett. 2005, 12, 13–18. [Google Scholar] [CrossRef]
- Berecz, B.; Mills, E.N.C.; Tamas, L.; Lang, F.; Shewry, P.R.; Mackie, A.R. Structural stability and surface activity of sunflower 2s albumins and nonspecific lipid transfer protein. J. Agric. Food Chem. 2010, 58, 6490–6497. [Google Scholar] [CrossRef]
- Edqvist, J.; Blomqvist, K.; Nieuwland, J.; Salminen, T.A. Plant lipid transfer proteins: Are we finally closing in on the roles of these enigmatic proteins? J. Lipid Res. 2018, 59, 1374–1382. [Google Scholar] [CrossRef]
- Carvalho, A.d.O.; Gomes, V.M. Role of plant lipid transfer proteins in plant cell physiology—A concise review. Peptides 2007, 28, 1144–1153. [Google Scholar] [CrossRef]
- Nieuwland, J.; Feron, R.; Huisman, B.A.H.; Fasolino, A.; Hilbers, C.W.; Derksen, J.; Mariani, C. Lipid transfer proteins enhance cell wall extension in tobacco. Plant Cell 2005, 17, 2009–2019. [Google Scholar] [CrossRef]
- Tsuboi, S.; Osafune, T.; Tsugeki, R.; Nishimura, M.; Yamada, M. Nonspecific lipid transfer protein in castor bean cotyledon cells subcellular localization and a possible role in lipid-metabolism. J. Biochem. 1992, 111, 500–508. [Google Scholar] [CrossRef]
- Jacq, A.; Pernot, C.; Martinez, Y.; Domergue, F.; Payre, B.; Jamet, E.; Burlat, V.; Pacquit, V.B. The Arabidopsis lipid transfer protein 2 (AtLTP2) is involved in cuticle-cell wall interface integrity and in etiolated hypocotyl permeability. Front. Plant Sci. 2017, 8, 263. [Google Scholar] [CrossRef] [PubMed]
- Deeken, R.; Saupe, S.; Klinkenberg, J.; Riedel, M.; Leide, J.; Hedrich, R.; Mueller, T.D. The nonspecific lipid transfer protein AtLtpI-4 is involved in suberin formation of Arabidopsis thaliana crown galls. Plant Physiol. 2016, 172, 1911–1927. [Google Scholar] [CrossRef] [PubMed]
- DeBono, A.; Yeats, T.H.; Rose, J.K.C.; Bird, D.; Jetter, R.; Kunst, L.; Samuelsa, L. Arabidopsis LTPG Is a glycosylphosphatidylinositol-anchored lipid transfer protein required for export of lipids to the plant surface. Plant Cell 2009, 21, 1230–1238. [Google Scholar] [CrossRef] [PubMed]
- Chassot, C.; Nawrath, C.; Metraux, J.P. Cuticular defects lead to full immunity to a major plant pathogen. Plant J. 2007, 49, 972–980. [Google Scholar] [CrossRef]
- Molina, A.; GarciaOlmedo, F. Enhanced tolerance to bacterial pathogens caused by the transgenic expression of barley lipid transfer protein LTP2. Plant J. 1997, 12, 669–675. [Google Scholar] [CrossRef]
- Diz, M.S.; Carvalho, A.O.; Ribeiro, S.F.F.; Da Cunha, M.; Beltramini, L.; Rodrigues, R.; Nascimento, V.V.; Machado, O.L.T.; Gomes, V.M. Characterisation, immunolocalisation and antifungal activity of a lipid transfer protein from chili pepper (Capsicum annuum) seeds with novel alpha-amylase inhibitory properties. Physiol. Plant. 2011, 142, 233–246. [Google Scholar] [CrossRef]
- Gonorazky, A.G.; Regente, M.C.; de la Canal, L. Stress induction and antimicrobial properties of a lipid transfer protein in germinating sunflower seeds. J. Plant Physiol. 2005, 162, 618–624. [Google Scholar] [CrossRef]
- Cammue Bruno, P.A.; Thevissen, K.; Hendriks, M.; Eggermont, K.; Goderis, I.J.; Proost, P.; Vandamme, J.; Osborn, R.W.; Guerbette, F.; Kader, J.C.; et al. A potent antimicrobial protein from onion seeds showing sequence homology to plant lipid transfer proteins. Plant Physiol. 1995, 109, 445–455. [Google Scholar] [CrossRef]
- Terras, F.R.; Goderis, I.J.; Van Leuven, F.; Vanderleyden, J.; Cammue, B.P.; Broekaert, W.F. In vitro antifungal activity of a radish (Raphanus sativus L.) seed protein homologous to nonspecific lipid transfer proteins. Plant Physiol. 1992, 100, 1055–1058. [Google Scholar] [CrossRef]
- Regente, M.C.; Giudici, A.M.; Villalain, J.; de la Canal, L. The cytotoxic properties of a plant lipid transfer protein involve membrane permeabilization of target cells. Lett. Appl. Microbiol. 2005, 40, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Kader, J.C. Lipid-transfer proteins in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 627–654. [Google Scholar] [CrossRef] [PubMed]
- Glazebrook, J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef]
- Sels, J.; Mathys, J.; De Coninck, B.M.A.; Cammue, B.P.A.; de Bolle, M.F.C. Plant pathogenesis-related (PR) proteins: A focus on PR peptides. Plant Physiol. Biochem. 2008, 46, 941–950. [Google Scholar] [CrossRef]
- Christeller, J.; Laing, W. Plant serine proteinase inhibitors. Protein Pept. Lett. 2005, 12, 439–447. [Google Scholar] [CrossRef]
- Haq, S.K.; Atif, S.M.; Khan, R.H. Protein proteinase inhibitor genes in combat against insects, pests, and pathogens: Natural and engineered phytoprotection. Arch. Biochem. Biophys. 2004, 431, 145–159. [Google Scholar] [CrossRef] [PubMed]
- Valueva, T.A.; Mosolov, V.V. Role of inhibitors of proteolytic enzymes in plant defense against phytopathogenic microorganisms. Biochem. Mosc. 2004, 69, 1305–1309. [Google Scholar] [CrossRef]
- Dunaevskii, Y.E.; Tsybina, T.A.; Belyakova, G.A.; Domash, V.I.; Sharpio, T.P.; Zabreiko, S.A.; Belozerskii, M.A. Proteinase inhibitors as antistress proteins in higher plants. Appl. Biochem. Microbiol. 2005, 41, 344–348. [Google Scholar] [CrossRef]
- Gutierrez-Campos, R.; Torres-Acosta, J.A.; Saucedo-Arias, L.J.; Gomez-Lim, M.A. The use of cysteine proteinase inhibitors to engineer resistance against potyviruses in transgenic tobacco plants. Nat. Biotechnol. 1999, 17, 1223–1226. [Google Scholar] [CrossRef]
- Vila, L.; Quilis, J.; Meynard, D.; Breitler, J.C.; Marfa, V.; Murillo, I.; Vassal, J.M.; Messeguer, J.; Guiderdoni, E.; San Segundo, B. Expression of the maize proteinase inhibitor (mpi) gene in rice plants enhances resistance against the striped stem borer (Chilo suppressalis): Effects on larval growth and insect gut proteinases. Plant Biotechnol. J. 2005, 3, 187–202. [Google Scholar] [CrossRef]
- Peng, J.H.; Black, L.L. Increased proteinase-inhibitor activity in response to infection of resistant tomato plants by Phytophthora infestans. Phytopathology 1976, 66, 958–963. [Google Scholar]
- Pautot, V.; Holzer, F.M.; Walling, L.L. Differential expression of tomato proteinase inhibitor-i and inhibitor-ii genes during bacterial pathogen invasion and wounding. Mol. Plant Microbe Interact. 1991, 4, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Terras, F.R.G.; Schoofs, H.M.E.; Thevissen, K.; Osborn, R.W.; Vanderleyden, J.; Cammue, B.P.A.; Broekaert, W.F. Synergistic enhancement of the antifungal activity of wheat and barley thionins by radish and oilseed rape-2S albumins and by barley trypsin-inhibitors. Plant Physiol. 1993, 103, 1311–1319. [Google Scholar] [CrossRef] [PubMed]
- Vieira Bard, G.C.; Nascimento, V.V.; Ribeiro, S.F.F.; Rodrigues, R.; Perales, J.; Teixeira-Ferreira, A.; Carvalho, A.O.; Fernandes, K.V.S.; Gomes, V.M. Characterization of peptides from Capsicum annuum hybrid seeds with inhibitory activity against α-amylase, serine proteinases and fungi. Protein J. 2015, 34, 122–129. [Google Scholar] [CrossRef] [PubMed]
- ten Have, A.; Dekkers, E.; Kay, J.; Phylip, L.H.; van Kan, J.A.L. An aspartic proteinase gene family in the filamentous fungus Botrytis cinerea contains members with novel features. Microbiology 2004, 150, 2475–2489. [Google Scholar] [CrossRef] [PubMed]
- Lorito, M.; Peterbauer, T.C.; Hayes, C.K.; Harman, G.E. Synergistic interaction between fungal cell-wall degrading enzymes and different antifungal compounds enhances inhibition of spore germination. Microbiology 1994, 140, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Machida, S.; Saito, M. Purification and characterization of membrane-bound chitin synthase. J. Biol. Chem. 1993, 268, 1702–1707. [Google Scholar] [CrossRef]
- Ramamoorthy, V.; Zhao, X.; Snyder, A.K.; Xu, J.; Shah, D.M. Two mitogen-activated protein kinase signalling cascades mediate basal resistance to antifungal plant defensins in Fusarium graminearum. Cell. Microbiol. 2010, 9, 1491–1506. [Google Scholar] [CrossRef] [PubMed]
- Rao, U.; Stec, B.; Teeter, M.M. Refinement of purothionins reveals solute particles important for lattice formation and toxicity. Part 1: Alpha1-purothionin revisited. Acta Crystallogr. Sect. D 1995, 51, 904–913. [Google Scholar] [CrossRef]
- Giudici, A.M.; Regente, M.C.; Villalain, J.; Pfuller, K.; Pfuller, U.; de la Canal, L. Mistletoe viscotoxins induce membrane permeabilization and spore death in phytopathogenic fungi. Physiol. Plant. 2004, 121, 2–7. [Google Scholar] [CrossRef]
- Fisher, M.C.; Henk, D.A.; Briggs, C.J.; Brownstein, J.S.; Madoff, L.C.; McCraw, S.L.; Gurr, S.J. Emerging fungal threats to animal, plant and ecosystem health. Nature 2012, 484, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Shah, P.; Hsiao, F.S.H.; Ho, Y.H.; Chen, C.S. The proteome targets of intracellular targeting antimicrobial peptides. Proteomics 2016, 16, 1225–1237. [Google Scholar] [CrossRef] [PubMed]
- Jarva, M.; Lay, F.T.; Thanh Kha, P.; Humble, C.; Poon, I.K.H.; Bleackley, M.R.; Anderson, M.A.; Hulett, M.D.; Kvansakul, M. X-ray structure of a carpet-like antimicrobial defensin-phospholipid membrane disruption complex. Nat. Commun. 2018, 9, 1962. [Google Scholar] [CrossRef] [PubMed]
- Saito, K.; Takakuwa, N.; Ohnishi, M.; Oda, Y. Presence of glucosylceramide in yeast and its relation to alkali tolerance of yeast. Appl. Microbiol. Biotechnol. 2006, 71, 515–521. [Google Scholar] [CrossRef]
- Tavares, P.M.; Thevissen, K.; Cammue, B.P.A.; Francois, I.E.J.A.; Barreto-Bergter, E.; Taborda, C.P.; Marques, A.F.; Rodrigues, M.L.; Nimrichter, L. In vitro activity of the antifungal plant defensin RsAFP2 against Candida isolates and its in vivo efficacy in prophylactic murine models of candidiasis. Antimicrob. Agents Chemother. 2008, 52, 4522–4525. [Google Scholar] [CrossRef]
- Noble, S.M.; French, S.; Kohn, L.A.; Chen, V.; Johnson, A.D. Systematic screens of a Candida albicans homozygous deletion library decouple morphogenetic switching and pathogenicity. Nat. Genet. 2010, 42, 590–598. [Google Scholar] [CrossRef]
- Garcia-Rubio, R.; de Oliveira, H.C.; Rivera, J.; Trevijano-Contador, N. The fungal cell wall: Candida, Cryptococcus, and Aspergillus species. Front. Microbiol. 2020, 10, 2993. [Google Scholar] [CrossRef]
- Rodrigues, M.L.; Travassos, L.R.; Miranda, K.R.; Franzen, A.J.; Rozental, S.; de Souza, W.; Alviano, C.S.; Barreto-Bergter, E. Human antibodies against a purified glucosylceramide from Cryptococcus neoformans inhibit cell budding and fungal growth. Infect. Immun. 2000, 68, 7049–7060. [Google Scholar] [CrossRef]
- Koo, J.C.; Lee, B.; Young, M.E.; Koo, S.C.; Cooper, J.A.; Baek, D.; Lim, C.O.; Lee, S.Y.; Yun, D.J.; Cho, M.J. Pn-AMP1, a plant defense protein, induces actin depolarization in yeasts. Plant Cell Physiol. 2004, 45, 1669–1680. [Google Scholar] [CrossRef]
- Tsai, P.W.; Yang, C.Y.; Chang, H.T.; Lan, C.Y. Human antimicrobial peptide LL-37 inhibits adhesion of Candida albicans by interacting with yeast cell-wall carbohydrates. PLoS ONE 2011, 6, e17755. [Google Scholar] [CrossRef]
- Li, H.; Velivelli, S.L.S.; Shah, D.M. Antifungal potency and modes of action of a novel olive tree defensin against closely related ascomycete fungal pathogens. Mol. Plant Microbe Interact. 2019, 32, 1649–1664. [Google Scholar] [CrossRef]
- Combest, M.M.; Moroz, N.; Tanaka, K.; Rogan, C.J.; Anderson, J.C.; Thura, L.; Rakotondrafara, A.M.; Goyer, A. StPIP1, a PAMP-induced peptide in potato, elicits plant defenses and is associated with disease symptom severity in a compatible interaction with Potato virus Y. J. Exp. Bot. 2021, 72, 4472–4488. [Google Scholar] [CrossRef] [PubMed]
- Reyes, M.I.; Nash, T.E.; Dallas, M.M.; Ascencio-Ibanez, J.T.; Hanley-Bowdoin, L. Peptide aptamers that bind to geminivirus replication proteins confer a resistance phenotype to tomato yellow leaf curl virus and tomato mottle virus infection in tomato. J. Virol. 2013, 87, 9691–9706. [Google Scholar] [CrossRef] [PubMed]
- Mendoza-Figueroa, J.S.; Kvarnheden, A.; Mendez-Lozano, J.; Rodriguez-Negrete, E.A.; Arreguin-Espinosa de los Monteros, R.; Soriano-Garcia, M. A peptide derived from enzymatic digestion of globulins from amaranth shows strong affinity binding to the replication origin of Tomato yellow leaf curl virus reducing viral replication in Nicotiana benthamiana. Pestic. Biochem. Physiol. 2018, 145, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Rudolph, C.; Schreier, P.H.; Uhrig, J.F. Peptide-mediated broad-spectrum plant resistance to tospoviruses. Proc. Natl. Acad. Sci. USA 2003, 100, 4429–4434. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.X.; Ng, T.B. Ginkbilobin, a novel antifungal protein from Ginkgo biloba seeds with sequence similarity to embryo-abundant protein. Biochem. Biophys. Res. Commun. 2000, 279, 407–411. [Google Scholar] [CrossRef]
- Wang, H.X.; Ng, T.B. Ascalin, a new anti-fungal peptide with human immunodeficiency virus type 1 reverse transcriptase-inhibiting activity from shallot bulbs. Peptides 2002, 23, 1025–1029. [Google Scholar] [CrossRef]
- Wong, J.H.; Ng, T.B. Lunatusin, a trypsin-stable antimicrobial peptide from lima beans (Phaseolus lunatus L.). Peptides 2005, 26, 2086–2092. [Google Scholar] [CrossRef]
- Wong, J.H.; Ng, T.B. Vulgarinin, a broad-spectrum antifungal peptide from haricot beans (Phaseolus vulgaris). Int. J. Biochem. Cell Biol. 2005, 37, 1626–1632. [Google Scholar] [CrossRef]
- Wachsman, M.B.; Castilla, V.; Coto, C.E. Inhibition of foot and mouth disease virus (FMDV) uncoating by a plant-derived peptide isolated from Melia azedarach L leaves. Arch. Virol. 1998, 143, 581–590. [Google Scholar] [CrossRef]
- Panya, A.; Yongpitakwattana, P.; Budchart, P.; Sawasdee, N.; Krobthong, S.; Paemanee, A.; Roytrakul, S.; Rattanabunyong, S.; Choowongkomon, K.; Yenchitsomanus, P.T. Novel bioactive peptides demonstrating anti-dengue virus activity isolated from the Asian medicinal plant Acacia Catechu. Chem. Biol. Drug Des. 2019, 93, 100–109. [Google Scholar] [CrossRef]
- Panya, A.; Sawasdee, N.; Songprakhon, P.; Tragoolpua, Y.; Rotarayanont, S.; Choowongkomon, K.; Yenchitsomanus, P.T. A synthetic bioactive peptide derived from the asian medicinal plant Acacia Catechu binds to dengue virus and inhibits cell entry. Viruses Basel 2020, 12, 1267. [Google Scholar] [CrossRef]
- Yamaji, Y.; Maejima, K.; Komatsu, K.; Shiraishi, T.; Okano, Y.; Himeno, M.; Sugawara, K.; Neriya, Y.; Minato, N.; Miura, C.; et al. Lectin-mediated resistance impairs plant virus infection at the cellular level. Plant Cell 2012, 24, 778–793. [Google Scholar] [CrossRef]
- Yoshida, T.; Shiraishi, T.; Hagiwara-Komoda, Y.; Komatsu, K.; Maejima, K.; Okano, Y.; Fujimoto, Y.; Yusa, A.; Yamaji, Y.; Namba, S. The Plant noncanonical antiviral resistance protein JAX1 inhibits potexviral replication by targeting the viral RNA-dependent RNA polymerase. J. Virol. 2019, 93, e01506-18. [Google Scholar] [CrossRef]
- Li, F.; Zhang, C.; Li, Y.; Wu, G.; Hou, X.; Zhou, X.; Wang, A. Beclin1 restricts RNA virus infection in plants through suppression and degradation of the viral polymerase. Nat. Commun. 2018, 9, 1268. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.Y.; Mendu, V.; Pogany, J.; Qin, J.; Nagy, P.D. The TPR domain in the host Cyp40-like cyclophilin binds to the viral replication protein and inhibits the assembly of the tombusviral replicase. PLoS Pathog. 2012, 8, e1002491. [Google Scholar] [CrossRef] [PubMed]
- Mookherjee, N.; Anderson, M.A.; Haagsman, H.P.; Davidson, D.J. Antimicrobial host defence peptides: Functions and clinical potential. Nat. Rev. Drug Discov. 2020, 19, 311–332. [Google Scholar] [CrossRef] [PubMed]
- Kota, S.; Sabbah, A.; Chang, T.H.; Harnack, R.; Xiang, Y.; Meng, X.; Bose, S. Role of human beta-defensin-2 during tumor necrosis factor-alpha/NF-kappa B- mediated innate antiviral response against human respiratory syncytial virus. J. Biol. Chem. 2008, 283, 22417–22429. [Google Scholar] [CrossRef]
- Robinson, W.E.; McDougall, B.; Tran, D.; Selsted, M.E. Anti-HIV-1 activity of indolicidin, an antimicrobial peptide from neutrophils. J. Leukoc. Biol. 1998, 63, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.G.; Nemerow, G.R. Mechanism of adenovirus neutralization by human alpha-defensins. Cell Host Microbe 2008, 3, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Lehrer, R.I.; Jung, G.; Ruchala, P.; Andre, S.; Gabius, H.J.; Lu, W. Multivalent binding of carbohydrates by the human alpha-defensin, HD5. J. Immunol. 2009, 183, 480–490. [Google Scholar] [CrossRef]
- Wang, W.; Owen, S.M.; Rudolph, D.L.; Cole, A.M.; Hong, T.; Waring, A.J.; Lal, R.B.; Lehrer, R.I. Activity of alpha- and theta-defensins against primary isolates of HIV-1. J. Immunol. 2004, 173, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Song, B.H.; Lee, G.C.; Moon, M.S.; Cho, Y.H.; Lee, C.H. Human cytomegalovirus binding to heparan sulfate proteoglycans on the cell surface and/or entry stimulates the expression of human leukocyte antigen class I. J. Gen. Virol. 2001, 82, 2405–2413. [Google Scholar] [CrossRef]
- Yasin, B.; Wang, W.; Pang, M.; Cheshenko, N.; Hong, T.; Waring, A.J.; Herold, B.C.; Wagar, E.A.; Lehrer, R.I. Theta defensins protect cells from infection by herpes simplex virus by inhibiting viral adhesion and entry. J. Virol. 2004, 78, 5147–5156. [Google Scholar] [CrossRef] [PubMed]
- Andersen, J.H.; Jenssen, H.; Sandvik, K.; Gutteberg, T.J. Anti-HSV activity of lactoferrin and lactoferricin is dependent on the presence of heparan sulphate at the cell surface. J. Med. Virol. 2004, 74, 262–271. [Google Scholar] [CrossRef]
- Sousa, F.H.; Casanova, V.; Findlay, F.; Stevens, C.; Svoboda, P.; Pohl, J.; Proudfoot, L.; Barlow, P.G. Cathelicidins display conserved direct antiviral activity towards rhinovirus. Peptides 2017, 95, 76–83. [Google Scholar] [CrossRef] [PubMed]
- de Lumen, B.O. Lunasin: A cancer-preventive soy peptide. Nutr. Rev. 2005, 63, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.J.; Lam, Y.; de Lumen, B.O. Barley lunasin suppresses ras-induced colony formation and inhibits core histone acetylation in mammalian cells. J. Agric. Food Chem. 2002, 50, 5903–5908. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.J.; Jin, B.J.; Kim, D.S.; Park, J.H.; Lee, J.B.; Kweon, D.H.; Chung, G.Y.; Seo, E.W.; Lumen, B. The cancer preventive peptide lunasin from wheat inhibits core histone acetylation. Cancer Lett. 2007, 255, 42–48. [Google Scholar] [CrossRef]
- Gerlach, S.L.; Rathinakumar, R.; Chakravarty, G.; Goransson, U.; Wimley, W.C.; Darwin, S.P.; Mondal, D. Anticancer and Chemosensitizing Abilities of cycloviolacin O2 from Viola odorata and psyle cyclotides from Psychotria leptothyrsa. Biopolymers 2010, 94, 617–625. [Google Scholar] [CrossRef]
- Ji, Y.; Majumder, S.; Millard, M.; Borra, R.; Bi, T.; Elnagar, A.Y.; Neamati, N.; Shekhtman, A.; Camarero, J.A. In vivo activation of the p53 tumor suppressor pathway by an engineered cyclotide. J. Am. Chem. Soc. 2013, 135, 11623–11633. [Google Scholar] [CrossRef]
- Ding, X.; Bai, D.; Qian, J. Novel cyclotides from Hedyotis biflora inhibit proliferation and migration of pancreatic cancer cell in vitro and in vivo. Med. Chem. Res. 2014, 23, 1406–1413. [Google Scholar] [CrossRef]
- Bleackley, M.R.; Payne, J.A.E.; Hayes, B.M.E.; Durek, T.; Craik, D.J.; Shafee, T.M.A.; Poon, I.K.H.; Hulett, M.D.; van der Weerden, N.L.; Anderson, M.A. Nicotiana alata defensin chimeras reveal differences in the mechanism of fungal and tumor cell killing and an enhanced antifungal variant. Antimicrob. Agents Chemother. 2016, 60, 6302–6312. [Google Scholar] [CrossRef]
- Kong, J.L.; Du, X.B.; Fan, C.X.; Xu, J.F.; Zheng, X.J. Determination of primary structure of a novel peptide from mistletoe and its antitumor activity. Yao Xue Xue Bao Acta Pharm. Sin. 2004, 39, 813–817. [Google Scholar]
- Li, S.S.; Gullbo, J.; Lindholm, P.; Larsson, R.; Thunberg, E.; Samuelsson, G.; Bohlin, L.; Claeson, P. Ligatoxin B, a new cytotoxic protein with a novel helix-turn-helix DNA-binding domain from the mistletoe Phoradendron liga. Biochem. J. 2002, 366, 405–413. [Google Scholar] [CrossRef]
- van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: Where they are and how they behave. Nat. Rev. Mol. Cell Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Johansson, S.; Gullbo, J.; Lindholm, P.; Ek, B.; Thunberg, E.; Samuelsson, G.; Larsson, R.; Bohlin, L.; Claeson, P. Small, novel proteins from the mistletoe Phoradendron tomentosum exhibit highly, selective cytotoxicity to human breast cancer cells. Cell. Mol. Life Sci. 2003, 60, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Loeza-Angeles, H.; Sagrero-Cisneros, E.; Lara-Zarate, L.; Villagomez-Gomez, E.; Lopez-Meza, J.E.; Ochoa-Zarzosa, A. Thionin Thi2.1 from Arabidopsis thaliana expressed in endothelial cells shows antibacterial, antifungal and cytotoxic activity. Biotechnol. Lett. 2008, 30, 1713–1719. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.H.; Ng, T.B. Sesquin, a potent defensin-like antimicrobial peptide from ground beans with inhibitory activities toward tumor cells and HIV-1 reverse transcriptase. Peptides 2005, 26, 1120–1126. [Google Scholar] [CrossRef]
- Wong, J.H.; Ng, T.B. Limenin, a defensin-like peptide with multiple exploitable activities from shelf beans. J. Pept. Sci. 2006, 12, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Ngai, P.H.K.; Ng, T.B. Coccinin, an antifungal peptide with antiproliferative and HIV-1 reverse transcriptase inhibitory activities from large scarlet runner beans. Peptides 2004, 25, 2063–2068. [Google Scholar] [CrossRef]
- Silva, O.N.; Porto, W.F.; Migliolo, L.; Mandal, S.M.; Gomes, D.G.; Holanda, H.H.S.; Silva, R.S.P.; Dias, S.C.; Costa, M.P.; Costa, C.R.; et al. Cn-AMP1: A new promiscuous peptide with potential for microbial infections treatment. Biopolymers 2012, 98, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.M.; Migliolo, L.; Das, S.; Mandal, M.; Franco, O.L.; Hazra, T.K. Identification and characterization of a bactericidal and proapoptotic peptide from cycas revoluta seeds with DNA binding properties. J. Cell. Biochem. 2012, 113, 184–193. [Google Scholar] [CrossRef]
- Wele, A.; Zhang, Y.J.; Ndoye, I.; Brouard, J.P.; Pousset, J.L.; Bodo, B. A cytotoxic cyclic heptapeptide from the seeds of Annona cherimola. J. Nat. Prod. 2004, 67, 1577–1579. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Gauri, S.S.; Mukhopadhyay, S.K.; Chatterjee, S.; Das, S.S.; Mandal, S.M.; Dey, S. Identification and structural characterization of a new pro-apoptotic cyclic octapeptide cyclosaplin from somatic seedlings of Santalum album L. Peptides 2014, 54, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Pinto, M.E.F.; Najas, J.Z.G.; Magalhaes, L.G.; Bobey, A.F.; Mendonca, J.N.; Lopes, N.P.; Leme, F.M.; Teixeira, S.P.; Trovo, M.; Andricopulo, A.D.; et al. Inhibition of breast cancer cell migration by cyclotides isolated from Pombalia calceolaria. J. Nat. Prod. 2018, 81, 1203–1208. [Google Scholar] [CrossRef]
- Chang, V.H.S.; Yang, D.H.A.; Lin, H.-H.; Pearce, G.; Ryan, C.A.; Chen, Y.C. IbACP, a sixteen-amino-acid peptide isolated from Ipomoea batatas leaves, induces carcinoma cell apoptosis. Peptides 2013, 47, 148–156. [Google Scholar] [CrossRef]
- Gaspar, D.; Veiga, A.S.; Castanho, M.R.B. From antimicrobial to anticancer peptides. A review. Front. Microbiol. 2013, 4. [Google Scholar] [CrossRef]
- Schweizer, F. Cationic amphiphilic peptides with cancer-selective toxicity. Eur. J. Pharmacol. 2009, 625, 190–194. [Google Scholar] [CrossRef]
- Wang, L.; Dong, C.; Li, X.; Han, W.; Su, X. Anticancer potential of bioactive peptides from animal sources (Review). Oncol. Rep. 2017, 38, 637–651. [Google Scholar] [CrossRef]
- Feire, A.L.; Koss, H.; Compton, T. Cellular integrins function as entry receptors for human cytomegalovirus via a highly conserved disintegrin-like domain. Proc. Natl. Acad. Sci. USA 2004, 101, 15470–15475. [Google Scholar] [CrossRef]
- Lin, P.; Wong, J.H.; Ng, T.B. A defensin with highly potent antipathogenic activities from the seeds of purple pole bean. Biosci. Rep. 2010, 30, 101–109. [Google Scholar] [CrossRef]
- Luna-Vital, D.A.; Liang, K.; Gonzalez de Mejia, E.; Loarca-Pina, G. Dietary peptides from the non-digestible fraction of Phaseolus vulgaris L. decrease angiotensin II-dependent proliferation in HCT116 human colorectal cancer cells through the blockade of the renin-angiotensin system. Food Funct. 2016, 7, 2409–2419. [Google Scholar] [CrossRef]
- Iqbal, A.; Khan, R.S.; Shehryar, K.; Imran, A.; Ali, F.; Attia, S.; Shah, S.; Mii, M. Antimicrobial peptides as effective tools for enhanced disease resistance in plants. Plant Cell Tissue Organ Cult. 2019, 139, 1–15. [Google Scholar] [CrossRef]
- Khan, R.S.; Iqbal, A.; Malak, R.; Shehryar, K.; Attia, S.; Ahmed, T.; Khan, M.A.; Arif, M.; Mii, M. Plant defensins: Types, mechanism of action and prospects of genetic engineering for enhanced disease resistance in plants. 3 Biotech 2019, 9, 192. [Google Scholar] [CrossRef]
- Lee, O.S.; Lee, B.; Park, N.; Koo, J.C.; Kim, Y.H.; Prasad, T.; Karigar, C.; Chun, H.J.; Jeong, B.R.; Kim, D.H.; et al. Pn-AMPs, the hevein-like proteins from Pharbitis nil confers disease resistance against phytopathogenic fungi in tomato, Lycopersicum esculentum. Phytochemistry 2003, 62, 1073–1079. [Google Scholar] [CrossRef]
- Jung, H.W.; Kim, K.D.; Hwang, B.K. Identification of pathogen-responsive regions in the promoter of a pepper lipid transfer protein gene (CALTPI) and the enhanced resistance of the CALTPI transgenic Arabidopsis against pathogen and environmental stresses. Planta 2005, 221, 361–373. [Google Scholar] [CrossRef]
- Jha, S.; Chattoo, B.B. Transgene stacking and coordinated expression of plant defensins confer fungal resistance in rice. Rice 2009, 2, 143–154. [Google Scholar] [CrossRef][Green Version]
- Coca, M.; Peñas, G.; Gómez, J.; Campo, S.; Bortolotti, C.; Messeguer, J.; Segundo, B.S. Enhanced resistance to the rice blast fungus Magnaporthe grisea conferred by expression of a cecropin A gene in transgenic rice. Planta 2006, 223, 392–406. [Google Scholar] [CrossRef] [PubMed]
- Bundó, M.; Montesinos, L.; Izquierdo, E.; Campo, S.; Mieulet, D.; Guiderdoni, E.; Rossignol, M.; Badosa, E.; Montesinos, E.; Segundo, B.S.; et al. Production of cecropin A antimicrobial peptide in rice seed endosperm. BMC Plant Biol. 2014, 14, 102. [Google Scholar] [CrossRef] [PubMed]
- Jan, P.S.; Huang, H.Y.; Chen, H.M. Expression of a synthesized gene encoding cationic peptide cecropin B in transgenic tomato plants protects against bacterial diseases. Appllied Environ. Microbiol. 2010, 76, 769–775. [Google Scholar] [CrossRef] [PubMed]
- Company, N.; Nadal, A.; La Paz, J.L.; Martínez, S.; Rasche, S.; Schillberg, S.; Pla, M. The production of recombinant cationic α-helical antimicrobial peptides in plant cells induces the formation of protein bodies derived from the endoplasmic reticulum. Plant Biotechnol. J. 2014, 12, 81–92. [Google Scholar] [CrossRef]
- Nadal, A.; Montero, M.; Company, N.; Badosa, E.; Messeguer, J.; Montesinos, L.; Montesinos, E.; Pla, M. Constitutive expression of transgenes encoding derivatives of the synthetic antimicrobial peptide BP100: Impact on rice host plant fitness. BMC Plant Biol. 2012, 12, 159. [Google Scholar] [CrossRef]
- Nuti, R.; Goud, N.S.; Saraswati, A.P.; Alvala, R.; Alvala, M. Antimicrobial peptides: A promising therapeutic strategy in tackling antimicrobial resistance. Curr. Med. Chem. 2017, 24, 4303–4314. [Google Scholar] [CrossRef]
- Greber, K.E.; Dawgul, M. Antimicrobial peptides under clinical trials. Curr. Top. Med. Chem. 2017, 17, 620–628. [Google Scholar] [CrossRef]
- Holaskova, E.; Galuszka, P.; Frebort, I.; Oz, M.T. Antimicrobial peptide production and plant-based expression systems for medical and agricultural biotechnology. Biotechnol. Adv. 2015, 33, 1005–1023. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.B.; Li, B.; Jin, S.; Daniell, H. Expression and characterization of antimicrobial peptides Retrocyclin-101 and Protegrin-1 in chloroplasts to control viral and bacterial infections. Plant Biotechnol. J. 2011, 9, 100–115. [Google Scholar] [CrossRef] [PubMed]
- Cabanos, C.; Ekyo, A.; Amari, Y.; Kato, N.; Kuroda, M.; Nagaoka, S.; Takaiwa, F.; Utsumi, S.; Maruyama, N. High-level production of lactostatin, a hypocholesterolemic peptide, in transgenic rice using soybean A1aB1b as carrier. Transgenic Res. 2013, 22, 621–629. [Google Scholar] [CrossRef] [PubMed]
- de Veer, S.J.; Kan, M.W.; Craik, D.J. Cyclotides: From structure to function. Chem. Rev. 2019, 119, 12375–12421. [Google Scholar] [CrossRef]
- Vlieghe, P.; Lisowski, V.; Martinez, J.; Khrestchatisky, M. Synthetic therapeutic peptides: Science and market. Drug Discov. Today 2010, 15, 40–56. [Google Scholar] [CrossRef]



| Source of Virus | Representative Peptide | Antiviral Activity | Mode of Action | References |
|---|---|---|---|---|
| Plant virus | StPIP1 | Potato Y virus (PVY) | StPIP1 induces the ROS production, callose deposition, and expression of defense-related genes in plants. | [112] |
| A22 and A64 | Tomato Golden Mosaic virus (TGMV) | Peptides interfere with virus replication by binding to the replication origin sequence (OriRep). | [113] | |
| AmPep1 | Tomato yellow leaf curl virus (TYLCV) Pepper yellow vein virus (PHYVV) | [114] | ||
| Dominant transacting peptide | Tomato chlorotic spot virus (TCSV) Groundnut ring spot virus (GRSV) Chrysanthemum stem necrotic virus (CSNV) Impatiens necrotic spot virus (INSV) Iris macular spot virus (IYSV) Physalis severe mottle virus (PSMV) Watermelon silver mottle virus (WSMV) | The peptide interacts with the nucleocapsid proteins (N) of different tospoviruses and induces host immune responses. | [115] | |
| Animal virus | NTP | Influenza A virus (H1N1) | NTP inhibits virus proliferation by blocking the neuramidase on the virus envelope, and inhibits the cytopathic effect induced by H1N1. | [16] |
| Respiratory syncytial virus (RSV) | NTP prevents RSV entry into host cells and proliferation by binding to the viral glycoproteins or inhibiting viral replication and assembly. | |||
| Ginkbilobin | HIV-1 | Peptides suppress the activity of HIV-1 reverse transcriptase. | [116,117,118,119] | |
| Ascalin | ||||
| Lunatusin | ||||
| Vulgarinin | ||||
| Meliacine | Foot-and-mouth disease virus (FMDV) | Meliacine prevents the hulling process of FMDV by inhibiting vacuolar acidification, and thus restrains the virus proliferation. | [120] | |
| Pep-RTYM | Dengue virus (DENV) | Pep-RTYM binds to the DENV particles to prevent viral interaction with host cell receptors and the subsequent nucleic acid release. | [121,122] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, X.; Wu, W.; Feng, L.; Treves, H.; Ren, M. Short Peptides Make a Big Difference: The Role of Botany-Derived AMPs in Disease Control and Protection of Human Health. Int. J. Mol. Sci. 2021, 22, 11363. https://doi.org/10.3390/ijms222111363
Luo X, Wu W, Feng L, Treves H, Ren M. Short Peptides Make a Big Difference: The Role of Botany-Derived AMPs in Disease Control and Protection of Human Health. International Journal of Molecular Sciences. 2021; 22(21):11363. https://doi.org/10.3390/ijms222111363
Chicago/Turabian StyleLuo, Xiumei, Wenxian Wu, Li Feng, Haim Treves, and Maozhi Ren. 2021. "Short Peptides Make a Big Difference: The Role of Botany-Derived AMPs in Disease Control and Protection of Human Health" International Journal of Molecular Sciences 22, no. 21: 11363. https://doi.org/10.3390/ijms222111363
APA StyleLuo, X., Wu, W., Feng, L., Treves, H., & Ren, M. (2021). Short Peptides Make a Big Difference: The Role of Botany-Derived AMPs in Disease Control and Protection of Human Health. International Journal of Molecular Sciences, 22(21), 11363. https://doi.org/10.3390/ijms222111363