Molecular Diversity of Anesthetic Actions Is Evident in Electroencephalogram Effects in Humans and Animals


Abstract
1. The Broad Molecular Targets of Anesthetics
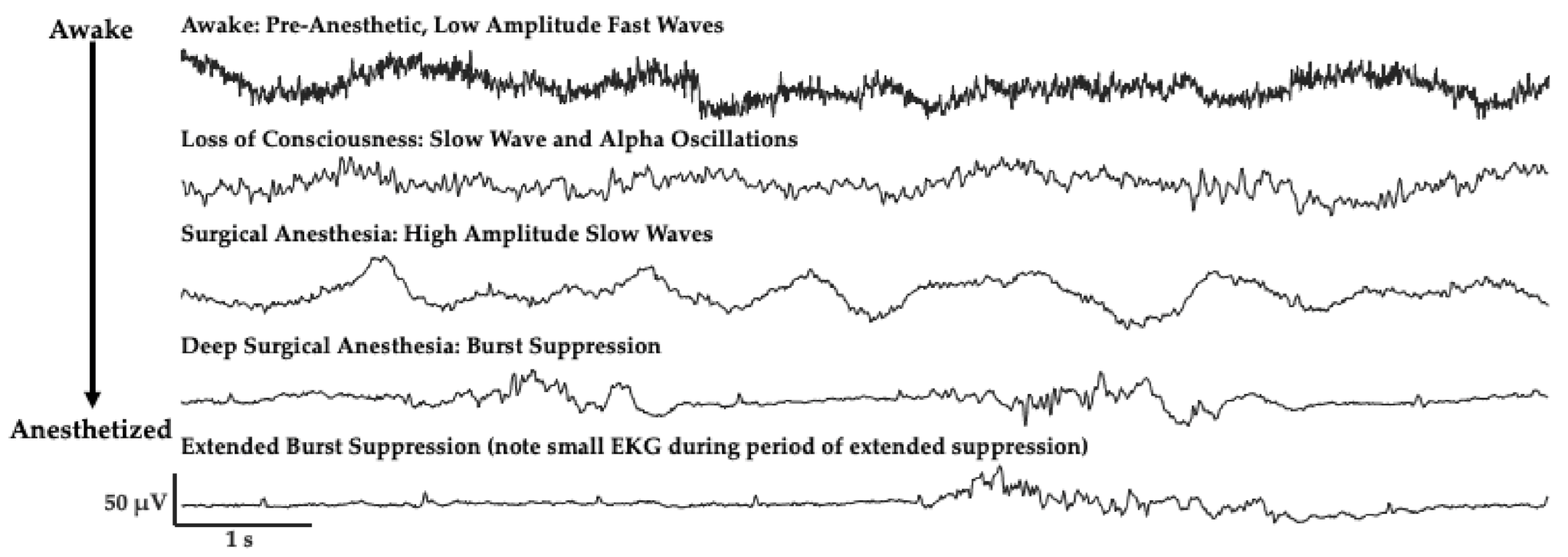
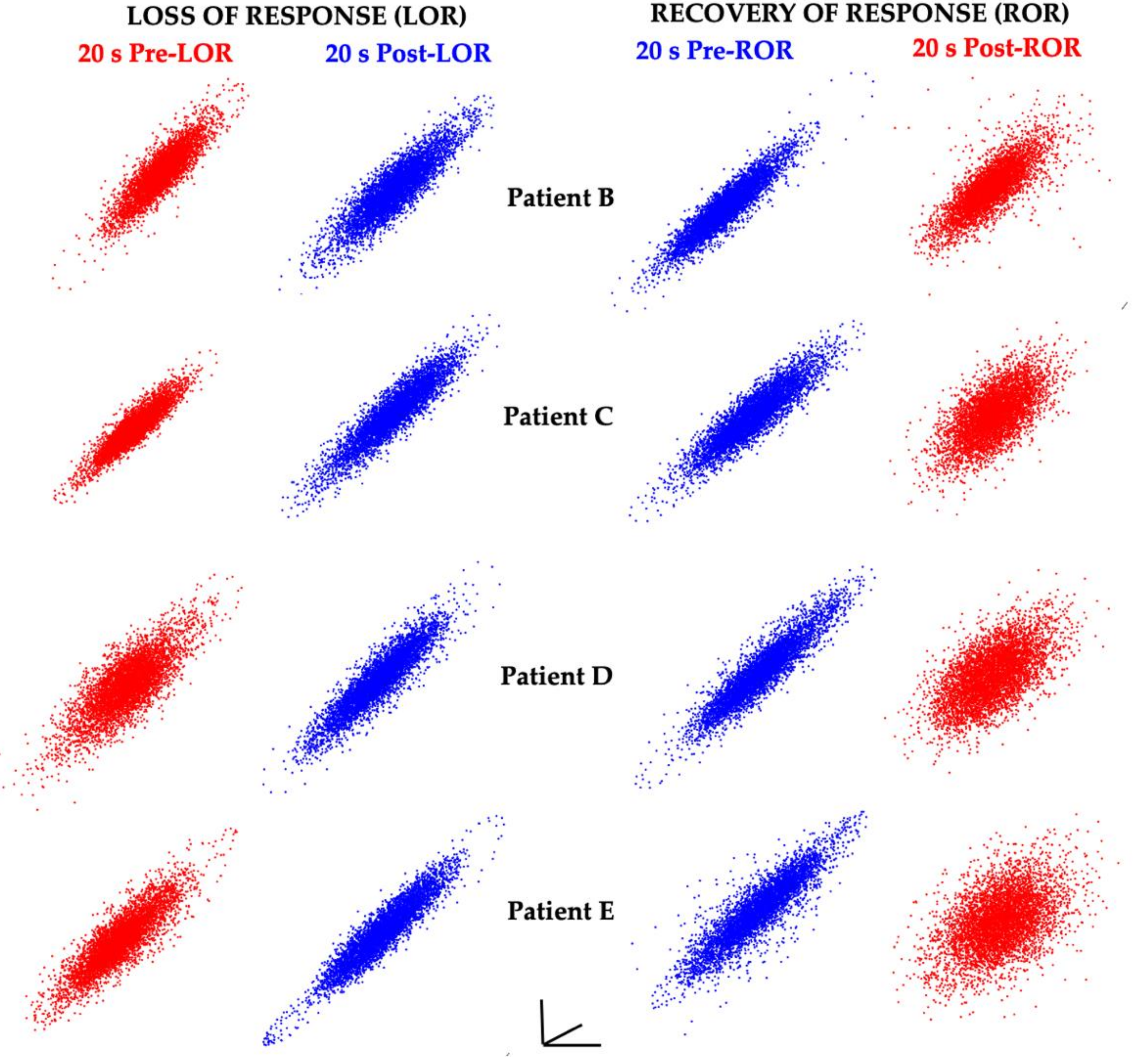
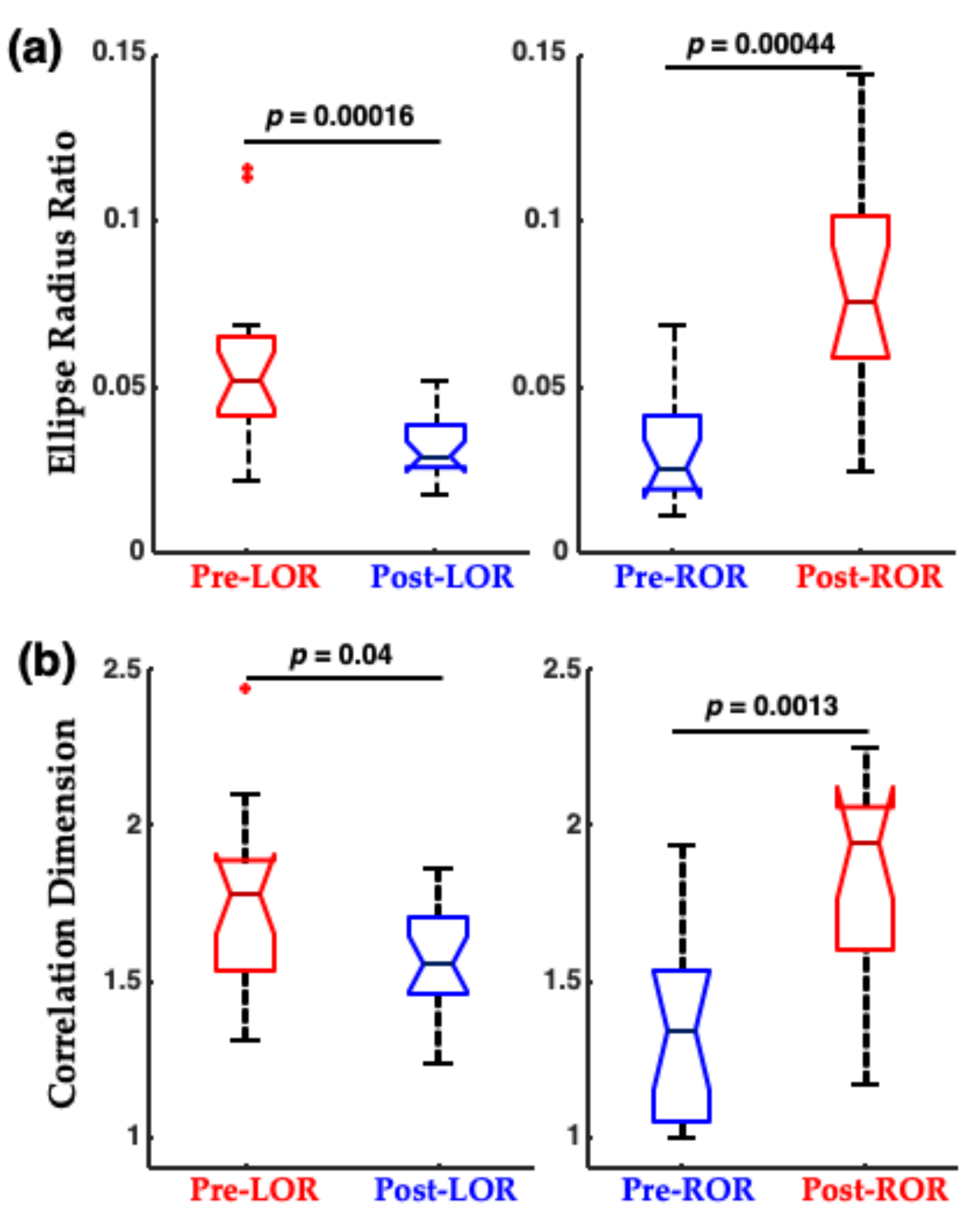
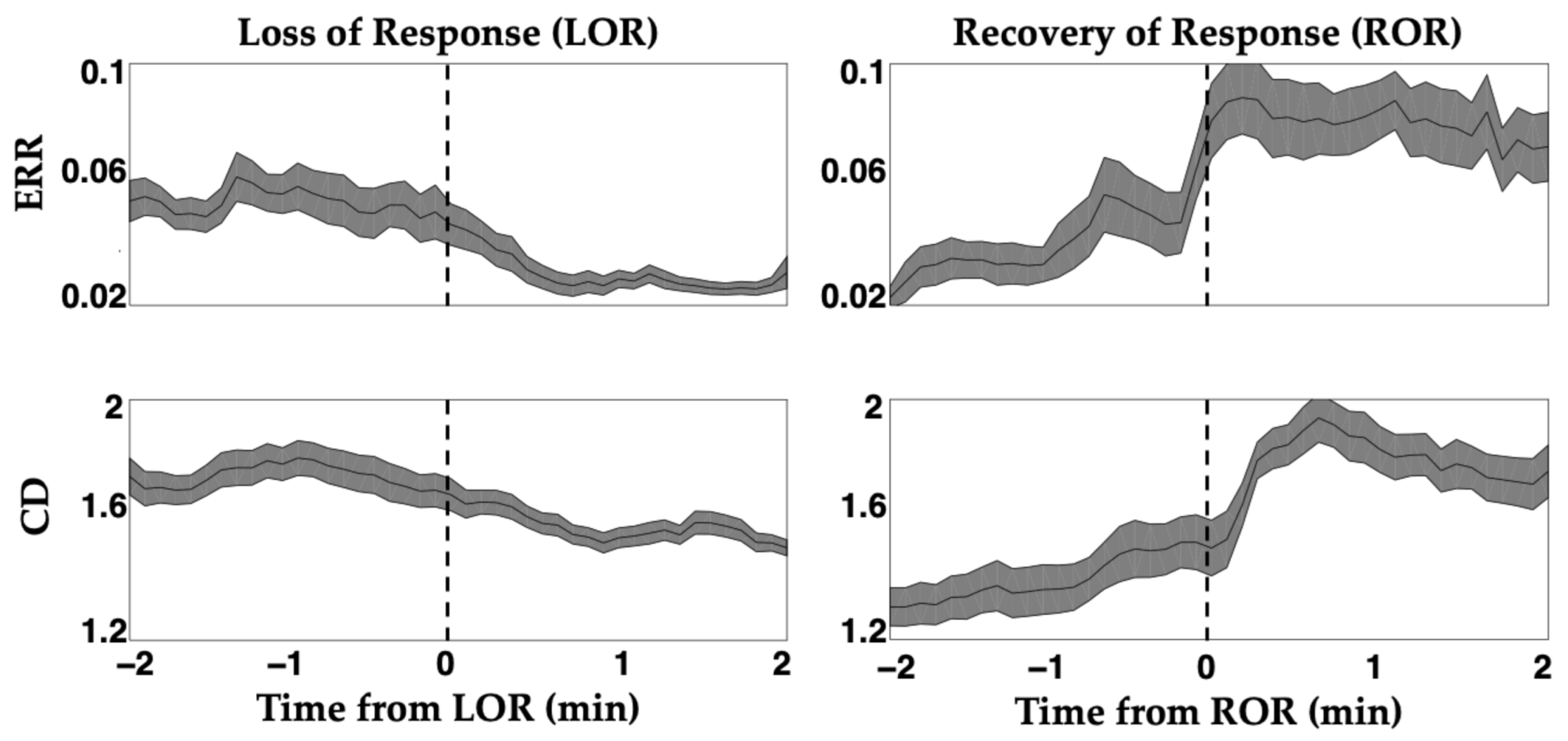
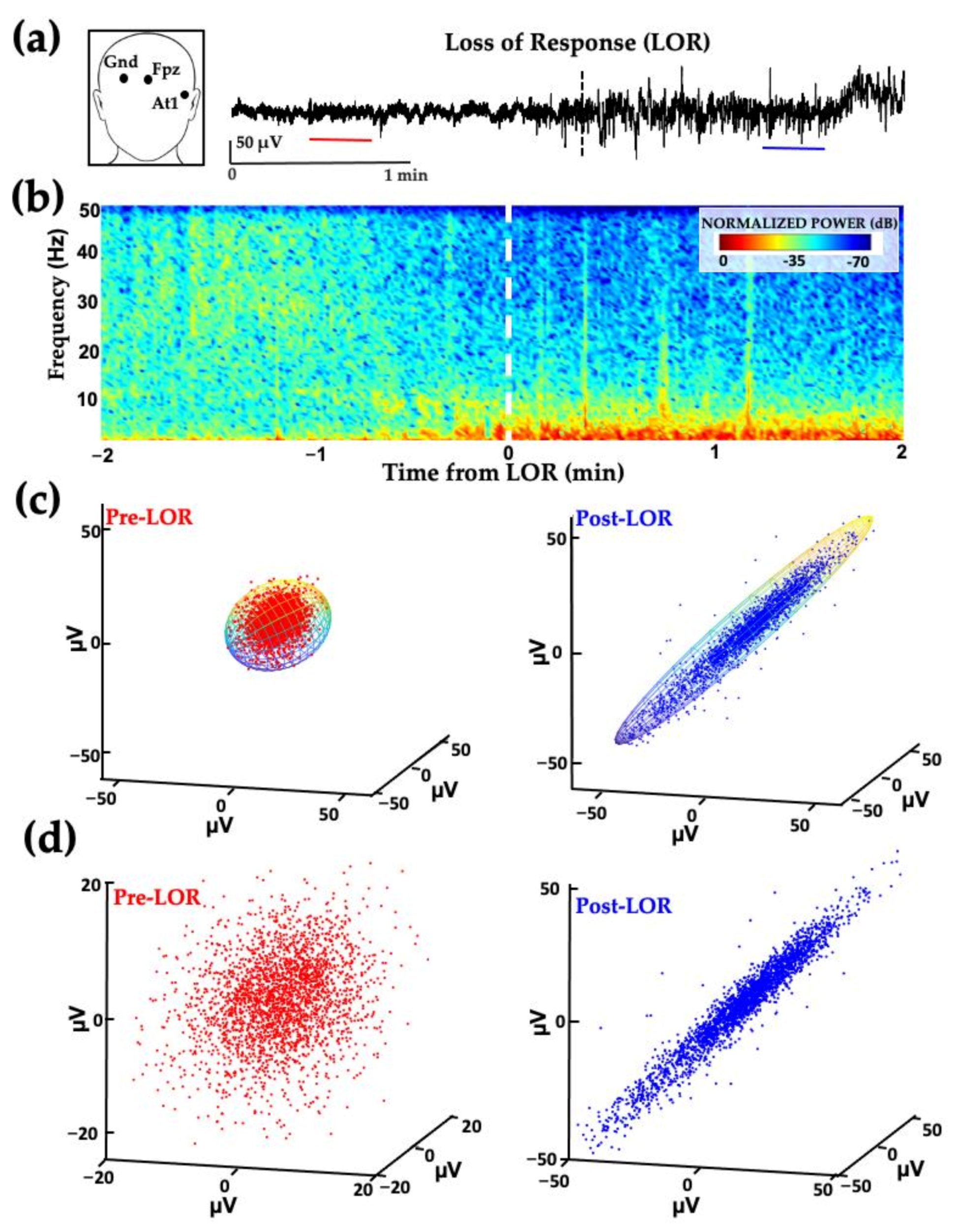
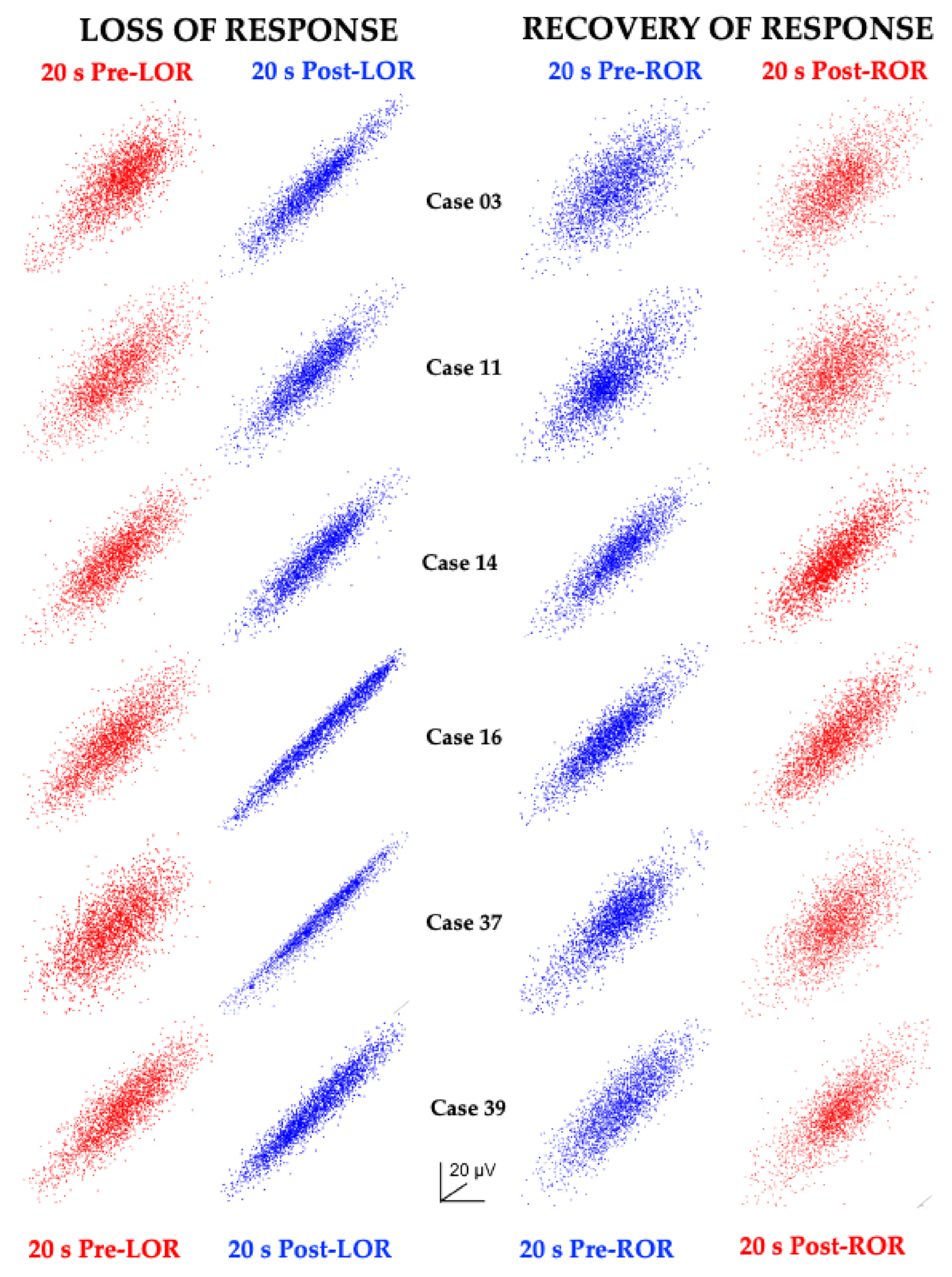
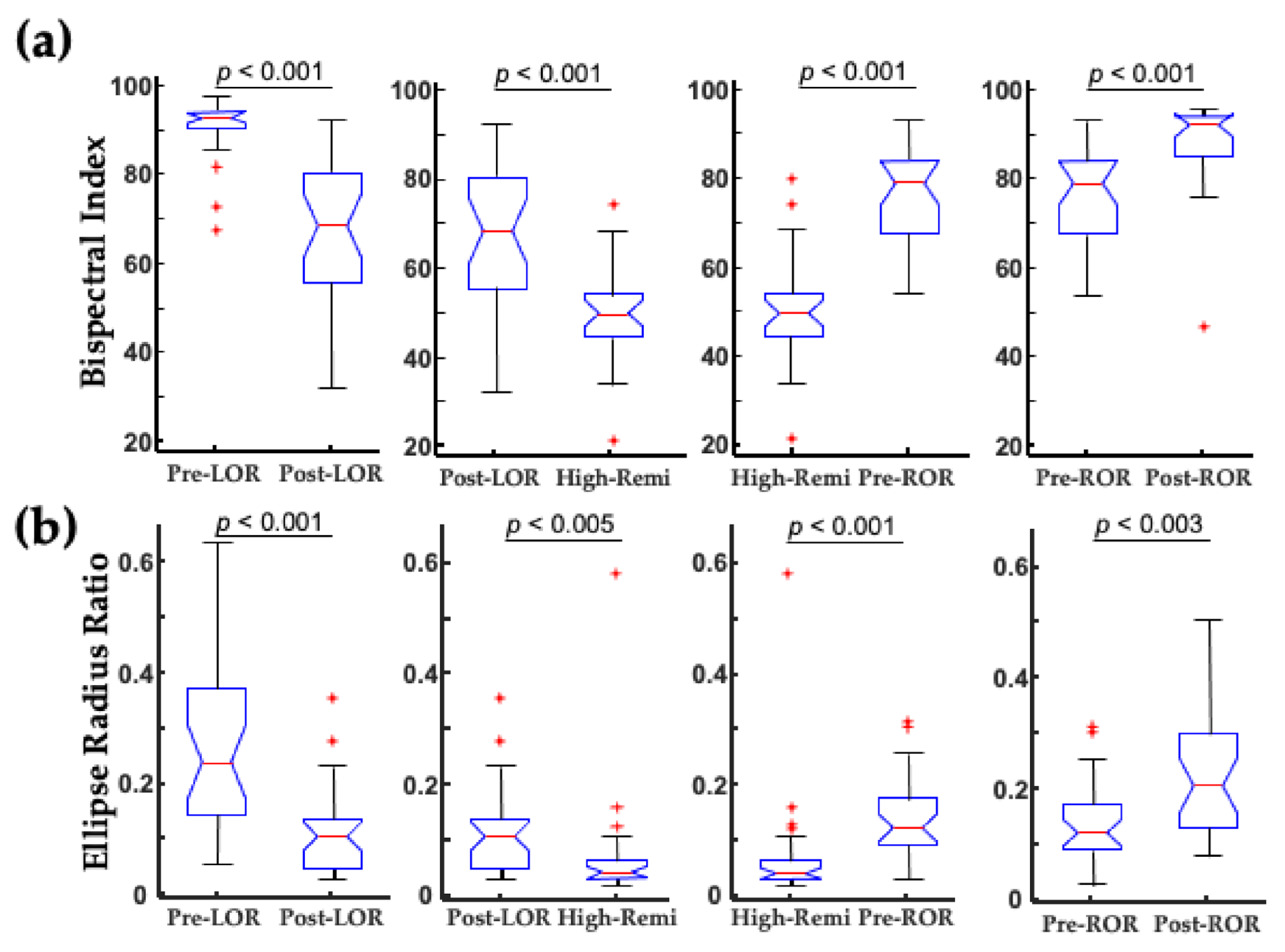
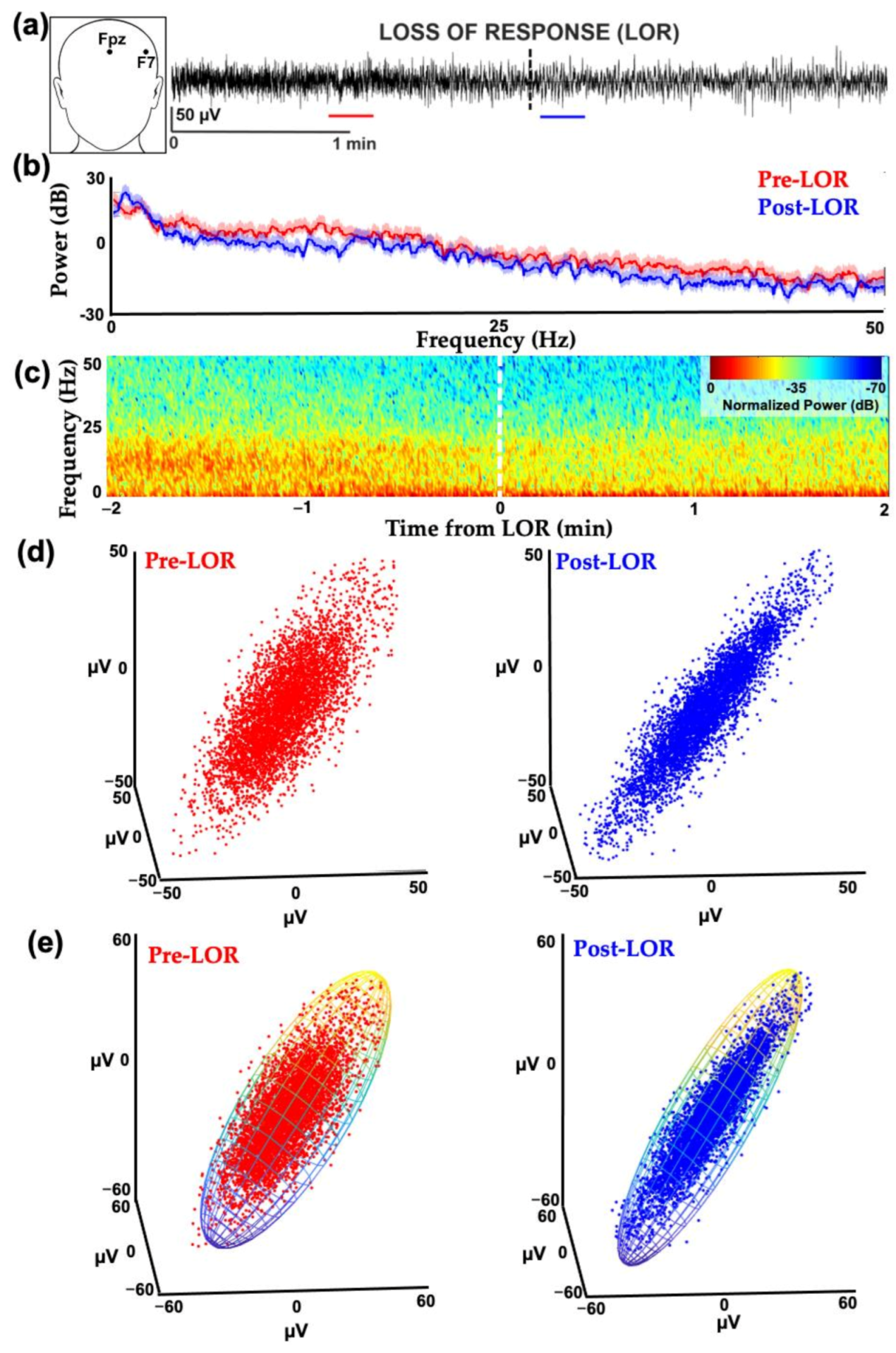
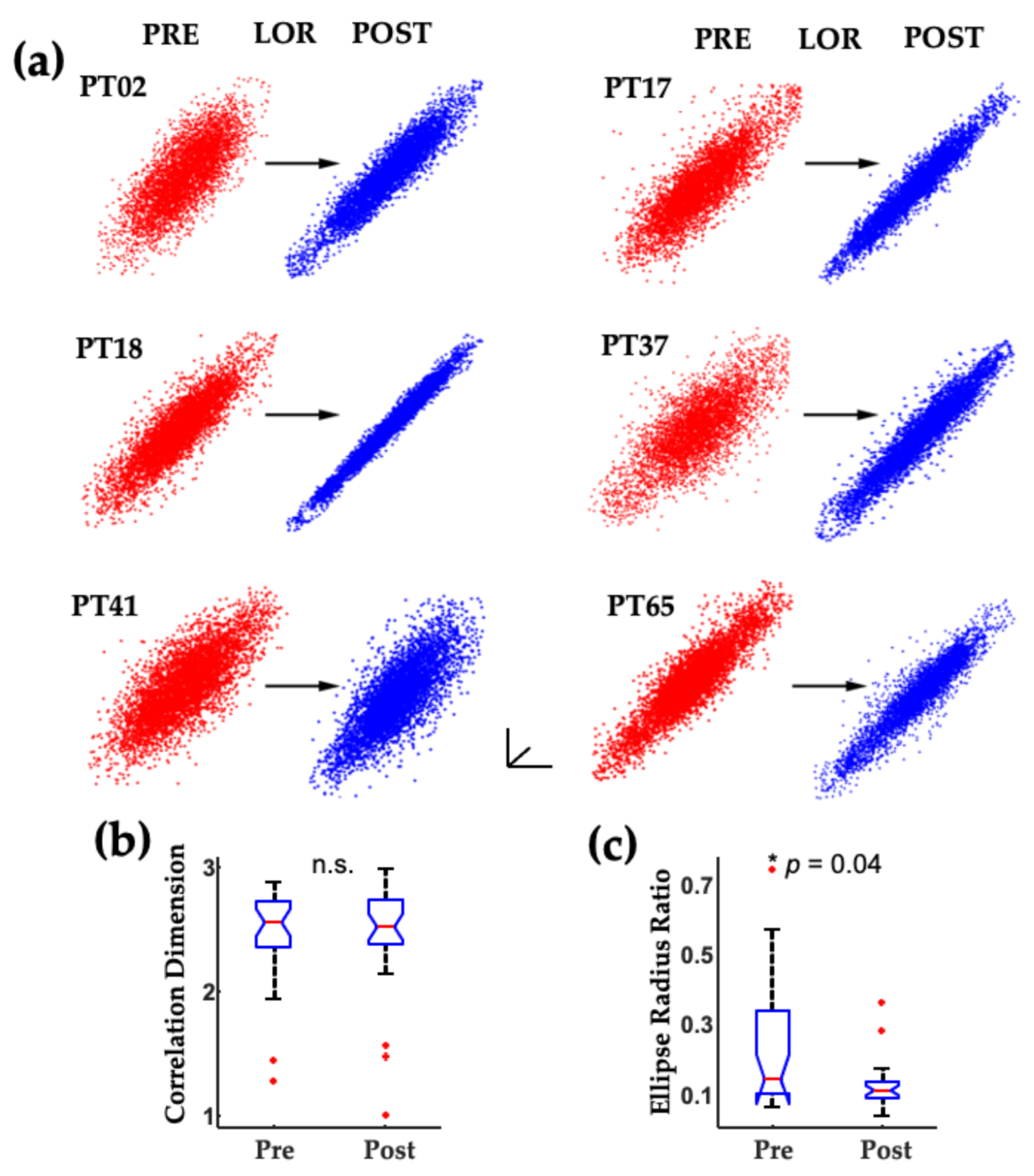
2. The Challenge of Finding Common Electrophysiology Biomarkers
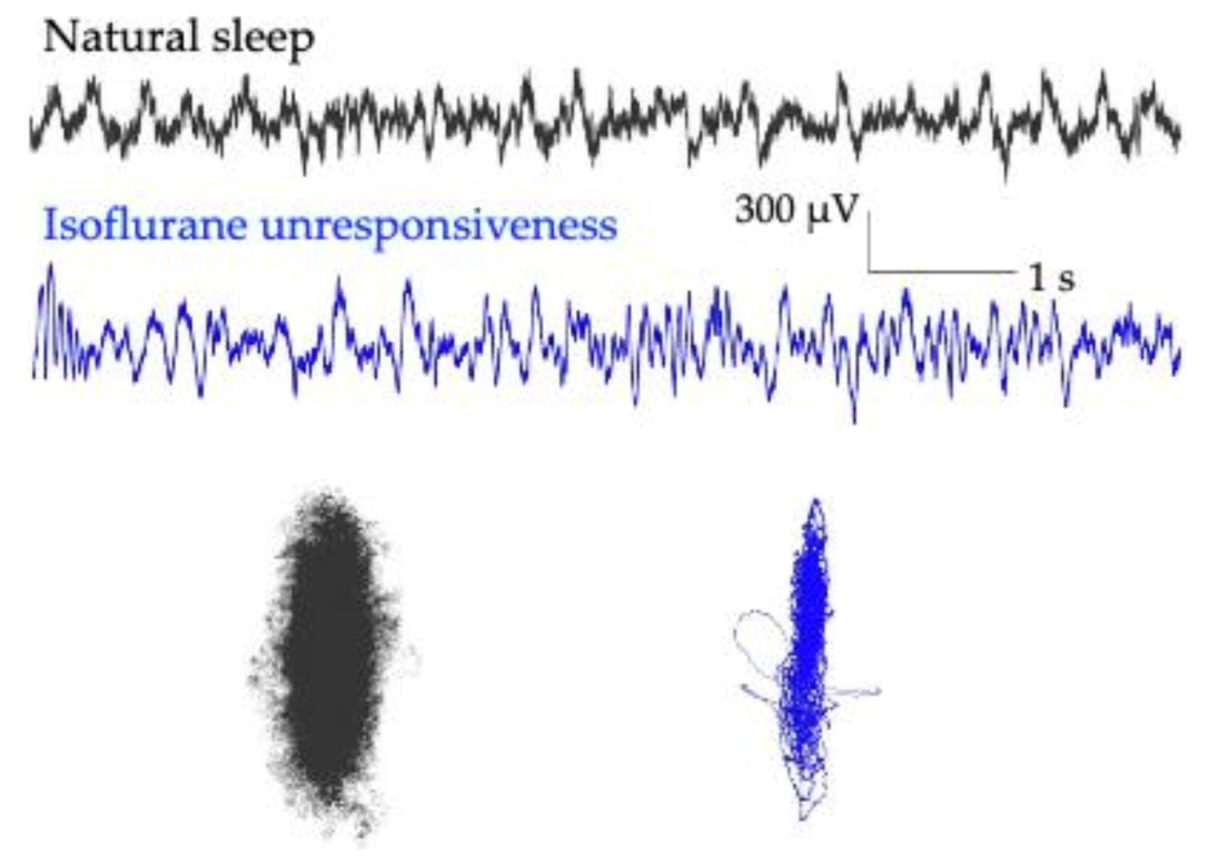
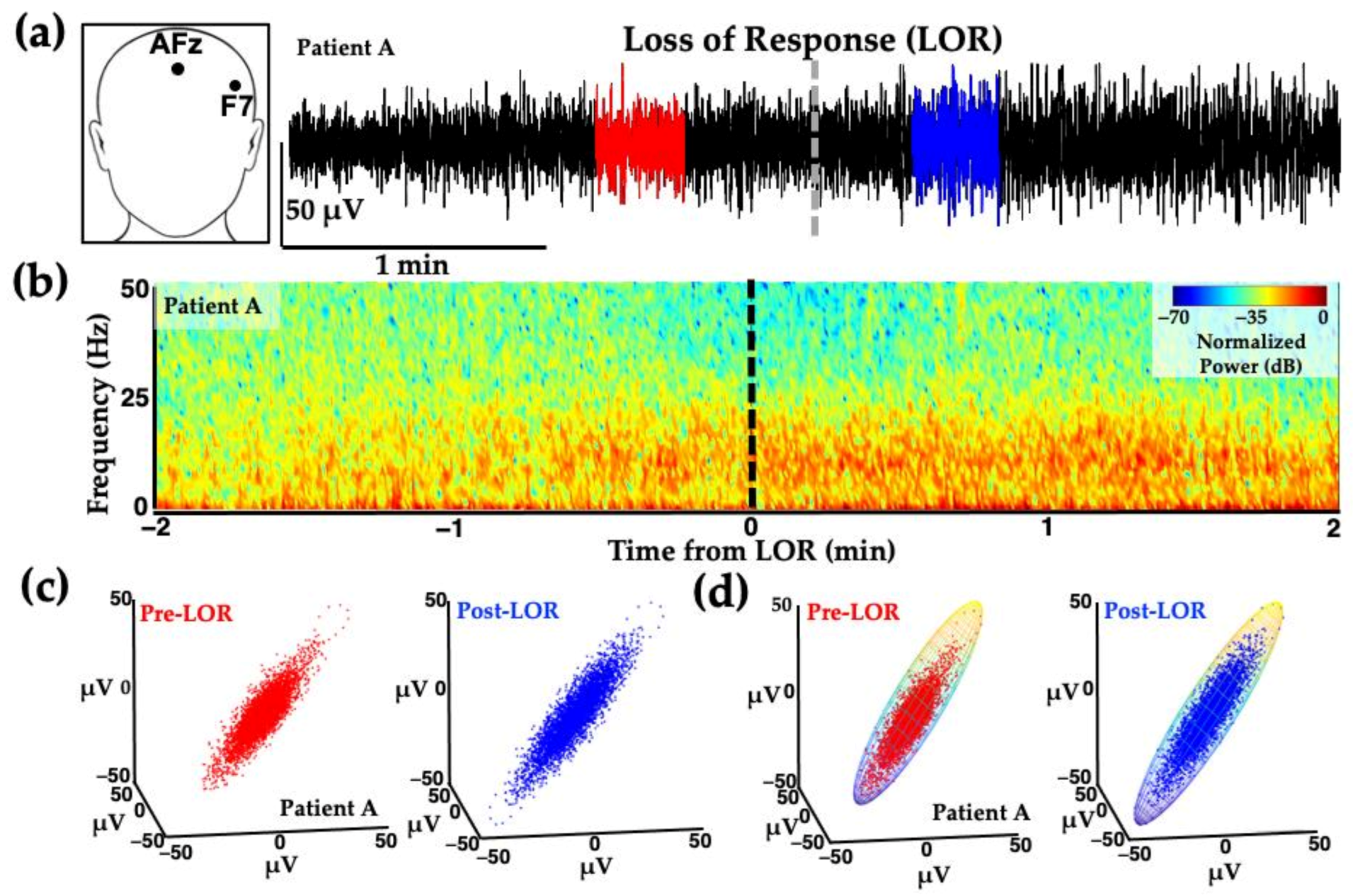
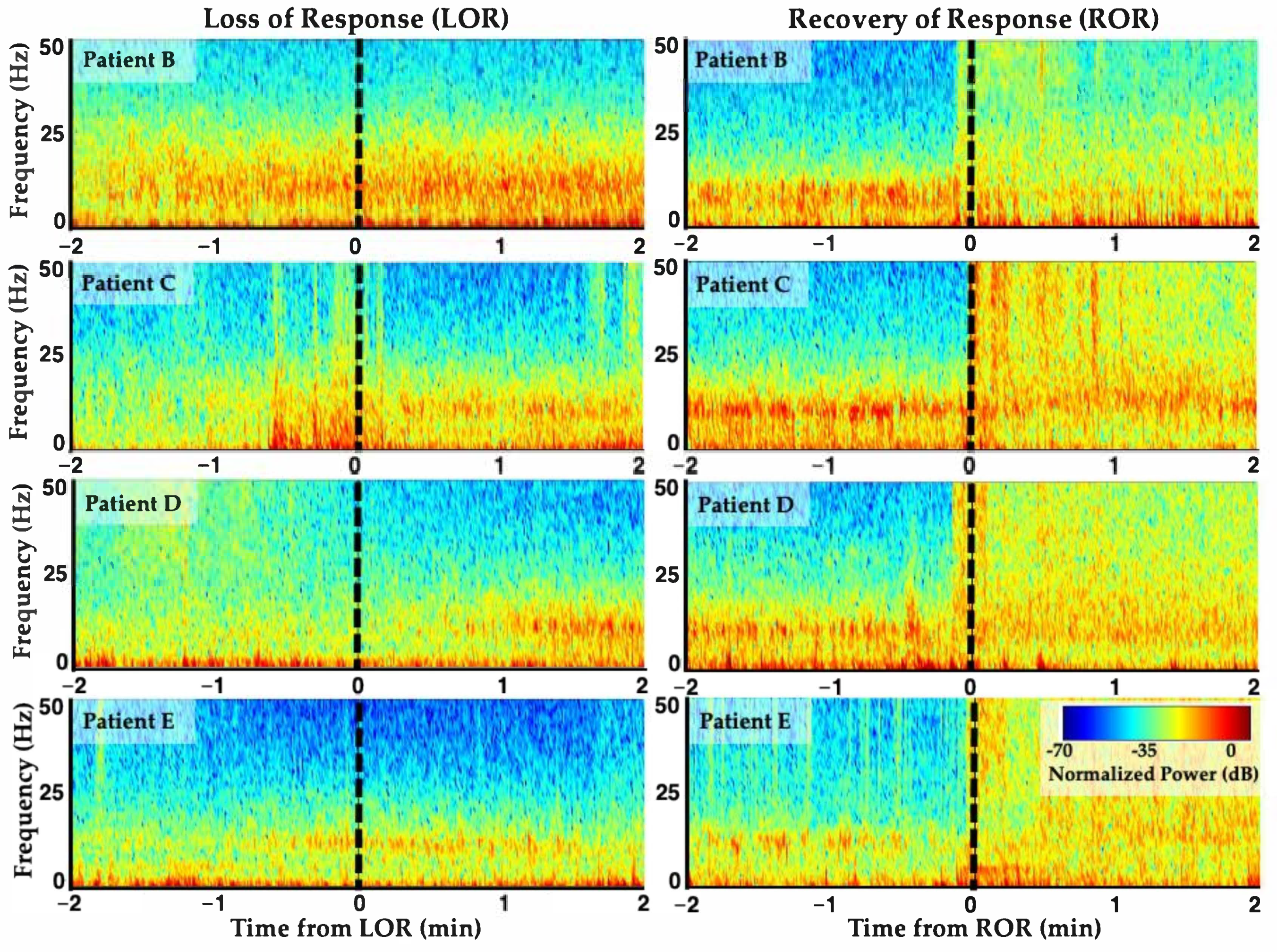
3. Unique Anesthetic Combinations Influence Diverse Molecular Targets
4. Certain Patient Cohorts Provide Additional Challenges Given Physiological and Metabolic Brain Changes
5. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| EEG | Electroencephalogram, electroencephalography |
| GABA | Gamma-aminobutyric acid |
| LOR | Loss of response |
| ROR | Recovery of response |
| CD | Correlation dimension |
| ERR | Ellipse radius ratio |
| N2O | Nitrous oxide |
| High-Remi | Highest blood concentration of remifentanil |
| BIS | Bispectral index |
References
- Hemmings, H.C.; Riegelhaupt, P.M.; Kelz, M.B.; Solt, K.; Eckenhoff, R.G.; Orser, B.A.; Goldstein, P.A. Towards a Comprehensive Understanding of Anesthetic Mechanisms of Action: A Decade of Discovery. Trends Pharmacol. Sci. 2019, 40, 464–481. [Google Scholar] [CrossRef]
- Hemmings, H.C.; Akabas, M.H.; Goldstein, P.A.; Trudell, J.R.; Orser, B.A.; Harrison, N.L. Emerging Molecular Mechanisms of General Anesthetic Action. Trends Pharmacol. Sci. 2005, 26, 503–510. [Google Scholar] [CrossRef]
- Brown, E.N.; Pavone, K.J.; Naranjo, M. Multimodal General Anesthesia: Theory and Practice. Anesth. Analg. 2018, 127, 1246–1258. [Google Scholar] [CrossRef] [PubMed]
- MacIver, M.B.; Bland, B.H. Chaos Analysis of EEG during Isoflurane-Induced Loss of Righting in Rats. Front. Syst. Neurosci. 2014, 8, 203. [Google Scholar] [CrossRef] [PubMed]
- Eagleman, S.L.; Drover, C.M.; Drover, D.R.; Ouellette, N.T.; MacIver, M.B. Remifentanil and Nitrous Oxide Anesthesia Produces a Unique Pattern of EEG Activity during Loss and Recovery of Response. Front. Hum. Neurosci. 2018, 12, 173. [Google Scholar] [CrossRef]
- Eagleman, S.L.; Vaughn, D.A.; Drover, D.R.; Drover, C.M.; Cohen, M.S.; Ouellette, N.T.; MacIver, M.B. Do Complexity Measures of Frontal EEG Distinguish Loss of Consciousness in Geriatric Patients Under Anesthesia? Front. Neurosci. 2018, 12, 645. [Google Scholar] [CrossRef]
- Eagleman, S.L.; Chander, D.; Reynolds, C.; Ouellette, N.T.; Maciver, M.B. Nonlinear Dynamics Captures Brain States at Different Levels of Consciousness in Patients Anesthetized with Propofol. PLoS ONE 2019, 14, e0223921. [Google Scholar] [CrossRef] [PubMed]
- Purdon, P.L.; Sampson, A.; Pavone, K.J.; Brown, E.N. Clinical Electroencephalography for Anesthesiologists: Part I: Background and Basic Signatures. Anesthesiology 2015, 123, 937–960. [Google Scholar] [CrossRef] [PubMed]
- Foster, B.L.; Liley, D.T.J. Effects of Nitrous Oxide Sedation on Resting Electroencephalogram Topography. Clin. Neurophysiol. 2013, 124, 417–423. [Google Scholar] [CrossRef]
- Yamamura, T.; Fukuda, M.; Takeya, H.; Goto, Y.; Furukawa, K. Fast Oscillatory EEG Activity Induced by Analgesic Concentrations of Nitrous Oxide in Man. Anesth. Analg. 1981, 60, 283–288. [Google Scholar] [CrossRef]
- Peyton, P.J.; Wu, C.Y. Nitrous Oxide–Related Postoperative Nausea and Vomiting Depends on Duration of Exposure. Anesthesiology 2014, 120, 1137–1145. [Google Scholar] [CrossRef] [PubMed]
- Kurdi, M.S.; Theerth, K.A.; Deva, R.S. Ketamine: Current Applications in Anesthesia, Pain, and Critical Care. Anesth. Essays Res. 2014, 8, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Naaz, S.; Ozair, E. Dexmedetomidine in Current Anaesthesia Practice—A Review. J. Clin. Diagn. Res. 2014, 8, GE01–GE04. [Google Scholar] [CrossRef] [PubMed]
- Mazanikov, M.; Udd, M.; Kylänpää, L.; Lindström, O.; Aho, P.; Halttunen, J.; Färkkilä, M.; Pöyhiä, R. Patient-Controlled Sedation with Propofol and Remifentanil for ERCP: A Randomized, Controlled Study. Gastrointest. Endosc. 2011, 73, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Egan, T.D.; Minto, C.F.; Hermann, D.J.; Barr, J.; Muir, K.T.; Shafer, S.L. Remifentanil versus Alfentanil: Comparative Pharmacokinetics and Pharmacodynamics in Healthy Adult Male Volunteers. Anesthesiology 1996, 84, 821–833. [Google Scholar] [CrossRef]
- Sivak, E.L.; Davis, P.J. Review of the Efficacy and Safety of Remifentanil for the Prevention and Treatment of Pain during and after Procedures and Surgery. Local Reg. Anesth. 2010, 3, 35–43. [Google Scholar]
- Buzsáki, G.; Anastassiou, C.A.; Koch, C. The Origin of Extracellular Fields and Currents-EEG, ECoG, LFP and Spikes. Nat. Rev. Neurosci. 2012, 13, 407–420. [Google Scholar] [CrossRef]
- Carlo, C.N.; Stevens, C.F. Structural Uniformity of Neocortex, Revisited. Proc. Natl. Acad. Sci. USA 2013, 110, 1488–1493. [Google Scholar] [CrossRef]
- Rampil, I.J. A Primer for EEG Signal Processing in Anesthesia. Anesthesiology 1998, 89, 980–1002. [Google Scholar] [CrossRef]
- Clark, D.L.; Rosner, B.S. Neurophysiologic Effects of General Anesthetics. I. The Electroencephalogram and Sensory Evoked Responses in Man. Anesthesiology 1973, 38, 564–582. [Google Scholar] [CrossRef]
- MacIver, M.B. Anesthetic Agent-Specific Effects on Synaptic Inhibition. Anesth. Analg. 2014, 119, 558–569. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Akeju, O.; Westover, M.B.; Pavone, K.J.; Sampson, A.L.; Hartnack, K.E.; Brown, E.N.; Purdon, P.L. Effects of Sevoflurane and Propofol on Frontal Electroencephalogram Power and Coherence. Anesthesiology 2014, 121, 990–998. [Google Scholar] [CrossRef] [PubMed]
- Purdon, P.L.; Pierce, E.T.; Mukamel, E.A.; Prerau, M.J.; Walsh, J.L.; Wong, K.F.K.; Salazar-Gomez, A.F.; Harrell, P.G.; Sampson, A.L.; Cimenser, A.; et al. Electroencephalogram Signatures of Loss and Recovery of Consciousness from Propofol. Proc. Natl. Acad. Sci. USA 2013, 110, E1142–E1151. [Google Scholar] [CrossRef] [PubMed]
- Ching, S.; Cimenser, A.; Purdon, P.L.; Brown, E.N.; Kopell, N.J. Thalamocortical Model for a Propofol-Induced-Rhythm Associated with Loss of Consciousness. Proc. Natl. Acad. Sci. USA 2010, 107, 22665–22670. [Google Scholar] [CrossRef]
- Watt, R.C.; Hameroff, S.R. Phase Space Electroencephalography (EEG): A New Mode of Intraoperative EEG Analysis. J. Clin. Monit. Comput. 1988, 5, 3–13. [Google Scholar] [CrossRef]
- Walling, P.T.; Hicks, K.N. Nonlinear Changes in Brain Dynamics during Emergence from Sevoflurane Anesthesia: Preliminary Exploration Using New Software. Anesthesiology 2006, 105, 927–935. [Google Scholar] [CrossRef]
- Frank, G.B.; Jhamandas, K. Effects of Drugs Acting Alone and in Combination on the Motor Activity of Intact Mice. Br. J. Pharmacol. 1970, 39, 696–706. [Google Scholar] [CrossRef]
- Franks, N.P. General Anaesthesia: From Molecular Targets to Neuronal Pathways of Sleep and Arousal. Nat. Rev. Neurosci. 2008, 9, 370–386. [Google Scholar] [CrossRef]
- Wasilczuk, A.Z.; Harrison, B.A.; Kwasniewska, P.; Ku, B.; Kelz, M.B.; McKinstry-Wu, A.R.; Proekt, A. Resistance to State Transitions in Responsiveness Is Differentially Modulated by Different Volatile Anaesthetics in Male Mice. Br. J. Anaesth. 2020, 125, 308–320. [Google Scholar] [CrossRef]
- McKinstry-Wu, A.R.; Wasilczuk, A.Z.; Harrison, B.A.; Bedell, V.M.; Sridharan, M.J.; Breig, J.J.; Pack, M.; Kelz, M.B.; Proekt, A. Analysis of Stochastic Fluctuations in Responsiveness Is a Critical Step toward Personalized Anesthesia. Elife 2019, 8, e50143. [Google Scholar] [CrossRef]
- Grassberger, P.; Procaccia, I. Characterization of Strange Attractors. Phys. Rev. Lett. 1983, 50, 346–349. [Google Scholar] [CrossRef]
- Albano, A.M.; Muench, J.; Schwartz, C.; Mees, A.I.; Rapp, P.E. Singular-Value Decomposition and the Grassberger-Procaccia Algorithm. Phys. Rev. A Gen. Phys. 1988, 38, 3017–3026. [Google Scholar] [CrossRef] [PubMed]
- Khachiyan, L.G. Polynomial Algorithms in Linear Programming. USSR Comput. Math. Math. Phys. 1980, 20, 53–72. [Google Scholar] [CrossRef]
- Kim, H.; Moon, J.-Y.; Mashour, G.A.; Lee, U. Mechanisms of Hysteresis in Human Brain Networks during Transitions of Consciousness and Unconsciousness: Theoretical Principles and Empirical Evidence. PLoS Comput. Biol. 2018, 14, e1006424. [Google Scholar] [CrossRef] [PubMed]
- Pleuvry, B.J. Hysteresis in Drug Response. Anaesth. Intensive Care Med. 2008, 9, 372–373. [Google Scholar] [CrossRef]
- Sepúlveda, P.O.; Carrasco, E.; Tapia, L.F.; Ramos, M.; Cruz, F.; Conget, P.; Olivares, Q.F.B.; Cortínez, I. Evidence of Hysteresis in Propofol Pharmacodynamics. Anaesthesia 2018, 73, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Hudetz, A.G.; Liu, X.; Pillay, S.; Boly, M.; Tononi, G. Propofol Anesthesia Reduces Lempel-Ziv Complexity of Spontaneous Brain Activity in Rats. Neurosci. Lett. 2016, 628, 132–135. [Google Scholar] [CrossRef]
- Tononi, G. Information Measures for Conscious Experience. Arch. Ital. Biol. 2001, 139, 367–371. [Google Scholar]
- Abásolo, D.; Simons, S.; Morgado da Silva, R.; Tononi, G.; Vyazovskiy, V.V. Lempel-Ziv Complexity of Cortical Activity during Sleep and Waking in Rats. J. Neurophysiol. 2015, 113, 2742–2752. [Google Scholar] [CrossRef]
- Sarasso, S.; Boly, M.; Napolitani, M.; Gosseries, O.; Charland-Verville, V.; Casarotto, S.; Rosanova, M.; Casali, A.G.; Brichant, J.-F.; Boveroux, P.; et al. Consciousness and Complexity during Unresponsiveness Induced by Propofol, Xenon, and Ketamine. Curr. Biol. CB 2015, 25, 3099–3105. [Google Scholar] [CrossRef]
- Casali, A.G.; Gosseries, O.; Rosanova, M.; Boly, M.; Sarasso, S.; Casali, K.R.; Casarotto, S.; Bruno, M.-A.; Laureys, S.; Tononi, G.; et al. A Theoretically Based Index of Consciousness Independent of Sensory Processing and Behavior. Sci. Transl. Med. 2013, 5, 198ra105. [Google Scholar] [CrossRef]
- Li, D.; Vlisides, P.E.; Kelz, M.B.; Avidan, M.S.; Mashour, G.A.; Blain-Moraes, S.; Golmirzaie, G.; Janke, E.; Picton, P.; Tarnal, V.; et al. Dynamic Cortical Connectivity during General Anesthesia in Healthy Volunteers. Anesthesiology 2019, 130, 870–884. [Google Scholar] [CrossRef] [PubMed]
- Vlisides, P.E.; Li, D.; Zierau, M.; Lapointe, A.P.; Ip, K.I.; McKinney, A.M.; Mashour, G.A. Dynamic Cortical Connectivity during General Anesthesia in Surgical Patients. Anesthesiology 2019, 130, 885–897. [Google Scholar] [CrossRef] [PubMed]
- Mashour, G.A.; Hudetz, A.G. Neural Correlates of Unconsciousness in Large-Scale Brain Networks. Trends Neurosci. 2018, 41, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Mashour, G.A.; Noh, G.-J.; Kim, S.; Lee, U. Reconfiguration of Network Hub Structure after Propofol-Induced Unconsciousness. Anesthesiology 2013, 119, 1347–1359. [Google Scholar] [CrossRef]
- Wang, D.J.J.; Jann, K.; Fan, C.; Qiao, Y.; Zang, Y.-F.; Lu, H.; Yang, Y. Neurophysiological Basis of Multi-Scale Entropy of Brain Complexity and Its Relationship With Functional Connectivity. Front. Neurosci. 2018, 12, 352. [Google Scholar] [CrossRef] [PubMed]
- Sleigh, J.; Harvey, M.; Voss, L.; Denny, B. Ketamine—More Mechanisms of Action than Just NMDA Blockade. Trends Anaesth. Crit. Care 2014, 4, 76–81. [Google Scholar] [CrossRef]
- Durieux, M.E. Inhibition by Ketamine of Muscarinic Acetylcholine Receptor Function. Anesth. Analg. 1995, 81, 57–62. [Google Scholar]
- Foster, B.L.; Liley, D.T.J. Nitrous Oxide Paradoxically Modulates Slow Electroencephalogram Oscillations. Anesth. Analg. 2011, 113, 758–765. [Google Scholar] [CrossRef]
- Kuhlmann, L.; Liley, D.T.J. Assessing Nitrous Oxide Effect Using Electroencephalographically-Based Depth of Anesthesia Measures Cortical State and Cortical Input. J. Clin. Monit. Comput. 2018, 32, 173–188. [Google Scholar] [CrossRef]
- Akeju, O.; Song, A.H.; Hamilos, A.E.; Pavone, K.J.; Flores, F.J.; Brown, E.N.; Purdon, P.L. Electroencephalogram Signatures of Ketamine Anesthesia-Induced Unconsciousness. Clin. Neurophysiol. 2016, 127, 2414–2422. [Google Scholar] [CrossRef]
- Blain-Moraes, S.; Lee, U.; Ku, S.; Noh, G.; Mashour, G.A. Electroencephalographic Effects of Ketamine on Power, Cross-Frequency Coupling, and Connectivity in the Alpha Bandwidth. Front. Syst. Neurosci. 2014, 8, 114. [Google Scholar] [CrossRef]
- Akeju, O.; Kim, S.-E.; Vazquez, R.; Rhee, J.; Pavone, K.J.; Hobbs, L.E.; Purdon, P.L.; Brown, E.N. Spatiotemporal Dynamics of Dexmedetomidine-Induced Electroencephalogram Oscillations. PLoS ONE 2016, 11, e0163431. [Google Scholar] [CrossRef]
- Absalom, A.R.; Menon, D.K. BIS and Spectral Entropy Monitoring during Sedation with Midazolam/Remifentanil and Dexmedetomidine/Remifentanil. Crit. Care (Lond. Engl.) 2009, 13, 137. [Google Scholar] [CrossRef]
- Pavone, K.J.; Akeju, O.; Sampson, A.L.; Ling, K.; Purdon, P.L.; Brown, E.N. Nitrous Oxide-Induced Slow and Delta Oscillations. Clin. Neurophysiol. 2016, 127, 556–564. [Google Scholar] [CrossRef]
- Rampil, I.J.; Kim, J.S.; Lenhardt, R.; Negishi, C.; Sessler, D.I. Bispectral EEG Index during Nitrous Oxide Administration. Anesthesiology 1998, 89, 671–677. [Google Scholar] [CrossRef]
- Doufas, A.G.; Bakhshandeh, M.; Haugh, G.S.; Bjorksten, A.R.; Greif, R.; Sessler, D.I. Automated Responsiveness Test and Bispectral Index Monitoring during Propofol and Propofol/N2O Sedation. Acta Anaesthesiol. Scand. 2003, 47, 951–957. [Google Scholar] [CrossRef]
- Drover, D.R.; Lemmens, H.J. Population Pharmacodynamics and Pharmacokinetics of Remifentanil as a Supplement to Nitrous Oxide Anesthesia for Elective Abdominal Surgery. Anesthesiology 1998, 89, 869–877. [Google Scholar] [CrossRef]
- Cornelissen, L.; Kim, S.-E.; Purdon, P.L.; Brown, E.N.; Berde, C.B. Age-Dependent Electroencephalogram (EEG) Patterns during Sevoflurane General Anesthesia in Infants. Elife 2015, 4, e06513. [Google Scholar] [CrossRef]
- Lee, J.M.; Akeju, O.; Terzakis, K.; Pavone, K.J.; Deng, H.; Houle, T.T.; Firth, P.G.; Shank, E.S.; Brown, E.N.; Purdon, P.L. A Prospective Study of Age-Dependent Changes in Propofol-Induced Electroencephalogram Oscillations in Children. Anesthesiology 2017, 127, 293–306. [Google Scholar] [CrossRef]
- Akeju, O.; Pavone, K.J.; Thum, J.A.; Firth, P.G.; Westover, M.B.; Puglia, M.; Shank, E.S.; Brown, E.N.; Purdon, P.L. Age-Dependency of Sevoflurane-Induced Electroencephalogram Dynamics in Children. Br. J. Anaesth. 2015, 115, i66–i76. [Google Scholar] [CrossRef] [PubMed]
- Rigouzzo, A.; Khoy-Ear, L.; Laude, D.; Louvet, N.; Moutard, M.-L.; Sabourdin, N.; Constant, I. EEG Profiles during General Anesthesia in Children: A Comparative Study between Sevoflurane and Propofol. Pediatr. Anesth. 2019, 29, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Mangoni, A.A.; Jackson, S.H.D. Age-Related Changes in Pharmacokinetics and Pharmacodynamics: Basic Principles and Practical Applications. Br. J. Clin. Pharmacol. 2004, 57, 6–14. [Google Scholar] [CrossRef] [PubMed]
- Kanonidou, Z.; Karystianou, G. Anesthesia for the Elderly. Hippokratia 2007, 11, 175–177. [Google Scholar] [PubMed]
- Dustman, R.E.; Shearer, D.E.; Emmerson, R.Y. Life-Span Changes in EEG Spectral Amplitude, Amplitude Variability and Mean Frequency. Clin. Neurophysiol. 1999, 110, 1399–1409. [Google Scholar] [CrossRef]
- Zappasodi, F.; Marzetti, L.; Olejarczyk, E.; Tecchio, F.; Pizzella, V. Age-Related Changes in Electroencephalographic Signal Complexity. PLoS ONE 2015, 10, e0141995. [Google Scholar] [CrossRef]
- Voytek, B.; Kramer, M.A.; Case, J.; Lepage, K.Q.; Tempesta, Z.R.; Knight, R.T.; Gazzaley, A. Age-Related Changes in 1/f Neural Electrophysiological Noise. J. Neurosci. 2015, 35, 13257–13265. [Google Scholar] [CrossRef]
- Yeatman, J.D.; Wandell, B.A.; Mezer, A.A. Lifespan Maturation and Degeneration of Human Brain White Matter. Nat. Commun. 2014, 5, 4932. [Google Scholar] [CrossRef]
- Bentourkia, M.; Bol, A.; Ivanoiu, A.; Labar, D.; Sibomana, M.; Coppens, A.; Michel, C.; Cosnard, G.; De Volder, A.G. Comparison of Regional Cerebral Blood Flow and Glucose Metabolism in the Normal Brain: Effect of Aging. J. Neurol. Sci. 2000, 181, 19–28. [Google Scholar] [CrossRef]
- Brown, E.N.; Purdon, P.L. The Aging Brain and Anesthesia. Curr. Opin. Anaesthesiol. 2013, 26, 414–419. [Google Scholar] [CrossRef]
- Drover, D.R.; Schmiesing, C.; Buchin, A.F.; Ortega, H.R.; Tanner, J.W.; Atkins, J.H.; Macario, A. Titration of Sevoflurane in Elderly Patients: Blinded, Randomized Clinical Trial, in Non-Cardiac Surgery after Beta-Adrenergic Blockade. J. Clin. Monit. Comput. 2011, 25, 175–181. [Google Scholar] [CrossRef]
- Ghosh, I.; Bithal, P.K.; Dash, H.H.; Chaturvedi, A.; Prabhakar, H. Both Clonidine and Metoprolol Modify Anesthetic Depth Indicators and Reduce Intraoperative Propofol Requirement. J. Anesth. 2008, 22, 131–134. [Google Scholar] [CrossRef] [PubMed]
- Zaugg, M.; Tagliente, T.; Silverstein, J.H.; Lucchinetti, E. Atenolol May Not Modify Anesthetic Depth Indicators in Elderly Patients—A Second Look at the Data. Can. J. Anesth. 2003, 50, 638–642. [Google Scholar] [CrossRef] [PubMed]
- Johansen, J.W. Esmolol Promotes Electroencephalographic Burst Suppression during Propofol/Alfentanil Anesthesia. Anesth. Analg. 2001, 93, 1526–1531. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Fleming, L.M.; Javitt, D.C.; Carter, C.S.; Kantrowitz, J.T.; Girgis, R.R.; Kegeles, L.S.; Ragland, J.D.; Maddock, R.J.; Lesh, T.A.; Tanase, C.; et al. A Multicenter Study of Ketamine Effects on Functional Connectivity: Large Scale Network Relationships, Hubs and Symptom Mechanisms. Neuroimage Clin. 2019, 22, 101739. [Google Scholar] [CrossRef]
- Hentschke, H.; Raz, A.; Krause, B.M.; Murphy, C.A.; Banks, M.I. Disruption of Cortical Network Activity by the General Anaesthetic Isoflurane. Br. J. Anaesth. 2017, 119, 685–696. [Google Scholar] [CrossRef] [PubMed]
- Mashour, G.A.; Hudetz, A.G. Bottom-Up and Top-Down Mechanisms of General Anesthetics Modulate Different Dimensions of Consciousness. Front. Neural Circuits 2017, 11, 44. [Google Scholar] [CrossRef] [PubMed]
- Lukatch, H.S.; MacIver, M.B. Synaptic Mechanisms of Thiopental-Induced Alterations in Synchronized Cortical Activity. Anesthesiology 1996, 84, 1425–1434. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Anesthetic Agent | Uses | Mechanism of Action | EEG Effects | Advantages | Disadvantages |
|---|---|---|---|---|---|
| Propofol | Common general anesthetic. Can be administered at low doses as a sedative. | GABA agonist [8]. | Alpha and beta oscillations during sedation. Sleep-like EEG pattern during general anesthesia. Burst suppression to isoelectricity at higher doses [8]. | Rapid onset and offset. | Hard-to-monitor without EEG. Can decrease blood pressure and cause breathing difficulties. |
| Halogenated Ethers (e.g., Sevoflurane) | Common general anesthetic. | Predominately suppresses glutamate-mediated excitation. | Sleep-like EEG pattern during general anesthesia. Burst suppression to isoelectricity at higher doses. | Rapid onset and offset. Easy-to-monitor with MAC values. | May lead to extended burst suppression and peri-operative neurocognitive decline in geriatric patients. |
| Nitrous Oxide | Supplemental anesthetic agent. Used for analgesia. | NMDA antagonist. | Maintenance of wake-like EEG activity in beta and gamma range [8,9,10]. | Rapid onset and offset. Few drug interactions. | Weak anesthetic agent. Increased risk of nausea [11]. |
| Ketamine | Can be used for sedation, as a supplemental anesthetic agent, and for antinociception. | Many molecular targets, including NMDA receptors [1,12]. | Beta and gamma oscillations at sedation [8]. Maintenance of wake-like EEG activity. | Bronchodilator, protects patients with reactive airways, preserves spontaneous respirations and airway [12]. | Frequent use could cause memory impairment. Increases muscle tone. Airway compromise may occur [12] |
| Dexmedetomidine | Used as a supplemental anesthetic agent and for analgesia. | Alpha-2-adrenoceptor agonist. Blocks norepinephrine release which activates GABA and other inhibitory projections [8]. | Maintenance of wake-like EEG activity. | Anesthetic sparing. Preserves respiratory function and reduces delirium. Cardiovascular sparing [13]. | May cause bradycardia, hypotension, hypertension, nausea, and dry mouth [13]. |
| Remifentanil | Used as a supplemental anesthetic agent [14] and for analgesia. | Mu-type-opioid receptor agonist. | Sleep-like EEG pattern during general anesthesia [15]. | Rapid onset and offset compared to other opioids. Can decrease overall general anesthesia needs [15]. | May increase nausea, respiratory depression, and hypoxia [16]. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eagleman, S.; MacIver, M.B. Molecular Diversity of Anesthetic Actions Is Evident in Electroencephalogram Effects in Humans and Animals. Int. J. Mol. Sci. 2021, 22, 495. https://doi.org/10.3390/ijms22020495
Eagleman S, MacIver MB. Molecular Diversity of Anesthetic Actions Is Evident in Electroencephalogram Effects in Humans and Animals. International Journal of Molecular Sciences. 2021; 22(2):495. https://doi.org/10.3390/ijms22020495
Chicago/Turabian StyleEagleman, Sarah, and M. Bruce MacIver. 2021. "Molecular Diversity of Anesthetic Actions Is Evident in Electroencephalogram Effects in Humans and Animals" International Journal of Molecular Sciences 22, no. 2: 495. https://doi.org/10.3390/ijms22020495
APA StyleEagleman, S., & MacIver, M. B. (2021). Molecular Diversity of Anesthetic Actions Is Evident in Electroencephalogram Effects in Humans and Animals. International Journal of Molecular Sciences, 22(2), 495. https://doi.org/10.3390/ijms22020495